
Pituitary Adenylate Cyclase-Activating Polypeptide (PACAP) in Physiological and Pathological Processes within the Gastrointestinal Tract: A Review


Abstract
:1. Introduction
2. Localization of PACAP in the Gastrointestinal Tract in Individual Species of Mammals
2.1. Rodents
2.2. Pigs
2.3. Humans
2.4. Other Species
3. Co-Localization of PACAP with Other Neuroactive Substance in the GI Tract
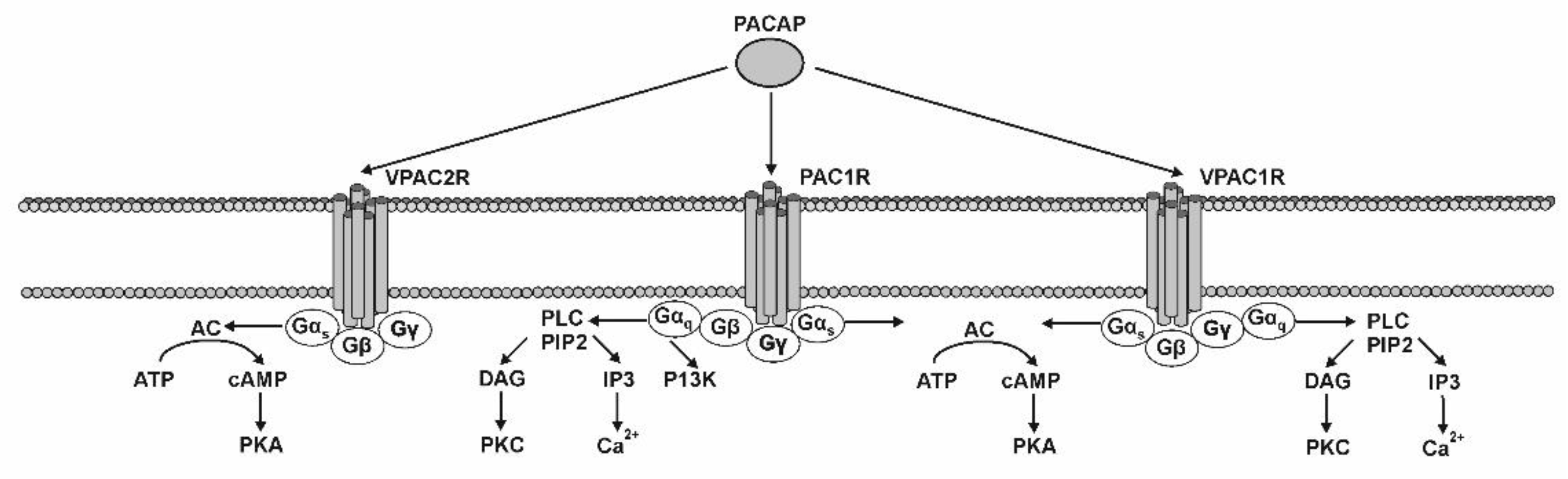
4. PACAP Receptors and Their Localization in the GI Tract
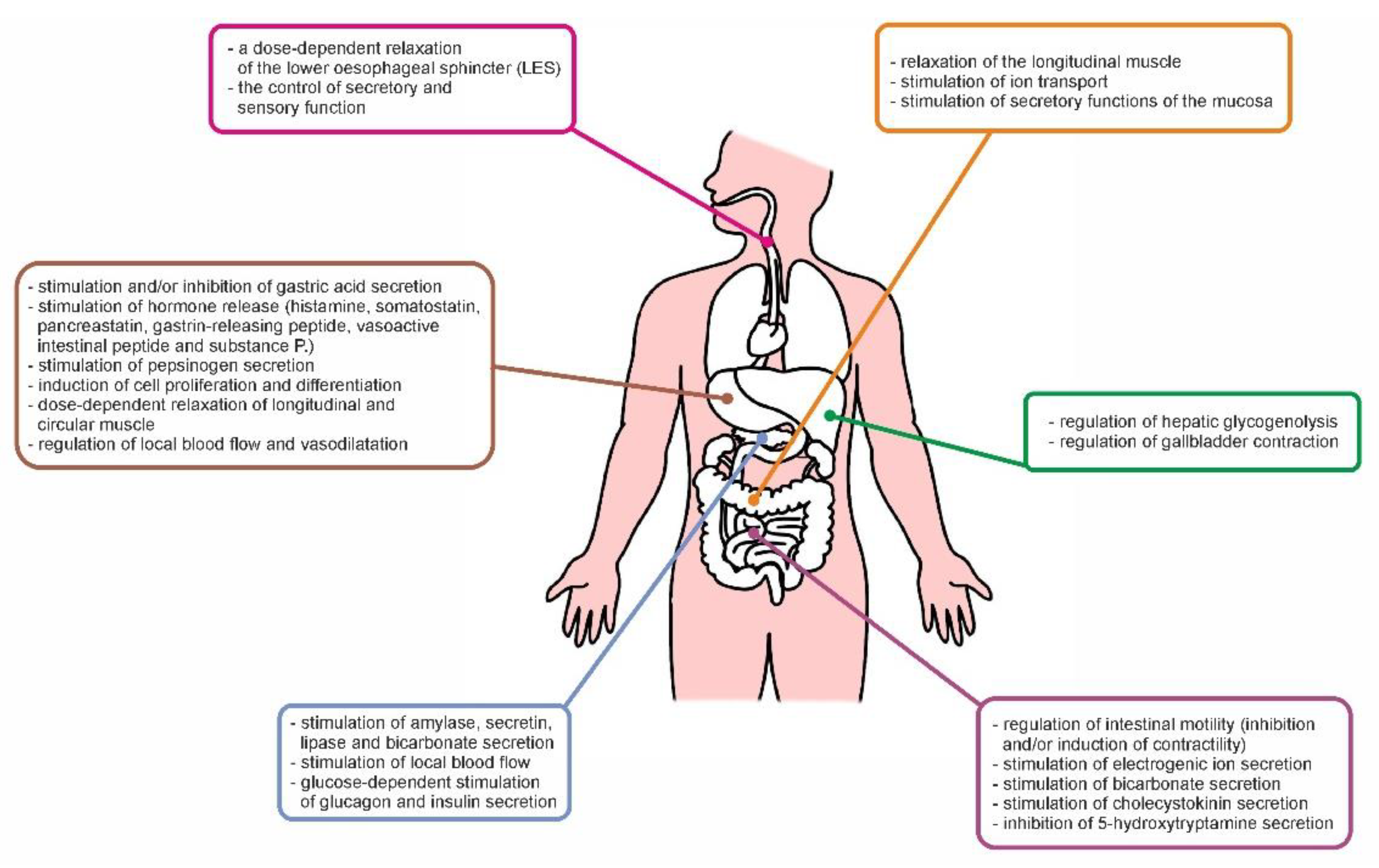
5. Physiological Role of PACAP in the GI Tract
6. PACAP Participation in Pathological Processes in the GI Tract
6.1. Inflammatory Condition
6.2. Injuries and Intoxications
6.3. Neoplastic Processes
6.4. Other Disorders of the GI Tract
7. Therapeutic Use of PACAP and Its Receptors Agonists
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Miyata, A.; Jiang, L.; Dahl, R.D.; Kitada, C.; Kubo, K.; Fujino, M.; Minamino, N.; Arimura, A. Isolation of a neuropeptide corresponding to the N-terminal 27 residues of the pituitary adenylate cyclase activating polypeptide with 38 residues (PACAP38). Biochem. Biophys. Res. Commun. 1990, 170, 643–648. [Google Scholar] [CrossRef]
- Cardoso, J.C.R.; Garcia, M.G.; Power, D.M. Tracing the Origins of the Pituitary Adenylate-Cyclase Activating Polypeptide (PACAP). Front. Neurosci. 2020, 14, 366. [Google Scholar] [CrossRef]
- Arimura, A. Receptors for pituitary adenylate cyclase-activating polypeptide: Comparison with vasoactive intestinal peptide receptors. Trends Endocrinol. Metab. 1992, 3, 288–294. [Google Scholar] [CrossRef]
- Vaudry, D.; Falluel-Morel, A.; Bourgault, S.; Basille, M.; Burel, D.; Wurtz, O.; Fournier, A.; Chow, B.K.; Hashimoto, H.; Galas, L.; et al. Pituitary adenylate cyclase-activating polypeptide and its receptors: 20 years after the discovery. Pharmacol. Rev. 2009, 61, 283–357. [Google Scholar] [CrossRef]
- Vaudry, D.; Gonzalez, B.J.; Basille, M.; Yon, L.; Fournier, A.; Vaudry, H. Pituitary adenylate cyclase-activating polypeptide and its receptors: From structure to functions. Pharmacol. Rev. 2000, 52, 269–324. [Google Scholar] [PubMed]
- Horvath, G.; Opper, B.; Reglodi, D. The Neuropeptide Pituitary Adenylate Cyclase-Activating Polypeptide (PACAP) is Protective in Inflammation and Oxidative Stress-Induced Damage in the Kidney. Int. J. Mol. Sci. 2019, 20, 4944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hannibal, J.; Ekblad, E.; Mulder, H.; Sundler, F.; Fahrenkrug, J. Pituitary adenylate cyclase activating polypeptide (PACAP) in the gastrointestinal tract of the rat: Distribution and effects of capsaicin or denervation. Cell Tissue Res. 1998, 291, 65–79. [Google Scholar] [CrossRef] [PubMed]
- Reglodi, D.; Illes, A.; Opper, B.; Schafer, E.; Tamas, A.; Horvath, G. Presence and Effects of Pituitary Adenylate Cyclase Activating Polypeptide Under Physiological and Pathological Conditions in the Stomach. Front. Endocrinol. (Lausanne) 2018, 9, 90. [Google Scholar] [CrossRef] [Green Version]
- Läuff, J.M.; Modlin, I.M.; Tang, L.H. Biological relevance of pituitary adenylate cyclase-activating polypeptide (PACAP) in the gastrointestinal tract. Regul. Pept. 1999, 84, 1–12. [Google Scholar] [CrossRef]
- Gonkowski, S.; Całka, J. Changes in pituitary adenylate cyclase-activating Peptide 27-like immunoreactive nervous structures in the porcine descending colon during selected pathological processes. J. Mol. Neurosci. 2012, 48, 777–787. [Google Scholar] [CrossRef]
- Palus, K.; Bulc, M.; Całka, J.; Zielonka, Ł.; Nowicki, M. Diabetes Affects the Pituitary Adenylate Cyclase-Activating Polypeptide (PACAP)-Like Immunoreactive Enteric Neurons in the Porcine Digestive Tract. Int. J. Mol. Sci. 2021, 22, 5727. [Google Scholar] [CrossRef]
- Gonkowski, S.; Obremski, K.; Całka, J. The Influence of Low Doses of Zearalenone on Distribution of Selected Active Substances in Nerve Fibers Within the Circular Muscle Layer of Porcine Ileum. J. Mol. Neurosci. 2015, 56, 878–886. [Google Scholar] [CrossRef] [Green Version]
- Thoene, M.; Rytel, L.; Dzika, E.; Wojtkiewicz, J. Increased PACAP- and DβH-Positive Hepatic Nerve Fibers after Bisphenol A Exposure. Toxics 2021, 9, 110. [Google Scholar] [CrossRef]
- Godlewski, J.; Łakomy, I.M. Changes in vasoactive intestinal peptide, pituitary adenylate cyclase-activating polypeptide and neuropeptide Y-ergic structures of the enteric nervous system in the carcinoma of the human large intestine. Folia Histochem. Cytobiol. 2010, 48, 208–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szanto, Z.; Sarszegi, Z.; Reglodi, D.; Nemeth, J.; Szabadfi, K.; Kiss, P.; Varga, A.; Banki, E.; Csanaky, K.; Gaszner, B.; et al. PACAP immunoreactivity in human malignant tumor samples and cardiac diseases. J. Mol. Neurosci. 2012, 48, 667–673. [Google Scholar] [CrossRef] [PubMed]
