
Key Enzymes Involved in the Synthesis of Hops Phytochemical Compounds: From Structure, Functions to Applications


Abstract
:1. Introduction
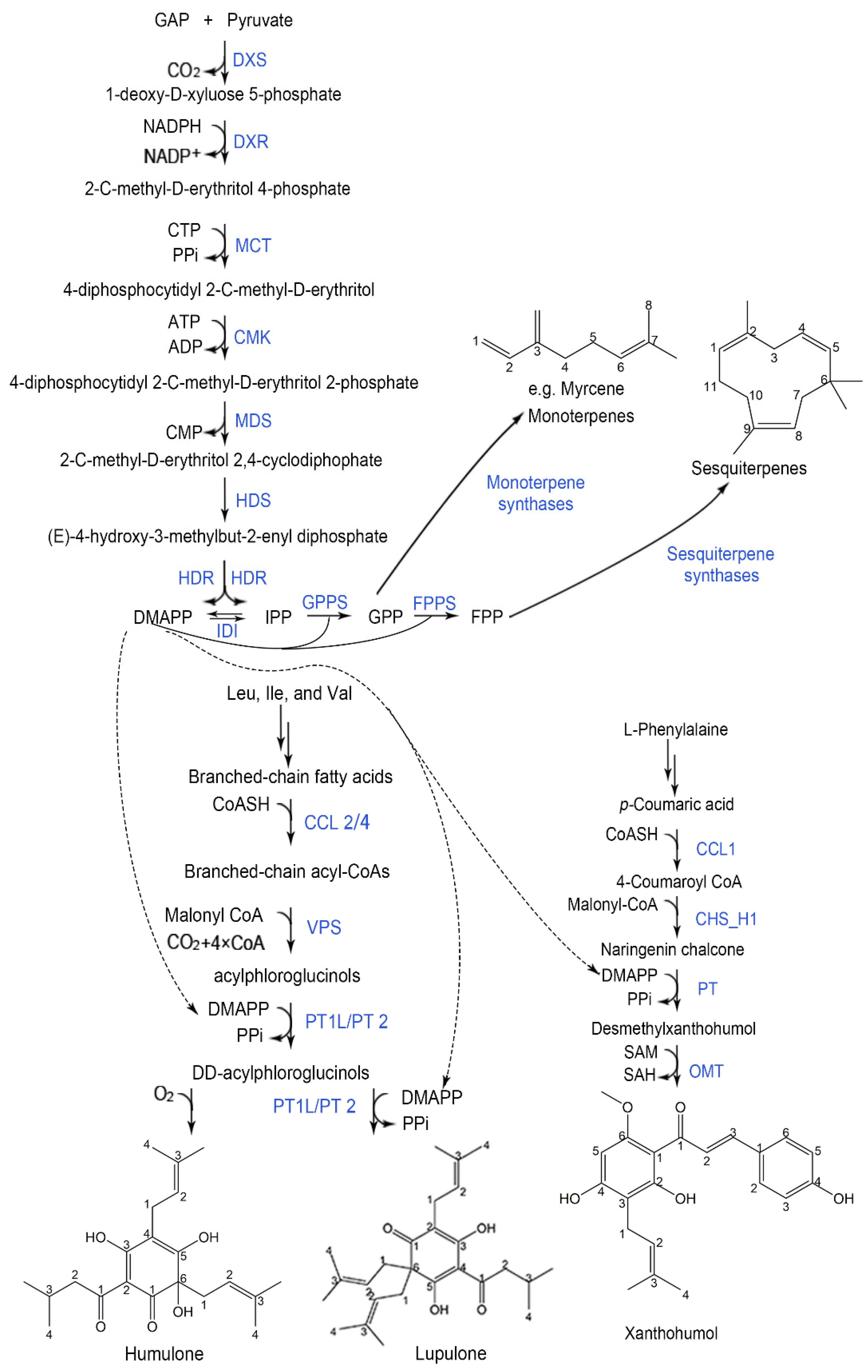
2. Hop Bitter Acids
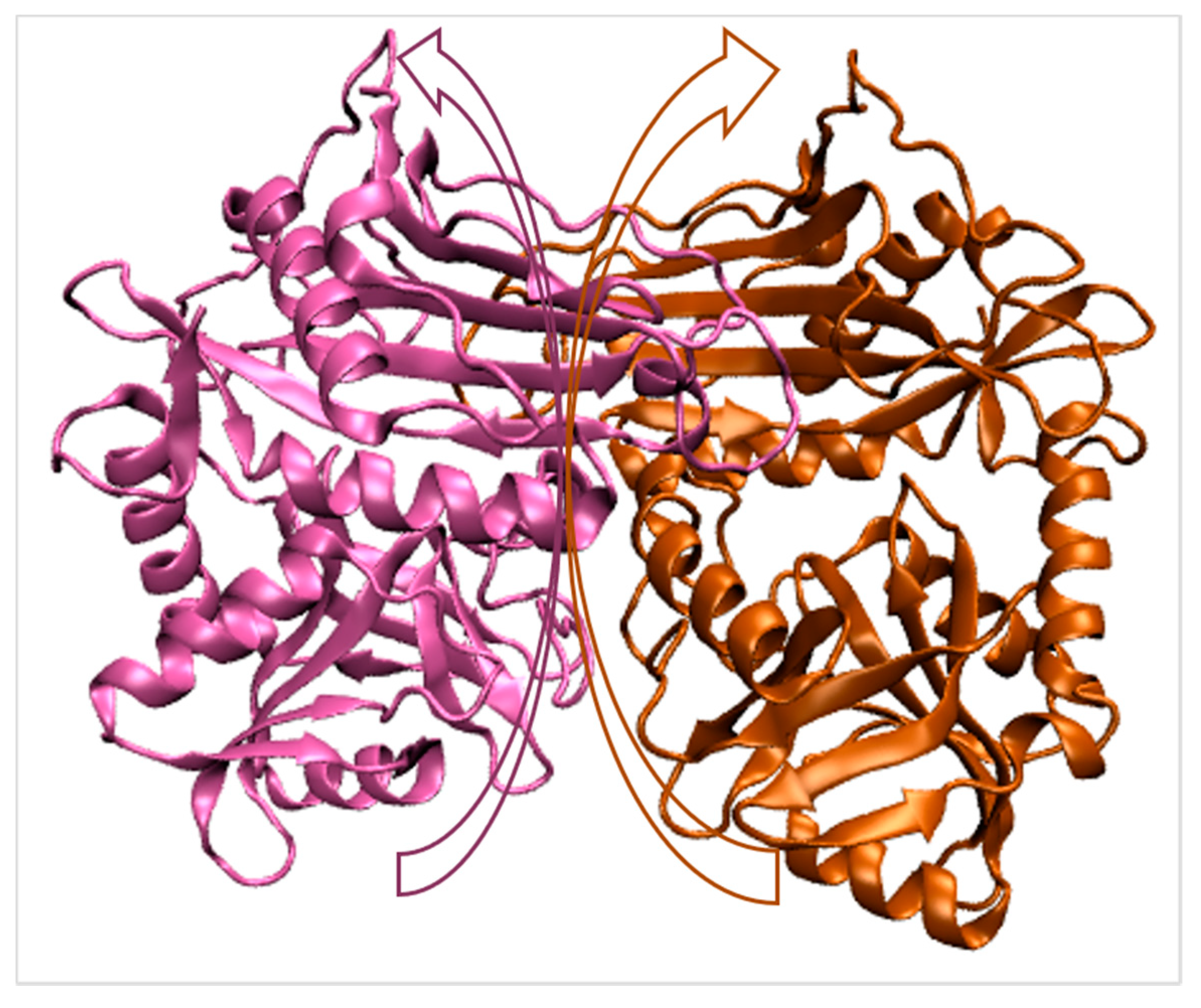
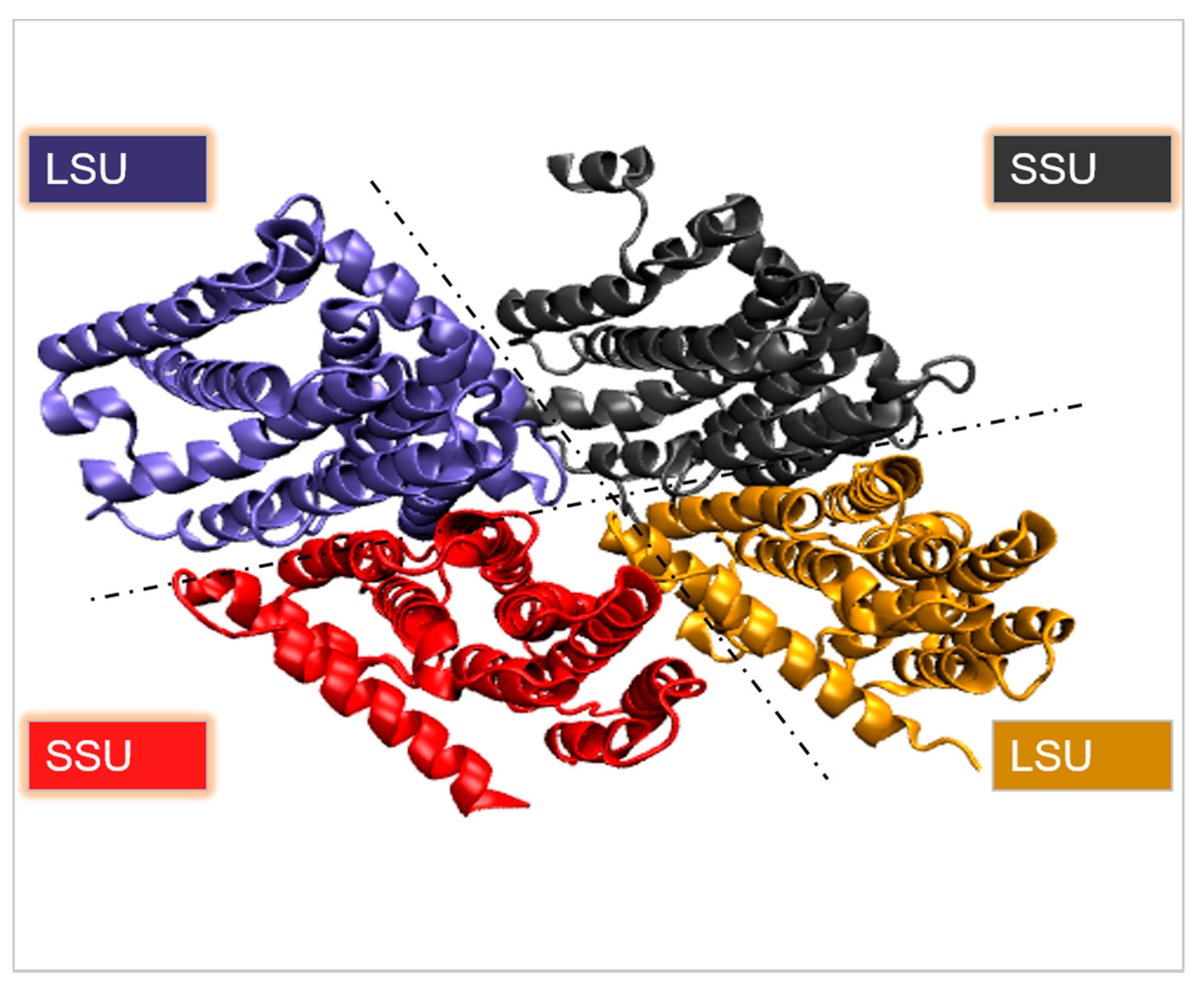
2.1. Branched-Chain Aminotransferase (BCAT)
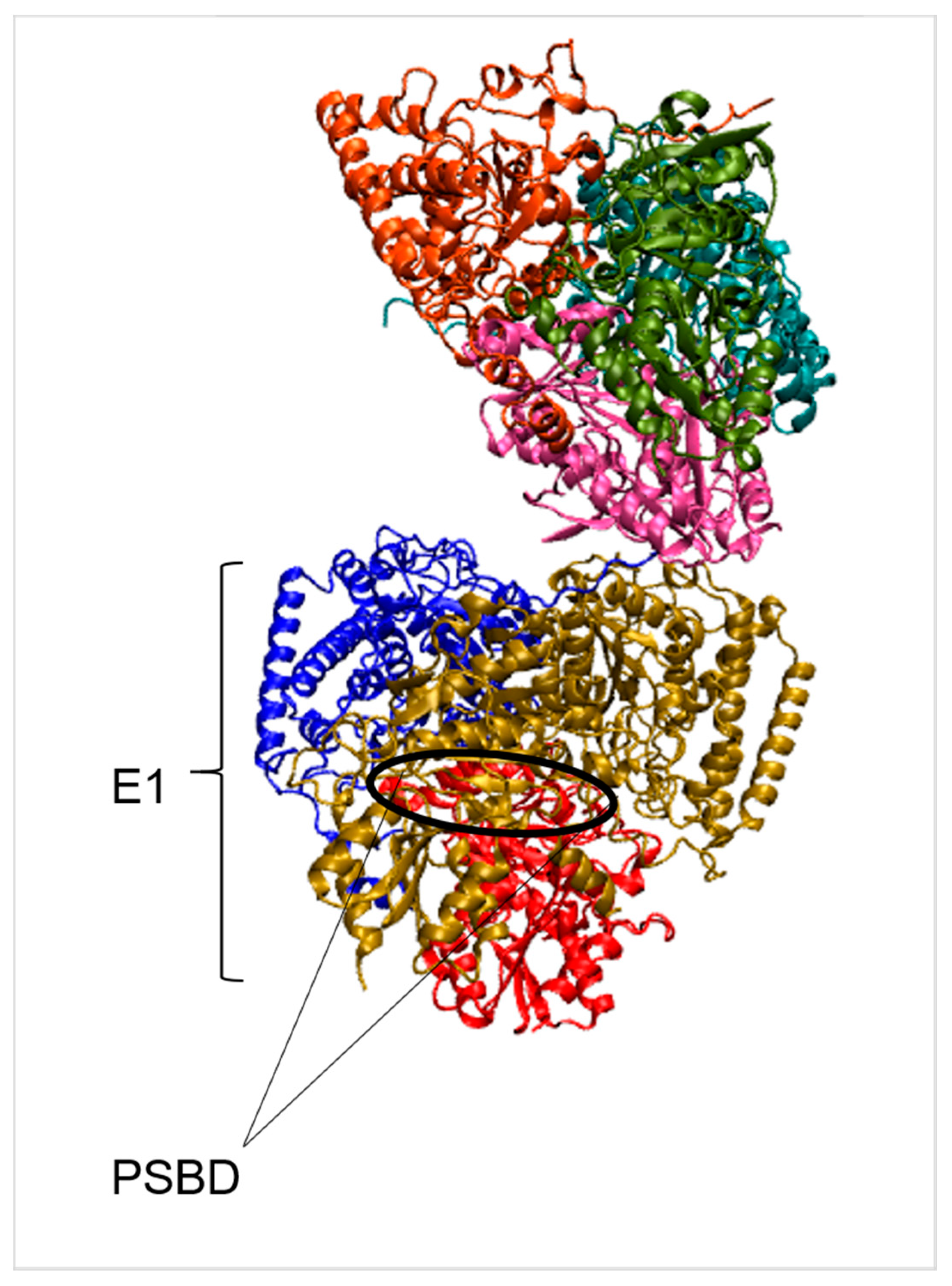
2.2. Branched-Chain Keto-Acid Dehydrogenase (BCKDH)
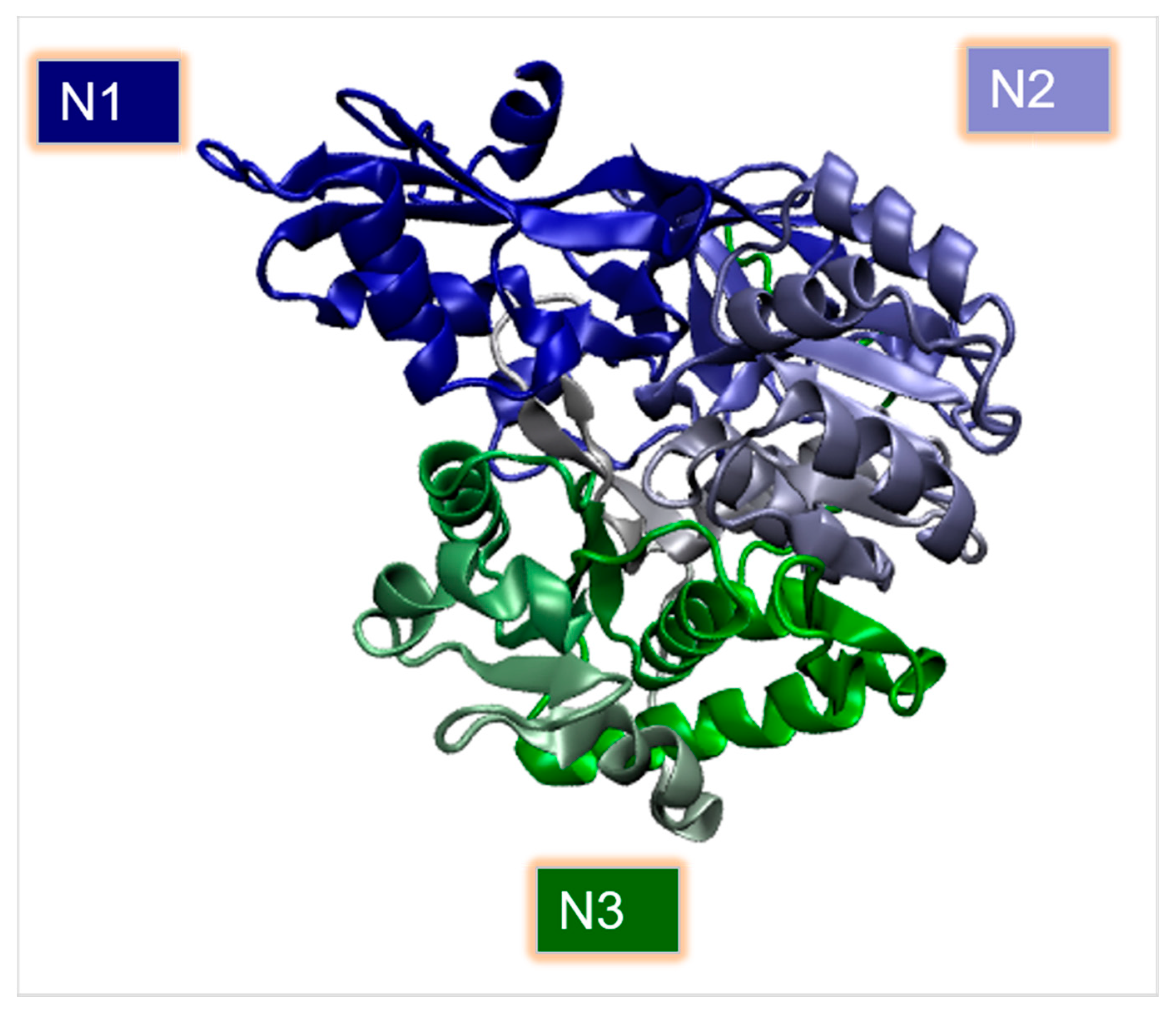
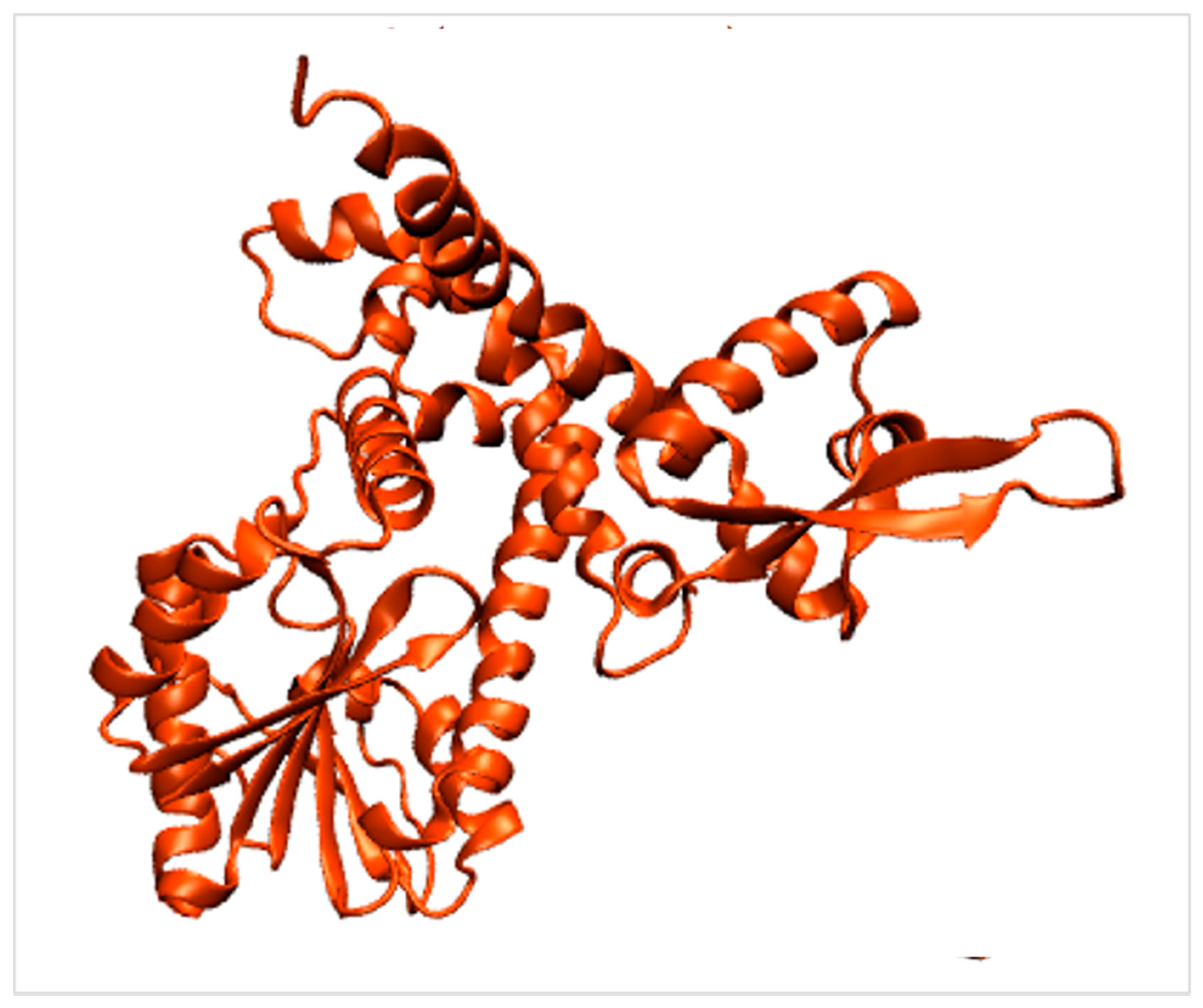
2.3. Carboxyl CoA Ligase (CCL)
2.4. Valerophenone Synthase (VPS)
2.5. Prenyltransferase (PT)
3. Terpene Compounds
3.1. 1-Deoxyxylulose-5-phosphate Synthase (DXS)
3.2. 4-Hydroxy-3-methylbut-2-enyl Diphosphate Synthase (HDS)/4-Hydroxy-3-methylbut-2-enyl Diphosphate Reductase (HDR)
3.3. Geranyl Diphosphate Synthase (GPPS)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hop Compounds | Enzyme | Organism | Expression System | Reference |
|---|---|---|---|---|
| Hop bitter acids | BCAT | Escherichia coli/Mice | - | [26] |
| BCKDH | Pseudomonas putida | - | [37] | |
| CCL2, CCL4 | Populus tomentosa | Escherichia coli | [50] | |
| VPS | Humulus lupulus | Escherichia coli | [57] | |
| PT | Humulus lupulus | insect cells | [59] | |
| Terpene compounds | DXS | Picea abies | - | [70] |
| GPPS | Mentha x piperita | - | [94] | |
| MTS | Humulus lupulus | - | - | |
| LIS, GES | Mint and basil | yeast | [95] | |
| Xanthohumol derivatives. | C4H | Humulus lupulus | yeast | [96] |
| CHS_H1 | Humulus lupulus | yeast | - | |
| CHIL | Humulus lupulus | yeast | - | |
| OMT | Medicago truncatula | Escherichia coli | [97,98] |
3.4. Monoterpene Synthase (MTS)
4. Xanthohumol Derivatives
4.1. Cinnamate 4-hydroxylase (C4H)
4.2. Naringenin-Chalcone Synthase (CHS)/Chalcone Isomerase (CHI)-Like Proteins
4.3. O-methyltransferase (OMT)
5. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AMP | Adenosine 5′-monophosphate |
| BCAA | Branched-chain amino acids |
| BCAT | Branched-chain aminotransferase |
| BCKDH | Branched-chain keto-acid dehydrogenase |
| C4H | Cinnamate 4-hydroxylase |
| CCL | Carboxyl CoA ligase |
| CHI | Chalcone isomerase |
| CHIL | Chalcone isomerase-like proteins |
| CHS | Chalcone synthase |
| DMAPP | Dimethylallyl diphosphate |
| DXR | 1-deoxy-d-xylulose-5-phosphate reductoisomerase |
| DXS | 1-deoxyxylulose-5-phosphate synthase |
| GES | Geraniol synthase |
| GGPP | Geranylgeranyl pyrophosphate |
| GPP | Geranyl diphosphate |
| GPPS | Geranyl diphosphate synthase |
| HDR | 4-hydroxy-3-methylbut-2-enyl diphosphate reductase |
| HDS | 4-hydroxy-3-methylbut-2-enyl diphosphate synthase |
| HlBCAT | Branched-chain aminotransferase in Humulus lupulus |
| HlCCL | Carboxyl CoA ligase gene in Humulus Lupulus |
| HlCHIL | Chalcone isomerase-like proteins in Humulus Lupulus |
| HlMTS | Monoterpene synthases in Humulus Lupulus |
| HlPT | Prenyltransferase in Humulus Lupulus |
| HMBPP | 4-hydroxy-3-methylbut-2-enyl diphosphate |
| IDI | IPP/DMAPP isomerase |
| IPP | Isopentenyl diphosphate |
| IspG | Isoprenoid synthesis G |
| IspH | Isoprenoid synthesis H |
| LIS | linalool synthase |
| MEP | Methylerythritol phosphate |
| MTS | Monoterpene synthase |
| MVA | Mevalonate |
| OMT1 | O-methyltransferase |
| PSBD | Peripheral subunit binding domain |
| PT | Prenyltransferase |
| VPS | Valerophenone synthase |
References
- Almaguer, C.; Schönberger, C.; Gastl, M.; Arendt, E.K.; Becker, T. Humulus lupulus—A story that begs to be told. A review. J. Inst. Brew. 2014, 120, 289–314. [Google Scholar] [CrossRef]
- Dresel, M.; Vogt, C.; Dunkel, A.; Hofmann, T. The Bitter chemodiversity of hops (Humulus lupulus L.). J. Agric. Food Chem. 2016, 64, 7789–7799. [Google Scholar] [CrossRef]
- Stevens, J.F.; Ivancic, M.; Hsu, V.L.; Deinzer, M.L. Prenylflavonoids from Humulus lupulus. Phytochemistry 1997, 44, 1575–1585. [Google Scholar] [CrossRef]
- Jaskula, B.; Kafarski, P.; Aerts, G.; De Cooman, L. A kinetic study on the isomerization of hop α-Acids. J. Agric. Food Chem. 2008, 56, 6408–6415. [Google Scholar] [CrossRef] [PubMed]
- Collin, S.; Jerkovic, V.; Bröhan, M.; Callemien, D. Polyphenols and Beer Quality. In Natural Products; Ramawat, K., Mérillon, J.M., Eds.; Springer: Berlin, Heidelberg, 2013; pp. 2333–2359. [Google Scholar]
