
Transcriptome Profiling of m6A mRNA Modification in Bovine Mammary Epithelial Cells Treated with Escherichia coli

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
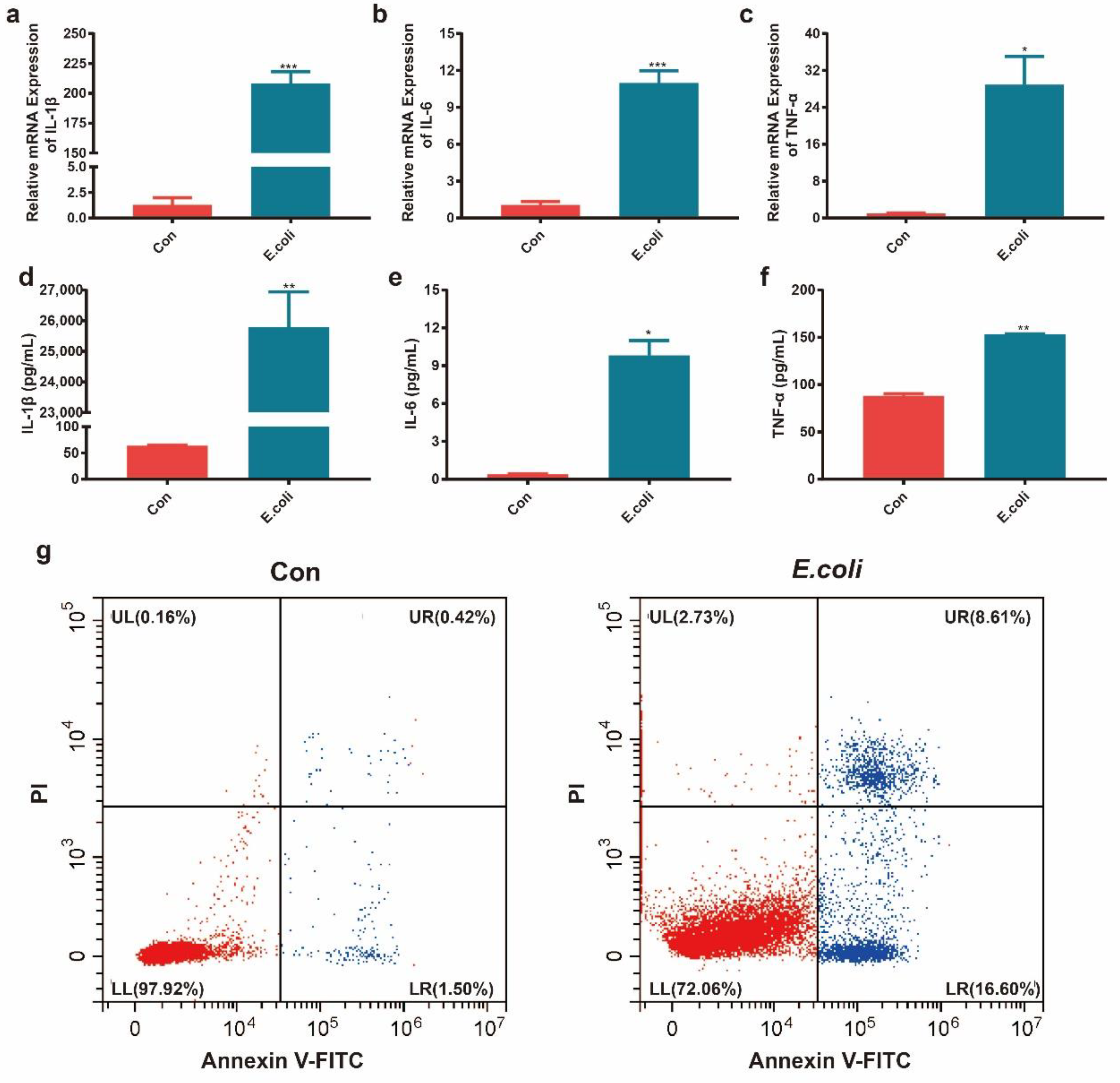
2.1. Establishment of Mastitis Model In Vitro
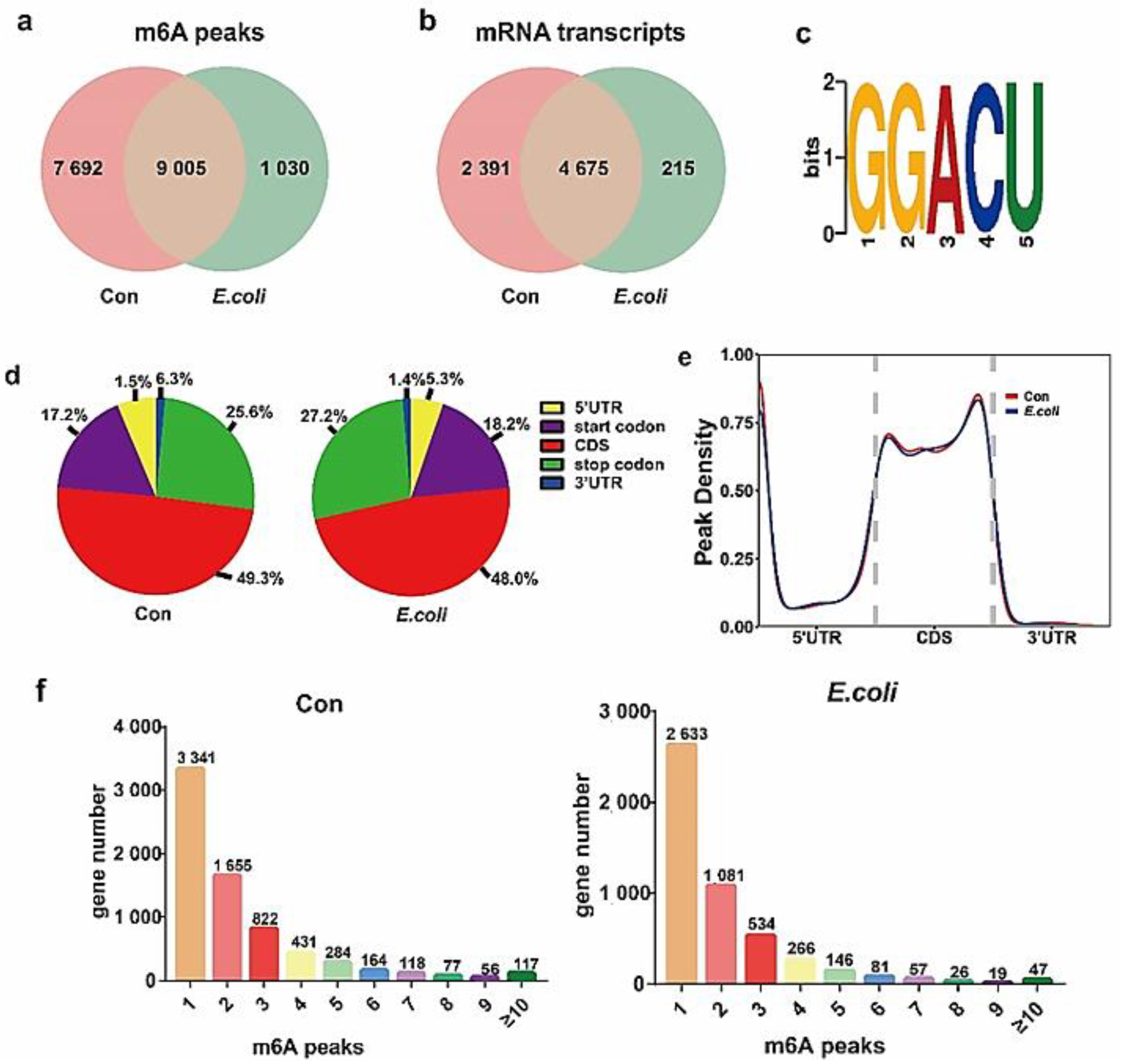
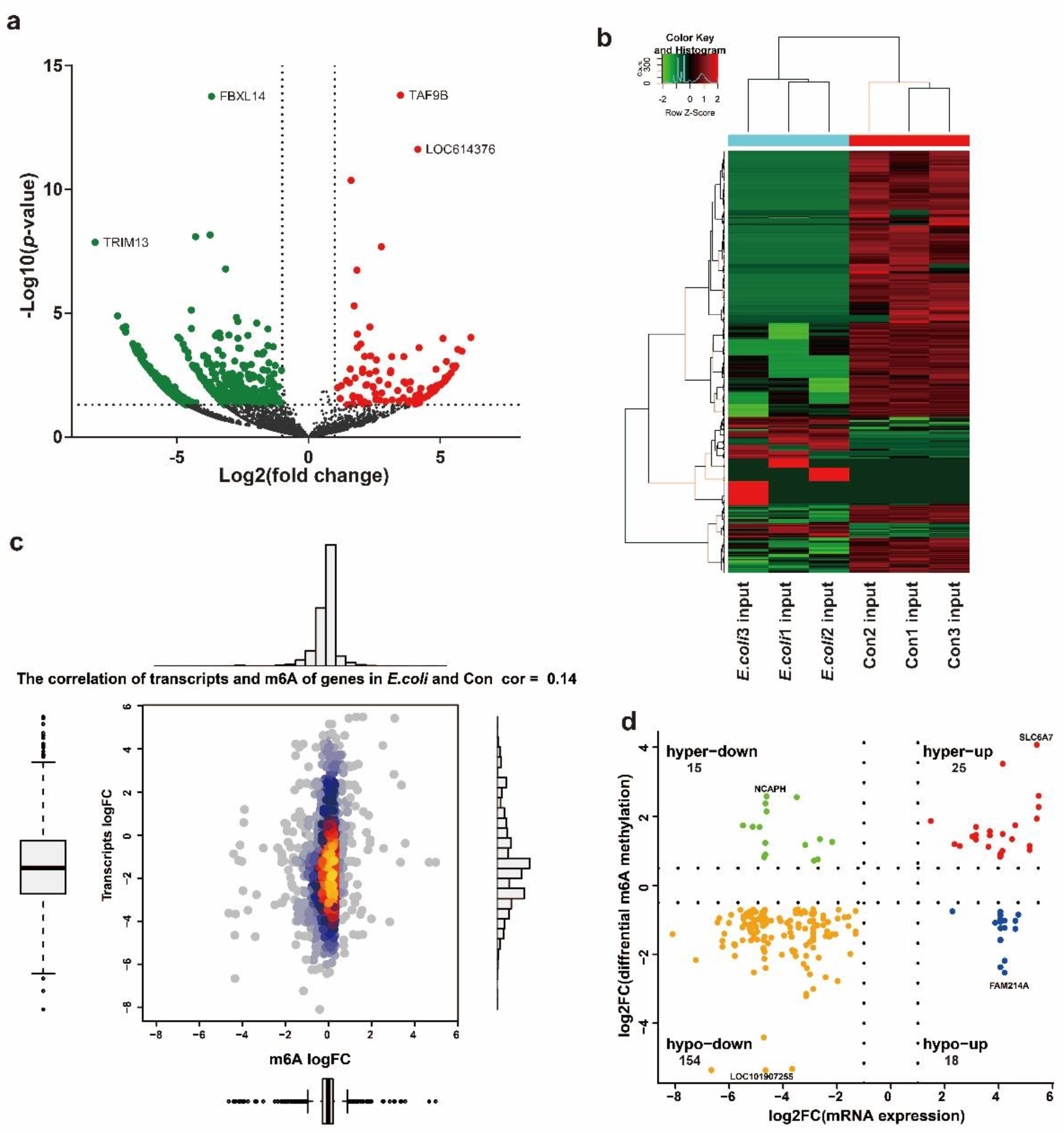
2.2. Profile of the m6A Modification in MAC-T Cells Treated with E. coli
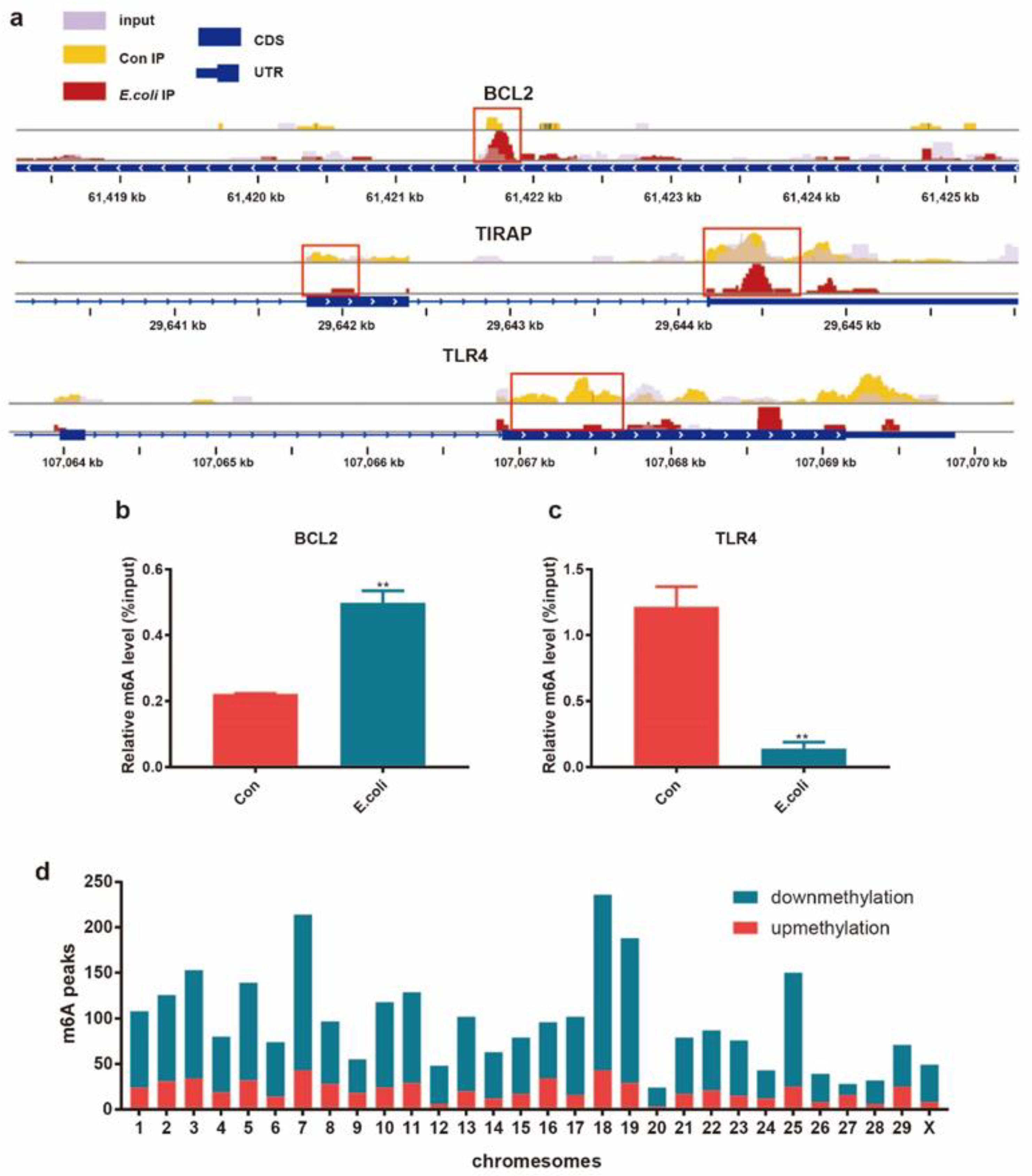
2.3. Differential m6A Modification between the E. coli Group and Control Group
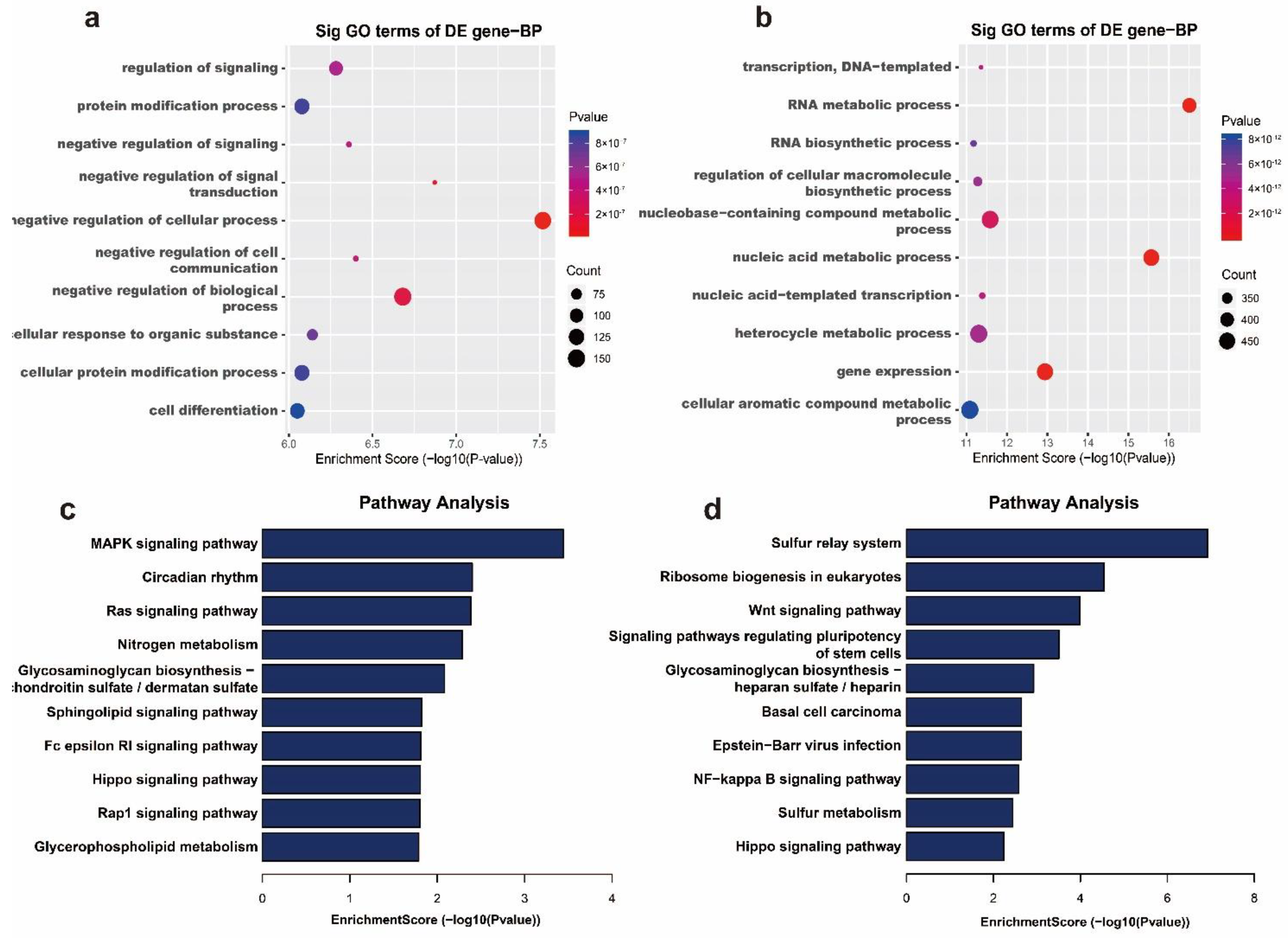
2.4. Differential m6A Modifications Participate in Important Biological Pathways
2.5. Conjoint Analysis of mRNA-Seq and MeRIP-Seq
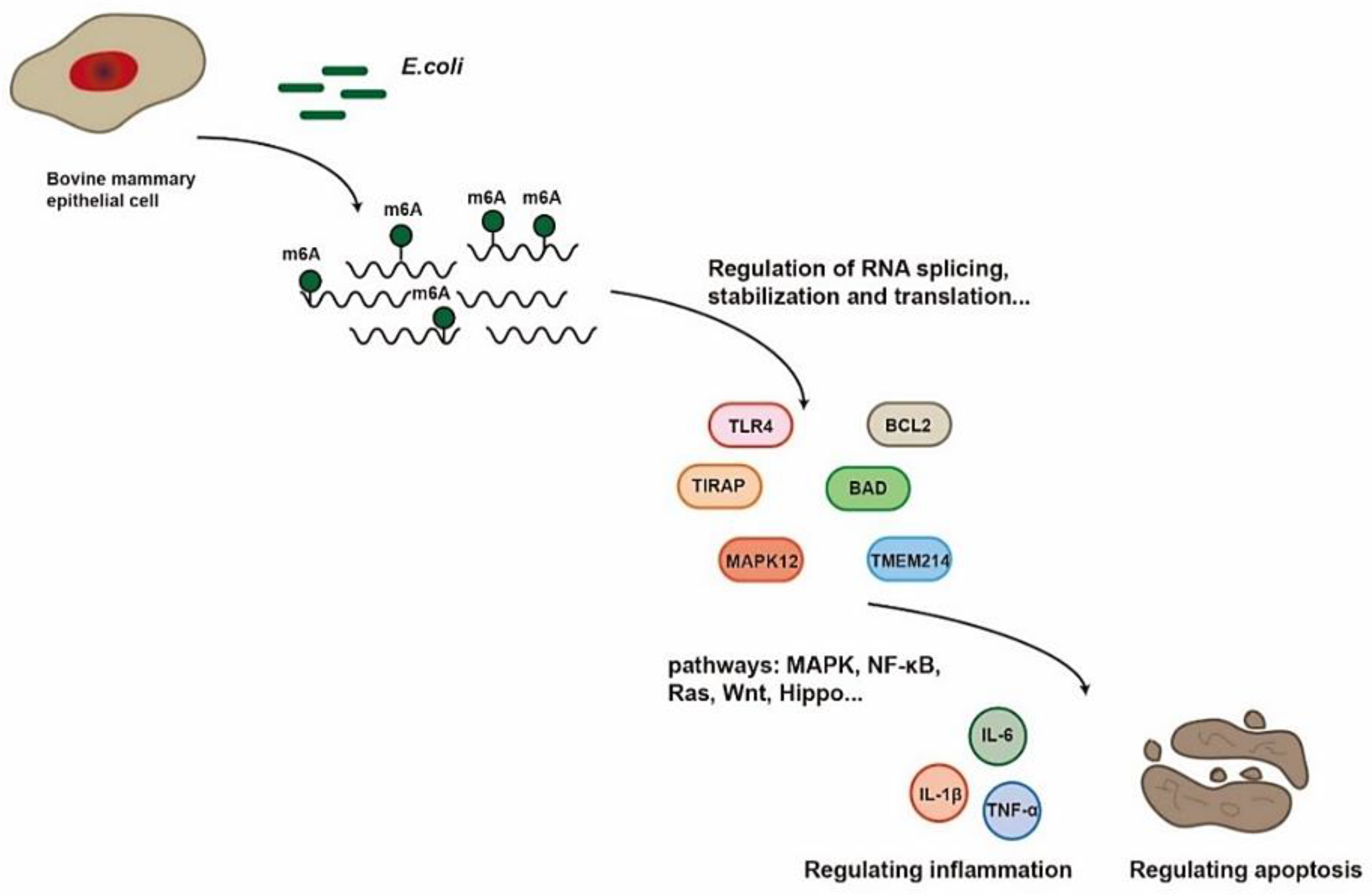
3. Discussion
4. Materials and Methods
4.1. Bacteria Strains and Cell Line
4.2. Sample Collection and RNA Extraction
4.3. Real-Time Quantitative PCR
4.4. Enzyme-Linked Immunosorbent Assay (ELISA)
4.5. Flow Cytometry
4.6. Methylated RNA Immunoprecipitation (MeRIP)
4.7. MeRIP-seq and mRNA-seq
4.8. Bioinformatics Analysis
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Monistero, V.; Graber, H.U.; Pollera, C.; Cremonesi, P.; Castiglioni, B.; Bottini, E.; Ceballos-Marquez, A.; Lasso-Rojas, L.; Kroemker, V.; Wente, N.; et al. Staphylococcus aureus Isolates from Bovine Mastitis in Eight Countries: Genotypes, Detection of Genes Encoding Different Toxins and Other Virulence Genes. Toxins 2018, 10, 247. [Google Scholar] [CrossRef] [Green Version]
- Rivas, A.L.; Tadevosyan, R.; Quimby, F.W.; Coksaygan, T.; Lein, D.H. Identification of subpopulations of bovine mammary-gland phagocytes and evaluation of sensitivity and specificity of morphologic and functional indicators of bovine mastitis. Can. J. Vet. Res. 2002, 66, 165. [Google Scholar] [PubMed]
- Günther, J.; Petzl, W.; Zerbe, H.; Schuberth, H.-J.; Koczan, D.; Goetze, L.; Seyfert, H.-M. Lipopolysaccharide priming enhances expression of effectors of immune defence while decreasing expression of pro-inflammatory cytokines in mammary epithelia cells from cows. BMC Genom. 2012, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Islam, A.; Takagi, M.; Fukuyama, K.; Komatsu, R.; Albarracin, L.; Nochi, T.; Suda, Y.; Ikeda-Ohtsubo, W.; Rutten, V.; Van Eden, W.; et al. Transcriptome Analysis of The Inflammatory Responses of Bovine Mammary Epithelial Cells: Exploring Immunomodulatory Target Genes for Bovine Mastitis. Pathogens 2020, 9, 200. [Google Scholar] [CrossRef]