- Nagahama, M.; Tsuzuki, M.; Mochizuki, T.; Iguchi, K.; Kuwahara, A. Light and electron microscopic studies of pituitary adenylate cyclase-activating peptide (PACAP)--immunoreactive neurons in the enteric nervous system of rat small and large intestine. Anat. Embryol. 1998, 198, 341–352. [Google Scholar] [CrossRef] [PubMed]
- Zeng, N.; Kang, T.; Lyu, R.M.; Wong, H.; Wen, Y.; Walsh, J.H.; Sachs, G.; Pisegna, J.R. The pituitary adenylate cyclase activating polypeptide type 1 receptor (PAC1-R) is expressed on gastric ECL cells: Evidence by immunocytochemistry and RT-PCR. Ann. N. Y. Acad. Sci. 1998, 865, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Kántor, O.; Heinzlmann, A.; Suzuki, N.; Vincze, E.; Kocsis, K.; Köves, K. Distribution of PACAP and its mRNA in several nonneural tissues of rats demonstrated by sandwich enzyme immunoassay and RT-PCR technique. Regul. Pept. 2002, 109, 103–105. [Google Scholar] [CrossRef]
- Sundler, F.; Ekblad, E.; Absood, A.; Håkanson, R.; Köves, K.; Arimura, A. Pituitary adenylate cyclase activating peptide: A novel vasoactive intestinal peptide-like neuropeptide in the gut. Neuroscience 1992, 46, 439–454. [Google Scholar] [CrossRef]
- Toyoshima, M.; Takeuchi, T.; Goto, H.; Mukai, K.; Shintani, N.; Hashimoto, H.; Baba, A.; Hata, F. Roles of PACAP and PHI as inhibitory neurotransmitters in the circular muscle of mouse antrum. Pflüg. Arch. 2006, 451, 559–568. [Google Scholar] [CrossRef]
- Portbury, A.L.; McConalogue, K.; Furness, J.B.; Young, H.M. Distribution of pituitary adenylyl cyclase-activating peptide (PACAP) immunoreactivity in neurons of the guinea-pig digestive tract and their projections in the ileum and colon. Cell Tissue Res. 1995, 279, 385–392. [Google Scholar] [CrossRef]
- Suzuki, M.; Nokihara, K.; Ando, E.; Naruse, S. Immunohistochemical comparison of localization of pituitary adenylate cyclase activating polypeptide (PACAP) and vasoactive intestinal polypeptide (VIP) in the enteric nerve plexus of the guinea pig jejunum. Biomed. Pept. Proteins Nucleic Acids 1996, 2, 19–22. [Google Scholar]
- Wu, M.; Majewski, M.; Wojtkiewicz, J.; Vanderwinden, J.M.; Adriaensen, D.; Timmermans, J.P. Anatomical and neurochemical features of the extrinsic and intrinsic innervation of the striated muscle in the porcine esophagus: Evidence for regional and species differences. Cell Tissue Res. 2003, 311, 289–297. [Google Scholar] [CrossRef]
- Calka, J. Increased expression of CART, nNOS, VIP, PACAP, SP and GAL in enteric neurons of the porcine stomach prepyloric region following hydrochloric acid infusion. Folia Histochem. Cytobiol. 2019, 57, 179–187. [Google Scholar] [CrossRef]
- Czajkowska, M.; Całka, J. Neurochemistry of Enteric Neurons Following Prolonged Indomethacin Administration in the Porcine Duodenum. Front. Pharmacol. 2020, 11, 564457. [Google Scholar] [CrossRef] [PubMed]
- Rytel, L.; Wojtkiewicz, J.; Snarska, A.; Mikołajczyk, A. Changes in the Neurochemical Characterization of Enteric Neurons in the Porcine Duodenum after Administration of Low-Dose Salmonella Enteritidis Lipopolysaccharides. J. Mol. Neurosci. 2020, 71, 1556–1566. [Google Scholar] [CrossRef]
- Uddman, R.; Luts, A.; Absood, A.; Arimura, A.; Ekelund, M.; Desai, H.; Håkanson, R.; Hambreaus, G.; Sundler, F. PACAP, a VIP-like peptide, in neurons of the esophagus. Regul. Pept. 1991, 36, 415–422. [Google Scholar] [CrossRef]
- Vincze, E.; Kántor, O.; Kiss, A.; Gonda, G.; Gombás, P.; Kiss, J.; Juhász, M.; Arimura, A.; Köves, K. Pituitary adenylate cyclase activating polypeptide (PACAP) is present in human and cat gastric glands. Peptides 1999, 20, 937–941. [Google Scholar] [CrossRef]
- Shen, Z.; Larsson, L.T.; Malmfors, G.; Absood, A.; Håkanson, R.; Sundler, F. A novel neuropeptide, pituitary adenylate cyclase-activating polypeptide (PACAP), in human intestine: Evidence for reduced content in Hirschsprung’s disease. Cell Tissue Res. 1992, 269, 369–374. [Google Scholar] [CrossRef]
- Simpson, J.; Sundler, F.; Humes, D.J.; Jenkins, D.; Scholefield, J.H.; Spiller, R.C. Post inflammatory damage to the enteric nervous system in diverticular disease and its relationship to symptoms. Neurogastroenterol. Motil. 2009, 21, 847-e58. [Google Scholar] [CrossRef]
- Kamińska, B.; Landowski, P.; Gonkowski, S.; Szlagatys-Sidorkiewicz, A.; Majewski, M.; Dobosz, M.; Ismail, H.; Korzon, M. Analysis of enteral nervous system in children with drug resistant ulcerative colitis. Med. Wieku Rozw. 2007, 11, 117–122. [Google Scholar] [PubMed]