- Drewnowski, A.; Gomez-Carneros, C. Bitter taste, phytonutrients, and the consumer: A review. Am. J. Clin. Nutr. 2000, 72, 1424–1435. [Google Scholar] [CrossRef] [PubMed]
- Floridi, S.; Montanari, L.; Marconi, O.; Fantozzi, P. Determination of free phenolic acids in wort and beer by coulometric array detection. J. Agric. Food Chem. 2003, 51, 1548–1554. [Google Scholar] [CrossRef] [PubMed]
- Yoshimaru, T.; Komatsu, M.; Tashiro, E.; Imoto, M.; Osada, H.; Miyoshi, Y.; Honda, J.; Sasa, M.; Katagiri, T. Xanthohumol suppresses oestrogen-signalling in breast cancer through the inhibition of BIG3-PHB2 interactions. Sci. Rep. 2015, 4, 7355. [Google Scholar] [CrossRef] [Green Version]
- Costa, R.; Rodrigues, I.; Guardão, L.; Rocha-Rodrigues, S.; Silva, C.; Magalhães, J.; Ferreira-de-Almeida, M.; Negrão, R.; Soares, R. Xanthohumol and 8-prenylnaringenin ameliorate diabetic-related metabolic dysfunctions in mice. J. Nutr. Biochem. 2017, 45, 39–47. [Google Scholar] [CrossRef]
- Hartkorn, A.; Hoffmann, F.; Ajamieh, H.; Vogel, S.; Heilmann, J.; Gerbes, A.L.; Vollmar, A.M.; Zahler, S. Antioxidant effects of xanthohumol and functional impact on hepatic ischemia-reperfusion injury. J. Nat. Prod. 2009, 72, 1741–1747. [Google Scholar] [CrossRef] [Green Version]
- Eyres, G.; Dufour, J. Hop essential oil: Analysis, chemical composition and odor characteristics. In Beer in Health and Disease Prevention; Academic Press: Cambridge, MA, USA, 2009; pp. 239–254. [Google Scholar]
- Sharp, D.C.; Townsend, M.S.; Qian, Y.; Shellhammer, T.H. Effect of harvest maturity on the chemical composition of cascade and willamette hops. J. Am. Soc. Brew. Chem. 2014, 72, 231–238. [Google Scholar] [CrossRef]
- Steinhaus, M.; Schieberle, P. Comparison of the most odor-active compounds in fresh and dried hop cones (Humulus lupulus L. variety spalter select) based on GC−olfactometry and odor dilution techniques. J. Agric. Food Chem. 2000, 48, 1776–1783. [Google Scholar] [CrossRef]
- Lermusieau, G.; Bulens, M.; Collin, S. Use of GC-olfactometry to identify the hop aromatic compounds in beer. J. Agric. Food Chem. 2001, 49, 3867–3874. [Google Scholar] [CrossRef]
- Stevens, R. The chemistry of hop constituents. Chem. Rev. 1982, 67, 19–71. [Google Scholar] [CrossRef]
- De Keukeleire, J.; Janssens, I.; Heyerick, A.; Ghekiere, G.; Cambie, J.; Roldán-Ruiz, I.; Van Bockstaele, E.; De Keukeleire, D. Relevance of organic farming and effect of climatological conditions on the formation of α-Acids, β-Acids, desmethylxanthohumol, and xanthohumol in hop (Humulus lupulus L.). J. Agric. Food Chem. 2007, 55, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Natsume, S.; Takagi, H.; Shiraishi, A.; Murata, J.; Toyonaga, H.; Patzak, J.; Takagi, M.; Yaegashi, H.; Uemura, A.; Mitsuoka, C.; et al. The draft genome of hop (Humulus lupulus), an essence for brewing. Plant Cell Physiol. 2015, 56, 428–441. [Google Scholar] [CrossRef] [Green Version]
- Skinner, R.N.; Kavanagh, T.E.; Hildebrand, R.P.; Clarke, B.J. Hop maturation and resin formation: Field trials. J. Sci. Food Agric. 1974, 25, 1121–1133. [Google Scholar] [CrossRef]
- Matsui, H.; Inui, T.; Oka, K.; Fukui, N. The influence of pruning and harvest timing on hop aroma, cone appearance, and yield. Food Chem. 2016, 202, 15–22. [Google Scholar] [CrossRef]
- Clark, S.M.; Vaitheeswaran, V.; Ambrose, S.J.; Purves, R.W.; Page, J.E. Transcriptome analysis of bitter acid biosynthesis and precursor pathways in hop (Humulus lupulus). BMC Plant Biol. 2013, 13, 12. [Google Scholar] [CrossRef] [Green Version]
- Binder, S.; Knill, T.; Schuster, J. Branched-chain amino acid metabolism in higher plants. Physiol. Plantarum. 2007, 129, 68–78. [Google Scholar] [CrossRef]
- Diebold, R.; Schuster, J.; Daschner, K.; Binder, S. The branched-chain amino acid transaminase gene family in Arabidopsis encodes plastid and mitochondrial proteins. Plant Physiol. 2002, 129, 540–550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maloney, G.S.; Kochevenko, A.; Tieman, D.M.; Tohge, T.; Krieger, U.; Zamir, D.; Taylor, M.G.; Fernie, A.R.; Klee, H.J. Characterization of the branched-chain amino acid aminotransferase enzyme family in tomato. Plant Physiol. 2010, 153, 925–936. [Google Scholar] [CrossRef] [Green Version]
