
Mast Cell Proteases Tryptase and Chymase Induce Migratory and Morphological Alterations in Bronchial Epithelial Cells


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
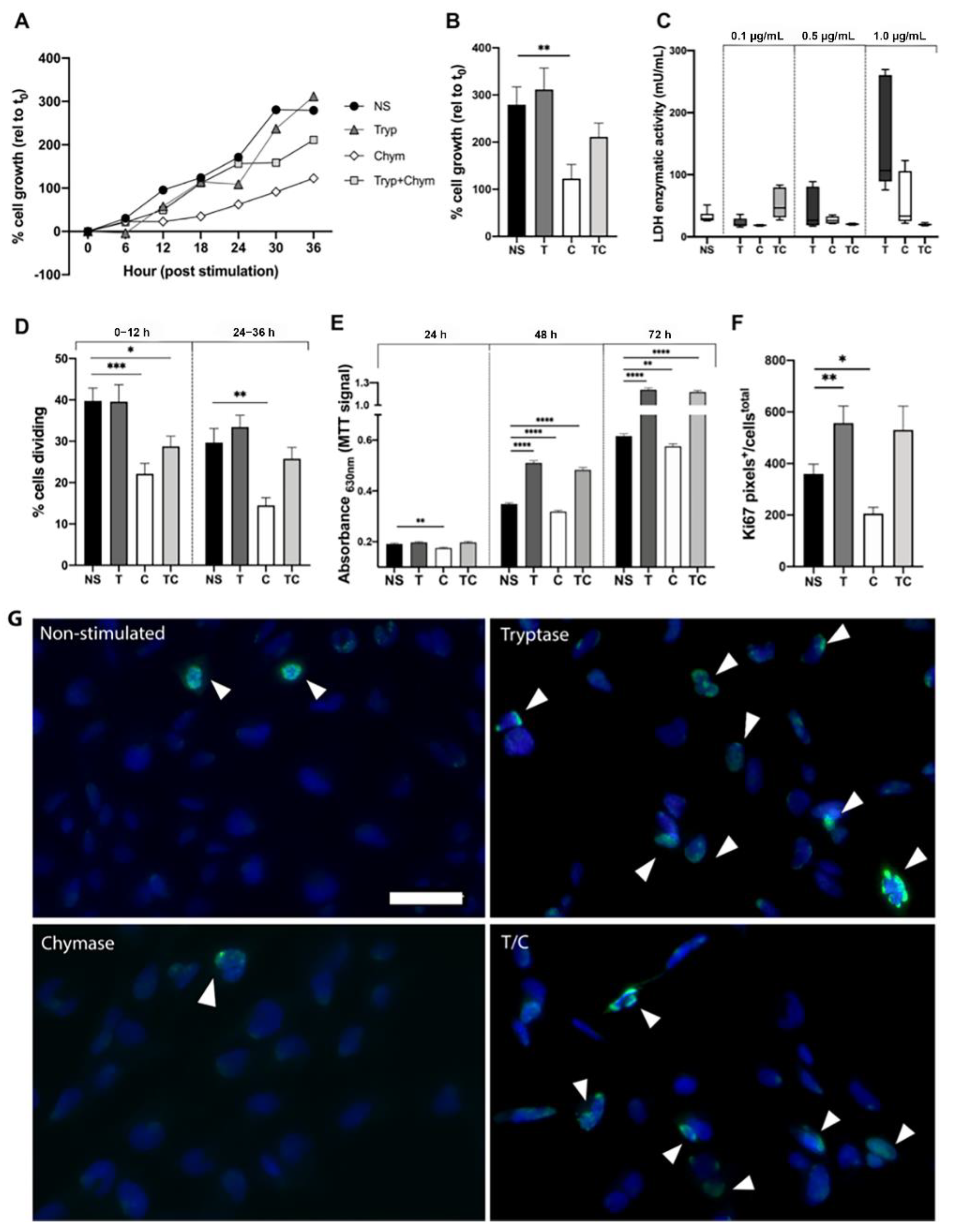
2.1. Mast Cell Proteases Alter Cell Growth and Division Rate in Bronchial Epithelial Cells
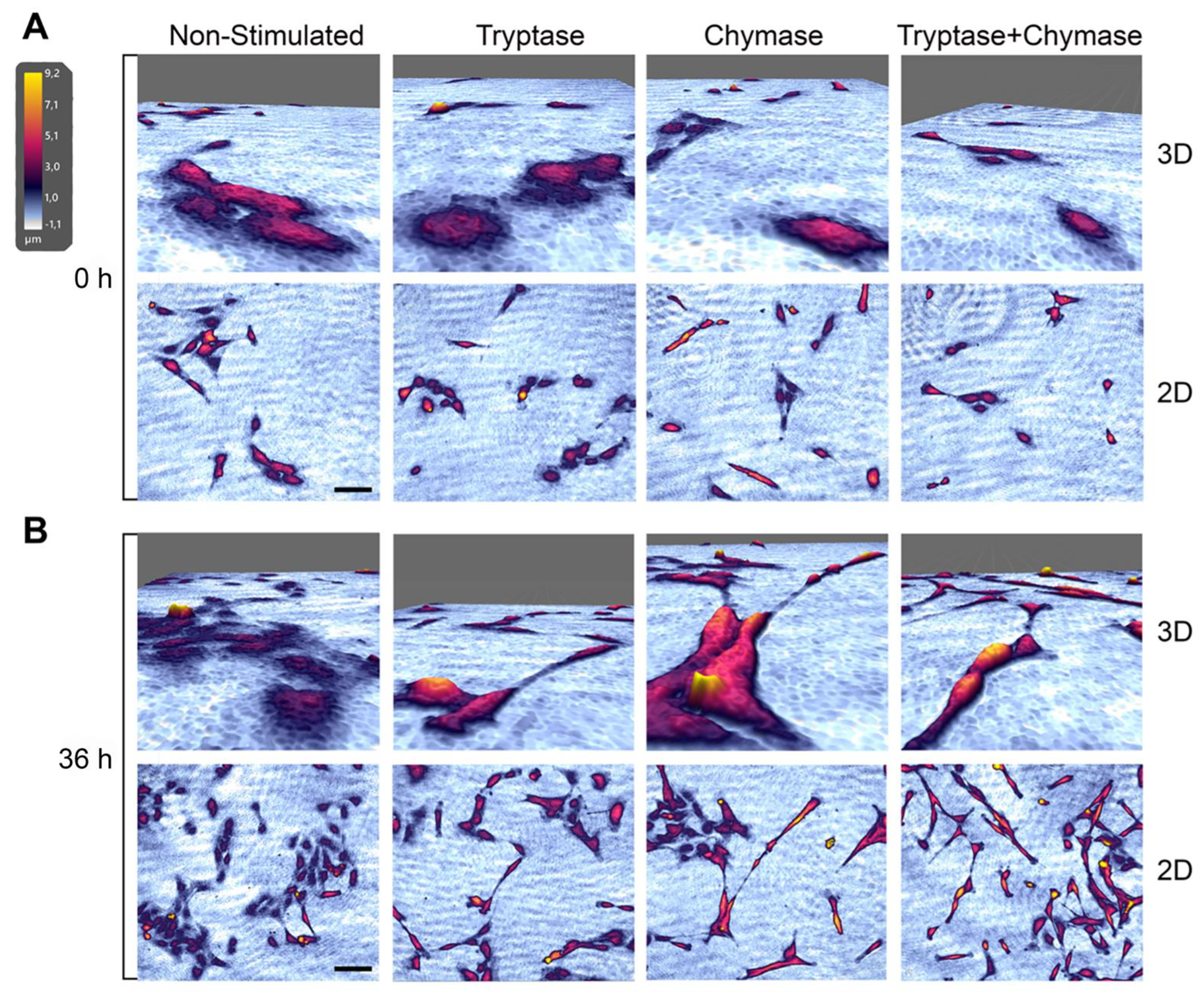
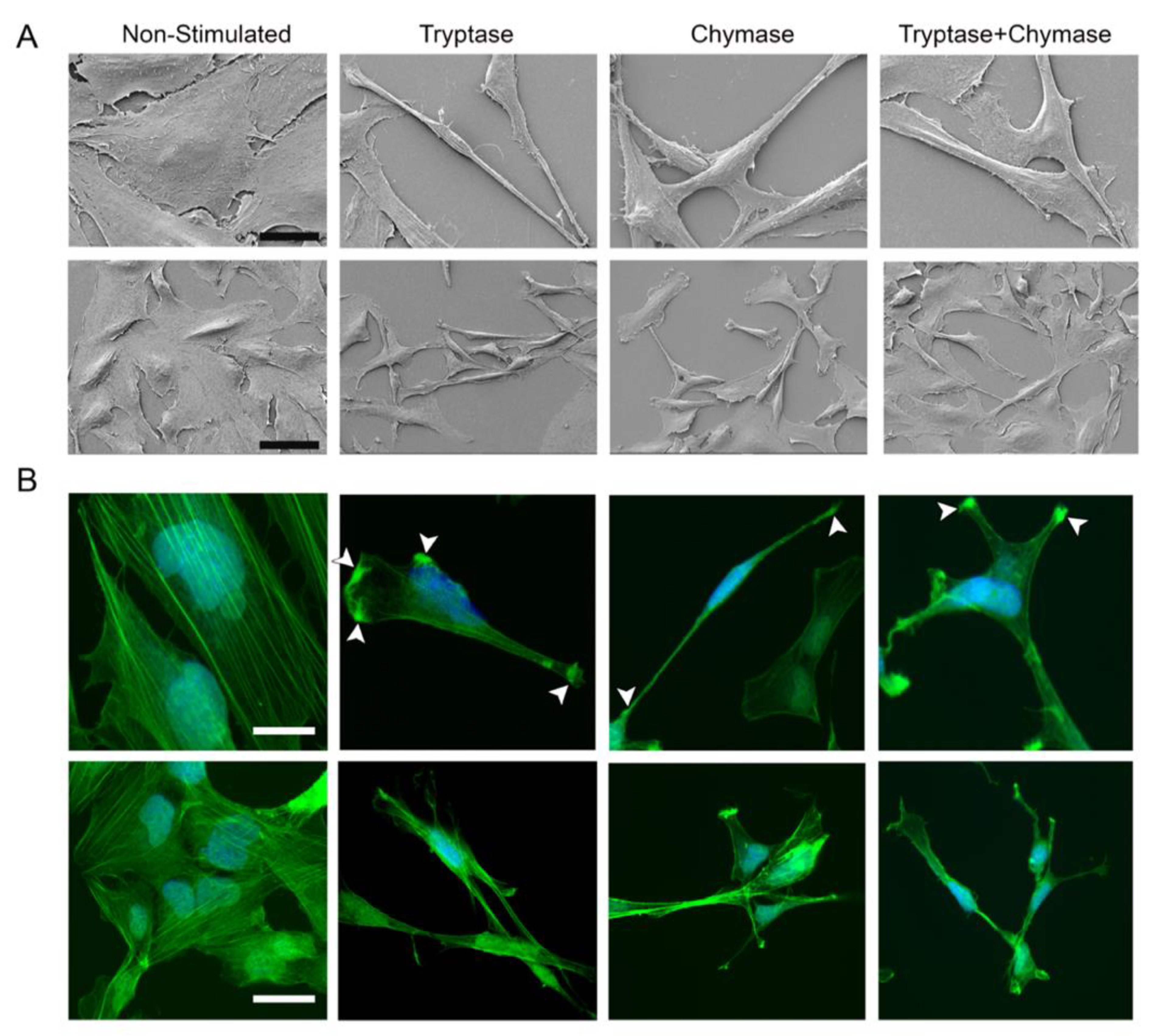
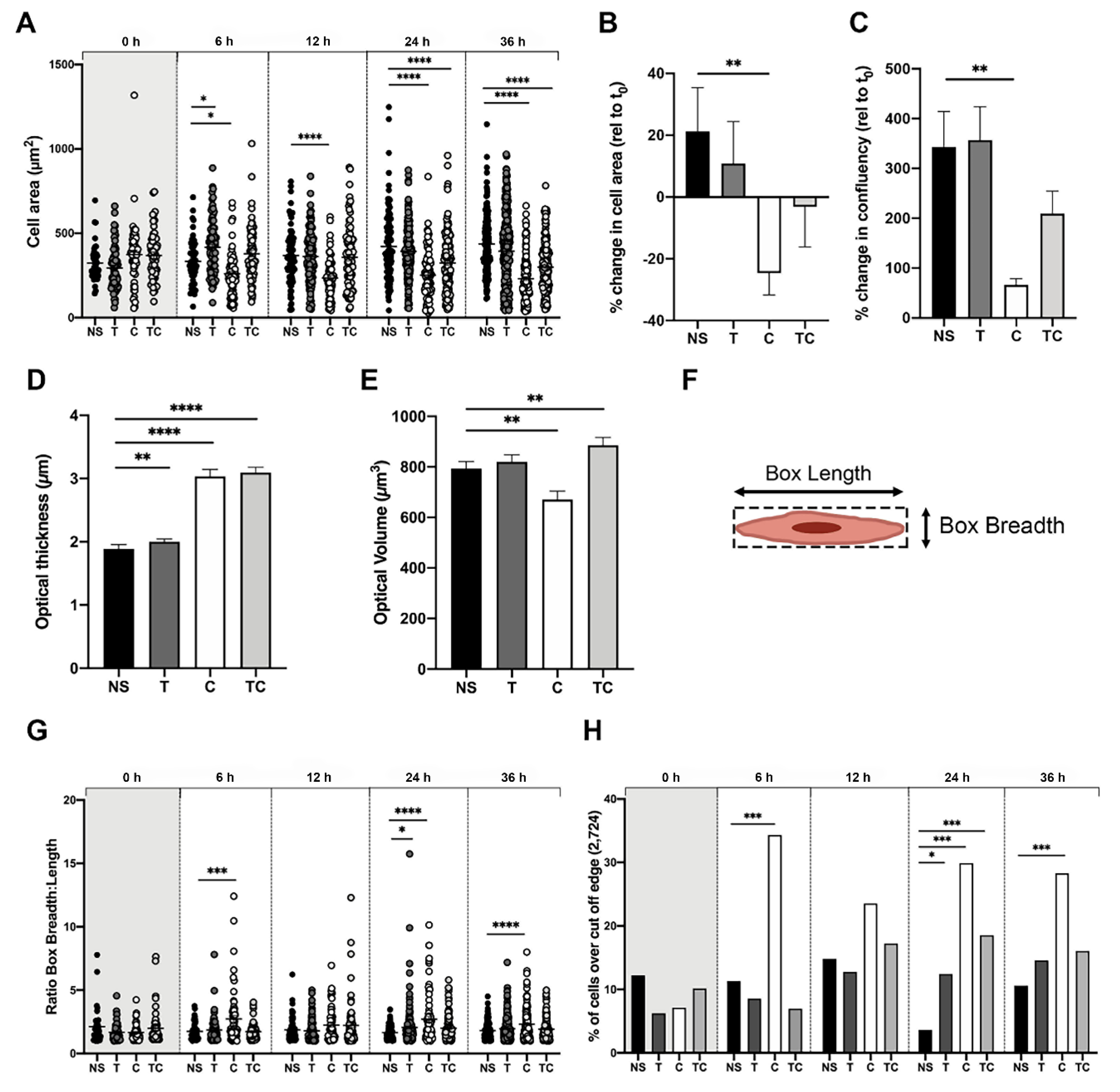
2.2. Bronchial Epithelial Cells Stimulated with MC Proteases Display Cell Elongation and Cytoskeletal Rearrangement
2.3. Tryptase and Chymase Induce Morphological Alterations in Bronchial Epithelial Cells
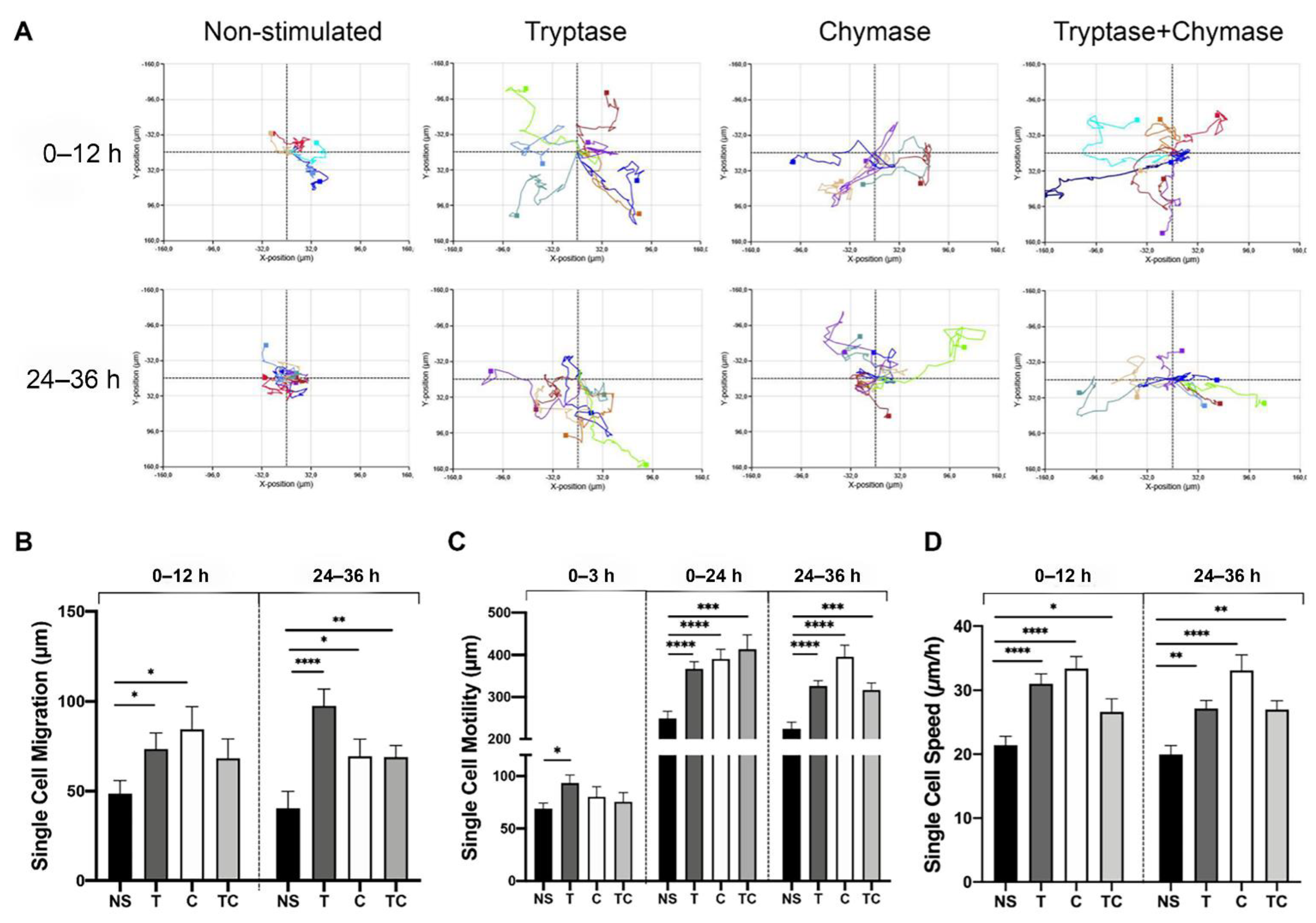