- Arciszewski, M.B.; Mozel, S.; Sienkiewicz, W. Pituitary adenylate cyclase-activating peptide-27 (PACAP-27) is co-stored with galanin, substance P and corticotropin releasing factor (CRF) in intrapancreatic ganglia of the sheep. Pol. J. Vet. Sci. 2015, 18, 343–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Köves, K.; Arimura, A.; Vigh, S.; Somogyvári-Vigh, A.; Miller, J. Immunohistochemical localization of PACAP in the ovine digestive system. Peptides 1993, 14, 449–455. [Google Scholar] [CrossRef]
- Gonkowski, S.; Kamińska, B.; Landowski, P.; Całka, J. Immunohistochemical distribution of cocaine- and amphetamine-regulated transcript peptide—Like immunoreactive (CART-LI) nerve fibers and various degree of co-localization with other neuronal factors in the circular muscle layer of human descending colon. Histol. Histopathol. 2013, 28, 851–858. [Google Scholar]
- Harmar, A.J.; Fahrenkrug, J.; Gozes, I.; Laburthe, M.; May, V.; Pisegna, J.R.; Vaudry, D.; Vaudry, H.; Waschek, J.A.; Said, S.I. Pharmacology and functions of receptors for vasoactive intestinal peptide and pituitary adenylate cyclase-activating polypeptide: IUPHAR review 1. Br. J. Pharmacol. 2012, 166, 4–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, C.; Remington, J.M.; May, V.; Li, J. Molecular Basis of Class B GPCR Selectivity for the Neuropeptides PACAP and VIP. Front. Mol. Biosci. 2021, 8, 644644. [Google Scholar] [CrossRef]
- Blechman, J.; Levkowitz, G. Alternative Splicing of the Pituitary Adenylate Cyclase-Activating Polypeptide Receptor PAC1: Mechanisms of Fine Tuning of Brain Activity. Front. Endocrinol. 2013, 4, 55. [Google Scholar] [CrossRef] [Green Version]
- Lakk, M.; Denes, V.; Gabriel, R. Pituitary Adenylate Cyclase-Activating Polypeptide Receptors Signal via Phospholipase C Pathway to Block Apoptosis in Newborn Rat Retina. Neurochem. Res. 2015, 40, 1402–1409. [Google Scholar] [CrossRef] [PubMed]
- D’Amico, A.G.; Maugeri, G.; Musumeci, G.; Reglodi, D.; D’Agata, V. PACAP and NAP: Effect of Two Functionally Related Peptides in Diabetic Retinopathy. J. Mol. Neurosci. 2021, 71, 1525–1535. [Google Scholar] [CrossRef] [PubMed]
- Gábriel, R.; Pöstyéni, E.; Dénes, V. Neuroprotective Potential of Pituitary Adenylate Cyclase Activating Polypeptide in Retinal Degenerations of Metabolic Origin. Front. Neurosci. 2019, 13, 1031. [Google Scholar] [CrossRef] [PubMed]
- Katsoulis, S.; Schmidt, W.E.; Schwarzhoff, R.; Folsch, U.R.; Jin, J.G.; Grider, J.R.; Makhlouf, G.M. Inhibitory transmission in guinea pig stomach mediated by distinct receptors for pituitary adenylate cyclase-activating peptide. J. Pharmacol. Exp. Ther. 1996, 278, 199–204. [Google Scholar]
- Läuffer, J.M.; Modlin, I.M.; Hinoue, T.; Kidd, M.; Zhang, T.; Schmid, S.W.; Tang, L.H. Pituitary adenylate cyclase-activating polypeptide modulates gastric enterochromaffin-like cell proliferation in rats. Gastroenterology 1999, 116, 623–635. [Google Scholar] [CrossRef]
- Naruse, S.; Nakamura, T.; Wei, M.; Ando, E.; Nokihara, K.; Wray, V.; Ozaki, T.; Kitagawa, M.; Hayakawa, T. Effects of PACAP-VIP hybrid peptides on gastric blood flow in conscious dogs. Ann. N. Y. Acad. Sci. 1996, 805, 511–515. [Google Scholar] [CrossRef] [PubMed]
- Katsoulis, S.; Clemens, A.; Schwörer, H.; Creutzfeldt, W.; Schmidt, W.E. PACAP is a stimulator of neurogenic contraction in guinea pig ileum. Am. J. Physiol. 1993, 265 Pt 1, G295–G302. [Google Scholar] [CrossRef]
- Katsoulis, S.; Clemens, A.; Schwörer, H.; Creutzfeldt, W.; Schmidt, W.E. Pituitary adenylate cyclase activating polypeptide (PACAP) is a potent relaxant of the rat ileum. Peptides 1993, 14, 587–592. [Google Scholar] [CrossRef]
- Mao, Y.K.; Wang, Y.F.; Moogk, C.; Fox-Threlkeld, J.E.; Xiao, Q.; McDonald, T.J.; Daniel, E.E. Locations and molecular forms of PACAP and sites and characteristics of PACAP receptors in canine ileum. Am. J. Physiol. 1998, 274, G217–G225. [Google Scholar] [CrossRef] [PubMed]
- Schwörer, H.; Clemens, A.; Katsoulis, S.; Köhler, H.; Creutzfeldt, W.; Schmidt, W.E. Pituitary adenylate cyclase-activating peptide is a potent modulator of human colonic motility. Scand. J. Gastroenterol. 1993, 28, 625–632. [Google Scholar] [CrossRef] [PubMed]
- Kishi, M.; Takeuchi, T.; Suthamnatpong, N.; Ishii, T.; Nishio, H.; Hata, F.; Takewaki, T. VIP- and PACAP-mediated nonadrenergic, noncholinergic inhibition in longitudinal muscle of rat distal colon: Involvement of activation of charybdotoxin- and apamin-sensitive K+ channels. Br. J. Pharmacol. 1996, 119, 623–630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Motomura, Y.; Chijiiwa, Y.; Iwakiri, Y.; Ochiai, T.; Nawata, H. Interactive mechanisms among pituitary adenylate cyclase-activating peptide, vasoactive intestinal peptide, and parathyroid hormone receptors in guinea pig cecal circular smooth muscle cells. Endocrinology 1998, 139, 2869–2878. [Google Scholar] [CrossRef]
