Novel Mechanism and Kinetics of Tetramethrin Degradation Using an Indigenous Gordonia cholesterolivorans A16

 ,
,  and
and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Isolation, Enrichment, and Characterization of the Bacterial Strain A16
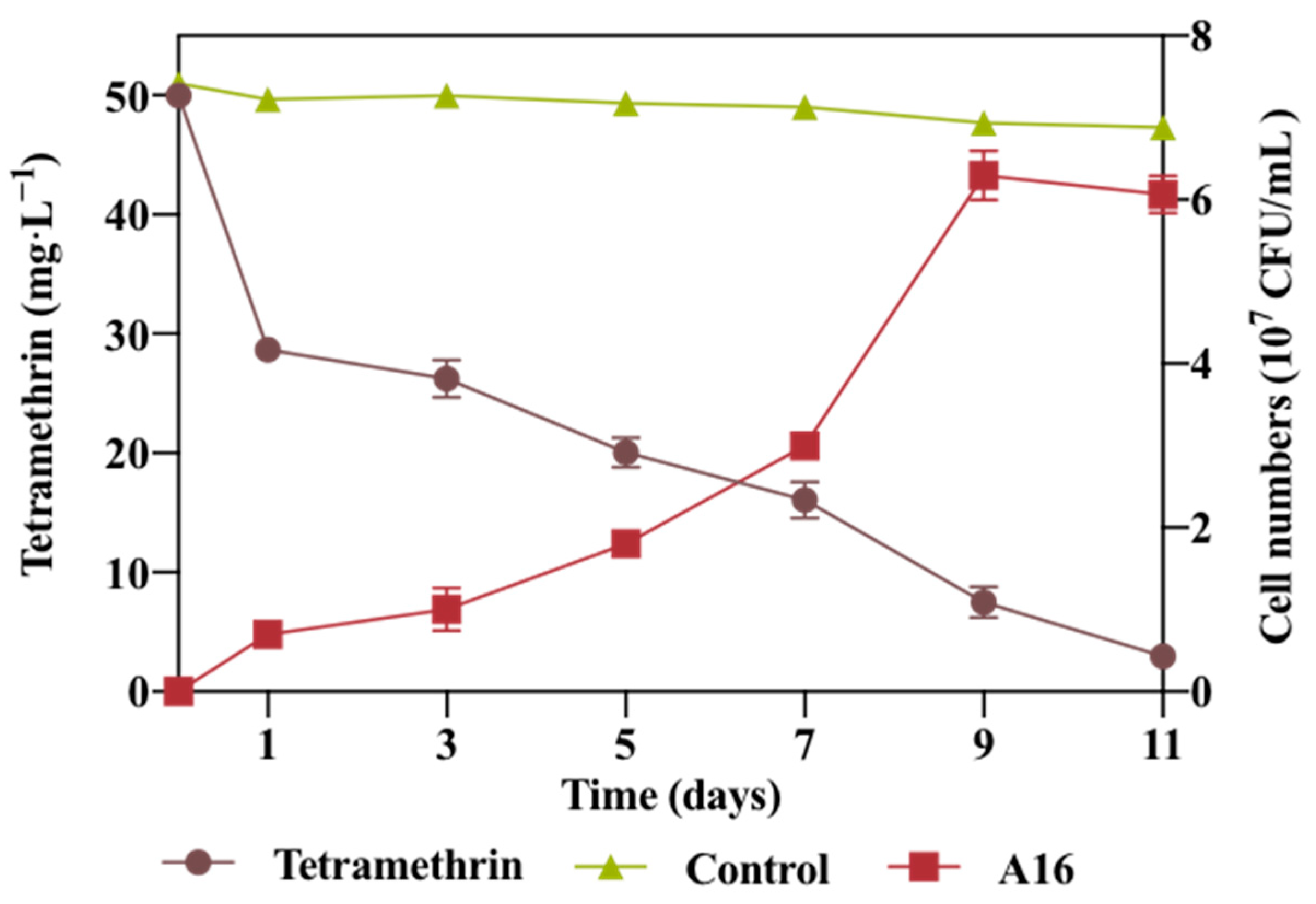
2.2. Growth Linked Degradation of Tetramethrin with Strain A16
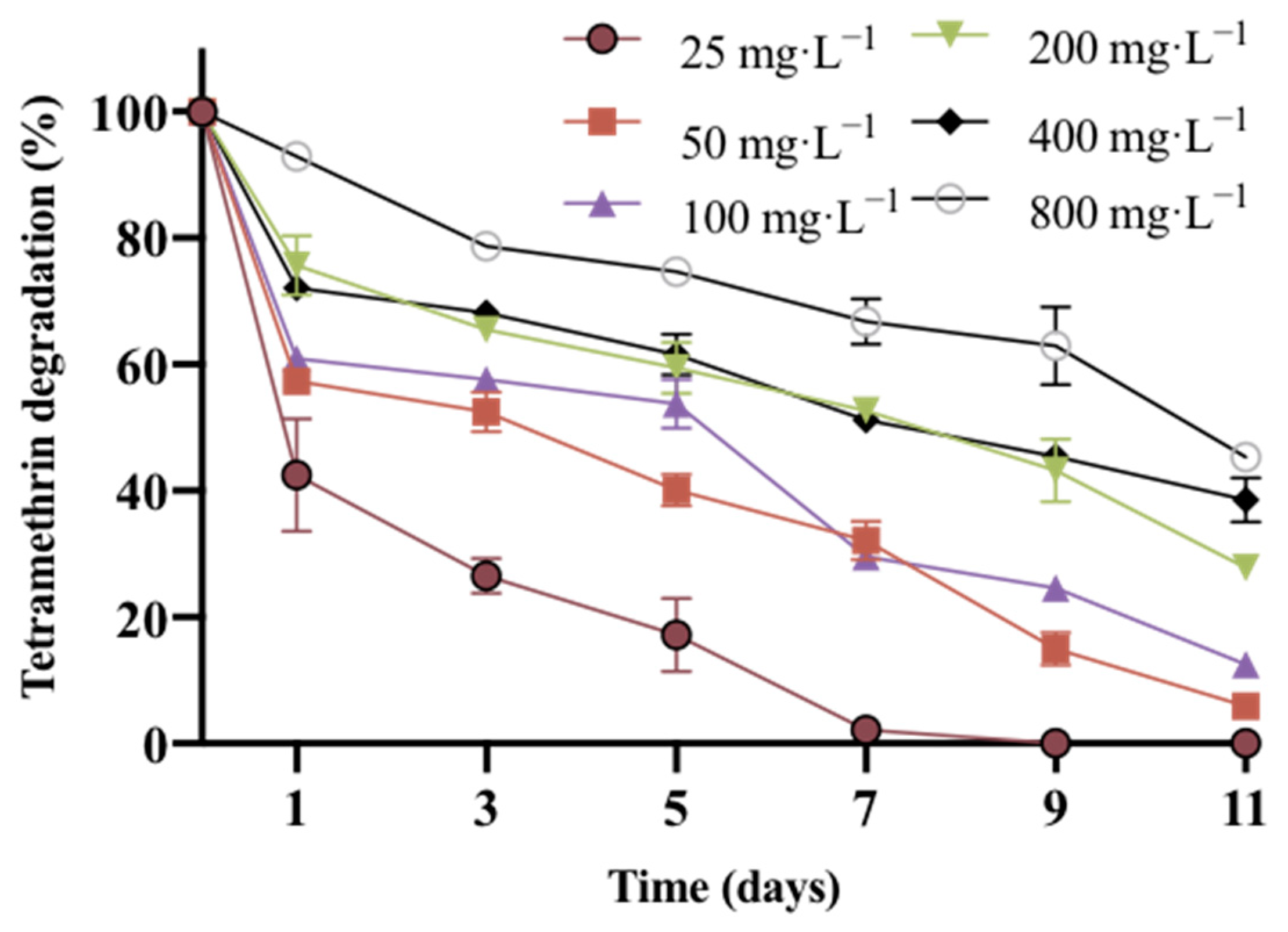
2.3. Degradation Kinetics of Strain A16 with Different Initial Concentrations of Tetramethrin
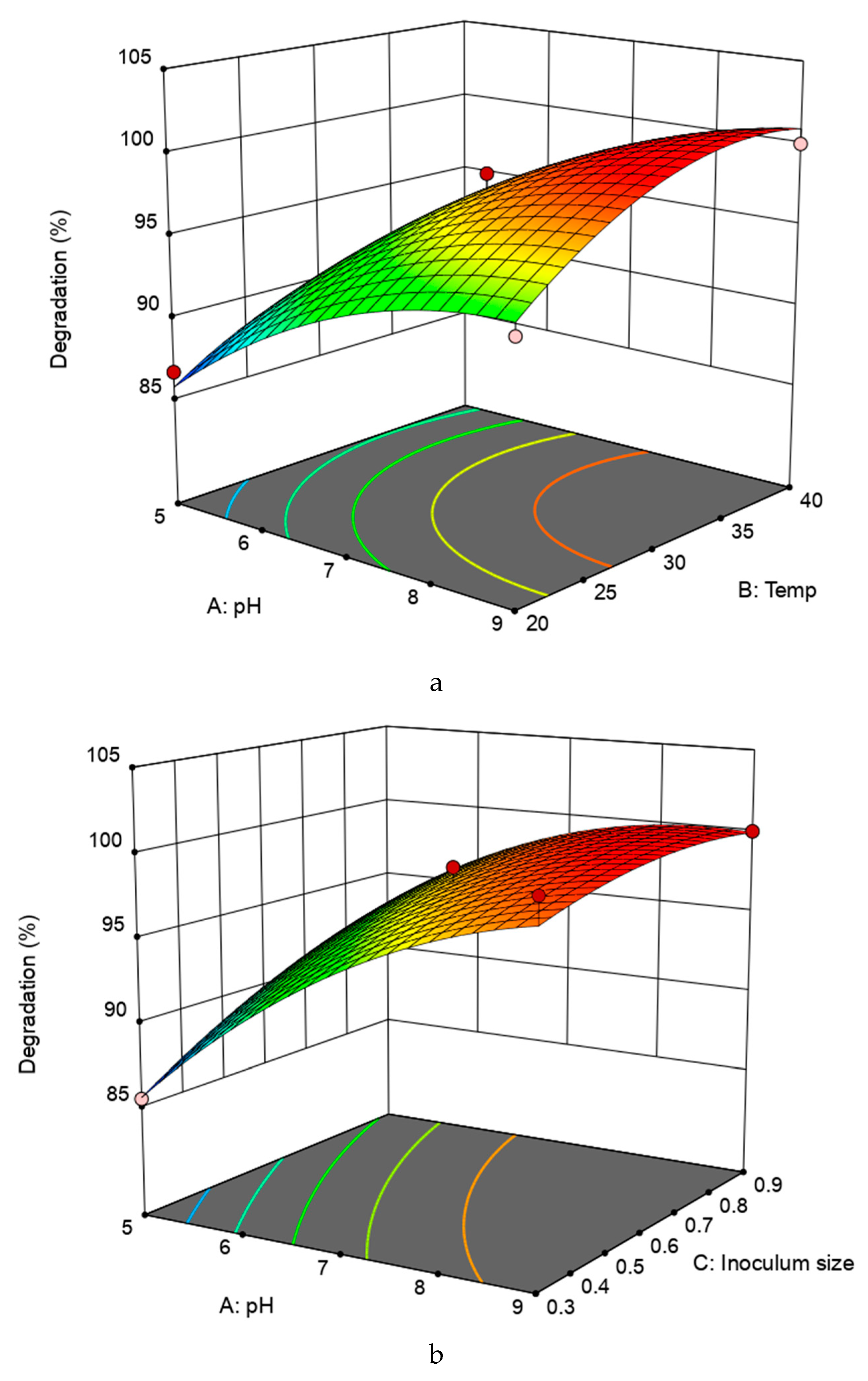
2.4. Optimization of Tetramethrin Degradation Conditions
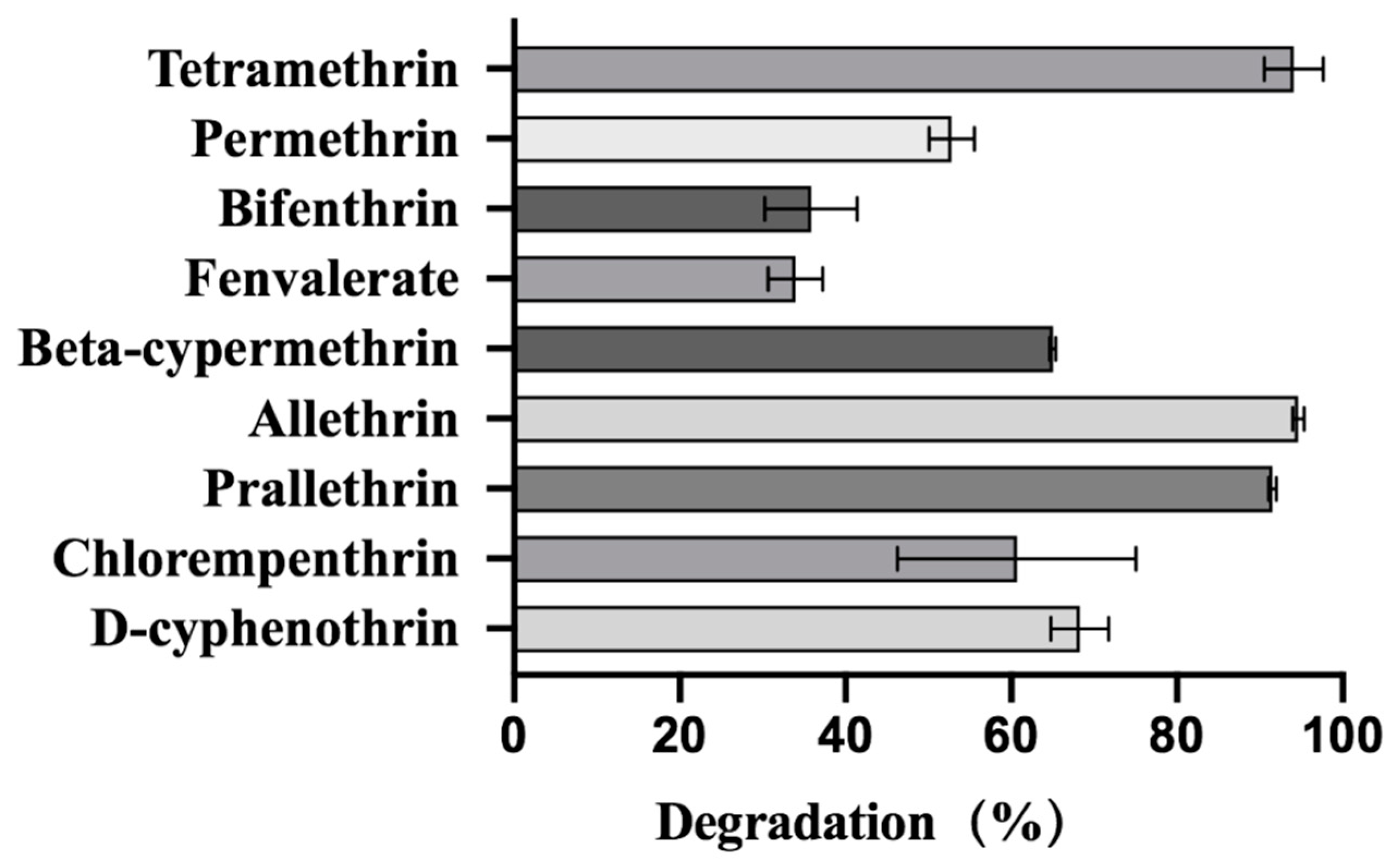
2.5. Degradation of Various Synthetic Pyrethroids by Strain A16
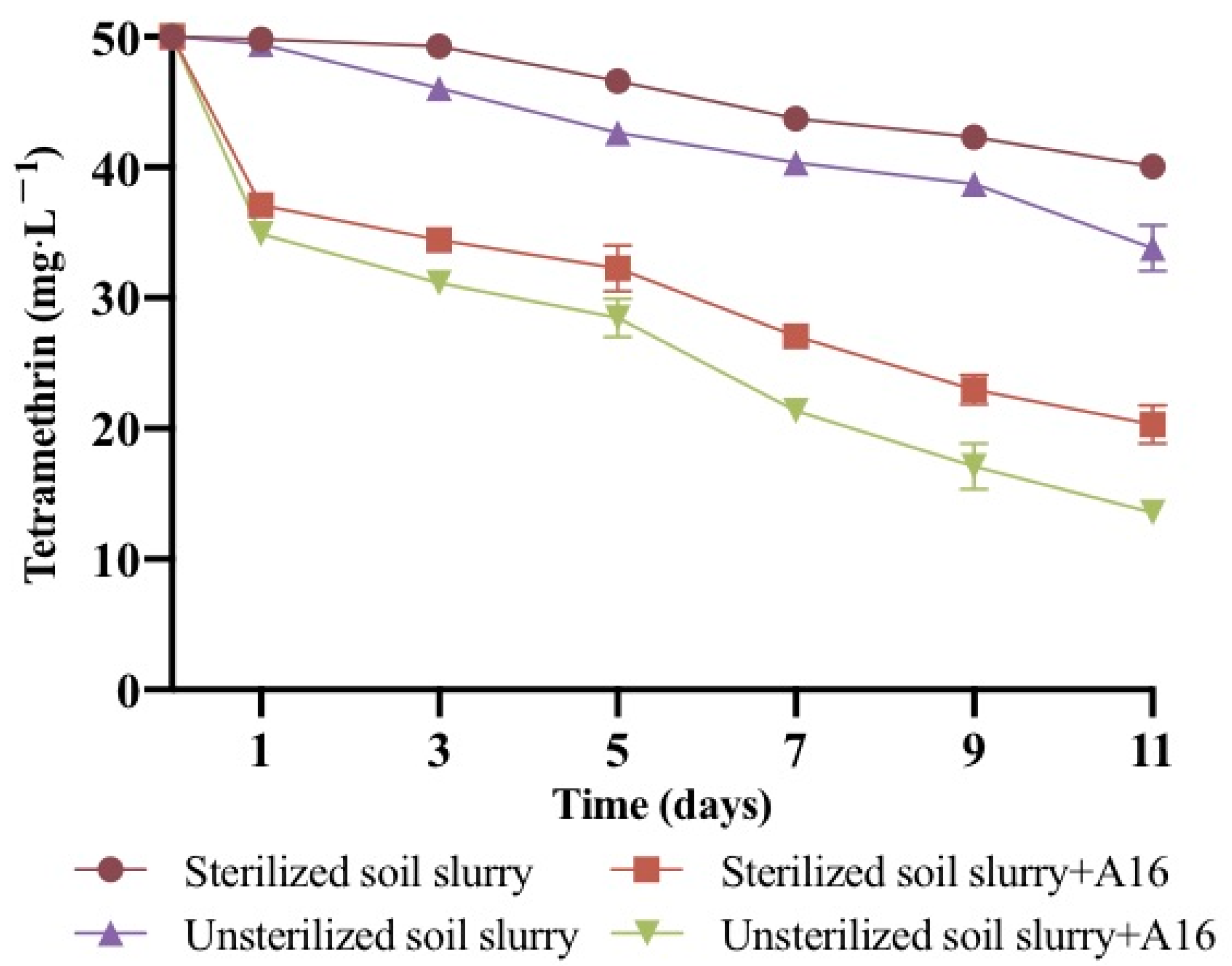
2.6. Degradation Kinetics of Tetramethrin in Soil Slurry
2.7. Metabolite Identification
3. Material and Methods
3.1. Chemicals and Media
3.2. Enrichment, Isolation and Screening of Tetramethrin-Degrading Bacterial Strain A16
3.3. Bacterial Identification
3.4. Growth and Degradation Assays
3.5. Different Initial Concentrations of Tetramethrin Degradation
3.6. Optimization of Tetramethrin Degradation Conditions
3.7. Degradation of Various Substrates by A16
3.8. Biodegradation of Tetramethrin in Soil Slurry
3.9. Identification of Tetramethrin Metabolites
3.10. Kinetic Analysis
3.11. Chemical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Lybrand, D.B.; Xu, H.; Last, R.L.; Pichersky, E. How Plants synthesize pyrethrins: Safe and biodegradable insecticides. Trends. Plant. Sci. 2020, 25, 1240–1251. [Google Scholar] [CrossRef]
- Chrustek, A.; Hołyńska-Iwan, I.; Dziembowska, I.; Bogusiewicz, J.; Wróblewski, M.; Cwynar, A. Current research on the safety of pyrethroids used as insecticides. Medicina 2018, 54, 61. [Google Scholar] [CrossRef] [Green Version]
- Bhatt, P.; Huang, Y.; Rene, E.R.; Kumar, A.J.; Chen, S. Mechanism of allethrin biodegradation by a newly isolated Sphingomonas trueperi strain CW3 from wastewater sludge. Bioresour. Technol. 2020, 305, 123074. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, Y.; Hou, Z.; Wang, X.; Wang, J.; Lu, Z. Biodegradation potential of deltamethrin by the Bacillus cereus strain Y1 in both culture and contaminated soil. Int. Biodeterior. Biodegrad. 2016, 106, 53–59. [Google Scholar] [CrossRef]
- Nasuti, C. Different effects of type I and type II pyrethroids on erythrocyte plasma membrane properties and enzymatic activity in rats. Toxicology 2003, 191, 233–244. [Google Scholar] [CrossRef]
- Zhai, Y.; Li, K.; Song, J.; Shi, Y.; Yan, Y. Molecular cloning, purification and biochemical characterization of a novel pyrethroid-hydrolyzing carboxylesterase gene from Ochrobactrum anthropi YZ-1. J. Hazard. Mater. 2012, 221–222, 206–212. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, T. Simultaneous determination of 18 pyrethroids in indoor air by gas chromatography/mass spectrometry. J. Chromatogr. A 2009, 1216, 5069–5076. [Google Scholar] [CrossRef] [PubMed]