- Kochevenko, A.; Klee, H.J.; Fernie, A.R.; Araújo, W.L. Molecular identification of a further branched-chain aminotransferase 7 (BCAT7) in tomato plants. J. Plant Physiol. 2012, 169, 437–443. [Google Scholar] [CrossRef]
- Okada, K.; Hirotsu, K.; Sato, M.; Hayashi, H.; Kagamiyama, H. Three-dimensional structure of Escherichia coli branched-chain amino acid aminotransferase at 2.5 angstrom resolution. J. Biochem. 1997, 121, 637–641. [Google Scholar] [CrossRef]
- Bertrand, S.M.; Ancellin, N.; Beaufils, B.; Bingham, R.P.; Borthwick, J.A.; Boullay, A.; Boursier, E.; Carter, P.S.; Chung, C.; Churcher, I.; et al. The Discovery of in vivo active mitochondrial branched-chain aminotransferase (BCATm) inhibitors by hybridizing fragment and HTS hits. J. Med. Chem. 2015, 58, 7140–7163. [Google Scholar] [CrossRef] [PubMed]
- Tremblay, L.W.; Blanchard, J.S. The 1.9Å structure of the branched-chain amino-acid transaminase (IlvE) from Mycobacterium tuberculosis. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2009, 65, 1071–1077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyko, K.M.; Stekhanova, T.N.; Nikolaeva, A.Y.; Mardanov, A.V.; Rakitin, A.L.; Ravin, N.V.; Bezsudnova, E.Y.; Popov, V.O. First structure of archaeal branched-chain amino acid aminotransferase from Thermoproteus uzoniensis specific for l-amino acids and R-amines. Extremophiles 2016, 20, 215–225. [Google Scholar] [CrossRef] [PubMed]
- Hutson, S. Structure and Function of Branched Chain Aminotransferases; Academic Press: Cambridge, MA, USA, 2001. [Google Scholar] [CrossRef]
- Okada, K.; Hirotsu, K.; Hayashi, H.; Kagamiyama, H. Structures of Escherichia coli branched-chain amino acid aminotransferase and its complexes with 4-methylvalerate and 2-methylleucine: Induced fit and substrate recognition of the enzyme. Biochemistry 2001, 40, 7453–7463. [Google Scholar] [CrossRef]
- Castell, A.; Mille, C.; Unge, T. Structural analysis of mycobacterial branched-chain aminotransferase: Implications for inhibitor design. Acta Crystallogr. Sect. D Biol. Crystallogr. 2010, 66, 549–557. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.D.; Lin, C.H.; Chuankhayan, P.; Huang, Y.C.; Hsieh, Y.C.; Huang, T.F.; Guan, H.H.; Liu, M.Y.; Chang, W.C.; Chen, C.J. Crystal structures of complexes of the branched-chain aminotransferase from Deinococcus radiodurans with α-ketoisocaproate and L-glutamate suggest the radiation resistance of this enzyme for catalysis. J. Bacteriol. 2012, 194, 6206–6216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slocombe, S.P.; Schauvinhold, I.; McQuinn, R.P.; Besser, K.; Welsby, N.A.; Harper, A.; Aziz, N.; Li, Y.; Larson, T.R.; Giovannoni, J.; et al. Transcriptomic and reverse genetic analyses of branched-chain fatty acid and acyl sugar production in Solanum pennellii and Nicotiana benthamiana. Plant Physiol. 2008, 148, 1830–1846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mooney, B.P.; Miernyk, J.A.; Randall, D.D. The complex fate of α-ketoacids. Annu. Rev. Plant Biol. 2002, 53, 357–375. [Google Scholar] [CrossRef]
- Lutziger, I.; Oliver, D.J. Characterization of two cDNAs encoding mitochondrial lipoamide dehydrogenase from arabidopsis. Plant Physiol. 2001, 127, 615–623. [Google Scholar] [CrossRef] [PubMed]
- Perham, R.N. Swinging arms and swinging domains in multifunctional enzymes: Catalytic machines for multistep reactions. Annu. Rev. Biochem. 2000, 69, 961–1004. [Google Scholar] [CrossRef] [PubMed]
- Frank, R.A.W.; Pratap, J.V.; Pei, X.Y.; Perham, R.N.; Luisi, B.F. The molecular origins of specificity in the assembly of a multienzyme complex. Structure 2005, 13, 1119–1130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perham, R.N.; Packman, L.C. 2-Oxo acid dehydrogenase multienzyme complexes: Domains, dynamics, and design. Ann. N. Y. Acad. Sci. 1989, 573, 1–20. [Google Scholar] [CrossRef]
- Perham, R.N. Domains, motifs, and linkers in 2-oxo acid dehydrogenase multienzyme complexes: A paradigm in the design of a multifunctional protein. Biochemistry 1991, 30, 8501–8512. [Google Scholar] [CrossRef]
- Lessard, I.A.D.; Fuller, C.; Perham, R.N. Competitive interaction of component enzymes with the peripheral subunit-binding domain of the pyruvate dehydrogenase multienzyme complex of Bacillus stearothermophilus: Kinetic analysis using surface plasmon resonance detection. Biochemistry 1996, 35, 16863–16870. [Google Scholar] [CrossRef]