- Rainard, P.; Repérant-Ferter, M.; Gitton, C.; Germon, P. Shielding Effect of Escherichia coli O-Antigen Polysaccharide on J5-Induced Cross-Reactive Antibodies. mSphere 2021, 6. [Google Scholar] [CrossRef] [PubMed]
- Vangroenweghe, F.; Lamote, I.; Burvenich, C. Physiology of the periparturient period and its relation to severity of clinical mastitis. Domest. Anim. Endocrinol. 2005, 29. [Google Scholar] [CrossRef]
- Burvenich, C.; Mehrzad, J.; Diez-Fraile, A.; Duchateau, L. Severity of E. coli mastitis is mainly determined by cow factors. Vet. Res. 2003, 34. [Google Scholar] [CrossRef] [Green Version]
- Ashraf, A.; Imran, M. Causes, types, etiological agents, prevalence, diagnosis, treatment, prevention, effects on human health and future aspects of bovine mastitis. Anim. Health Res. Rev. 2020. [Google Scholar] [CrossRef]
- Jensen, K.; Günther, J.; Talbot, R.; Petzl, W.; Zerbe, H.; Schuberth, H.-J.; Seyfert, H.-M.; Glass, E.J. Escherichia coli- and Staphylococcus aureus-induced mastitis differentially modulate transcriptional responses in neighbouring uninfected bovine mammary gland quarters. BMC Genom. 2013, 14. [Google Scholar] [CrossRef] [Green Version]
- Ibeagha-Awemu, E.; Ibeagha, A.E.; Messier, S.; Zhao, X. Proteomics, genomics, and pathway analyses of Escherichia coli and Staphylococcus aureus infected milk whey reveal molecular pathways and networks involved in mastitis. J. Proteome Res. 2010, 9. [Google Scholar] [CrossRef]
- Günther, J.; Petzl, W.; Bauer, I.; Ponsuksili, S.; Zerbe, H.; Schuberth, H.-J.; Brunner, R.M.; Seyfert, H.-M. Differentiating Staphylococcus aureus from Escherichia coli mastitis: S. aureus triggers unbalanced immune-dampening and host cell invasion immediately after udder infection. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Long, E.; Capuco, A.V.; Wood, D.L.; Sonstegard, T.; Tomita, G.; Paape, M.J.; Zhao, X. Escherichia coli induces apoptosis and proliferation of mammary cells. Cell Death Differ. 2001, 8. [Google Scholar] [CrossRef] [Green Version]
- Deng, X.; Chen, K.; Luo, G.-Z.; Weng, X.; Ji, Q.; Zhou, T.; He, C. Widespread occurrence of N6-methyladenosine in bacterial mRNA. Nucleic Acids Res. 2015, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, K.; Jaffrey, S.R. The dynamic epitranscriptome: N6-methyladenosine and gene expression control. Nat. Rev. Mol. Cell Biol. 2014, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desrosiers, R.; Friderici, K.; Rottman, F. Identification of methylated nucleosides in messenger RNA from Novikoff hepatoma cells. Proc. Natl. Acad. Sci. USA 1974, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perry, R.P.; Kelley, D.E.; Friderici, K.; Rottman, F. The methylated constituents of L cell messenger RNA: Evidence for an unusual cluster at the 5′ terminus. Cell 1975, 4. [Google Scholar] [CrossRef]