- Diegelmann, C.; Weber, J.; Heinzel-Wieland, R.; Kemme, M. Characterization of a cypermethrin-degrading Methylobacterium sp. strain A-1 and molecular cloning of its carboxylesterase gene: Cypermethrin biodegradation. J. Basic Microbiol. 2015, 55, 1245–1254. [Google Scholar] [CrossRef]
- Liu, H.; Hussain, S.A.; Ali, D.; Omar, S.Y.A.; Shaik, U.; Alghamdi, H.A.H. Induced alteration of rat erythrocyte membrane with effect of pyrethroid based compounds. Saudi J. Biol. Sci. 2020, 27, 3669–3675. [Google Scholar] [CrossRef] [PubMed]
- Köprücü, K.; Aydın, R. The toxic effects of pyrethroid deltamethrin on the common carp (Cyprinus carpio L.) embryos and larvae. Pestic. Biochem. Phys. 2004, 80, 47–53. [Google Scholar] [CrossRef]
- Hellinghausen, G.; Readel, E.R.; Wahab, M.F.; Lee, J.T.; Lopez, D.A.; Weatherly, C.A. Mass spectrometry-sompatible enantiomeric separations of 100 pesticides using core–shell chiral stationary phases and evaluation of iterative curve fitting models for overlapping peaks. Chromatographia 2019, 82, 221–233. [Google Scholar] [CrossRef]
- García, M.Á.; Menéndez-López, N.; Boltes, K.; Castro-Puyana, M.; Marina, M.L. A capillary micellar electrokinetic chromatography method for the stereoselective quantitation of bioallethrin in biotic and abiotic samples. J. Chromatogr. A 2017, 1510, 108–116. [Google Scholar] [CrossRef]
- Ma, Y.; Chen, L.; Lu, X.; Chu, H.; Xu, C.; Liu, W. Enantioselectivity in aquatic toxicity of synthetic pyrethroid insecticide fenvalerate. Ecotoxicol. Environ. Saf. 2009, 72, 1913–1918. [Google Scholar] [CrossRef] [PubMed]
- Ham, J.; You, S.; Lim, W.; Song, G. Bifenthrin impairs the functions of leydig and sertoli cells in mice via mitochondrion-endoplasmic reticulum dysregulation. Environ. Pollut. 2020, 266, 115174. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Qiu, Y.; Yao, T.; Ma, Y.; Zhang, H.; Yang, X. Evaluation of seven chemical pesticides by mixed microbial culture (PCS-1): Degradation ability, microbial community, and medicago sativa phytotoxicity. J. Hazard. Mater. 2020, 389, 121834. [Google Scholar] [CrossRef]
- Bhatt, P.; Huang, Y.; Zhan, H.; Chen, S. Insight Into microbial applications for the biodegradation of pyrethroid insecticides. Front. Microbiol. 2019, 10, 1778. [Google Scholar] [CrossRef] [PubMed]
- Vaze, V.K. Biodegradation of Pesticide Cypermethrin by phosphatase and esterase enzymes produced by actinomycetes. Res. J. Pharm. Technol. 2017, 10, 445. [Google Scholar] [CrossRef]
- Castro-Gutiérrez, V.; Fuller, E.; Thomas, J.C.; Sinclair, C.J.; Johnson, S.; Helgason, T. Genomic basis for pesticide degradation revealed by selection, isolation and characterisation of a library of metaldehyde-degrading strains from soil. Soil Biol. Biochem. 2020, 142, 107702. [Google Scholar] [CrossRef]
- Zhao, J.; Jia, D.; Du, J.; Chi, Y.; Yao, K. Substrate regulation on co-metabolic degradation of β-cypermethrin by Bacillus licheniformis B-1. AMB Express 2019, 9, 83. [Google Scholar] [CrossRef] [PubMed]