2.4. Tryptase and Chymase Induce Enhanced Cell Migration, Motility and Speed in Bronchial Epithelial Cells
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Tryptase and Chymase Activity Measurements
4.3. LDH Assay
4.4. RNA Extraction and RT2 PCR Array
4.5. MTT Assay
4.6. Immunocytochemistry
4.7. Scanning Electron Microscopy (SEM)
4.8. Live Cell Imaging: Migration, Morphology and Proliferation
- ○
- Cell growth over time (%): The cell count was based on the automatic cell identification in the HStudio and describes the number of cells per captured image. This represents the percentage cell growth relative to the starting point, which was calculated by dividing the cell count at a certain time point with the starting cell count for that specific focus point. The chosen time points were 0, 6, 12, 24 and 36 h.
- ○
- Confluency (%): The confluence was based on the cell area identification and describes the total area of a frame that is covered by cells. The confluency at 36 h was divided by the confluency at t0 and represents the increase of cell confluency at 36 h relative to starting value.
- ○
- Dividing cells: The percentage of dividing cells was obtained by calculating the number of dividing cells within the time interval divided by the starting number of cells at the beginning of the time interval.
- ○
- Cell Area (µm2): cell area was obtained using cell area identification application in HStudio and was based on calibration and calculation of single pixels. The single-cell area at different time points over 36 h as well as the relative change in cell area at 36 h relative to the starting value was plotted.
- ○
- Cell elongation: Cell elongation is represented as a ratio between box length and box breadth. It was calculated by dividing the cell box length with box breadth for each cell and then individually plotted at different time points over 36 h. The 10% percentile was calculated by collecting all cell ratios of the non-stimulated cell across all time points and analyzed in GraphPad Prism version 9.0 (GraphPad Software, La Jolla, CA, USA). The obtained value was then used as cut-off threshold, and the percentage of cells above and below this threshold was calculated and plotted.
