
Brain Metabolic Profile after Intranasal vs. Intraperitoneal Clomipramine Treatment in Rats with Ultrasound Model of Depression

 , ,
, ,
Abstract
:1. Introduction
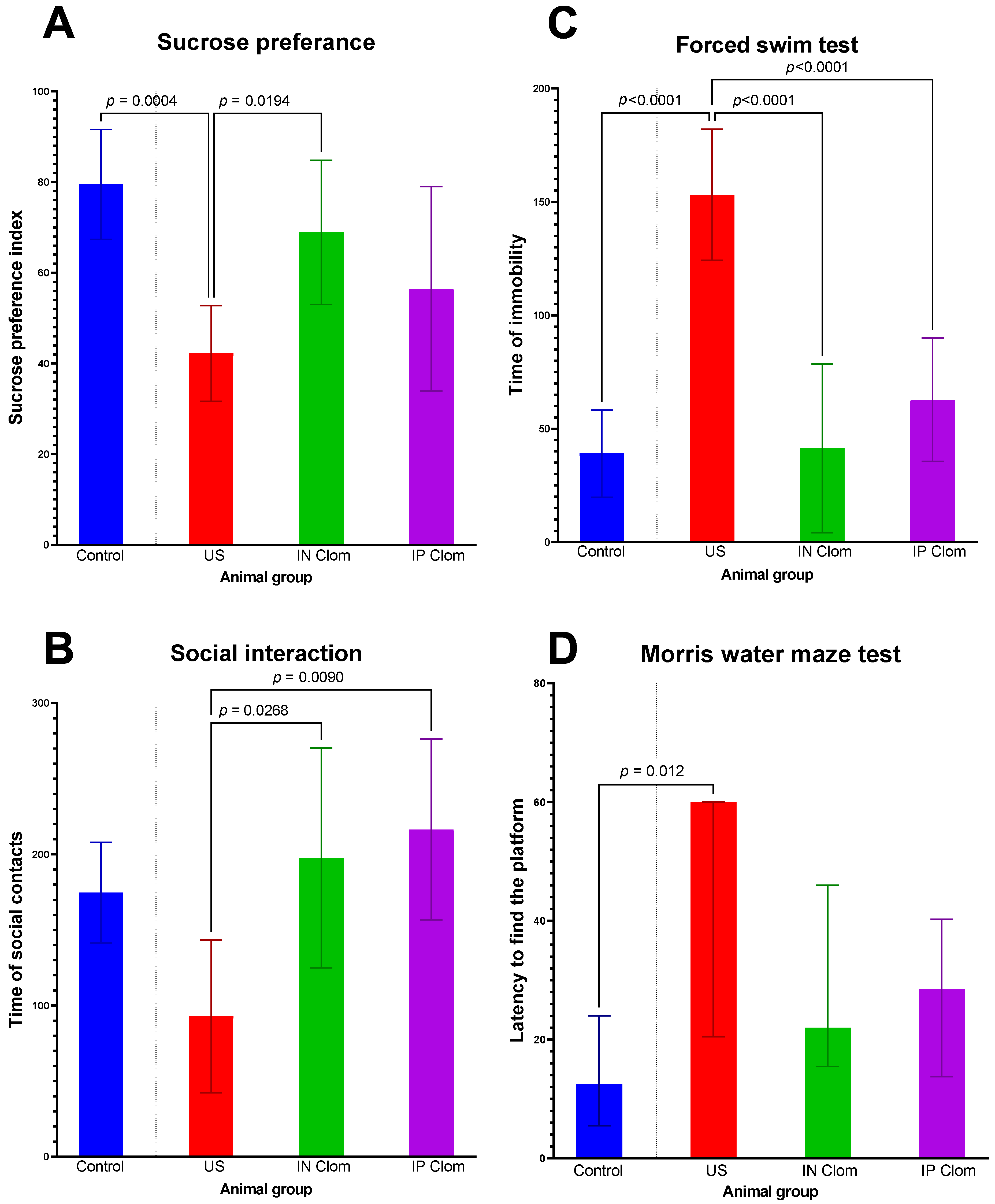
- We accessed the depressive-like animal state and also their cognitive functions after regular exposure to the variable frequency US waves in groups that got no treatment, and those that got treatment by clomipramine applied intranasally and intraperitoneally.
- We analyzed the amount and composition of 28 metabolites in rats’ frontal cortex (FC) and hippocampus in a normal and depressed-like state after treatment and with no treatment at all.
- We studied metabolic pathway changes related to depressive-like state and tried to find the metabolic pathway which could made the greatest contribution to metabolomic changes in depression.
- We made a comparative analysis of metabolic pathway changes depending on intranasal and intraperitoneal routes of drug administration.