- Cycoń, M.; Piotrowska-Seget, Z. Pyrethroid-degrading microorganisms and their potential for the bioremediation of contaminated soils: A review. Front. Microbiol. 2016, 7, 1463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gangola, S.; Sharma, A.; Bhatt, P.; Khati, P.; Chaudhary, P. Presence of esterase and laccase in Bacillus subtilis facilitates biodegradation and detoxification of cypermethrin. Sci. Rep. 2018, 8, 12755. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Hu, C.; Zhang, R.; Sun, A.; Li, D.; Shi, X. Mechanism study of cyfluthrin biodegradation by Photobacterium ganghwense with comparative metabolomics. Appl. Microbiol. Biotechnol. 2019, 103, 473–488. [Google Scholar] [CrossRef]
- Zhan, H.; Wang, H.; Liao, L.; Feng, Y.; Fan, X.; Zhang, L. Kinetics and novel degradation pathway of permethrin in Acinetobacter baumannii ZH-14. Front. Microbiol. 2018, 9, 98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, J.; Long, X.; Zhang, S.; Qin, Q.; Gan, L.; Tian, Y. Screening cyhalothrin degradation strains from locust epiphytic bacteria and studying Paracoccus acridae SCU-M53 cyhalothrin degradation process. Environ. Sci. Pollut. Res. Int. 2018, 25, 11505–11515. [Google Scholar] [CrossRef] [PubMed]
- Suleiman, M.; Schröder, C.; Kuhn, M.; Simon, A.; Stahl, A.; Frerichs, H. Microbial biofilm formation and degradation of octocrylene, a UV absorber found in sunscreen. Commun. Biol. 2019, 2, 430. [Google Scholar] [CrossRef] [Green Version]
- Drzyzga, O.; Fernández de las Heras, L.; Morales, V.; Navarro Llorens, J.M.; Perera, J. Cholesterol degradation by Gordonia cholesterolivorans. Appl. Environ. Microbiol. 2011, 77, 4802–4810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurniati, T.H.; Rusmana, I.; Suryani, A.; Mubarik, N.R. Degradation of polycyclic aromatic hydrocarbon pyrene by biosurfactant-producing bacteria Gordonia cholesterolivorans AMP 10. J. Bio. Bio. Edu. 2016, 8, 336. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Hu, Q.B.; Hu, M.Y.; Luo, J.J.; Weng, Q.F.; Lai, K.P. Isolation and characterization of a fungus able to degrade pyrethroids and 3-phenoxybenzaldehyde. Bioresour. Technol. 2011, 102, 8110–8116. [Google Scholar] [CrossRef]
- Birolli, W.G.; Vacondio, B.; Alvarenga, N.; Seleghim, M.H.R.; Porto, A.L.M. Enantioselective biodegradation of the pyrethroid (±)-lambda-cyhalothrin by marine-derived fungi. Chemosphere 2018, 197, 651–660. [Google Scholar] [CrossRef]
- Zhao, J.; Chim, Y.; Liu, F.; Jia, D.; Yao, K. Effects of two surfactants and beta-cyclodextrin on beta-cypermethrin degradation by Bacillus licheniformis B-1. J. Agric. Food Chem. 2015, 63, 10729–10735. [Google Scholar] [CrossRef]
- Xiao, Y.; Chen, S.H.; Gao, Y.; Hu, W.; Hu, M.Y.; Zhong, G.H. Isolation of a novel beta-cypermethrin degrading strain Bacillus subtilis BSF01 and its biodegradation pathway. Appl. Microbiol. Biotechnol. 2015, 99, 2849–2859. [Google Scholar] [CrossRef]
- Deng, W.; Lin, D.; Yao, K.; Yuan, H.; Wang, Z.; Li, J.; Zou, L.; Han, X.; Zhou, K.; He, L.; et al. Characterization of a novel β-cypermethrin-degrading Aspergillus niger YAT strain and the biochemical degradation pathway of β-cypermethrin. Appl. Microbiol. Biotechnol. 2015, 99, 8187–8198. [Google Scholar] [CrossRef]