- Roundtree, I.A.; Evans, M.E.; Pan, T.; He, C. Dynamic RNA Modifications in Gene Expression Regulation. Cell 2017, 169. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Li, Y.; Toth, J.I.; Petroski, M.D.; Zhang, Z.; Zhao, J.C. N6-methyladenosine modification destabilizes developmental regulators in embryonic stem cells. Nat. Cell Biol. 2014, 16. [Google Scholar] [CrossRef]
- Liu, J.; Yue, Y.; Han, D.; Wang, X.; Fu, Y.; Zhang, L.; Jia, G.; Yu, M.; Lu, Z.; Deng, X.; et al. A METTL3-METTL14 complex mediates mammalian nuclear RNA N6-adenosine methylation. Nat. Chem. Biol. 2014, 10. [Google Scholar] [CrossRef] [Green Version]
- Wu, B.; Li, L.; Huang, Y.; Ma, J.; Min, J. Readers, writers and erasers of N(6)-methylated adenosine modification. Curr. Opin. Struct. Biol. 2017, 47. [Google Scholar] [CrossRef]
- Shi, H.; Zhang, X.; Weng, Y.-L.; Lu, Z.; Liu, Y.; Lu, Z.; Li, J.; Hao, P.; Zhang, Y.; Zhang, F.; et al. m(6)A facilitates hippocampus-dependent learning and memory through YTHDF1. Nature 2018, 563. [Google Scholar] [CrossRef]
- Visvanathan, A.; Somasundaram, K. mRNA Traffic Control Reviewed: N6-Methyladenosine (m(6) A) Takes the Driver’s Seat. Bioessays 2018, 40. [Google Scholar] [CrossRef]
- Laffleur, B.; Basu, U. Biology of RNA Surveillance in Development and Disease. Trends Cell Biol. 2019, 29. [Google Scholar] [CrossRef]
- Chen, Y.G.; Chen, R.; Ahmad, S.; Verma, R.; Kasturi, S.P.; Amaya, L.; Broughton, J.P.; Kim, J.; Cadena, C.; Pulendran, B.; et al. N6-Methyladenosine Modification Controls Circular RNA Immunity. Mol. Cell 2019, 76. [Google Scholar] [CrossRef] [PubMed]
- Yu, R.; Li, Q.; Feng, Z.; Cai, L.; Xu, Q. m6A Reader YTHDF2 Regulates LPS-Induced Inflammatory Response. Int. J. Mol. Sci. 2019, 20, 1323. [Google Scholar] [CrossRef] [Green Version]
- Zhu, S.; Lu, Y. Dexmedetomidine Suppressed the Biological Behavior of HK-2 Cells Treated with LPS by Down-Regulating ALKBH5. Inflammation 2020, 43. [Google Scholar] [CrossRef]
- Wei, C.M.; Gershowitz, A.; Moss, B. 5′-Terminal and internal methylated nucleotide sequences in HeLa cell mRNA. Biochemistry 1976, 15. [Google Scholar] [CrossRef] [PubMed]