2. Results
2.1. Behaviour Tests
2.2. Metabolomic Analysis in Brain
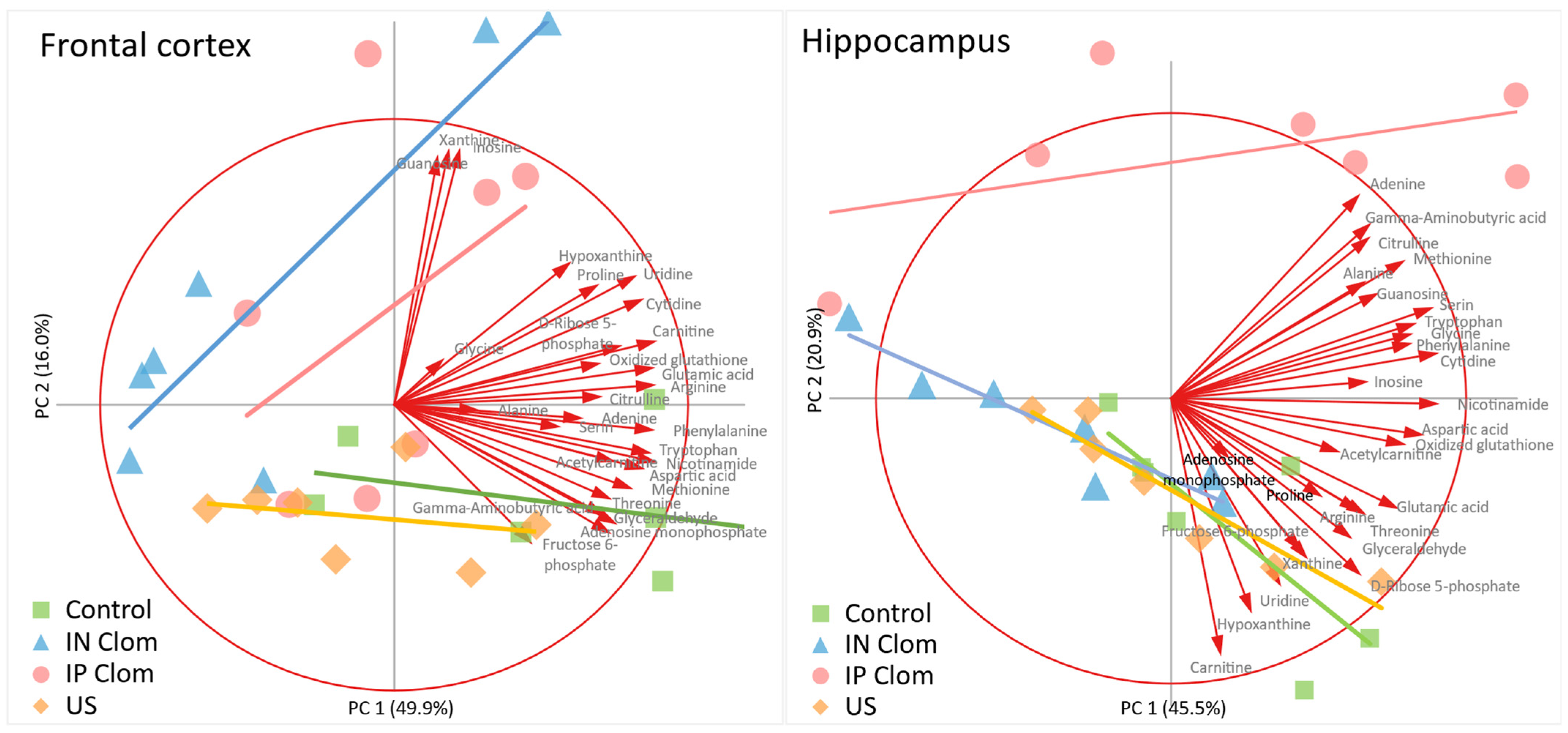
2.3. Principal Component Analysis (PCA)
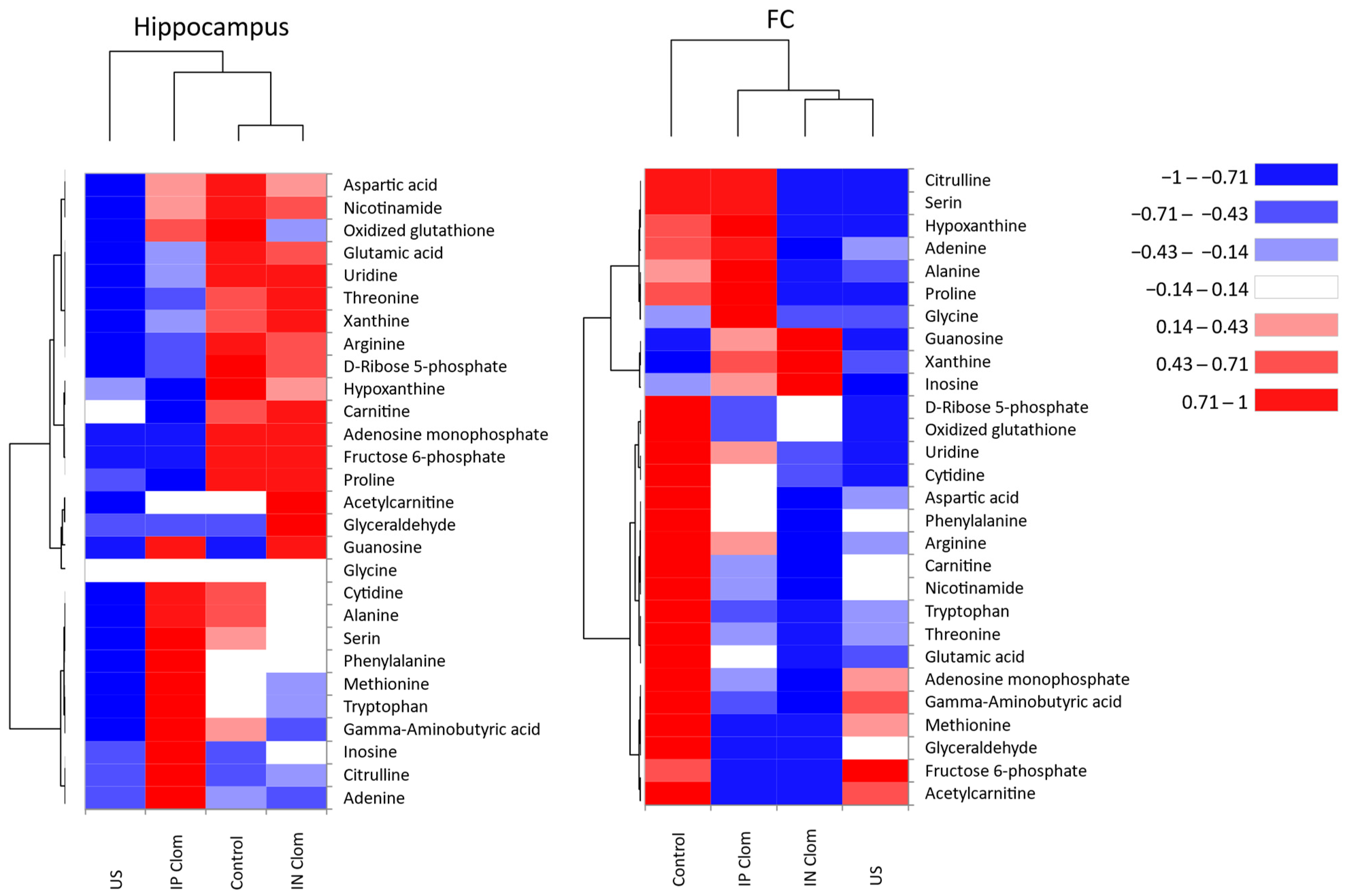
2.4. Heatmap Visualization
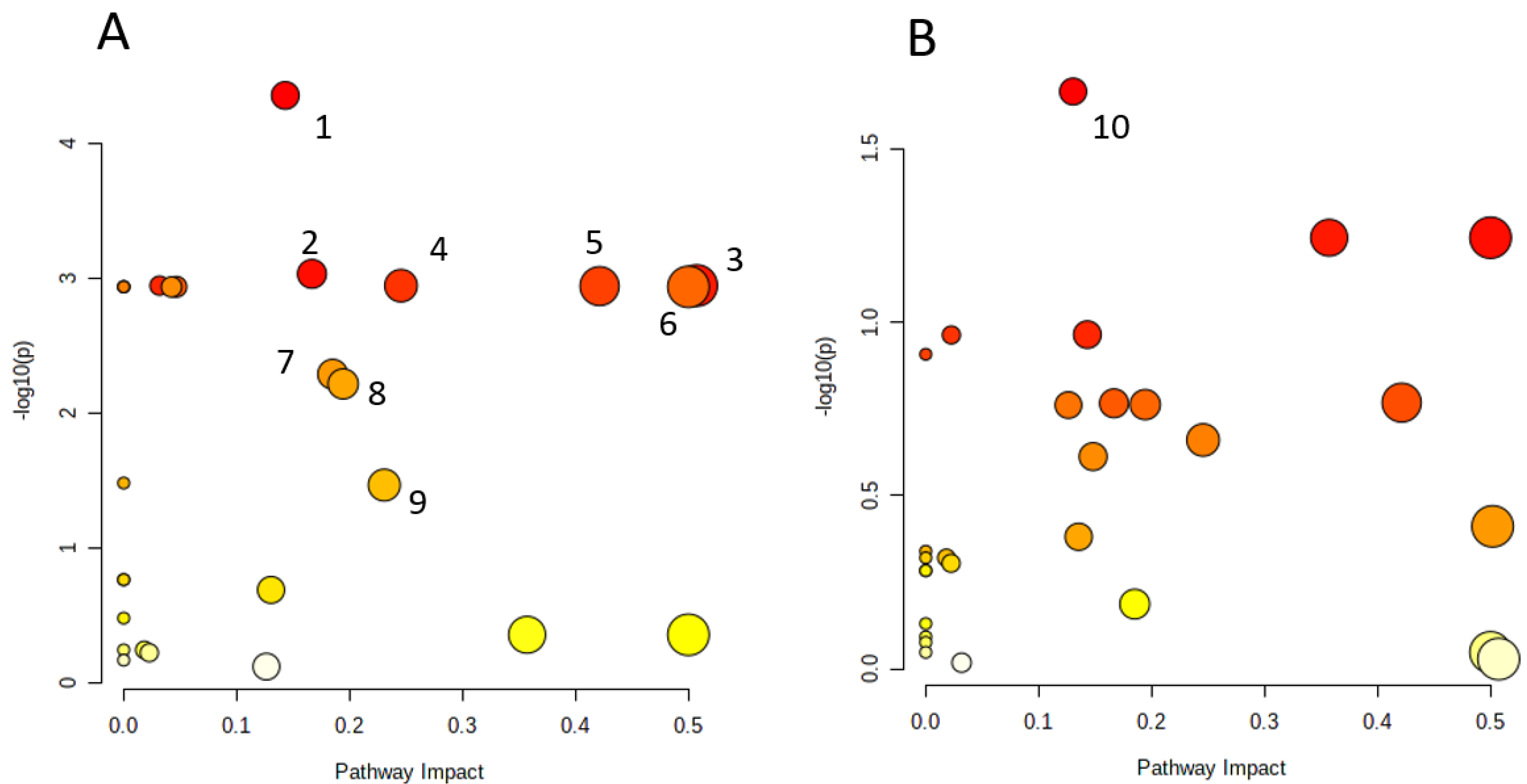
2.5. Metabolic Pathways Analysis
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Experimental Groups and Procedures
4.3. Behaviour Test
4.3.1. Social Interaction Test
4.3.2. Sucrose Preference Test
4.3.3. Forced Swim Test
4.3.4. Morris Water Maze Test
4.4. Tissue Sample Collection and Pretreatment
4.5. Sample Analysis
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviation
| CNS | Central nervous system |
| GABA | γ-aminobutyric acid |
| IN | Intranasal |
| IP | Intraperitoneal |
| MRM | Multiple-reaction monitoring |
| NAM | Nicotinamide |
| PCA | Principal component analysis |
| FC | Frontal cortex |
| SAM | S-Adenosyl methionine |
| US | Ultrasound |
References
- Malhi, G.S.; Mann, J.J. Depression. Lancet 2018, 392, 2299–2312. [Google Scholar] [CrossRef]
- Kessler, R.C.; Bromet, E.J. The Epidemiology of Depression Across Cultures. Annu. Rev. Public Health 2013, 34, 119–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pariante, C.M.; Lightman, S.L. The HPA axis in major depression: Classical theories and new developments. Trends Neurosci. 2008, 31, 464–468. [Google Scholar] [CrossRef] [PubMed]
- Willner, P. The chronic mild stress (CMS) model of depression: History, evaluation and usage. Neurobiol. Stress 2017, 6, 78–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, P.; Wang, K.; Lu, C.; Dong, L.; Chen, Y.; Wang, Q.; Shi, Z.; Yang, Y.; Chen, S.; Liu, X. Effects of the chronic restraint stress induced depression on reward-related learning in rats. Behav. Brain Res. 2017, 321, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Palma, B.D.; Suchecki, D.; Tufik, S. Differential effects of acute cold and footshock on the sleep of rats. Brain Res. 2000, 861, 97–104. [Google Scholar] [CrossRef]
- Morales-Medina, J.; Iannitti, T.; Freeman, A.; Caldwell, H. The olfactory bulbectomized rat as a model of depression: The hippocampal pathway. Behav. Brain Res. 2017, 317, 562–575. [Google Scholar] [CrossRef]
- Becker, M.; Pinhasov, A.; Ornoy, A. Animal Models of Depression: What Can They Teach Us about the Human Disease? Diagnostics 2021, 11, 123. [Google Scholar] [CrossRef]