- Schwörer, H.; Katsoulis, S.; Creutzfeldt, W.; Schmidt, W.E. Pituitary adenylate cyclase activating peptide, a novel VIP-like gut-brain peptide, relaxes the guinea-pig taenia caeci via apamin-sensitive potassium channels. Naunyn Schmiedebergs Arch. Pharmacol. 1992, 346, 511–514. [Google Scholar] [CrossRef]
- Guijarro, L.G.; Rodriguez-Henche, N.; García-López, E.; Noguerales, F.; Dapena, M.A.; Juarranz, M.G.; Prieto, J.C. Receptors for pituitary adenylate cyclase-activating peptide in human liver. J. Clin. Endocrinol. Metab. 1995, 80, 2451–2457. [Google Scholar] [PubMed]
- Robberecht, P.; Gourlet, P.; Cauvin, A.; Buscail, L.; De Neef, P.; Arimura, A.; Christophe, J. PACAP and VIP receptors in rat liver membranes. Am. J. Physiol. 1991, 260, G97–G102. [Google Scholar] [CrossRef]
- Parkman, H.P.; Pagano, A.P.; Ryan, J.P. Dual effects of PACAP on guinea pig gallbladder muscle via PACAP-preferring and VIP/PACAP-preferring receptors. Am. J. Physiol. 1997, 272, G1433–G1438. [Google Scholar] [CrossRef] [PubMed]
- Buscail, L.; Gourlet, P.; Cauvin, A.; De Neef, P.; Gossen, D.; Arimura, A.; Miyata, A.; Coy, D.H.; Robberecht, P.; Christophe, J. Presence of highly selective receptors for PACAP (pituitary adenylate cyclase activating peptide) in membranes from the rat pancreatic acinar cell line AR 4-2. J. FEBS Lett. 1990, 262, 77–81. [Google Scholar] [CrossRef]
- Schulz, S.; Röcken, C.; Mawrin, C.; Weise, W.; Höllt, V.; Schulz, S. Immunocytochemical identification of VPAC1, VPAC2, and PAC1 receptors in normal and neoplastic human tissues with subtype-specific antibodies. Clin. Cancer Res. 2004, 10, 8235–8242. [Google Scholar] [CrossRef] [Green Version]
- Miampamba, M.; Germano, P.M.; Arli, S.; Wong, H.H.; Scott, D.; Taché, Y.; Pisegna, J.R. Expression of pituitary adenylate cyclase-activating polypeptide and PACAP type 1 receptor in the rat gastric and colonic myenteric neurons. Regul. Pept. 2002, 105, 145–154. [Google Scholar] [CrossRef]
- Waschek, J.A.; Richards, M.L.; Bravo, D.T. Differential expression of VIP/PACAP receptor genes in breast, intestinal, and pancreatic cell lines. Cancer Lett. 1995, 92, 143–149. [Google Scholar] [CrossRef]
- Chakder, S.; Rattan, S. Involvement of pituitary adenylate cyclase-activating peptide in opossum internal anal sphincter relaxation. Am. J. Physiol. 1998, 275, G769–G777. [Google Scholar] [CrossRef] [PubMed]
- Jayawardena, D.; Guzman, G.; Gill, R.K.; Alrefai, W.A.; Onyuksel, H.; Dudeja, P.K. Expression and localization of VPAC1, the major receptor of vasoactive intestinal peptide along the length of the intestine. Am. J. Physiol. Gastrointest. Liver Physiol. 2017, 313, G16–G25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fung, C.; Boesmans, W.; Cirillo, C.; Foong, J.P.P.; Bornstein, J.C.; Vanden Berghe, P. VPAC Receptor Subtypes Tune Purinergic Neuron-to-Glia Communication in the Murine Submucosal Plexus. Front. Cell Neurosci. 2017, 11, 118. [Google Scholar] [CrossRef] [Green Version]
- Bertrand, G.; Puech, R.; Maisonnasse, Y.; Bockaert, J.; Loubatières-Mariani, M.M. Comparative effects of PACAP and VIP on pancreatic endocrine secretions and vascular resistance in rat. Br. J. Pharmacol. 1996, 117, 764–770. [Google Scholar] [CrossRef] [Green Version]
- Inagaki, N.; Yoshida, H.; Mizuta, M.; Mizuno, N.; Fujii, Y.; Gonoi, T.; Miyazaki, J.; Seino, S. Cloning and functional characterization of a third pituitary adenylate cyclase-activating polypeptide receptor subtype expressed in insulin-secreting cells. Proc. Natl. Acad. Sci. USA 1994, 91, 2679–2683. [Google Scholar] [CrossRef] [Green Version]
- Murthy, K.S.; Jin, J.G.; Grider, J.R.; Makhlouf, G.M. Characterization of PACAP receptors and signaling pathways in rabbit gastric muscle cells. Am. J. Physiol. 1997, 272, G1391–G1399. [Google Scholar] [CrossRef]
- Teng, B.; Murthy, K.S.; Kuemmerle, J.F.; Grider, J.R.; Makhlouf, G.M. Selective expression of vasoactive intestinal peptide (VIP)2/pituitary adenylate cyclase-activating polypeptide (PACAP)3 receptors in rabbit and guinea pig gastric and tenia coli smooth muscle cells. Regul. Pept. 1998, 77, 127–134. [Google Scholar] [CrossRef]
- Robberecht, P.; De Neef, P.; Lefebvre, R.A. Influence of selective VIP receptor agonists in the rat gastric fundus. Eur. J. Pharmacol. 1998, 359, 77–80. [Google Scholar] [CrossRef]
- Ny, L.; Larsson, B.; Alm, P.; Ekström, P.; Fahrenkrug, J.; Hannibal, J.; Andersson, K.E. Distribution and effects of pituitary adenylate cyclase activating peptide in cat and human lower oesophageal sphincter. Br. J. Pharmacol. 1995, 116, 2873–2880. [Google Scholar] [CrossRef] [PubMed]