- Schibler, U.; Kelley, D.E.; Perry, R.P. Comparison of methylated sequences in messenger RNA and heterogeneous nuclear RNA from mouse L cells. J. Mol. Biol. 1977, 115. [Google Scholar] [CrossRef]
- Zhang, H.; Shi, X.; Huang, T.; Zhao, X.; Chen, W.; Gu, N.; Zhang, R. Dynamic landscape and evolution of m6A methylation in human. Nucleic Acids Res. 2020, 48. [Google Scholar] [CrossRef] [PubMed]
- Petzl, W.; Zerbe, H.; Günther, J.; Seyfert, H.-M.; Hussen, J.; Schuberth, H.-J. Pathogen-specific responses in the bovine udder. Models and immunoprophylactic concepts. Res. Vet. Sci. 2018, 116. [Google Scholar] [CrossRef]
- Zhang, S.-Y.; Zhang, S.-W.; Fan, X.-N.; Meng, J.; Chen, Y.; Gao, S.-J.; Huang, Y. Global analysis of N6-methyladenosine functions and its disease association using deep learning and network-based methods. PLoS Comput. Biol. 2019, 15. [Google Scholar] [CrossRef] [Green Version]
- Siddiqui, W.A.; Ahad, A.; Ahsan, H. The mystery of BCL2 family: Bcl-2 proteins and apoptosis: An update. Arch. Toxicol. 2015, 89. [Google Scholar] [CrossRef] [PubMed]
- Kale, J.; Osterlund, E.J.; Andrews, D. BCL-2 family proteins: Changing partners in the dance towards death. Cell Death Differ. 2018, 25. [Google Scholar] [CrossRef] [Green Version]
- Liang, Y.-J.; Yang, W.-X. Kinesins in MAPK cascade: How kinesin motors are involved in the MAPK pathway? Gene 2019, 684. [Google Scholar] [CrossRef] [PubMed]
- Egaestel, M. MAPK-Activated Protein Kinases (MKs): Novel Insights and Challenges. Front. Cell Dev. Biol. 2015, 3. [Google Scholar] [CrossRef] [Green Version]
- Kawai, T.; Akira, S. TLR signaling. Semin. Immunol. 2007, 19. [Google Scholar] [CrossRef]
- Ryu, J.-K.; Kim, S.J.; Rah, S.-H.; Kang, J.I.; Jung, H.E.; Lee, D.; Lee, H.K.; Lee, J.-O.; Park, B.S.; Yoon, T.-Y.; et al. Reconstruction of LPS Transfer Cascade Reveals Structural Determinants within LBP, CD14, and TLR4-MD2 for Efficient LPS Recognition and Transfer. Immunity 2017, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sui, X.; Kong, N.; Ye, L.; Han, W.; Zhou, J.; Zhang, Q.; He, C.; Pan, H. p38 and JNK MAPK pathways control the balance of apoptosis and autophagy in response to chemotherapeutic agents. Cancer Lett. 2014, 344. [Google Scholar] [CrossRef]
- Yue, J.; López, J.M. Understanding MAPK Signaling Pathways in Apoptosis. Int. J. Mol. Sci. 2020, 21, 2346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oeckinghaus, A.; Hayden, M.; Ghosh, S. Crosstalk in NF-κB signaling pathways. Nat. Immunol. 2011, 12. [Google Scholar] [CrossRef]