- Yang, L.; Zhao, Y.; Wang, Y.; Liu, L.; Zhang, X.; Li, B.; Cui, R. The Effects of Psychological Stress on Depression. Curr. Neuropharmacol. 2015, 13, 494–504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miragaia, A.S.; Wertheimer, G.S.D.O.; Consoli, A.C.; Cabbia, R.; Longo, B.M.; Girardi, C.E.N.; Suchecki, D. Maternal Deprivation Increases Anxiety- and Depressive-Like Behaviors in an Age-Dependent Fashion and Reduces Neuropeptide Y Expression in the Amygdala and Hippocampus of Male and Female Young Adult Rats. Front. Behav. Neurosci. 2018, 12, 159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farbstein, D.; Hollander, N.; Peled, O.; Apter, A.; Fennig, S.; Haberman, Y.; Gitman, H.; Yaniv, I.; Shkalim, V.; Pick, C.G.; et al. Social isolation in mice: Behavior, immunity, and tumor growth. Stress 2021, 24, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.Y.; Zhou, X.Y.; Yang, L.N.; Wang, H.Y.; Zhang, Y.Q.; Pu, J.C.; Liu, L.X.; Gui, S.W.; Zeng, L.; Chen, J.J.; et al. Social defeat stress causes depression-like behavior with metabolite changes in the frontal cortex of rats. PLoS ONE 2017, 12, e0176725. [Google Scholar] [CrossRef] [Green Version]
- Morozova, A.; Zubkov, E.; Strekalova, T.; Кекелидзе, З.; Storozeva, Z.; Schroeter, C.A.; Bazhenova, N.; Lesch, K.-P.; Cline, B.H.; Chekhonin, V. Ultrasound of alternating frequencies and variable emotional impact evokes depressive syndrome in mice and rats. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2016, 68, 52–63. [Google Scholar] [CrossRef] [PubMed]
- Zorkina, Y.A.; Zubkov, E.; Morozova, A.Y.; Ushakova, V.; Chekhonin, V.P. The Comparison of a New Ultrasound-Induced Depression Model to the Chronic Mild Stress Paradigm. Front. Behav. Neurosci. 2019, 13, 146. [Google Scholar] [CrossRef]
- Xu, H.-B.; Fang, L.; Hu, Z.-C.; Chen, Y.-C.; Chen, J.-J.; Li, F.-F.; Lu, J.; Mu, J.; Xie, P. Potential clinical utility of plasma amino acid profiling in the detection of major depressive disorder. Psychiatry Res. 2012, 200, 1054–1057. [Google Scholar] [CrossRef]
- Steffens, D.C.; Krishnan, K.R.R.; Karoly, E.D.; Mitchell, M.W.; O’Connor, C.M.; Kaddurah-Daouk, R.; Jiang, W. Metabolomic Differences in Heart Failure Patients With and Without Major Depression. J. Geriatr. Psychiatry Neurol. 2010, 23, 138–146. [Google Scholar] [CrossRef]
- Liu, L.; Zhou, X.; Zhang, Y.; Liu, Y.; Yang, L.; Pu, J.; Zhu, D.; Zhou, C.; Xie, P. The identification of metabolic disturbances in the prefrontal cortex of the chronic restraint stress rat model of depression. Behav. Brain Res. 2016, 305, 148–156. [Google Scholar] [CrossRef]
- Chen, J.-J.; Bai, S.-J.; Li, W.-W.; Zhou, C.-J.; Zheng, P.; Fang, L.; Wang, H.-Y.; Liu, Y.-Y.; Xie, P. Urinary biomarker panel for diagnosing patients with depression and anxiety disorders. Transl. Psychiatry 2018, 8, 192. [Google Scholar] [CrossRef] [Green Version]
- Macdonald, K.; Krishnan, A.; Cervenka, E.; Hu, G.; Guadagno, E.; Trakadis, Y. Biomarkers for major depressive and bipolar disorders using metabolomics: A systematic review. Am. J. Med Genet. Part B: Neuropsychiatr. Genet. 2019, 180, 122–137. [Google Scholar] [CrossRef] [PubMed]