- Zhan, H.; Huang, Y.; Lin, Z.; Bhatt, P.; Chen, S. New insights into the microbial degradation and catalytic mechanism of synthetic pyrethroids. Environ. Res. 2020, 182, 109138. [Google Scholar] [CrossRef]
- Zhao, J.; Jia, D.; Chi, Y.; Yao, K. Co-metabolic enzymes and pathways of 3-phenoxybenzoic acid degradation by Aspergillus oryzae M-4. Ecotoxicol. Environ. Saf. 2020, 189, 109953. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.; Hu, K.; Li, J.; Zhu, Y.; Liu, A.; Yao, K.; Liu, S. Current insights into the microbial degradation for pyrethroids: Strain safety, biochemical pathway, and genetic engineering. Chemosphere 2021, 279, 130542. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, P.; Rene, E.R.; Kumar, A.J.; Gangola, S.; Kumar, G.; Sharma, A. Fipronil degradation kinetics and resource recovery potential of Bacillus sp. strain FA4 isolated from a contaminated agricultural field in Uttarakhand, India. Chemosphere 2021, 276, 130156. [Google Scholar] [CrossRef] [PubMed]
- Birolli, W.G.; Arai, M.S.; Nitschke, M.; Porto, A.L.M. The pyrethroid (±)-lambda-cyhalothrin enantioselective biodegradation by a bacterial consortium. Pestic. Biochem. Phys. 2019, 156, 129–137. [Google Scholar] [CrossRef]
- Huang, Y.; Lin, Z.; Zhang, W.; Pang, S.; Bhatt, P.; Rene, E.R.; Kumar, A.J.; Chen, S. New insights into the microbial degradation of D-cyphenothrin in contaminated water/soil environments. Microorganisms 2020, 8, 473. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.; Chi, Y.; Wu, S.; Jia, D.; Yao, K. Simultaneous degradation of cypermethrin and its metabolite, 3-phenoxybenzoic acid, by the cooperation of Bacillus licheniformis B-1 and Sphingomonas sp. SC-1. J. Agric. Food Chem. 2014, 62, 8256–8262. [Google Scholar] [CrossRef]
- Cycoń, M.; Mrozik, A.; Piotrowska-Seget, Z. Bioaugmentation as a strategy for the remediation of pesticide-polluted soil: A review. Chemosphere 2017, 172, 52–71. [Google Scholar] [CrossRef]
- Chen, S.; Luo, J.; Hu, M.; Lai, K.; Geng, P.; Huang, H. Enhancement of cypermethrin degradation by a coculture of Bacillus cereus ZH-3 and Streptomyces aureus HP-S-01. Bioresour. Technol. 2013, 110, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Birolli, W.G.; Lima, R.N.; Porto, A.L.M. Applications of marine-derived microorganisms and their enzymes in biocatalysis and biotransformation, the underexplored potentials. Front. Microbiol. 2019, 10, 1453. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.C.; Liu, Y.H.; Wang, Z.Y.; Zhang, X.Y.; Li, H.; Liang, W.Q.; Luo, N.; Hu, J.M.; Lu, J.Q.; Luan, T.G.; et al. Molecular cloning, purification, and biochemical characterization of a novel pyrethroid-hydrolyzing esterase from Klebsiella sp. strain ZD112. J. Agric. Food Chem. 2006, 54, 836–842. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.H.; Luo, J.J.; Hu, M.Y.; Geng, P.; Zhang, Y.B. Microbial detoxification of bifenthrin by a novel yeast and its potential for contaminated soils treatment. PLoS ONE 2012, 7, e30862. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, Y.; Yoshida, M.; Sugano, T.; Shibata, A.; Kodaka, R.; Fujisawa, T.; Katagi, T. Behavior of cyphenothrin in aquatic environment. J. Pestic. Sci. 2017, 42, 17–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Jia, L.; Wang, S.H.; Qu, J.; Xu, L.L.; Shi, H.H.; Yan, Y.C. Biodegradation of beta-cypermethrin by two Serratia spp. with different cell surface hydrophobicity. Bioresour. Technol. 2010, 101, 3423–3429. [Google Scholar] [CrossRef]
- Bhatt, P.; Rene, E.R.; Kumar, A.J.; Zhang, W.; Chen, S. Binding interaction of allethrin with esterase: Bioremediation potential and mechanism. Bioresour. Technol. 2020, 315, 123845. [Google Scholar] [CrossRef]
- Birolli, W.G.; Dos Santos, A.; Pilau, E.; Rodrigues-Filho, E. New role for a commercially available bioinsecticide: Bacillus thuringiensis Berliner biodegrades the pyrethroid cypermethrin. Environ. Sci. Technol. 2021, 55, 4792–4803. [Google Scholar] [CrossRef] [PubMed]
- Tallur, P.N.; Megadi, V.B.; Ninnekar, H.Z. Biodegradation of cypermethrin by Micrococcus sp. strain CPN 1. Biodegradation 2008, 19, 77–82. [Google Scholar] [CrossRef]
- Chen, S.; Chang, C.; Deng, Y.; An, S.; Dong, Y.H.; Zhou, J.; Hu, M.; Zhong, G.; Zhang, L.H. Fenpropathrin biodegradation pathway in Bacillus sp. DG-02 and its potential for bioremediation of pyrethroid-contaminated soils. J. Agric. Food. Chem. 2014, 62, 2147–2157. [Google Scholar] [CrossRef]
- Lin, Q.S.; Chen, S.H.; Hu, M.Y.; Rizwan-ul-Haq, M.; Yang, L.; Li, H. Biodegradation of cypermethrin by a newly isolated actinomycetes HU-S-01 from wastewater sludge. Int. J. Environ. Sci. Technol. 2011, 8, 45–56. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.Z.; Ma, Y.; Zhou, W.Y.; Zheng, J.W.; He, J. Biodegradation of synthetic pyrethroids by Ochrobactrum tritici strain pyd-1. World J. Microbiol. Biotechnol. 2011, 27, 2315–2324. [Google Scholar] [CrossRef]
- Chen, S.; Geng, P.; Xiao, Y.; Hu, M. Bioremediation of β-cypermethrin and 3-phenoxybenzaldehyde contaminated soils using Streptomyces aureus HP-S-01. Appl. Microbiol. Biotechnol. 2012, 94, 505–515. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Chi, Y.; Xu, Y.; Jia, D.; Yao, K. Co-metabolic degradation of β-cypermethrin and 3-phenoxybenzoic acid by co-culture of Bacillus licheniformis B-1 and Aspergillus oryzae M-4. PLoS ONE 2016, 11, e166796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, Y.; Lu, Q.; Yi, X.; Zhong, G.; Liu, J. Synergistic degradation of pyrethroids by the quorum sensing-regulated carboxylesterase of Bacillus subtilis BSF01. Front. Bioeng. Biotechnol. 2020, 8, 889. [Google Scholar] [CrossRef]
- Cycoń, M.; Żmijowska, A.; Piotrowska-Seget, Z. Enhancement of deltamethrin degradation by soil bioaugmentation with two different strains of Serratia marcescens. Int. J. Environ. Sci. Technol. 2014, 11, 1305–1316. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Dong, Y.H.; Chang, C.; Deng, Y.; Zhang, X.F.; Zhong, G.; Song, H.; Hu, M.; Zhang, L.H. Characterization of a novel cyfluthrin-degrading bacterial strain Brevibacterium aureum and its biochemical degradation pathway. Bioresour. Technol. 2013, 132, 16–23. [Google Scholar] [CrossRef]