- Hawkins, C.F.; Borges, A.; Perham, R.N. A common structural motif in thiamin pyrophosphate-binding enzymes. FEBS Lett. 1989, 255, 77–82. [Google Scholar] [CrossRef] [Green Version]
- Fries, M.; Chauhan, H.J.; Domingo, G.J.; Jung, H.; Perham, R.N. Site-directed mutagenesis of a loop at the active site of E1 (α2β2) of the pyruvate dehydrogenase complex. A possible common sequence motif. Eur. J. Biochem. 2003, 270, 861–870. [Google Scholar] [CrossRef]
- Hawes, J.W.; Schnepf, R.J.; Jenkins, A.E.; Shimomura, Y.; Popov, K.M.; Harris, R.A. Roles of amino acid residues surrounding phosphorylation site 1 of branched-chain alpha-ketoacid dehydrogenase (BCKDH) in catalysis and phosphorylation site recognition by BCKDH kinase. J. Biol. Chem. 1995, 270, 31071–31076. [Google Scholar] [CrossRef] [Green Version]
- Binder, S. Branched-Chain Amino Acid Metabolism in Arabidopsis thaliana. Arab. Book 2010, 8, e0137. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Zhang, F.; Liu, B.; Huhman, D.V.; Sumner, L.W.; Dixon, R.A.; Wang, G. Characterization of the formation of branched short-chain fatty acid: CoAs for bitter acid biosynthesis in hop glandular trichomes. Mol. Plant 2013, 6, 1301–1317. [Google Scholar] [CrossRef]
- Wang, G.; Tian, L.; Aziz, N.; Broun, P.; Dai, X.; He, J.; King, A.; Zhao, P.X.; Dixon, R.A. Terpene biosynthesis in glandular trichomes of hop. Plant Physiol. 2008, 148, 1254–1266. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Ban, Z.; Qin, H.; Ma, L.; King, A.J.; Wang, G. A heteromeric membrane-bound prenyltransferase complex from hop catalyzes three sequential aromatic prenylations in the bitter acid pathway. Plant Physiol. 2015, 167, 650–659. [Google Scholar] [CrossRef]
- Shockey, J.; Browse, J. Genome-level and biochemical diversity of the acyl-activating enzyme superfamily in plants. Plant J. 2011, 66, 143–160. [Google Scholar] [CrossRef] [PubMed]
- Gulick, A.M. Conformational Dynamics in the Acyl-CoA Synthetases, Adenylation domains of non-ribosomal peptide synthetases, and firefly luciferase. ACS Chembiol. 2009, 4, 811–827. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Gai, Y.; Yin, L.; Wang, X.; Feng, C.; Feng, L.; Li, D.; Jiang, X.; Wang, D. Crystal structures of a Populus tomentosa 4-Coumarate: CoA ligase shed light on its enzymatic mechanisms. Plant Cell 2010, 22, 3093–3104. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Nair, S.K. Structural basis for specificity and flexibility in a plant 4-Coumarate: CoA ligase. Structure 2015, 23, 2032–2042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cerenak, A.; Satovic, Z.; Javornik, B. Genetic mapping of hop (Humulus lupulus L.) applied to the detection of QTLs for alpha-acid content. Genome 2006, 49, 485–494. [Google Scholar] [CrossRef]
- Paniego, N.B.; Zuurbier, K.W.M.; Fung, S.Y.; van der Heijden, R.; Scheffer, J.J.C.; Verpoorte, R. Phlorisovalerophenone synthase, a novel polyketide synthase from hop (Humulus lupulus L.) cones. Eur. J. Biochem. 1999, 262, 612–616. [Google Scholar] [CrossRef] [PubMed]
- Okada, Y.; Yamazaki, Y.; Suh, D.; Sankawa, U.; Ito, K. Bifunctional Activities of Valerophenone Synthase in Hop (Humulus Lupulus L.). J. Am. Soc. Brew. Chem. 2001, 59, 163–166. [Google Scholar]
- Okada, Y.; Sano, Y.; Kaneko, T.; Abe, I.; Noguchi, H.; Ito, K. Enzymatic reactions by five chalcone synthase homologs from hop (Humulus lupulus L). Biosci. Biotechnol. Biochem. 2004, 68, 1142–1145. [Google Scholar] [CrossRef]
- Hagemann, M.H.; Born, U.; Weber, G.; Wunsche, J.N. Gene structure and allelic variations of hop (Humulus lupulus L.) bitter acid biosynthesis genes. Brew. Sci. 2019, 72, 207–216. [Google Scholar]
- Zhou, W.; Zhuang, Y.; Bai, Y.; Bi, H.; Liu, T.; Ma, Y. Biosynthesis of phlorisovalerophenone and 4-hydroxy-6-isobutyl-2-pyrone in Escherichia coli from glucose. Microb. Cell Fact. 2016, 15, 149. [Google Scholar] [CrossRef] [Green Version]
- Tsurumaru, Y.; Sasaki, K.; Miyawaki, T.; Momma, T.; Umemoto, N.; Yazaki, K. An aromatic prenyltransferase-like gene HlPT-1 preferentially expressed in lupulin glands of hop. Plant Biotechnol. 2010, 27, 199–204. [Google Scholar] [CrossRef] [Green Version]