- Felley, C.P.; Qian, J.M.; Mantey, S.; Pradhan, T.; Jensen, R.T. Chief cells possess a receptor with high affinity for PACAP and VIP that stimulates pepsinogen release. Am. J. Physiol. 1992, 263 Pt 1, G901–G907. [Google Scholar] [CrossRef]
- Håkanson, R.; Chen, D.; Lindström, E.; Bernsand, M.; Norlén, P. Control of secretion from rat stomach ECL cells in situ and in primary culture. Scand. J. Clin. Lab. Investig. Suppl. 2001, 234, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Sandvik, A.K.; Cui, G.; Bakke, I.; Munkvold, B.; Waldum, H.L. PACAP stimulates gastric acid secretion in the rat by inducing histamine release. Am. J. Physiol. Gastrointest. Liver Physiol. 2001, 281, G997–G1003. [Google Scholar] [CrossRef] [PubMed]
- Piqueras, L.; Taché, Y.; Martínez, V. Peripheral PACAP inhibits gastric acid secretion through somatostatin release in mice. Br. J. Pharmacol. 2004, 142, 67–78. [Google Scholar] [CrossRef]
- Oh, D.S.; Lieu, S.N.; Yamaguchi, D.J.; Tachiki, K.; Lambrecht, N.; Ohning, G.V.; Sachs, G.; Germano, P.M.; Pisegna, J.R. PACAP regulation of secretion and proliferation of pure populations of gastric ECL cells. J. Mol. Neurosci. 2005, 26, 85–97. [Google Scholar] [CrossRef]
- Läuffer, J.M.; Tang, L.H.; Zhang, T.; Hinoue, T.; Rahbar, S.; Odo, M.; Modlin, I.M.; Kidd, M. PACAP mediates the neural proliferative pathway of Mastomys enterochromaffin-like cell transformation. Regul. Pept. 2001, 102, 157–164. [Google Scholar] [CrossRef]
- Van Assche, G.; Depoortere, I.; de Vos, R.; Geboes, K.; Janssens, J.J.; Collins, S.M.; Peeters, T.L. Region-specific antiproliferative effect of VIP and PACAP-(1-38) on rabbit enteric smooth muscle. Am. J. Physiol. 1999, 276, G303–G310. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, T.; Toyoshima, M.; Mukai, K.; Hagi, K.; Matsui, M.; Nakajima, H.; Azuma, Y.T.; Hata, F. Involvement of M(2) muscarinic receptors in relaxant response of circular muscle of mouse gastric antrum. Neurogastroenterol. Motil. 2006, 18, 226–233. [Google Scholar] [CrossRef] [PubMed]
- Ekblad, E.; Sundler, F. Distinct receptors mediate pituitary adenylate cyclase-activating peptide- and vasoactive intestinal peptide-induced relaxation of rat ileal longitudinal muscle. Eur. J. Pharmacol. 1997, 334, 61–66. [Google Scholar] [CrossRef]
- Onaga, T.; Harada, Y.; Okamoto, K. Pituitary adenylate cyclase-activating polypeptide (PACAP) induces duodenal phasic contractions via the vagal cholinergic nerves in sheep. Regul. Pept. 1998, 77, 69–76. [Google Scholar] [CrossRef]
- Costa, M.; Spencer, N.J.; Brookes, S.J.H. The role of enteric inhibitory neurons in intestinal motility. Auton. Neurosci. 2021, 235, 102854. [Google Scholar]
- Cox, H.M. Pituitary adenylate cyclase activating polypeptides, PACAP-27 and PACAP-38: Stimulators of electrogenic ion secretion in the rat small intestine. Br. J. Pharmacol. 1992, 106, 498–502. [Google Scholar] [CrossRef] [Green Version]
- Fuchs, M.; Adermann, K.; Raab, H.R.; Forssmann, W.G.; Kuhn, M. Pituitary adenylate cyclase-activating polypeptide: A potent activator of human intestinal ion transport. Ann. N. Y. Acad. Sci. 1996, 805, 640–647. [Google Scholar] [CrossRef]
- Takeuchi, K.; Takehara, K.; Kato, S.; Yagi, K. PACAPs stimulate duodenal bicarbonate secretion at PACAP receptors in the rat. Am. J. Physiol. 1997, 272, G646–G653. [Google Scholar] [CrossRef]
- Chang, C.H.; Chey, W.Y.; Braggins, L.; Coy, D.H.; Chang, T.M. Pituitary adenylate cyclase-activating polypeptide stimulates cholecystokinin secretion in STC-1 cells. Am. J. Physiol. 1996, 271, G516–G523. [Google Scholar] [CrossRef] [PubMed]
- Racké, K.; Reimann, A.; Schwörer, H.; Kilbinger, H. Regulation of 5-HT release from enterochromaffin cells. Behav. Brain Res. 1996, 73, 83–87. [Google Scholar] [CrossRef]
- Chang, C.H.; Chey, W.Y.; Erway, B.; Coy, D.H.; Chang, T.M. Modulation of secretin release by neuropeptides in secretin-producing cells. Am. J. Physiol. 1998, 275, G192–G202. [Google Scholar] [CrossRef] [PubMed]
- McConalogue, K.; Furness, J.B.; Vremec, M.A.; Holst, J.J.; Tornøe, K.; Marley, P.D. Histochemical, pharmacological, biochemical and chromatographic evidence that pituitary adenylyl cyclase activating peptide is involved in inhibitory neurotransmission in the taenia of the guinea-pig caecum. J. Auton. Nerv. Syst. 1995, 50, 311–322. [Google Scholar] [CrossRef]
- Yokota, C.; Kawai, K.; Ohashi, S.; Watanabe, Y.; Yamashita, K. PACAP stimulates glucose output from the perfused rat liver. Peptides 1995, 16, 55–60. [Google Scholar] [CrossRef]
- Alonso, R.M.; Rodríguez, A.M.; García, L.J.; López, M.A.; Calvo, J.J. Comparison between the effects of VIP and the novel peptide PACAP on the exocrine pancreatic secretion of the rat. Pancreas 1994, 9, 123–128. [Google Scholar] [CrossRef]