- ○
- Optical volume (µm3): represents the single cell volume and is calculated from the phase shift and is independent of cell shape.
- ○
- Optical thickness (µm): represents the maximum cell thickness and is obtained from maximum phase shift.
- ○
- Cell movement: Cell movement is categorized both by non-directional cell motility and by directional cell migration.
- ▪
- Migration (µm): measurement of the linear distance that a cell moves between starting position A and end position B of the cell path. The parameter indicates the directness of cell movement.
- ▪
- Motility (µm): measurement of the total distance traveled per individual cell. This measurement will quantify the full path of the movement of the cell. However, it will not measure how far from the starting position the cell has moved, but rather the overall activity of movement. Cell motility is regarded as random cell movement, occurring in almost every cell culture.
- ▪
- Cell speed (µm/h): measurement of how far the cell moves in µm per time unit (hour). Data was based on the movement of a cell from one analysis frame to the next and divided by the time between the frames (15 min).
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| C | chymase |
| CF | cystic fibrosis |
| COPD | chronic obstructive pulmonary disease |
| ECM | extracellular matrix |
| EZR | ezrin |
| IPF | idiopathic pulmonary fibrosis |
| LDH | lactate dehydrogenase |
| MC | mast cell |
| MMP | matrix metalloproteinase |
| MTT | 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide |
| NS | non-stimulated |
| OD | optical density |
| PLAUR | urokinase plasminogen activator surface receptor |
| PLD1 | phospholipase D1 |
| RASA1 | RAS p21 protein activator 1 |
| RHO | transforming protein RhoA |
| SEM | scanning electron microscopy |
| STAT3 | signal transducer and activator of transcription 3 |
| T | tryptase |
| TC | tryptase-chymase |
| WASF1 | WASP-family verprolin homologous protein 1 |
References
- Bradding, P.; Walls, A.F.; Holgate, S.T. The role of the mast cell in the pathophysiology of asthma. J. Allergy Clin. Immunol. 2006, 117, 1277–1284. [Google Scholar] [CrossRef] [PubMed]
- Andersson, C.; Tufvesson, E.; Diamant, Z.; Bjermer, L. Revisiting the role of the mast cell in asthma. Curr. Opin. Pulm. Med. 2016, 22, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Metcalfe, D.D.; Baram, D.; Mekori, Y.A. Mast cells. Physiol. Rev. 1997, 77, 1033–1079. [Google Scholar] [CrossRef] [PubMed]
- Enerbäck, L.; Enerbäck, L.; Norrby, K.; Norrby, K. The Mast Cells. Curr. Top Pathol. 1989, 79, 169–204. [Google Scholar]
- Moon, T.C.; Laurent, C.D.S.; E Morris, K.; Marcet, C.; Yoshimura, T.; Sekar, Y.; Befus, A.D. Advances in mast cell biology: New understanding of heterogeneity and function. Mucosal Immunol. 2009, 3, 111–128. [Google Scholar] [CrossRef] [Green Version]
- Irani, A.A.; Schechter, N.M.; Craig, S.S.; DeBlois, G.; Schwartz, L.B. Two types of human mast cells that have distinct neutral protease compositions. Proc. Natl. Acad. Sci. USA 1986, 83, 4464–4468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersson, C.K.; Mori, M.; Bjermer, L.; Lofdahl, C.-G.; Erjefalt, J.S. Novel site-specific mast cell subpopulations in the human lung. Thorax 2009, 64, 297–305. [Google Scholar] [CrossRef] [Green Version]
- Bradding, P. Human lung mast cell heterogeneity. Thorax 2009, 64, 278–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balzar, S.; Chu, H.W.; Strand, M.; Wenzel, S. Relationship of Small Airway Chymase-Positive Mast Cells and Lung Function in Severe Asthma. Am. J. Respir. Crit. Care Med. 2005, 171, 431–439. [Google Scholar] [CrossRef]
- Fajt, M.; Balzar, S.; Wenzel, S. Increased Presence of Mast Cells in the Airway Lumen in Severe Asthma. Am. J. Respir. Crit. Care Med. 2009, 179, A2507. [Google Scholar]
- Andersson, C.K.; Mori, M.; Bjermer, L.; Löfdahl, C.-G.; Erjefält, J.S. Alterations in Lung Mast Cell Populations in Patients with Chronic Obstructive Pulmonary Disease. Am. J. Respir. Crit. Care Med. 2010, 181, 206–217. [Google Scholar] [CrossRef]
- Andersson, C.K.; Andersson-Sjöland, A.; Mori, M.; Hallgren, O.; Pardo, A.; Eriksson, L.; Bjermer, L.; Löfdahl, C.-G.; Selman, M.; Westergren-Thorsson, G.; et al. Activated MCTC mast cells infiltrate diseased lung areas in cystic fibrosis and idiopathic pulmonary fibrosis. Respir. Res. 2011, 12, 139. [Google Scholar] [CrossRef] [Green Version]