- Tsurumaru, Y.; Sasaki, K.; Miyawaki, T.; Uto, Y.; Momma, T.; Umemoto, N.; Momose, M.; Yazaki, K. HlPT-1, a membrane-bound prenyltransferase responsible for the biosynthesis of bitter acids in hops. Biochem. Biophys. Res. Commun. 2012, 417, 393–398. [Google Scholar] [CrossRef]
- Ban, Z.; Qin, H.; Mitchell, A.J.; Liu, B.; Zhang, F.; Weng, J.; Dixon, R.A.; Wang, G. Noncatalytic chalcone isomerase-fold proteins in Humulus lupulus are auxiliary components in prenylated flavonoid biosynthesis. Proc. Natl. Acad. Sci. USA 2018, 115, E5223–E5232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Champagne, A.; Boutry, M. A comprehensive proteome map of glandular trichomes of hop (Humulus lupulus L.) female cones: Identification of biosynthetic pathways of the major terpenoid-related compounds and possible transport proteins. Protemics 2017, 17, 1600411. [Google Scholar] [CrossRef] [PubMed]
- Tholl, D. Biosynthesis and Biological Functions of Terpenoids in Plants. Adv. Biochem. Eng. Biotechnol. 2015, 148, 63–106. [Google Scholar]
- Lange, B.M. The Evolution of plant secretory structures and emergence of terpenoid chemical diversity. Annu. Rev. Plant Biol. 2015, 66, 139–159. [Google Scholar] [CrossRef]
- Booth, J.K.; Page, J.E.; Bohlmann, J. Terpene synthases from Cannabis sativa. PLoS ONE 2017, 12, e0173911. [Google Scholar]
- Dudareva, N.; Martin, D.; Kish, C.M.; Kolosova, N.; Gorenstein, N.; Faldt, J.; Miller, B.; Bohlmann, J. (E)-beta-ocimene and myrcene synthase genes of floral scent biosynthesis in snapdragon: Function and expression of three terpene synthase genes of a new terpene synthase subfamily. Plant Cell 2003, 15, 1227–1241. [Google Scholar] [CrossRef] [Green Version]
- Lois, L.M.; Rodriguez-Concepcion, M.; Gallego, F.; Campos, N.; Boronat, A. Carotenoid biosynthesis during tomato fruit development: Regulatory role of 1-deoxy-D-xylulose 5-phosphate synthase. Plant J. 2000, 22, 503–513. [Google Scholar] [CrossRef]
- Walter, M.H.; Fester, T.; Strack, D. Arbuscular mycorrhizal fungi induce the non-mevalonate methylerythritol phosphate pathway of isoprenoid biosynthesis correlated with accumulation of the ’yellow pigment’ and other apocarotenoids. Plant J. 2000, 21, 571–578. [Google Scholar] [CrossRef] [PubMed]
- Estevez, J.M.; Cantero, A.; Reindl, A.; Reichler, S.; Leon, P. 1-deoxy-D-xylulose-5-phosphate synthase, a limiting enzyme for plastidic isoprenoid biosynthesis in plants. J. Biol. Chem. 2001, 276, 22901–22909. [Google Scholar] [CrossRef] [Green Version]
- Walter, M.H.; Hans, J.; Strack, D. Two distantly related genes encoding 1-deoxy-D-xylulose 5-phosphate synthases: Differential regulation in shoots and apocarotenoid-accumulating mycorrhizal roots. Plant J. 2002, 31, 243–254. [Google Scholar] [CrossRef] [PubMed]
- Phillips, M.A.; Walter, M.H.; Ralph, S.G.; Dabrowska, P.; Luck, K.; Urós, E.M.; Boland, W.; Strack, D.; Rodríguez-Concepción, M.; Bohlmann, J.; et al. Functional identification and differential expression of 1-deoxy-d-xylulose 5-phosphate synthase in induced terpenoid resin formation of Norway spruce (Picea abies). Plant Mol. Biol. 2007, 65, 243–257. [Google Scholar] [CrossRef] [PubMed]
- Estevez, J.M.; Cantero, A.; Romero, C.; Kawaide, H.; Jimenez, L.F.; Kuzuyama, T.; Seto, H.; Kamiya, Y.; Leon, P. Analysis of the expression of CLA1, a gene that encodes the 1-deoxyxylulose 5-phosphate synthase of the 2-C-methyl-D-erythritol-4-phosphate pathway in Arabidopsis. Plant Physiol. 2000, 124, 95–103. [Google Scholar] [CrossRef] [Green Version]
- Tong, Y.; Su, P.; Zhao, Y.; Zhang, M.; Wang, X.; Liu, Y.; Zhang, X.; Gao, W.; Huang, L. Molecular cloning and characterization of DXS and DXR genes in the terpenoid biosynthetic pathway of Tripterygium wilfordii. Int. J. Mol. Sci. 2015, 16, 25516–25535. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Li, K.; Zhang, C.; Gai, J.; Yu, D. Identification and characterization of class 1 DXS gene encoding 1-deoxy-d-xylulose-5-phosphate synthase, the first committed enzyme of the MEP pathway from soybean. Mol. Biol. Rep. 2009, 36, 879–887. [Google Scholar] [CrossRef]
- Rohdich, F.; Hecht, S.; Gartner, K.; Adam, P.; Krieger, C.; Amslinger, S.; Arigoni, D.; Bacher, A.; Eisenreich, W. Studies on the nonmevalonate terpene biosynthetic pathway: Metabolic role of IspH (LytB) protein. Proc. Natl. Acad. Sci. USA 2002, 99, 1158–1163. [Google Scholar] [CrossRef] [Green Version]