- Schmidt, W.E.; Seebeck, J.; Höcker, M.; Schwarzhoff, R.; Schäfer, H.; Fornefeld, H.; Morys-Wortmann, C.; Fölsch, U.R.; Creutzfeldt, W. PACAP and VIP stimulate enzyme secretion in rat pancreatic acini via interaction with VIP/PACAP-2 receptors: Additive augmentation of CCK/carbachol-induced enzyme release. Pancreas 1993, 8, 476–487. [Google Scholar] [CrossRef] [PubMed]
- Kashimura, J.; Shimosegawa, T.; Iguchi, K.; Mochizuki, T.; Yanaihara, N.; Koizumi, M.; Toyota, T. The stimulatory effects and binding characteristics of PACAP27 in rat dispersed pancreatic acini. Tohoku J. Exp. Med. 1993, 171, 243–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mungan, Z.; Ertan, A.; Hammer, R.A.; Arimura, A. Effect of pituitary adenylate cyclase activating polypeptide on rat pancreatic exocrine secretion. Peptides 1991, 12, 559–562. [Google Scholar] [CrossRef]
- Yada, T.; Sakurada, M.; Ihida, K.; Nakata, M.; Murata, F.; Arimura, A.; Kikuchi, M. Pituitary adenylate cyclase activating polypeptide is an extraordinarily potent intra-pancreatic regulator of insulin secretion from islet beta-cells. J. Biol. Chem. 1994, 269, 1290–1293. [Google Scholar] [CrossRef]
- Yokota, C.; Kawai, K.; Ohashi, S.; Watanabe, Y.; Suzuki, S.; Yamashita, K. Stimulatory effects of pituitary adenylate cyclase-activating polypeptide (PACAP) on insulin and glucagon release from the isolated perfused rat pancreas. Acta Endocrinol. 1993, 129, 473–479. [Google Scholar] [CrossRef] [PubMed]
- Heimesaat, M.M.; Dunay, I.R.; Schulze, S.; Fischer, A.; Grundmann, U.; Alutis, M.; Kühl, A.A.; Tamas, A.; Toth, G.; Dunay, M.P.; et al. Pituitary adenylate cyclase-activating polypeptide ameliorates experimental acute ileitis and extra-intestinal sequelae. PLoS ONE 2014, 9, e108389. [Google Scholar] [CrossRef] [Green Version]
- Bereswill, S.; Escher, U.; Grunau, A.; Kühl, A.A.; Dunay, I.R.; Tamas, A.; Reglodi, D.; Heimesaat, M.M. Pituitary Adenylate Cyclase-Activating Polypeptide-A Neuropeptide as Novel Treatment Option for Subacute Ileitis in Mice Harboring a Human Gut Microbiota. Front. Immunol. 2019, 10, 554. [Google Scholar] [CrossRef]
- Illes, A.; Horvath, G.; Schafer, E.; Kerenyi, M. Effect of PACAP on Bacterial Adherence and Cytokine Expression in Intestinal Cell Cultures. Int. J. Pept. Res. Ther. 2019, 25, 1011–1018. [Google Scholar] [CrossRef] [Green Version]
- Heimesaat, M.M.; Reifenberger, G.; Vicena, V.; Illes, A.; Horvath, G.; Tamas, A.; Fulop, B.D.; Bereswill, S.; Reglodi, D. Intestinal Microbiota Changes in Mice Lacking Pituitary Adenylate Cyclase Activating Polypeptide (PACAP)—Bifidobacteria Make the Difference. Eur. J. Microbiol. Immunol. 2017, 7, 187–199. [Google Scholar] [CrossRef] [Green Version]
- Kun, J.; Szitter, I.; Kemény, A.; Perkecz, A.; Kereskai, L.; Pohóczky, K.; Vincze, A.; Gódi, S.; Szabó, I.; Szolcsányi, J.; et al. Upregulation of the transient receptor potential ankyrin 1 ion channel in the inflamed human and mouse colon and its protective roles. PLoS ONE 2014, 9, e108164. [Google Scholar] [CrossRef]
- Azuma, Y.T.; Hagi, K.; Shintani, N.; Kuwamura, M.; Nakajima, H.; Hashimoto, H.; Baba, A.; Takeuchi, T. PACAP provides colonic protection against dextran sodium sulfate induced colitis. J. Cell. Physiol. 2008, 216, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Nemetz, N.; Abad, C.; Lawson, G.; Nobuta, H.; Chhith, S.; Duong, L.; Tse, G.; Braun, J.; Waschek, J.A. Induction of colitis and rapid development of colorectal tumors in mice deficient in the neuropeptide PACAP. Int. J. Cancer 2008, 122, 1803–1809. [Google Scholar] [CrossRef] [Green Version]
- Heimesaat, M.M.; Mousavi, S.; Kløve, S.; Genger, C.; Weschka, D.; Tamas, A.; Reglodi, D.; Bereswill, S. Pituitary Adenylate Cyclase-Activating Polypeptide Alleviates Intestinal, Extra-Intestinal and Systemic Inflammatory Responses during Acute Campylobacter jejuni-induced Enterocolitis in Mice. Pathogens 2020, 9, 805. [Google Scholar] [CrossRef]
- Horvath, G.; Illes, A.; Heimesaat, M.M.; Bardosi, A.; BHardosi, S.; Tamas, A.; Fulop, B.D.; Opper, B.; Nemeth, J.; Ferencz, A.; et al. Protective Intestinal Effects of Pituitary Adenylate Cyclase Activating Polypeptide. In Pituitary Adenylate Cyclase Activating Polypeptide—PACAP, Current Topics in Neurotoxicity 11; Reglodi, D., Tamas, A., Eds.; Springer: New York, NY, USA, 2016; pp. 271–288. [Google Scholar]
- Nedvig, K.; Szabó, G.; Csukás, D.; Sándor, J.; Németh, J.; Kovács, K.; Reglődi, D.; Kemény, A.; Wéber, G.; Ferencz, A. [Examination of cytoprotective and anti-inflammatory effect of PACAP-38 on small bowel autotransplantation]. Magy. Seb. 2013, 66, 250–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferencz, A.; Rácz, B.; Tamás, A.; Nedvig, K.; Németh, J.; Kalmár-Nagy, K.; Horváth, O.P.; Wéber, G.; Röth, E.; Reglödi, D. Changes and effect of PACAP-38 on intestinal ischemia-reperfusion and autotransplantation. Transplant. Proc. 2009, 41, 57–59. [Google Scholar] [CrossRef]