- Andersson, C.K.; Bergqvist, A.; Mori, M.; Mauad, T.; Bjermer, L.; Erjefält, J.S. Mast cell–associated alveolar inflammation in patients with atopic uncontrolled asthma. J. Allergy Clin. Immunol. 2011, 127, 905–912.e7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersson, C.K.; Tufvesson, E.; Aronsson, D.; Bergqvist, A.; Mori, M.; Bjermer, L.; Erjefält, J.S. Alveolar mast cells shift to an FcepsilonRI-expressing phenotype in mild atopic asthma: A novel feature in allergic asthma pathology. Allergy 2011, 66, 1590–1597. [Google Scholar] [CrossRef]
- Lai, Y.; Altemeier, W.A.; Vandree, J.; Piliponsky, A.M.; Johnson, B.; Appel, C.L.; Frevert, C.W.; Hyde, D.M.; Ziegler, S.F.; Smith, D.E.; et al. Increased density of intraepithelial mast cells in patients with exercise-induced bronchoconstriction regulated through epithelially derived thymic stromal lymphopoietin and IL-33. J. Allergy Clin. Immunol. 2013, 133, 1448–1455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paivandy, A.; Pejler, G. Novel Strategies to Target Mast Cells in Disease. J. Innate Immun. 2021, 13, 131–147. [Google Scholar] [CrossRef]
- Montefort, S.; Roberts, J.A.; Beasley, R.; Holgate, S.T.; Roche, W.R. The site of disruption of the bronchial epithelium in asthmatic and non-asthmatic subjects. Thorax 1992, 47, 499–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davies, D.E. The Role of the Epithelium in Airway Remodeling in Asthma. Proc. Am. Thorac. Soc. 2009, 6, 678–682. [Google Scholar] [CrossRef] [Green Version]
- Lloyd, C.M.; Saglani, S. Asthma and allergy: The emerging epithelium. Nat. Med. 2010, 16, 273–274. [Google Scholar] [CrossRef] [Green Version]
- Edwards, M.R.; Regamey, N.; Vareille, M.; Kieninger, E.; Gupta, A.; Shoemark, A.; Saglani, S.; Sykes, A.; MacIntyre, J.; Davies, J.; et al. Impaired innate interferon induction in severe therapy resistant atopic asthmatic children. Mucosal Immunol. 2012, 6, 797–806. [Google Scholar] [CrossRef] [Green Version]
- Jackson, D.J.; Makrinioti, H.; Rana, B.M.; Shamji, B.W.; Trujillo-Torralbo, M.B.; Footitt, J.; Del-Rosario, J.; Telcian, A.G.; Nikonova, A.; Zhu, J.; et al. IL-33-dependent type 2 inflammation during rhinovirus-induced asthma exacerbations in vivo. Am. J. Respir. Crit. Care Med. 2014, 190, 1373–1382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sykes, A.; Macintyre, J.; Edwards, M.R.; Del Rosario, A.; Haas, J.; Gielen, V.; Kon, O.M.; McHale, M.; Johnston, S.L. Rhinovirus-induced interferon production is not deficient in well controlled asthma. Thorax 2013, 69, 240–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farne, H.A.; Johnston, S.L. Immune mechanisms of respiratory viral infections in asthma. Curr. Opin. Immunol. 2017, 48, 31–37. [Google Scholar] [CrossRef]
- Grainge, C.L.; Davies, D.E. Epithelial Injury and Repair in Airways Diseases. Chest 2013, 144, 1906–1912. [Google Scholar] [CrossRef] [PubMed]
- Caughey, G.H. Mast cell tryptases and chymases in inflammation and host defense. Immunol. Rev. 2007, 217, 141–154. [Google Scholar] [CrossRef] [PubMed]
- Pejler, G. The emerging role of mast cell proteases in asthma. Eur. Respir. J. 2019, 54, 1900685. [Google Scholar] [CrossRef]
- Gela, A.; Kasetty, G.; Jovic, S.; Ekoff, M.; Nilsson, G.; Mörgelin, M.; Kjellström, S.; Pease, J.E.; Schmidtchen, A.; Egesten, A. Eotaxin-3 (CCL26) exerts innate host defense activities that are modulated by mast cell proteases. Allergy 2015, 70, 161–170. [Google Scholar] [CrossRef]
- Amin, K. The role of mast cells in allergic inflammation. Respir. Med. 2012, 106, 9–14. [Google Scholar] [CrossRef] [Green Version]
- Cairns, J.A.; Walls, A.F. Mast cell tryptase is a mitogen for epithelial cells. Stimulation of IL-8 production and intercellular adhesion molecule-1 expression. J. Immunol. 1996, 156, 275–283. [Google Scholar]
- Huttunen, M.; Harvima, I.T. Mast cell tryptase and chymase in chronic leg ulcers: Chymase is potentially destructive to epithelium and is controlled by proteinase inhibitors. Br. J. Dermatol. 2005, 152, 1149–1160. [Google Scholar] [CrossRef]
- Li, Q.; Jie, Y.; Wang, C.; Zhang, Y.; Guo, H.; Pan, Z. Tryptase compromises corneal epithelial barrier function. Cell Biochem. Funct. 2014, 32, 183–187. [Google Scholar] [CrossRef]