- Rohdich, F.; Zepeck, F.; Adam, P.; Hecht, S.; Kaiser, J.; Laupitz, R.; Grawert, T.; Amslinger, S.; Eisenreich, W.; Bacher, A.; et al. The deoxyxylulose phosphate pathway of isoprenoid biosynthesis: Studies on the mechanisms of the reactions catalyzed by IspG and IspH protein. Proc. Natl. Acad. Sci. USA 2003, 100, 1586–1591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tritsch, D.; Hemmerlin, A.; Bach, T.J.; Rohmer, M. Plant isoprenoid biosynthesis via the MEP pathway: In vivo IPP/DMAPP ratio produced by (E)-4-hydroxy-3-methylbut-2-enyl diphosphate reductase in tobacco BY-2 cell cultures. FEBS Lett. 2010, 584, 129–134. [Google Scholar] [CrossRef] [Green Version]
- Phillips, M.A.; D’Auria, J.C.; Gershenzon, J.; Pichersky, E. The Arabidopsis thaliana type I isopentenyl diphosphate isomerases are targeted to multiple subcellular compartments and have overlapping functions in isoprenoid biosynthesis. Plant Cell 2008, 20, 677–696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, W.; Ma, T.; Liu, M.; Qu, J.; Liu, Z.; Zhang, H.; Shi, B.; Fu, S.; Ma, J.; Lai, L.T.F.; et al. Modular enzyme assembly for enhanced cascade biocatalysis and metabolic flux. Nat. Commun. 2019, 10, 4248. [Google Scholar] [CrossRef]
- Zhu, F.; Zhong, X.; Hu, M.; Lu, L.; Deng, Z.; Liu, T. In vitro reconstitution of mevalonate pathway and targeted engineering of farnesene overproduction in Escherichia coli. Biotechnol. Bioeng. 2014, 111, 1396–1405. [Google Scholar] [CrossRef]
- Albrecht, M.; Misawa, N.; Sandmann, G. Metabolic engineering of the terpenoid biosynthetic pathway of Escherichia coli for production of the carotenoids β-carotene and zeaxanthin. Biotechnol. Lett. 1999, 21, 791–795. [Google Scholar] [CrossRef]
- Kim, S.W.; Keasling, J.D. Metabolic engineering of the non-mevalonate isopentenyl diphosphate synthesis pathway in Escherichia coli enhances lycopene production. Biotechnol. Bioeng. 2001, 72, 408–415. [Google Scholar] [CrossRef]
- Leonard, E.; Ajikumar, P.K.; Thayer, K.; Xiao, W.H.; Mo, J.D.; Tidor, B.; Stephanopoulos, G.; Prather, K.L.J. Combining metabolic and protein engineering of a terpenoid biosynthetic pathway for overproduction and selectivity control. Proc. Natl. Acad. Sci. USA 2010, 107, 13654–13659. [Google Scholar] [CrossRef] [Green Version]
- Muñoz-Bertomeu, J.; Arrillaga, I.; Ros, R.; Segura, J. Up-Regulation of 1-Deoxy-d-Xylulose-5-Phosphate synthase enhances production of essential Oils in transgenic spike lavender. Plant Physiol. 2006, 142, 890–900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- You, M.K.; Lee, Y.J.; Kim, J.K.; Baek, S.A.; Jeon, Y.A.; Lim, S.H.; Ha, S.H. The organ-specific differential roles of rice DXS and DXR, the first two enzymes of the MEP pathway, in carotenoid metabolism in Oryza sativa leaves and seeds. BMC Plant Biol. 2020, 20, 167. [Google Scholar] [CrossRef] [Green Version]
- Martin, V.J.J.; Pitera, D.J.; Withers, S.T.; Newman, J.D.; Keasling, J.D. Engineering a mevalonate pathway in Escherichia coli for production of terpenoids. Nat. Biotechnol. 2003, 21, 796–802. [Google Scholar] [CrossRef]
- Tabata, K.; Hashimoto, S. Production of mevalonate by a metabolically-engineered Escherichia coli. Biotechnol. Lett. 2004, 26, 1487–1491. [Google Scholar] [CrossRef]
- Tsuruta, H.; Paddon, C.J.; Eng, D.; Lenihan, J.R.; Horning, T.; Anthony, L.C.; Regentin, R.; Keasling, J.D.; Renninger, N.S.; Newman, J.D. High-level production of Amorpha-4,11-Diene, a precursor of the antimalarial agent artemisinin, in Escherichia coli. PLoS ONE 2009, 4, e4489. [Google Scholar] [CrossRef]
- Yoon, S.; Lee, S.; Das, A.; Ryu, H.; Jang, H.; Kim, J.; Oh, D.; Keasling, J.D.; Kim, S. Combinatorial expression of bacterial whole mevalonate pathway for the production of β-carotene in E. coli. J. Biotechnol. 2009, 140, 218–226. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Yang, J.; Qin, B.; Li, Y.; Sun, Y.; Su, S.; Xian, M. Biosynthesis of isoprene in Escherichia coli via methylerythritol phosphate (MEP) pathway. Appl. Microbiol. Biot. 2011, 90, 1915–1922. [Google Scholar] [CrossRef] [PubMed]