- Nobis, A.; Zalewski, D.; Waszkiewicz, N. Peripheral Markers of Depression. J. Clin. Med. 2020, 9, 3793. [Google Scholar] [CrossRef]
- Bhattacharyya, S.; Ahmed, A.T.; Arnold, M.; Liu, D.; Luo, C.; Zhu, H.; Mahmoudiandehkordi, S.; Neavin, D.; Louie, G.; Dunlop, B.W.; et al. Metabolomic signature of exposure and response to citalopram/escitalopram in depressed outpatients. Transl. Psychiatry 2019, 9, 173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaddurah-Daouk, R.; Boyle, S.H.; Matson, W.; Sharma, S.; Matson, S.; Zhu, H.; Bogdanov, M.B.; Churchill, E.; Krishnan, R.R.; Rush, A.; et al. Pretreatment metabotype as a predictor of response to sertraline or placebo in depressed outpatients: A proof of concept. Transl. Psychiatry 2011, 1, e26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cipriani, A.; Furukawa, T.; Salanti, G.; Chaimani, A.; Atkinson, L.; Ogawa, Y.; Leucht, S.; Ruhe, H.G.; Turner, E.H.; Higgins, J.; et al. Comparative efficacy and acceptability of 21 antidepressant drugs for the acute treatment of adults with major depressive disorder: A systematic review and network meta-analysis. Lancet 2018, 391, 1357–1366. [Google Scholar] [CrossRef] [Green Version]
- Hamilton, P.J.; Chen, E.Y.; Tolstikov, V.; Peña, C.J.; Picone, J.A.; Shah, P.; Panagopoulos, K.; Strat, A.N.; Walker, D.M.; Lorsch, Z.S.; et al. Chronic stress and antidepressant treatment alter purine metabolism and beta oxidation within mouse brain and serum. Sci. Rep. 2020, 10, 18134. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yuan, S.; Pu, J.; Yang, L.; Zhou, X.; Liu, L.; Jiang, X.; Zhang, H.; Teng, T.; Tian, L.; et al. Integrated Metabolomics and Proteomics Analysis of Hippocampus in a Rat Model of Depression. Neuroscience 2018, 371, 207–220. [Google Scholar] [CrossRef]
- Tian, L.; Pu, J.; Liu, Y.; Gui, S.; Zhong, X.; Song, X.; Xu, S.; Zhang, H.; Wang, H.; Zhou, W.; et al. Metabolomic analysis of animal models of depression. Metab. Brain Dis. 2020, 35, 979–990. [Google Scholar] [CrossRef]
- Seo, J.-S.; Wei, J.; Qin, L.; Kim, Y.; Yan, Z.; Greengard, P. Cellular and molecular basis for stress-induced depression. Mol. Psychiatry 2020, 22, 1440–1447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kraus, C.; Castrén, E.; Kasper, S.; Lanzenberger, R. Serotonin and neuroplasticity—Links between molecular, functional and structural pathophysiology in depression. Neurosci. Biobehav. Rev. 2017, 77, 317–326. [Google Scholar] [CrossRef] [Green Version]
- Martín-Hernández, D.; Tendilla-Beltrán, H.; Madrigal, J.; García-Bueno, B.; Leza, J.C.; Caso, J.R. Chronic Mild Stress Alters Kynurenine Pathways Changing the Glutamate Neurotransmission in Frontal Cortex of Rats. Mol. Neurobiol. 2019, 56, 490–501. [Google Scholar] [CrossRef]