- Faul, F.; Erdfelder, E.; Lang, A.-G.; Buchner, A. G*Power 3: A flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Behav. Res. Methods 2007, 39. [Google Scholar] [CrossRef]
- Berulava, T.; Buchholz, E.; Elerdashvili, V.; Pena, T.; Islam, R.; Lbik, D.; Mohamed, B.A.; Renner, A.; Von Lewinski, D.; Sacherer, M.; et al. Changes in m6A RNA methylation contribute to heart failure progression by modulating translation. Eur. J. Heart Fail. 2020, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danial, N.N.; Walensky, L.D.; Zhang, C.-Y.; Choi, C.S.; Fisher, J.K.; Molina, A.J.A.; Datta, S.R.; Pitter, K.L.; Bird, G.H.; Wikstrom, J.D.; et al. Dual role of proapoptotic BAD in insulin secretion and beta cell survival. Nat. Med. 2008, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sastry, K.S.R.; Al-Muftah, M.A.; Li, P.; Al-Kowari, M.K.; Wang, E.; Chouchane, A.I.; Kizhakayil, D.; Kulik, G.; Marincola, F.M.; Haoudi, A.; et al. Targeting proapoptotic protein BAD inhibits survival and self-renewal of cancer stem cells. Cell Death Differ. 2014, 21. [Google Scholar] [CrossRef] [Green Version]
- Yan, J.; Zhang, H.; Xiang, J.; Zhao, Y.; Yuan, X.; Sun, B.; Lin, A. The BH3-only protein BAD mediates TNFα cytotoxicity despite concurrent activation of IKK and NF-κB in septic shock. Cell Res. 2018, 28. [Google Scholar] [CrossRef]
- Li, C.; Wei, J.; Li, Y.; He, X.; Zhou, Q.; Yan, J.; Zhang, J.; Liu, Y.; Shu, H.-B. Transmembrane Protein 214 (TMEM214) mediates endoplasmic reticulum stress-induced caspase 4 enzyme activation and apoptosis. J. Biol. Chem. 2013, 288. [Google Scholar] [CrossRef] [Green Version]
- Niu, Y.; Zhao, X.; Wu, Y.-S.; Li, M.-M.; Wang, X.-J.; Yang, Y.-G. N6-methyl-adenosine (m6A) in RNA: An old modification with a novel epigenetic function. Genom. Proteom. Bioinform. 2013, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, X.; Chai, G.; Wu, Y.; Li, J.; Chen, F.; Liu, J.; Luo, G.; Tauler, J.; Du, J.; Lin, S.; et al. RNA m(6)A methylation regulates the epithelial mesenchymal transition of cancer cells and translation of Snail. Nat. Commun. 2019, 10. [Google Scholar] [CrossRef]
- Gilbert, F.B.; Cunha, P.; Jensen, K.; Glass, E.J.; Foucras, G.; Robert-Granié, C.; Rupp, R.; Rainard, P. Differential response of bovine mammary epithelial cells to Staphylococcus aureus or Escherichia coli agonists of the innate immune system. Vet. Res. 2013, 44. [Google Scholar] [CrossRef] [Green Version]