- Zhang, X.Q.; Hao, X.; Huo, S.; Lin, W.; Xia, X.; Liu, K.; Duan, B. Isolation and identification of the Raoultella ornithinolytica-ZK4 degrading pyrethroid pesticides within soil sediment from an abandoned pesticide plant. Arch. Microbiol. 2019, 201, 1207–1217. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Wang, S.; Yan, Y. Isomerization and biodegradation of beta-cypermethrin by Pseudomonas aeruginosa CH7 with biosurfactant production. Bioresour. Technol. 2011, 102, 7139–7146. [Google Scholar] [CrossRef]
- Yang, J.; Feng, Y.; Zhan, H.; Liu, J.; Yang, F.; Zhang, K.; Zhang, L.; Chen, S. Characterization of a pyrethroid-degrading Pseudomonas fulva strain P31 and biochemical degradation pathway of D-phenothrin. Front. Microbiol. 2018, 9, 1003. [Google Scholar] [CrossRef]
- Sevilla-Morán, B.; Calvo, L.; López-Goti, C.; Alonso-Prados, J.L.; Sandín-España, P. Photodegradation behaviour of sethoxydim and its commercial formulation PoastR under environmentally relevant conditions in aqueous media. Study of photoproducts and their toxicity. Chemosphere 2017, 168, 501–507. [Google Scholar] [CrossRef] [PubMed]
- Villaverde, J.J.; Santín-Montanyá, I.; Sevilla-Morán, B.; Alonso-Prados, J.L.; Sandín-España, P. Assessing the effects of alloxydim phototransformation products by QSAR models and a phototoxicity study. Molecules 2018, 23, 993. [Google Scholar] [CrossRef] [Green Version]
- Villaverde, J.J.; Sevilla-Morán, B.; Sandín-España, P.; López-Goti, C.; Alonso-Prados, J.L. Challenges of biopesticides under the European Regulation (EC) No. 1107/2009: An overview of new trends in residue analysis. Stud. Nat. Prod. Chem. 2014, 43, 437–482. [Google Scholar]
- Mishra, S.; Zhang, W.; Lin, Z.; Pang, S.; Huang, Y.; Bhatt, P.; Chen, S. Carbofuran toxicity and its microbial degradation in contaminated environments. Chemosphere 2020, 259, 127419. [Google Scholar] [CrossRef]
- Huang, Y.; Zhang, W.; Pang, S.; Chen, J.; Bhatt, P.; Mishra, S.; Chen, S. Insights into the microbial degradation and catalytic mechanisms of chlorpyrifos. Environ. Res. 2021, 194, 110660. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, P.; Joshi, T.; Bhatt, K.; Zhang, W.; Huang, Y.; Chen, S. Binding interaction of glyphosate with glyphosate oxidoreductase and C–P lyase: Molecular docking and molecular dynamics simulation studies. J. Hazard. Mater. 2021, 409, 124927. [Google Scholar] [CrossRef]
- Tang, A.; Wang, B.; Liu, Y.; Li, Q.; Tong, Z.; Wei, Y. Biodegradation and extracellular enzymatic activities of Pseudomonas aeruginosa strain GF31 on β-cypermethrin. Environ. Sci. Pollut. Res. Int. 2015, 22, 13049–13057. [Google Scholar] [CrossRef]
- Liu, X.; Liang, M.; Liu, Y.; Fan, X. Directed evolution and secretory expression of a pyrethroid-hydrolyzing esterase with enhanced catalytic activity and thermostability. Microb. Cell Factories 2017, 16, 81. [Google Scholar] [CrossRef]