- Kirby, J.; Dietzel, K.L.; Wichmann, G.; Chan, R.; Antipov, E.; Moss, N.; Baidoo, E.E.K.; Jackson, P.; Gaucher, S.P.; Gottlieb, S.; et al. Engineering a functional 1-deoxy-D-xylulose 5-phosphate (DXP) pathway in Saccharomyces cerevisiae. Metab. Eng. 2016, 38, 494–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, G.; Dixon, R.A. Heterodimeric geranyl (geranyl)diphosphate synthase from hop (Humulus lupulus) and the evolution of monoterpene biosynthesis. Proc. Natl. Acad. Sci. USA 2008, 106, 9914–9919. [Google Scholar] [CrossRef] [Green Version]
- Burke, C.C.; Wildung, M.R.; Croteau, R. Geranyl diphosphate synthase: Cloning, expression, and characterization of this prenyltransferase as a heterodimer. Proc. Natl. Acad. Sci. USA 1999, 96, 13062–13067. [Google Scholar] [CrossRef] [Green Version]
- Tholl, D.; Kish, C.M.; Orlova, I.; Sherman, D.; Gershenzon, J.; Pichersky, E.; Dudareva, N. Formation of monoterpenes in Antirrhinum majus and Clarkia breweri flowers involves heterodimeric geranyl diphosphate synthases. Plant Cell 2004, 16, 977–992. [Google Scholar] [CrossRef] [Green Version]
- Chang, T.; Hsieh, F.; Ko, T.; Teng, K.; Liang, P.; Wang, A.H.J. Structure of a heterotetrameric geranyl pyrophosphate synthase from Mint (Mentha piperita) reveals inter subunit regulation. Plant Cell 2010, 22, 454–467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denby, C.M.; Li, R.A.; Vu, V.T.; Costello, Z.; Lin, W.; Chan, L.J.G.; Williams, J.; Donaldson, B.; Bamforth, C.W.; Petzold, C.J.; et al. Industrial brewing yeast engineered for the production of primary flavor determinants in hopped beer. Nat. Commun. 2018, 9, 965. [Google Scholar] [CrossRef]
- Schoch, G.A.; Attias, R.; Belghazi, M.; Dansette, P.M.; Werck-Reichhart, D. Engineering of a water-soluble plant cytochrome P450, CYP73A1, and NMR-Based orientation of natural and alternate substrates in the active site. Plant Physiol. 2003, 133, 1198–1208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.; Deavours, B.E.; Richard, S.B.; Ferrer, J.; Blount, J.W.; Huhman, D.; Dixon, R.A.; Noel, J.P. Structural basis for dual functionality of isoflavonoid O-methyltransferases in the evolution of plant defense responses. Plant Cell 2007, 18, 3656–3669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wen, L.; Jiang, Y.; Yang, J.; Zhao, Y.; Tian, M.; Yang, B. Structure, bioactivity, and synthesis of methylated flavonoids. Ann. N. Y. Acad. Sci. 2017, 1398, 120–129. [Google Scholar] [CrossRef]
- Martin, D.M.; Bohlmann, J. Identification of Vitis vinifera (-)-alpha-terpineol synthase by in silico screening of full-length cDNA ESTs and functional characterization of recombinant terpene synthase. Phytochemistry 2004, 65, 1223–1229. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Wei, L.; Lin, J.; Hua, Q. Enhancing linalool production by engineering oleaginous yeast Yarrowia lipolytica. Bioresour. Technol. 2017, 245, 1641–1644. [Google Scholar] [CrossRef]
- Eriksen, R.L.; Padgitt-Cobb, L.K.; Townsend, M.S.; Henning, J.A. Gene expression for secondary metabolite biosynthesis in hop (Humulus lupulus L.) leaf lupulin glands exposed to heat and low-water stress. Sci. Rep. 2021, 11, 5138. [Google Scholar] [CrossRef]
- De Keukeleire, J.; Ooms, G.; Heyerick, A.; Roldan-Ruiz, I.; Van Bockstaele, E.; De Keukeleire, D. Formation and accumulation of α-Acids, β-Acids, desmethylxanthohumol, and xanthohumol during flowering of hops (Humulus lupulus L.). J. Agric. Food Chem. 2003, 51, 4436–4441. [Google Scholar] [CrossRef]
- Stevens, J.F.; Taylor, A.W.; Clawson, J.E.; Deinzer, M.L. Fate of xanthohumol and related prenylflavonoids from hops to beer. J. Agric. Food Chem. 1999, 47, 2421–2428. [Google Scholar] [CrossRef] [PubMed]
- Hansel, R.; Schulz, J. Desmethylxanthohumol: Isolierung aus hopfen und Cyclisierung zu flavanonen. Arch. Pharm. 2010, 321, 37–40. [Google Scholar] [CrossRef]
- Chadwick, L.R.; Pauli, G.F.; Farnsworth, N.R. The pharmacognosy of Humulus lupulus L. (hops) with an emphasis on estrogenic properties. Phytomedicine 2006, 13, 119–131. [Google Scholar] [CrossRef] [Green Version]
- Milligan, S.R.; Kalita, J.C.; Heyerick, A.; Rong, H.; De Cooman, L.; De Keukeleire, D. Identification of a potent phytoestrogen in hops (Humulus lupulus L.) and beer. J. Clin. Endocr. Metab. 1999, 84, 2249–2252. [Google Scholar] [CrossRef] [PubMed]