- Eskelund, A.; Li, Y.; Budac, D.P.; Müller, H.K.; Gulinello, M.; Sanchez, C.; Wegener, G. Drugs with antidepressant properties affect tryptophan metabolites differently in rodent models with depression-like behavior. J. Neurochem. 2017, 142, 118–131. [Google Scholar] [CrossRef]
- Kurbat, M.N. L-glutamate: A modern view on a well-known amino acid. Neurochem. J. 2009, 3, 173–178. [Google Scholar] [CrossRef]
- Zink, M.; Vollmayr, B.; Gebicke-Haerter, P.J.; Henn, F.A. Reduced expression of glutamate transporters vGluT1, EAAT2 and EAAT4 in learned helpless rats, an animal model of depression. Neuropharmacology 2010, 58, 465–473. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.P.; Wang, C.; Xu, J.P. Chronic unpredictable mild stress induced depression-like behaviours and glutamate-glutamine cycling dysfunctions in both blood and brain of mice. Pharm. Biol. 2019, 57, 280–286. [Google Scholar] [CrossRef]
- Moriguchi, S.; Takamiya, A.; Noda, Y.; Horita, N.; Wada, M.; Tsugawa, S.; Plitman, E.; Sano, Y.; Tarumi, R.; ElSalhy, M.; et al. Glutamatergic neurometabolite levels in major depressive disorder: A systematic review and meta-analysis of proton magnetic resonance spectroscopy studies. Mol. Psychiatry 2019, 24, 952–964. [Google Scholar] [CrossRef] [Green Version]
- Son, H.; Baek, J.H.; Go, B.S.; Jung, D.H.; Sontakke, S.B.; Chung, H.J.; Lee, D.H.; Roh, G.S.; Kang, S.S.; Cho, G.J.; et al. Glutamine has antidepressive effects through increments of glutamate and glutamine levels and glutamatergic activity in the medial frontal cortex. Neuropharmacology 2018, 143, 143–152. [Google Scholar] [CrossRef]
- Yoon, S.J.; Lyoo, I.K.; Haws, C.; Kim, T.S.; Cohen, B.M.; Renshaw, P.F. Decreased glutamate/glutamine levels may mediate cytidine’s efficacy in treating bipolar depression: A longitudinal proton magnetic resonance spectroscopy study. Neuropsychopharmacology 2009, 34, 1810–1818. [Google Scholar] [CrossRef] [Green Version]
- Henter, I.D.; de Sousa, R.T.; Zarate, C.A., Jr. Glutamatergic Modulators in Depression. Harv. Rev. Psychiatry 2018, 26, 307–319. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; He, W.; Huang, Y.; Zeng, Z.; Yang, X.; Huang, H.; Wen, J.; Cao, Y.; Sun, H. Hippocampal metabolic alteration in rat exhibited susceptibility to prenatal stress. J. Affect. Disord. 2019, 259, 458–467. [Google Scholar] [CrossRef]