- Ferencz, A.; Nedvig, K.; Fekecs, T.; Rácz, B.; Wéber, G.; Hashimoto, H.; Baba, A.; Helyes, Z.; Reglödi, D. Comparison of intestinal cold preservation injury on pituitary adenylate cyclase-activating polypeptide in knockout and wild-type mice. Transplant. Proc. 2010, 42, 2290–2292. [Google Scholar] [CrossRef]
- Illes, A.; Opper, B.; Reglodi, D.; Kerenyi, M.; Czetany, P.; Boronkai, A.; Schafer, E.; Toth, G.; Fabian, E.; Horvath, G. Effects of pituitary adenylate cyclase activating polypeptide on small intestinal INT 407 cells. Neuropeptides 2017, 65, 106–113. [Google Scholar] [CrossRef]
- Sadanandan, N.; Cozene, B.; Park, Y.J.; Farooq, J.; Kingsbury, C.; Wang, Z.J.; Moscatello, A.; Saft, M.; Cho, J.; Gonzales-Portillo, B.; et al. Pituitary Adenylate Cyclase-Activating Polypeptide: A Potent Therapeutic Agent in Oxidative Stress. Antioxidants 2021, 10, 354. [Google Scholar] [CrossRef]
- Ekelund, M.; Ekblad, E. Intestinal adaptation in atrophic rat ileum is accompanied by supersensitivity to vasoactive intestinal peptide, pituitary adenylate cyclase-activating peptide and nitric oxide. Scand. J. Gastroenterol. 2001, 36, 251–257. [Google Scholar] [CrossRef]
- Mei, Q.; Sundler, F. Changes in pituitary adenylate cyclase activating peptide and vasoactive intestinal peptide innervation of rat oxyntic mucosa during ulcer healing. Neuropeptides 1998, 32, 527–535. [Google Scholar] [CrossRef]
- Czajkowska, M.; Rychlik, A.; Całka, J. Long-term treatment with naproxen changes the chemical coding of the porcine intramural duodenum neurons. Ann. Anat. 2020, 227, 151425. [Google Scholar] [CrossRef] [PubMed]
- Le, S.V.; Yamaguchi, D.J.; McArdle, C.A.; Tachiki, K.; Pisegna, J.R.; Germano, P. PAC1 and PACAP expression, signaling, and effect on the growth of HCT8, human colonic tumor cells. Regul. Pept. 2002, 109, 115–125. [Google Scholar] [CrossRef]
- Ramos-Álvarez, I.; Mantey, S.A.; Nakamura, T.; Nuche-Berenguer, B.; Moreno, P.; Moody, T.W.; Maderdrut, J.L.; Coy, D.H.; Jensen, R.T. A structure-function study of PACAP using conformationally restricted analogs: Identification of PAC1 receptor-selective PACAP agonists. Peptides 2015, 66, 26–42. [Google Scholar] [CrossRef] [Green Version]
- Bourgault, S.; Vaudry, D.; Botia, B.; Couvineau, A.; Laburthe, M.; Vaudry, H.; Fournier, A. Novel stable PACAP analogs with potent activity towards the PAC1 receptor. Peptides 2008, 29, 919–932. [Google Scholar] [CrossRef]
- Inooka, H.; Endo, S.; Kitada, C.; Mizuta, E.; Fujino, M. Pituitary adenylate cyclase-activating polypeptide (PACAP) with 27 residues. Conformation determined by 1:H NMR and CD spectroscopies and distance geometry in 25% methanol solution. Int J. Pept. Protein Res. 1992, 40, 456–464. [Google Scholar] [CrossRef] [PubMed]
- Wray, V.; Kokoschke, C.; Nokihara, K.; Naruse, S. Solution structure of pituitary adenylate cyclase-activating polypeptide by nuclear magnetic resonance spectroscopy. Biochemistry 1993, 32, 5832–5841. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Species | Localization | References |
|---|---|---|
| Oesophagus | ||
| Rat | nerve fibers in the circular and longitudinal muscle layers, myenteric and submucous plexuses and in the mucosa | [7] |
| Pig | nerve terminals in the striated muscle | [23] |
| Human | neurons in the myenteric ganglia intraganglionic nerve fibers in the myenteric ganglia nerve fibers in the circular and longitudinal muscle layers | [27] [27] [28] |
| Cat | nerve fibers in the myenteric ganglia and muscle layers | [19] |
| Sheep | nerve fibers in the myenteric ganglia and muscle layers | [19,33] |
| Ferret | nerve fibers in the myenteric ganglia and muscle layers | [19] |
| Stomach | ||
| Rat | nerve fibers in the circular and longitudinal muscle layers, myenteric and submucous plexuses and in the mucosa neurons in the myenteric ganglia the enterochromaffin-like (ECL) cells in the mucosa | [7,18] [7,18] [17] |
| Mouse | nerve fibers in the mucosa nerve fibers in the circular muscle layer in the antrum | [19] [20] |
| Pig | neurons in the myenteric and submucous plexuses in the corpus and prepyloric area | [11,24] |
| Human | nerve fibers in the mucosa and muscle layers cells in the gastric glands | [19] [28] |
| Cat | nerve fibers in the myenteric ganglia, muscle layers and mucosa cells in the gastric glands | [19,28] |
| Sheep | nerve fibers in the myenteric ganglia and muscle layers | [19,33] |
| Ferret | nerve fibers in the myenteric ganglia and muscle layers | [19] |
| small intestine | ||