- Sirotkin, A.V.; Harrath, A.H. Phytoestrogens and their effects. Eur. J. Pharmacol. 2014, 741, 230–236. [Google Scholar] [CrossRef] [PubMed]
- Abdi, F.; Rahnemaei, F.A.; Roozbeh, N.; Pakzad, R. Impact of phytoestrogens on treatment of urogenital menopause symptoms: A systematic review of randomized clinical trials. Eur. J. Obstet. Gynecol. Reprod. Biol. 2021, 261, 222–235. [Google Scholar] [CrossRef] [PubMed]
- Stevens, J.F.; Page, J.E. Xanthohumol and related prenylflavonoids from hops and beer: To your good health! Phytochemistry 2004, 65, 1317–1330. [Google Scholar] [CrossRef] [PubMed]
- Novák, P.; Matouek, J.; Bíza, J. Valerophenone synthase-like chalcone synthase homologues in Humulus Lupulus. Biol. Plantarm. 2003, 46, 375–381. [Google Scholar] [CrossRef]
- Ngaki, M.N.; Louie, G.V.; Philippe, R.N.; Manning, G.; Pojer, F.; Bowman, M.E.; Li, L.; Larsen, E.; Wurtele, E.S.; Noel, J.P. Evolution of the chalcone-isomerase fold from fatty-acid binding to stereospecific catalysis. Nature 2012, 485, 530–533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Werck-Reichhart, D.; Feyereisen, R. Cytochromes P450: A success story. Genome Biol. 2000, 1, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Dixon, R.A. Natural products and plant disease resistance. Nature 2001, 411, 843–847. [Google Scholar] [CrossRef]
- Blount, J.W.; Korth, K.L.; Masoud, S.A.; Rasmussen, S.; Lamb, C.; Dixon, R.A. Altering expression of cinnamic acid 4-hydroxylase in transgenic plants provides evidence for a feedback loop at the entry point into the phenylpropanoid pathway1. Plant Physiol. 2000, 122, 107–116. [Google Scholar] [CrossRef]
- Acker, R.V.; Vanholme, R.; Storme, V. Lignin biosynthesis perturbations affect secondary cell wall composition and saccharification yield in Arabidopsis thaliana. Biotechnol. Biofuels 2013, 6, 1–17. [Google Scholar]
- Sewalt, V.J.H.; Ni, W.; Blount, J.W.; Jung, H.C.; Masoud, S.A.; Howles, P.A.; Lamb, C.; Dixon, R.A. Reduced lignin content and altered lignin composition in transgenic tobacco down-regulated in expression of L-phenylalanine ammonia-lyase or cinnamate 4-hydroxylase. Plant Physiol. 1997, 115, 41–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, B.; Lewis, K.M.; Abril, A.; Davydov, D.R.; Vermerris, W.; Sattler, S.E.; Kang, C. Structure and function of the cytochrome P450 monooxygenase cinnamate 4-hydroxylase from sorghum bicolor. Plant Physiol. 2020, 183, 957–973. [Google Scholar] [CrossRef] [PubMed]
- Matoušek, J.; Novák, P.; Bříza, J.; Patzak, J.; Niedermeierová, H. Cloning and characterisation of chs-specific DNA and cDNA sequences from hop (Humulus lupulus L.). Plant Sci. 2002, 162, 1007–1018. [Google Scholar] [CrossRef]
- Nagel, J.; Culley, L.K.; Lu, Y.; Liu, E.; Matthews, P.D.; Stevens, J.F.; Page, J.E. EST Analysis of hop glandular trichomes identifies an O-methyltransferase that catalyzes the biosynthesis of xanthohumol. Plant Cell 2008, 20, 186–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morita, Y.; Takagi, K.; Fukuchi-Mizutani, M.; Ishiguro, K.; Tanaka, Y.; Nitasaka, E.; Nakayama, M.; Saito, N.; Kagami, T.; Hoshino, A.; et al. A chalcone isomerase-like protein enhances flavonoid production and flower pigmentation. Plant J. 2014, 78, 294–304. [Google Scholar] [CrossRef]
- Balasubramani, S.; Lv, S.; Chen, Q.; Zhou, Z.; Moorthy, M.D.S.; Sathish, D.; Moola, A.K. A systematic review of the O-methyltransferase gene expression. Plant Gene 2021, 27, 100295. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hong, K.; Wang, L.; Johnpaul, A.; Lv, C.; Ma, C. Key Enzymes Involved in the Synthesis of Hops Phytochemical Compounds: From Structure, Functions to Applications. Int. J. Mol. Sci. 2021, 22, 9373. https://doi.org/10.3390/ijms22179373
Hong K, Wang L, Johnpaul A, Lv C, Ma C. Key Enzymes Involved in the Synthesis of Hops Phytochemical Compounds: From Structure, Functions to Applications. International Journal of Molecular Sciences. 2021; 22(17):9373. https://doi.org/10.3390/ijms22179373
Chicago/Turabian StyleHong, Kai, Limin Wang, Agbaka Johnpaul, Chenyan Lv, and Changwei Ma. 2021. "Key Enzymes Involved in the Synthesis of Hops Phytochemical Compounds: From Structure, Functions to Applications" International Journal of Molecular Sciences 22, no. 17: 9373. https://doi.org/10.3390/ijms22179373