- Akimoto, H.; Oshima, S.; Ohara, K.; Negishi, A.; Hiroyama, H.; Nemoto, T.; Kobayashi, D. High-Resolution Magic-Angle Spinning-1H-NMR Spectroscopy-Based Metabolic Profiling of Hippocampal Tissue in Rats with Depression-Like Symptoms. Biol. Pharm. Bull. 2017, 40, 789–796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Tang, G.; Cheng, K.; Yang, D.; Chen, G.; Liu, Z.; Zhang, R.; Zhou, J.; Fang, L.; Fang, Z.; et al. Peripheral blood mononuclear cell-based metabolomic profiling of a chronic unpredictable mild stress rat model of depression. Mol. BioSyst. 2014, 10, 2994–3001. [Google Scholar] [CrossRef]
- Lener, M.S.; Niciu, M.J.; Ballard, E.D.; Park, M.; Park, L.T.; Nugent, A.C.; Zarate, C.A., Jr. Glutamate and Gamma-Aminobutyric Acid Systems in the Pathophysiology of Major Depression and Antidepressant Response to Ketamine. Biol. Psychiatry 2017, 81, 886–897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duman, R.S.; Sanacora, G.; Krystal, J.H. Altered Connectivity in Depression: GABA and Glutamate Neurotransmitter Deficits and Reversal by Novel Treatments. Neuron 2019, 102, 75–90. [Google Scholar] [CrossRef]
- Luscher, B.; Shen, Q.; Sahir, N. The GABAergic deficit hypothesis of major depressive disorder. Mol. Psychiatry 2011, 16, 383–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dean, B.; Thomas, N.; Scarr, E.; Udawela, M. Evidence for impaired glucose metabolism in the striatum, obtained postmortem, from some subjects with schizophrenia. Transl. Psychiatry 2016, 6, e949. [Google Scholar] [CrossRef] [Green Version]
- Özaslan, M.S.; Balcı, N.; Demir, Y.; Gürbüz, M.; Küfrevioğlu, Ö.İ. Inhibition effects of some antidepressant drugs on pentose phosphate pathway enzymes. Environ. Toxicol. Pharmacol. 2019, 72, 103244. [Google Scholar] [CrossRef]
- Hwang, E.S.; Song, S.B. Possible Adverse Effects of High-Dose Nicotinamide: Mechanisms and Safety Assessment. Biomolecules 2020, 10, 687. [Google Scholar] [CrossRef]
- Liu, Z.; Li, C.; Fan, X.; Kuang, Y.; Zhang, X.; Chen, L.; Song, J.; Zhou, Y.; Takahashi, E.; He, G.; et al. Nicotinamide, a vitamin B3 ameliorates depressive behaviors independent of SIRT1 activity in mice. Mol. Brain 2020, 13, 162. [Google Scholar] [CrossRef] [PubMed]
- Fricker, R.A.; Green, E.L.; Jenkins, S.I.; Griffin, S.M. The Influence of Nicotinamide on Health and Disease in the Central Nervous System. Int. J. Tryptophan. Res. 2018, 11. [Google Scholar] [CrossRef] [Green Version]
- Aman, Y.; Qiu, Y.; Tao, J.; Fang, E.F. Therapeutic potential of boosting nad+ in aging and age-related diseases. Transl. Med. Aging 2018, 2, 30–37. [Google Scholar] [CrossRef]
- Gasperi, V.; Sibilano, M.; Savini, I.; Catani, M.V. Niacin in the Central Nervous System: An Update of Biological Aspects and Clinical Applications. Int. J. Mol. Sci. 2019, 20, 974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez, Y.; Li, X.; Liu, G.; Bin, P.; Yan, W.; Más, D.; Valdivié, M.; Hu, C.A.; Ren, W.; Yin, Y. The role of methionine on metabolism, oxidative stress, and diseases. Amino Acids 2017, 49, 2091–2098. [Google Scholar] [CrossRef] [PubMed]