- Jin, W.; Ibeagha-Awemu, E.M.; Liang, G.; Beaudoin, F.; Zhao, X.; Guan, L.L. Transcriptome microRNA profiling of bovine mammary epithelial cells challenged with Escherichia coli or Staphylococcus aureus bacteria reveals pathogen directed microRNA expression profiles. BMC Genom. 2014, 15. [Google Scholar] [CrossRef] [Green Version]
- Günther, J.; Koczan, D.; Yang, W.; Nürnberg, G.; Repsilber, D.; Schuberth, H.-J.; Park, Z.; Maqbool, N.; Molenaar, A.; Seyfert, H.-M. Assessment of the immune capacity of mammary epithelial cells: Comparison with mammary tissue after challenge with Escherichia coli. Vet. Res. 2009, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Günther, J.; Koy, M.; Berthold, A.; Schuberth, H.-J.; Seyfert, H.-M. Comparison of the pathogen species-specific immune response in udder derived cell types and their models. Vet. Res. 2016, 47. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Liu, T.; Meyer, C.A.; Eeckhoute, J.; Johnson, D.S.; Bernstein, B.E.; Nusbaum, C.; Myers, R.M.; Brown, M.; Li, W.; et al. Model-based analysis of ChIP-Seq (MACS). Genome Biol. 2008, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thorvaldsdóttir, H.; Robinson, J.T.; Mesirov, J.P. Integrative Genomics Viewer (IGV): High-performance genomics data visualization and exploration. Brief Bioinform 2013, 14. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Peak Region | Peak Start | Peak End | Chromosome | Log10 (p-Value) | Log2 (Fold Change) | Regulation |
|---|---|---|---|---|---|---|---|
| TOR1AIP1 | startC | 61,095,201 | 61,095,890 | NC_037343.1 | −9.64 | 1.09 | hyper |
| SYTL2 | CDS | 9,831,641 | 9,831,860 | NC_037356.1 | −9.42 | 1.21 | hyper |
| HERC6 | startC | 36,366,461 | 36,366,789 | NC_037333.1 | −8.98 | 1.44 | hyper |
| BHLHE41 | startC | 83,818,001 | 83,818,465 | NC_037332.1 | −8.9 | 2.57 | hyper |
| GPR155 | CDS | 22,358,286 | 22,358,700 | NC_037329.1 | −8.84 | 2.45 | hyper |
| IGFBP5 | stopC | 104,660,241 | 104,660,804 | NC_037329.1 | −8.82 | 1.44 | hyper |
| TSPAN18 | 3′UTR | 75,001,041 | 75,002,400 | NC_037342.1 | −8.68 | 1.74 | hyper |
| CENPU | 3′UTR | 15,119,101 | 15,119,520 | NC_037354.1 | −8.64 | 1.73 | hyper |
| AKAP13 | CDS | 16,514,463 | 16,514,712 | NC_037348.1 | −8.6 | 1.08 | hyper |
| RFX2 | 3′UTR | 18,396,043 | 18,397,660 | NC_037334.1 | −8.58 | 1.03 | hyper |
| CLCA2 | CDS | 57,619,160 | 57,619,431 | NC_037330.1 | −10.75 | 7.39 | hypo |
| NUDT19 | 3′UTR | 43,175,361 | 43,175,740 | NC_037345.1 | −10.52 | 4.43 | hypo |
| FZD2 | CDS | 44,329,981 | 44,330,600 | NC_037346.1 | −10.50 | 4.07 | hypo |
| LOC101907255 | 5′UTR | 41,829,801 | 41,830,640 | NC_037353.1 | −10.32 | 7.74 | hypo |