- Wilcz-Villega, E.M.; McClean, S.; O’Sullivan, M.A. Mast Cell Tryptase Reduces Junctional Adhesion Molecule-A (JAM-A) Expression in Intestinal Epithelial Cells: Implications for the Mechanisms of Barrier Dysfunction in Irritable Bowel Syndrome. Am. J. Gastroenterol. 2013, 108, 1140–1151. [Google Scholar] [CrossRef]
- Rastogi, P.; Young, D.M.; McHowat, J. Tryptase activates calcium-independent phospholipase A2 and releases PGE2 in airway epithelial cells. Am. J. Physiol. Cell. Mol. Physiol. 2008, 295, L925–L932. [Google Scholar] [CrossRef] [Green Version]
- Arai, R.; Usui-Ouchi, A.; Ito, Y.; Mashimo, K.; Murakami, A.; Ebihara, N. Effects of Secreted Mast Cell Mediators on Retinal Pigment Epithelial Cells: Focus on Mast Cell Tryptase. Mediat. Inflamm. 2017, 2017, 3124753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Metz, M.; Siebenhaar, F.; Maurer, M. Mast cell functions in the innate skin immune system. Immunobiology 2008, 213, 251–260. [Google Scholar] [CrossRef]
- Abraham, S.N.; John, A.S. Mast cell-orchestrated immunity to pathogens. Nat. Rev. Immunol. 2010, 10, 440–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hofmann, A.M.; Abraham, S.N. New roles for mast cells in pathogen defense and allergic disease. Discov. Med. 2010, 9, 79–83. [Google Scholar]
- Balzar, S.; Fajt, M.L.; Comhair, S.A.A.; Erzurum, S.C.; Bleecker, E.; Busse, W.W.; Castro, M.; Gaston, B.; Israel, E.; Schwartz, L.B.; et al. Mast Cell Phenotype, Location, and Activation in Severe Asthma. Am. J. Respir. Crit. Care Med. 2011, 183, 299–309. [Google Scholar] [CrossRef] [PubMed]
- Fajt, M.L.; Wenzel, S.E. Mast Cells, Their Subtypes, and Relation to Asthma Phenotypes. Ann. Am. Thorac. Soc. 2013, 10, S158–S164. [Google Scholar] [CrossRef] [PubMed]
- Brightling, C.E.; Bradding, P.; Symon, F.A.; Holgate, S.T.; Wardlaw, A.J.; Pavord, I.D. Mast-Cell Infiltration of Airway Smooth Muscle in Asthma. N. Engl. J. Med. 2002, 346, 1699–1705. [Google Scholar] [CrossRef]
- Zanini, A.; Chetta, A.; Saetta, M.; Baraldo, S.; D’Ippolito, R.; Castagnaro, A.; Neri, M.; Olivieri, D. Chymase-positive mast cells play a role in the vascular component of airway remodeling in asthma. J. Allergy Clin. Immunol. 2007, 120, 329–333. [Google Scholar] [CrossRef]
- Schwartz, L.B.; Irani, A.M.; Roller, K.; Castells, M.C.; Schechter, N.M. Quantitation of histamine, tryptase, and chymase in dispersed human T and TC mast cells. J. Immunol. 1987, 138, 2611–2615. [Google Scholar]
- Bergqvist, A.; Andersson, C.K.; Hoffmann, H.-J.; Mori, M.; Shikhagaie, M.; Krohn, I.K.; Dahl, R.; Bjermer, L.; Erjefält, J.S. Marked Epithelial Cell Pathology and Leukocyte Paucity in Persistently Symptomatic Severe Asthma. Am. J. Respir. Crit. Care Med. 2013, 188, 1475–1477. [Google Scholar] [CrossRef]
- Groschwitz, K.R.; Wu, D.; Osterfeld, H.; Ahrens, R.; Hogan, S.P. Chymase-mediated intestinal epithelial permeability is regulated by a protease-activating receptor/matrix metalloproteinase-2-dependent mechanism. Am. J. Physiol. Liver Physiol. 2013, 304, G479–G489. [Google Scholar] [CrossRef] [Green Version]
- Albert-Bayo, M.; Paracuellos, I.; González-Castro, A.M.; Rodríguez-Urrutia, A.; Rodríguez-Lagunas, M.J.; Alonso-Cotoner, C.; Santos, J.; Vicario, M. Intestinal Mucosal Mast Cells: Key Modulators of Barrier Function and Homeostasis. Cells 2019, 8, 135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knight, P.A.; Brown, J.K.; Pemberton, A.D. Innate immune response mechanisms in the intestinal epithelium: Potential roles for mast cells and goblet cells in the expulsion of adultTrichinella spiralis. Parasitol. 2008, 135, 655–670. [Google Scholar] [CrossRef] [PubMed]
- Terakawa, M.; Tomimori, Y.; Goto, M.; Hayashi, Y.; Oikawa, S.; Fukuda, Y. Eosinophil migration induced by mast cell chymase is mediated by extracellular signal-regulated kinase pathway. Biochem. Biophys. Res. Commun. 2005, 332, 969–975. [Google Scholar] [CrossRef] [PubMed]
- Bagher, M.; Larsson-Callerfelt, A.-K.; Rosmark, O.; Hallgren, O.; Bjermer, L.; Westergren-Thorsson, G. Mast cells and mast cell tryptase enhance migration of human lung fibroblasts through protease-activated receptor 2. Cell Commun. Signal. 2018, 16, 59. [Google Scholar] [CrossRef]