- Bhatt, P.; Bhandari, G.; Bhatt, K.; Maithani, D.; Mishra, S.; Gangola, S.; Bhatt, R.; Huang, Y.; Chen, S. Plasmid-mediated catabolism for the removal of xenobiotics from the environment. J. Hazard. Mater. 2021, 420, 126618. [Google Scholar] [CrossRef] [PubMed]
- Maloney, S.E.; Maule, A.; Smith, A.R. Microbial transformation of the pyrethroid insecticides: Permethrin, deltamethrin, fastac, fenvalerate, and fluvalinate. Appl. Environ. Microb. 1988, 54, 2874–2876. [Google Scholar] [CrossRef] [Green Version]
- Bhatt, P.; Bhatt, K.; Sharma, A.; Zhang, W.; Mishra, S.; Chen, S. Biotechnological basis of microbial consortia for the removal of pesticides from the environment. Crit. Rev. Biotechnol. 2021, 41, 317–338. [Google Scholar] [CrossRef]
- Wang, B.; Guo, P.; Hang, B.; Li, L.; He, J.; Wang, B.; Li, S.P. Cloning of a novel pyrethroid-hydrolyzing carboxylesterase gene from Sphingobium sp. strain JZ-1. Appl. Environ. Microbiol. 2009, 75, 5496–5500. [Google Scholar] [CrossRef] [Green Version]
- Bhatt, P.; Bhatt, K.; Huang, Y.; Lin, Z.; Chen, S. Esterase is a powerful tool for the biodegradation of pyrethroid insecticides. Chemosphere 2020, 244, 125507. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Navarro, A.I.; Perrella, L.; Cedergreen, N. Can organophosphates and carbamates cause synergism by inhibiting esterases responsible for biotransformation of pyrethroids? Environ. Sci. Technol. 2021, 55, 1585–1593. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, P.; Zhou, X.; Huang, Y.; Zhang, W.; Chen, S. Characterization of the role of esterases in the biodegradation of organophosphate, carbamate, and pyrethroid pesticides. J. Hazard. Mater. 2021, 411, 125026. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Gao, Y.; Sun, B.; Ran, T.; Zeng, L.; He, J.; He, J.; Wang, W. Pyrethroid carboxylesterase PytH from Sphingobium faniae JZ-2: Structure and catalytic mechanism. Appl. Environ. Microbiol. 2020, 86, e02971-19. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Feng, Y.; Zhang, W.; Pang, S.; Lin, Z.; Zhang, Y.; Huang, Y.; Bhatt, P.; Chen, S. Kinetics and new mechanism of azoxystrobin biodegradation by an Ochrobactrum anthropi strain SH14. Microorganisms 2020, 8, 625. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, P.; Huang, Y.; Zhang, W.; Sharma, A.; Chen, S. Enhanced cypermethrin degradation kinetics and metabolic pathway in Bacillus thuringiensis strain SG4. Microorganisms 2020, 8, 223. [Google Scholar] [CrossRef] [Green Version]
- Birolli, W.G.; Alvarenga, N.; Seleghim, M.H.R.; Porto, A.L.M. Biodegradation of the pyrethroid pesticide esfenvalerate by marine-derived fungi. Mar. Biotechnol. 2016, 18, 511–520. [Google Scholar] [CrossRef]
- Bhatt, P.; Zhang, W.; Lin, Z.; Pang, S.; Huang, Y.; Chen, S. Biodegradation of allethrin by a novel fungus Fusarium proliferatum strain CF2, isolated from contaminated soils. Microorganims 2020, 8, 593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aiello, F.; Simons, M.G.; van Velde, J.W.; Dani, P. New insights into the degradation path of deltamethrin. Molecules 2021, 26, 3811. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Run | X1 | X2 | X3 | Y |