- Bottiglieri, T. S-Adenosyl-L-methionine (SAMe): From the bench to the bedside--molecular basis of a pleiotrophic molecule. Am. J. Clin. Nutr. 2002, 76, 1151S–1157S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galizia, I.; Oldani, L.; Macritchie, K.; Amari, E.; Dougall, D.; Jones, T.N.; Lam, R.W.; Massei, G.J.; Yatham, L.N.; Young, A.H. S-adenosyl methionine (SAMe) for depression in adults. Cochrane Database Syst. Rev. 2016, 10, CD011286. [Google Scholar] [CrossRef] [Green Version]
- Karas Kuželički, N. S-Adenosyl Methionine in the Therapy of Depression and Other Psychiatric Disorders. Drug Dev. Res. 2016, 77, 346–356. [Google Scholar] [CrossRef]
- Ibba, M.; Soll, D. Aminoacyl-tRNA synthesis. Annu. Rev. Biochem. 2000, 69, 617–650. [Google Scholar] [CrossRef] [PubMed]
- Nie, A.; Sun, B.; Fu, Z.; Yu, D. Roles of aminoacyl-tRNA synthetases in immune regulation and immune diseases. Cell Death Dis. 2019, 10, 901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Yan, B.; Zhao, B.; Fan, Y.; He, X.; Yang, L.; Ma, Q.; Zheng, J.; Wang, W.; Bai, L.; et al. Assessing the Causal Effects of Human Serum Metabolites on 5 Major Psychiatric Disorders. Schizophr. Bull. 2020, 46, 804–813. [Google Scholar] [CrossRef] [PubMed]
- Cao, B.; Deng, R.; Wang, D.; Li, L.; Ren, Z.; Xu, L.; Gao, X. Association between arginine catabolism and major depressive disorder: A protocol for the systematic review and meta-analysis of metabolic pathway. Medicine 2020, 99, e21068. [Google Scholar] [CrossRef]
- Guelzim, N.; Mariotti, F.; Martin, P.G.; Lasserre, F.; Pineau, T.; Hermier, D. A role for PPARα in the regulation of arginine metabolism and nitric oxide synthesis. Amino Acids 2011, 41, 969–979. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.R.; Zhang, Y.; Rao, Y.B.; Chen, X.; Lou, H.F.; Zhang, Y.; Xie, H.Y.; Fang, P.; Hu, L.W. The changes in, and relationship between, plasma nitric oxide and corticotropin-releasing hormone in patients with major depressive disorder. Clin. Exp. Pharmacol. Physiol. 2018, 45, 10–15. [Google Scholar] [CrossRef]
- Jesse, C.R.; Bortolatto, C.F.; Savegnago, L.; Rocha, J.B.; Nogueira, C.W. Involvement of L-arginine-nitric oxide-cyclic guanosine monophosphate pathway in the antidepressant-like effect of tramadol in the rat forced swimming test. Prog. Neuropsychopharmacol. Biol. Psychiatry 2008, 32, 1838–1843. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Wang, Y.; Wu, Z.; Lan, T.; Tian, Y.; Chen, X.; Li, Y.; Dang, R.; Bai, M.; Cheng, K.; et al. Metabolomic abnormalities of purine and lipids implicated olfactory bulb dysfunction of CUMS depressive rats. Metab. Brain Dis. 2020, 35, 649–659. [Google Scholar] [CrossRef] [PubMed]