| LOC101903326 | CDS | 1,089,721 | 1,090,900 | NC_037341.1 | −10.32 | 5.05 | hypo |
| HGFAC | CDS | 115,444,475 | 115,444,760 | NC_037333.1 | −10.27 | 2.78 | hypo |
| GPR156 | CDS | 65,018,452 | 65,018,905 | NC_037328.1 | −10.11 | 3.88 | hypo |
| AOAH | CDS | 60,547,451 | 60,547,800 | NC_037331.1 | −10.11 | 3.33 | hypo |
| AAR2 | 3′UTR | 65,379,401 | 65,379,780 | NC_037340.1 | −10.09 | 3.98 | hypo |
| STRBP | 3′UTR | 94,269,361 | 94,269,860 | NC_037338.1 | −10.09 | 4.54 | hypo |
| Gene Name | Log2 (Fold Change) | Log10 (p-Value) | Regulation |
|---|---|---|---|
| IER3 | 6.164040553 | −4.02169998 | up |
| PTGS2 | 5.826757014 | −3.467281725 | up |
| ERO1A | 5.669337971 | −3.536666229 | up |
| PRDX5 | 5.586646738 | −2.881586777 | up |
| HNRNPC | 5.556401381 | −2.802666029 | up |
| CA4 | 5.481535598 | −2.86568667 | up |
| SLC6A7 | 5.414589103 | −2.589570251 | up |
| TMCC3 | 5.325086008 | −2.488894255 | up |
| DDIT4 | 5.280240307 | −2.400378604 | up |
| SLC2A1 | 5.237809067 | −3.050012716 | up |
| TRIM13 | −8.101651804 | −7.866625717 | down |
| CD70 | −7.242189071 | −4.903814594 | down |
| DDX39A | −7.027795258 | −4.414552565 | down |
| YEATS2 | −6.971898132 | −4.337112177 | down |
| RASGRP1 | −6.946317893 | −4.286252914 | down |
| PRPF38A | −6.946269946 | −4.239872373 | down |
| CELSR3 | −6.935875444 | −4.464239943 | down |
| AQP11 | −6.664329625 | −3.773669927 | down |
| GAPDHS | −6.638835912 | −3.724064388 | down |
| LOC112442013 | −6.591469341 | −3.591651156 | down |
| Gene Name | Change | Chromosome | m6A Modification Change | mRNA Expression Change | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Peak Start | Peak End | Peak Length | Peak Region | logFC | Log10 (p-Value) | logFC | p-Value | Strand | |||
| MAP3K2 | hyper-up | NC_037329.1 | 5,150,437 | 5,150,820 | 383 | 5′UTR | 2.49 | −6.97 | 4.071 | 0.021139 | + |
| ECI1 | hyper-up | NC_037352.1 | 1,765,100 | 1,765,420 | 320 | CDS | 2.33 | −5.30 | 4.07 | 0.023957 | + |
| ALK | hyper-up | NC_037338.1 | 70,660,381 | 70,661,760 | 1379 | CDS | 1.98 | −6.28 | 3.11 | 0.002504 | − |
| CA4 | hyper-up | NC_037346.1 | 12,807,579 | 12,807,920 | 341 | CDS | 13.42 | −7.90 | 5.48 | 0.001362 | + |
| SEMA6B | hyper-up | NC_037334.1 | 19,583,616 | 19,585,160 | 1544 | CDS | 3.81 | −6.16 | 4.49 | 0.022398 | + |
| HK1 | hyper-down | NC_037355.1 | 25,767,278 | 25,767,520 | 242 | CDS | 1.84 | −5.07 | −4.66 | 0.047763 | + |
| GAB2 | hyper-down | NC_037356.1 | 17,787,967 | 17,788,070 | 103 | 3′UTR | 1.85 | −5.68 | −2.71 | 0.042939 | + |