- Bagher, M.; Rosmark, O.; Rendin, L.E.; Nybom, A.; Wasserstrom, S.; Müller, C.; Zhou, X.-H.; Dellgren, G.; Hallgren, O.; Bjermer, L.; et al. Crosstalk between Mast Cells and Lung Fibroblasts Is Modified by Alveolar Extracellular Matrix and Influences Epithelial Migration. Int. J. Mol. Sci. 2021, 22, 506. [Google Scholar] [CrossRef] [PubMed]
- Ebihara, N.; Funaki, T.; Murakami, A.; Takai, S.; Miyazaki, M. Mast Cell Chymase Decreases the Barrier Function and Inhibits the Migration of Corneal Epithelial Cells. Curr. Eye Res. 2005, 30, 1061–1069. [Google Scholar] [CrossRef]
- Kofford, M.W.; Schwartz, L.B.; Schechter, N.M.; Yager, D.R.; Diegelmann, R.F.; Graham, M.F. Cleavage of Type I Procollagen by Human Mast Cell Chymase Initiates Collagen Fibril Formation and Generates a Unique Carboxyl-terminal Propeptide. J. Biol. Chem. 1997, 272, 7127–7131. [Google Scholar] [CrossRef] [Green Version]
- Tchougounova, E.; Lundequist, A.; Fajardo, I.; Winberg, J.-O.; Åbrink, M.; Pejler, G. A Key Role for Mast Cell Chymase in the Activation of Pro-matrix Metalloprotease-9 and Pro-matrix Metalloprotease-2. J. Biol. Chem. 2005, 280, 9291–9296. [Google Scholar] [CrossRef] [Green Version]
- Vartio, T.; Seppä, H.; Vaheri, A. Susceptibility of soluble and matrix fibronectins to degradation by tissue proteinases, mast cell chymase and cathepsin G. J. Biol. Chem. 1981, 256, 471–477. [Google Scholar] [CrossRef]
- Atiakshin, D.; Buchwalow, I.; Tiemann, M. Mast cell chymase: Morphofunctional characteristics. Histochem. Cell Biol. 2019, 152, 253–269. [Google Scholar] [CrossRef] [PubMed]
- Barton, S.J.; Koppelman, G.H.; Vonk, J.M.; Browning, C.A.; Nolte, I.M.; Stewart, C.E.; Bainbridge, S.; Mutch, S.; Rose-Zerilli, M.J.; Postma, D.S.; et al. PLAUR polymorphisms are associated with asthma, PLAUR levels, and lung function decline. J. Allergy Clin. Immunol. 2009, 123, 1391–1400.e17. [Google Scholar] [CrossRef]
- Stewart, C.E.; Nijmeh, H.S.; E Brightling, C.; Sayers, I. uPAR regulates bronchial epithelial repair in vitro and is elevated in asthmatic epithelium. Thorax 2012, 67, 477–487. [Google Scholar] [CrossRef] [Green Version]
- Yin, L.-M.; Duan, T.-T.; Ulloa, L.; Yang, Y.-Q. Ezrin Orchestrates Signal Transduction in Airway Cells. Rev. Physiol. Biochem. Pharmacol. 2017, 174, 1–23. [Google Scholar]
- Lessey, E.C.; Guilluy, C.; Burridge, K. From Mechanical Force to RhoA Activation. Biochemisty 2012, 51, 7420–7432. [Google Scholar] [CrossRef]
- Loirand, G. Rho Kinases in Health and Disease: From Basic Science to Translational Research. Pharmacol. Rev. 2015, 67, 1074–1095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavanaugh, K.E.; Staddon, M.F.; Munro, E.; Banerjee, S.; Gardel, M.L. RhoA Mediates Epithelial Cell Shape Changes via Mechanosensitive Endocytosis. Dev. Cell 2020, 52, 152–166.e5. [Google Scholar] [CrossRef]
- Acharya, B.R.; Espenel, C.; Libanje, F.; Raingeaud, J.; Morgan, J.; Jaulin, F.; Kreitzer, G. KIF17 regulates RhoA-dependent actin remodeling at epithelial cell-cell adhesions. J. Cell Sci. 2016, 129, 957–970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddel, R.R.; Ke, Y.; Kaighn, M.E.; Malan-Shibley, L.; Lechner, J.F.; Rhim, J.S.; Harris, C.C. Human bronchial epithelial cells neoplastically transformed by v-Ki-ras: Altered response to inducers of terminal squamous differentiation. Oncogene Res. 1988, 3, 401–408. [Google Scholar] [PubMed]
- Atiakshin, D.; Buchwalow, I.; Horny, P.; Tiemann, M. Protease profile of normal and neoplastic mast cells in the human bone marrow with special emphasis on systemic mastocytosis. Histochem Cell Biol. 2021. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Berlin, F.; Mogren, S.; Tutzauer, J.; Andersson, C.K. Mast Cell Proteases Tryptase and Chymase Induce Migratory and Morphological Alterations in Bronchial Epithelial Cells. Int. J. Mol. Sci. 2021, 22, 5250. https://doi.org/10.3390/ijms22105250
Berlin F, Mogren S, Tutzauer J, Andersson CK. Mast Cell Proteases Tryptase and Chymase Induce Migratory and Morphological Alterations in Bronchial Epithelial Cells. International Journal of Molecular Sciences. 2021; 22(10):5250. https://doi.org/10.3390/ijms22105250
Chicago/Turabian StyleBerlin, Frida, Sofia Mogren, Julia Tutzauer, and Cecilia K. Andersson. 2021. "Mast Cell Proteases Tryptase and Chymase Induce Migratory and Morphological Alterations in Bronchial Epithelial Cells" International Journal of Molecular Sciences 22, no. 10: 5250. https://doi.org/10.3390/ijms22105250