|---|---|---|---|---|
| 1 | 5 | 20 | 0.6 | 86.7 |
| 2 | 9 | 20 | 0.6 | 93.6 |
| 3 | 5 | 40 | 0.6 | 90.7 |
| 4 | 9 | 40 | 0.6 | 100 |
| 5 | 5 | 30 | 0.3 | 85.5 |
| 6 | 5 | 30 | 0.9 | 91.5 |
| 7 | 9 | 30 | 0.3 | 100 |
| 8 | 9 | 30 | 0.9 | 100 |
| 9 | 7 | 20 | 0.3 | 88.4 |
| 10 | 7 | 40 | 0.3 | 93.7 |
| 11 | 7 | 20 | 0.9 | 94.2 |
| 12 | 7 | 40 | 0.9 | 100 |
| 13 | 7 | 30 | 0.6 | 97.1 |
| 14 | 7 | 30 | 0.6 | 96.6 |
| 15 | 7 | 30 | 0.6 | 98.5 |
| Source | DF | SS | MS | F-Value | p-Value * |
|---|---|---|---|---|---|
| Model | 9 | 341.1 | 37.9 | 15.3 | 0.0039 |
| X1 | 1 | 194.1 | 194.1 | 78.4 | 0.0003 |
| X2 | 1 | 57.4 | 57.4 | 23.2 | 0.0048 |
| X3 | 1 | 41.0 | 41.0 | 16.5 | 0.0096 |
| X1X2 | 1 | 1.3 | 1.3 | 0.55 | 0.4887 |
| X1X3 | 1 | 9.0 | 9.0 | 3.6 | 0.1139 |
| X2X3 | 1 | 0.06 | 0.06 | 0.02 | 0.8799 |
| X12 | 1 | 18.5 | 18.5 | 7.5 | 0.0408 |
| X22 | 1 | 21.0 | 21.0 | 8.5 | 0.0331 |
| X32 | 1 | 2.9 | 2.9 | 1.2 | 0.3235 |
| Residual | 7 | 12.3 | 2.4 | ||
| Lack of Fit | 3 | 10.3 | 3.4 | 3.4 | 0.2320 |
| Pure Error | 4 | 2 | 0.99 | ||
| Cor Total | 16 | 353.4 |
| Treatments | Regression Equation | k | R2 | t1/2 |
|---|---|---|---|---|
| Sterilized soil slurry | Ct = 50.6e−0.0048t | 0.0048 | 0.9304 | 144.4 ± 1.1 |
| Unsterilized soil slurry | Ct = 51.1e−0.0054t | 0.0054 | 0.8425 | 128.3 ± 0.7 |
| Sterilized soil slurry + A16 | Ct = 51.2e−0.0373t | 0.0373 | 0.7004 | 18.6 ± 1.0 |
| Unsterilized soil slurry + A16 | Ct = 50.1e−0.0396t | 0.0396 | 0.7999 | 17.5 ± 1.5 |
| Code | RT (Minutes) | m/z | Compound Structure | Chemical Name |
|---|---|---|---|---|
| A | 26.0 | 331.2 |  | Tetramethrin |
| B | 23.1 | 154.1 |  | Cyclopropylmethanol |
| C | 25.9 | 207.1 |  | 2-Butyl-4,5,6,7-tetrahydro-1H-isoindole-1,3(2H)-dione |
| D | 23.5 | 71.0 |  | Acrylamide |
| E | 21.3 | 87.1 |  | N-ethylacetamide |
| Independent Variables | Code | Code Levels of Variables | ||
|---|---|---|---|---|
| −1 | 0 | 1 | ||
| Temperature (°C) | X1 | 20 | 30 | 40 |
| pH | X2 | 5 | 7 | 9 |
| Inoculum size (OD600) | X3 | 0.3 | 0.6 | 0.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, Y.; Huang, Y.; Pang, S.; Zhou, T.; Lin, Z.; Yu, H.; Zhang, G.; Bhatt, P.; Chen, S. Novel Mechanism and Kinetics of Tetramethrin Degradation Using an Indigenous Gordonia cholesterolivorans A16. Int. J. Mol. Sci. 2021, 22, 9242. https://doi.org/10.3390/ijms22179242
Guo Y, Huang Y, Pang S, Zhou T, Lin Z, Yu H, Zhang G, Bhatt P, Chen S. Novel Mechanism and Kinetics of Tetramethrin Degradation Using an Indigenous Gordonia cholesterolivorans A16. International Journal of Molecular Sciences. 2021; 22(17):9242. https://doi.org/10.3390/ijms22179242
Chicago/Turabian StyleGuo, Yuxin, Yaohua Huang, Shimei Pang, Tianhao Zhou, Ziqiu Lin, Hongxiao Yu, Guorui Zhang, Pankaj Bhatt, and Shaohua Chen. 2021. "Novel Mechanism and Kinetics of Tetramethrin Degradation Using an Indigenous Gordonia cholesterolivorans A16" International Journal of Molecular Sciences 22, no. 17: 9242. https://doi.org/10.3390/ijms22179242