| MAP2K1 | hyper-down | NC_037337.1 | 13,276,353 | 13,276,712 | 359 | 5′UTR | 3.52 | −5.06 | −2.18 | 0.023033 | + |
| CTSB | hyper-down | NC_037335.1 | 7,566,076 | 7,566,228 | 152 | 5′UTR | 1.73 | −5.81 | −5.22 | 0.015551 | + |
| EIF2AK4 | hyper-down | NC_037337.1 | 35,645,637 | 35,645,717 | 80 | CDS | 1.69 | −5.17 | −3.56 | 0.00288 | + |
| USP1 | hypo-down | NC_037330.1 | 83,118,461 | 83,118,920 | 459 | 3′UTR | 1.60 | −6.82 | −1.67 | 0.023734 | − |
| DDX58 | hypo-down | NC_037335.1 | 11,604,221 | 11,604,700 | 479 | 3′UTR | 3.30 | −8.74 | −5.00 | 0.021294 | + |
| FBN1 | hypo-down | NC_037337.1 | 61,917,881 | 61,918,380 | 499 | CDS | 4.14 | −7.63 | −2.17 | 0.001185 | + |
| BAD | hypo-down | NC_037356.1 | 42,573,697 | 42,574,197 | 500 | 3′UTR | 1.56 | −5.22 | −4.35 | 0.045042 | − |
| MAPK12 | hypo-down | NC_037332.1 | 119,619,701 | 119,620,440 | 739 | 3′UTR | 6.15 | −6.27 | −4.73 | 0.043358 | − |
| TMEM214 | hypo-up | NC_037338.1 | 72,593,801 | 72,594,100 | 299 | 3′UTR | 1.60 | −7.89 | 4.14 | 0.042666 | − |
| STAT2 | hypo-up | NC_037332.1 | 57,007,910 | 57,008,239 | 329 | CDS | 2.11 | −7.11 | 2.28 | 0.046055 | + |
| ACOX3 | hypo-up | NC_037333.1 | 114,662,000 | 114,662,840 | 840 | CDS | 2.14 | −5.17 | 4.06 | 0.045482 | − |
| RASSF6 | hypo-up | NC_037333.1 | 88,624,201 | 88,624,540 | 339 | 3′UTR | 2.79 | −7.76 | 4.61 | 0.017354 | − |
| ZNF385A | hypo-up | NC_037332.1 | 25,706,604 | 25,707,959 | 1355 | CDS | 2.87 | −6.82 | 4.06 | 0.034321 | + |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, T.; Lin, C.; Zhu, Y.; Xu, H.; Yin, Y.; Wang, C.; Tang, X.; Song, T.; Guo, A.; Chen, Y.; et al. Transcriptome Profiling of m6A mRNA Modification in Bovine Mammary Epithelial Cells Treated with Escherichia coli. Int. J. Mol. Sci. 2021, 22, 6254. https://doi.org/10.3390/ijms22126254
Li T, Lin C, Zhu Y, Xu H, Yin Y, Wang C, Tang X, Song T, Guo A, Chen Y, et al. Transcriptome Profiling of m6A mRNA Modification in Bovine Mammary Epithelial Cells Treated with Escherichia coli. International Journal of Molecular Sciences. 2021; 22(12):6254. https://doi.org/10.3390/ijms22126254
Chicago/Turabian StyleLi, Ting, Changjie Lin, Yifan Zhu, Haojun Xu, Yiya Yin, Chaohao Wang, Xin Tang, Tongxing Song, Aizhen Guo, Yingyu Chen, and et al. 2021. "Transcriptome Profiling of m6A mRNA Modification in Bovine Mammary Epithelial Cells Treated with Escherichia coli" International Journal of Molecular Sciences 22, no. 12: 6254. https://doi.org/10.3390/ijms22126254