| Rat | neurons in the myenteric and submucous plexuses nerve fibers in the circular and longitudinal muscle layers, myenteric and submucous plexuses and in the mucosa | [7,9,16] |
| Mouse | nerve fibers in the myenteric ganglia and smooth muscle | [19] |
| Guinea pig | nerve fibers in the circular and longitudinal muscle layers, myenteric and submucous plexuses and around blood vessels of the submucosa neurons in the myenteric ganglia | [21,22] |
| Pig | neurons in the myenteric plexus, outer submucous plexus, and inner submucous plexus nerve fibers in the mucosa and both muscular layers in the ileum | [10,11,12,25,26] |
| human | neurons in the myenteric and submucous ganglia nerve fibers in the circular and longitudinal muscle layers, in the mucosa and in the enteric ganglia | [19] |
| Cat | nerve fibers in the myenteric ganglia and muscle layers | [19] |
| Sheep | nerve fibers in the myenteric ganglia and muscle layers and arterial walls | [19,33] |
| Ferret | nerve fibers in the myenteric ganglia and muscle layers | [19] |
| large intestine | ||
| Rat | neurons in the myenteric and submucous plexuses nerve fibers in the circular and longitudinal muscle layers, myenteric and submucous plexuses and in the mucosa | [7,9,16] |
| Guinea pig | nerve fibers in the circular and longitudinal muscle layers, myenteric and submucous plexuses and around blood vessels of the submucosa neurons in the myenteric ganglia | [21,22] |
| Pig | neurons in the myenteric plexus, outer submucous plexus, and inner submucous plexus in the descending colon nerve fibers in the circular and longitudinal muscle layers, in the mucosa and in the enteric plexuses | [10,11] |
| Human | neurons in the myenteric plexus and submucous plexus nerve fibers in both enteric plexuses, mucosa and muscular layers | [14,29,30] [31] |
| Cat | nerve fibers in the myenteric ganglia and muscle layers | [19] |
| Sheep | nerve fibers in the myenteric ganglia and muscle layers | [19,33] |
| Ferret | nerve fibers in the myenteric ganglia and muscle layers | [19] |
| Pancreas | ||
| Rat | endocrine parts of pancreas | [18] |
| Sheep | nerve fibers in the islet of Langerhans and small arteries neurons in the intrapancreatic ganglia | [32,33] |
| Organ | Species | Disease Model | References |
|---|---|---|---|
| Stomach | Rat | extrinsic denervation experimental ulcers | [7] [107] |
| Pig | experimentally-induces hyperacidity diabetes | [24] [11] | |
| Small intestine | Rat | intestinal autotranslantation ischemia-reperfusion | [101] [102] |
| Pig | diabetes | [11] | |
| Mouse | cold and warm preservation in the course of intestinal transplantation | [103] | |
| Intestinal INT407 cells | Human | lipopolysaccharide (LPS) exposure and bacterial adherence (Escherichia coli, Salmonella Typhimurium, Klebsiella pneumoniae, Enterococcus faecalis) oxidative stress | [94] [104] |
| Duodenum | Pig | naproxen and indomethacin administration | [25,108] |
| Ileum | Mouse | acute ileitis subacute ileitis | [92] [93] |
| Rat | a dysfunctional (atrophic) intestine | [106] | |
| Pig | zearalenone intoxication | [12] | |
| Large intestine | Human | inflammatory bowel disease carcinoma Hirschsprung’s disease | [100] [14] [29] |
| Colon | Mouse | dextran sulphate sodium (DSS)-induced colitis acute Campylobacter jejuni-induced enterocolitis | [96,97,98] [99] |
| Human | symptomatic diverticular disease drug-resistant ulcerative colitis malignant tumor | [30] [31] [15] | |
| Pig | chemically-induced inflammation proliferative enteropathy axotomy of caudal colonic nerves diabetes | [10] [10] [10] [11] | |
| Colonic Caco-2 cells | Human | lipopolysaccharide (LPS) exposure and bacterial adherence (Escherichia coli, Salmonella Typhimurium, Klebsiella pneumoniae, Enterococcus faecalis) | [94] |
| HCT8 human colonic tumor cell | Pig | tumor | [109] |
| Liver | Pig | bisphenol A dietary exposure | [13] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karpiesiuk, A.; Palus, K. Pituitary Adenylate Cyclase-Activating Polypeptide (PACAP) in Physiological and Pathological Processes within the Gastrointestinal Tract: A Review. Int. J. Mol. Sci. 2021, 22, 8682. https://doi.org/10.3390/ijms22168682
Karpiesiuk A, Palus K. Pituitary Adenylate Cyclase-Activating Polypeptide (PACAP) in Physiological and Pathological Processes within the Gastrointestinal Tract: A Review. International Journal of Molecular Sciences. 2021; 22(16):8682. https://doi.org/10.3390/ijms22168682
Chicago/Turabian StyleKarpiesiuk, Aleksandra, and Katarzyna Palus. 2021. "Pituitary Adenylate Cyclase-Activating Polypeptide (PACAP) in Physiological and Pathological Processes within the Gastrointestinal Tract: A Review" International Journal of Molecular Sciences 22, no. 16: 8682. https://doi.org/10.3390/ijms22168682