- Kaddurah-Daouk, R.; Yuan, P.; Boyle, S.H.; Matson, W.; Wang, Z.; Zeng, Z.B.; Zhu, H.; Dougherty, G.G.; Yao, J.K.; Chen, G.; et al. Cerebrospinal fluid metabolome in mood disorders-remission state has a unique metabolic profile. Sci. Rep. 2012, 2, 667. [Google Scholar] [CrossRef]
- Ali-Sisto, T.; Tolmunen, T.; Toffol, E.; Viinamäki, H.; Mäntyselkä, P.; Valkonen-Korhonen, M.; Honkalampi, K.; Ruusunen, A.; Velagapudi, V.; Lehto, S.M. Purine metabolism is dysregulated in patients with major depressive disorder. Psychoneuroendocrinology 2016, 70, 25–32. [Google Scholar] [CrossRef] [Green Version]
- Vaváková, M.; Ďuračková, Z.; Trebatická, J. Markers of Oxidative Stress and Neuroprogression in Depression Disorder. Oxidative Med. Cell. Longev. 2015, 2015, 898393. [Google Scholar] [CrossRef] [Green Version]
- Gałecki, P. Systems Biology of Free Radicals and Antioxidants; Oxidative Stress in Depression; Springer: Berlin/Heidelberg, Germany, 2014; pp. 2369–2395. [Google Scholar]
- Liu, T.; Zhong, S.; Liao, X.; Chen, J.; He, T.; Lai, S.; Jia, Y. A Meta-Analysis of Oxidative Stress Markers in Depression. PLoS ONE 2015, 10, e0138904. [Google Scholar] [CrossRef]
- Eren, I.; Naziroğlu, M.; Demirdaş, A.; Celik, O.; Uğuz, A.C.; Altunbaşak, A.; Ozmen, I.; Uz, E. Venlafaxine modulates depression-induced oxidative stress in brain and medulla of rat. Neurochem. Res. 2007, 32, 497–505. [Google Scholar] [CrossRef]
- Gorlova, A.; Pavlov, D.; Zubkov, E.; Zorkina, Y.; Inozemtsev, A.; Morozova, A.; Chekhonin, V. Alteration of oxidative stress markers and behavior of rats in a novel model of depression. Acta Neurobiol. Exp. 2019, 79, 232–237. [Google Scholar] [CrossRef] [Green Version]
- Bouvier, E.; Brouillard, F.; Molet, J.; Claverie, D.; Cabungcal, J.H.; Cresto, N.; Doligez, N.; Rivat, C.; Do, K.Q.; Bernard, C.; et al. Nrf2-dependent persistent oxidative stress results in stress-induced vulnerability to depression. Mol. Psychiatry 2017, 22, 1701–1713, Erratum in: Mol. Psychiatry 2017, 22, 1795.doi:10.1038/mp.2016.144. [Google Scholar] [CrossRef] [Green Version]
- Ushakova, V.M.; Morozova, A.Y.; Reznik, A.M.; Kostyuk, G.P.; Chekhonin, V.P. Molecular Biological Aspects of Depressive Disorders: A Modern View. Mol. Biol. 2020, 54, 725–749. (In Russian) [Google Scholar] [CrossRef]
- Addinsoft. XLSTAT Statistical and Data Analysis Solution New York, USA. 2021. Available online: https://www.xlstat.com (accessed on 1 July 2021).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolites | PubChem CIDs | Structure | Control | IN Clom | IP Clom | US | F and p-Value | FDR |
|---|---|---|---|---|---|---|---|---|
| Proline | 614 | Hipp | 0.97 ± 0.13 | 1.08 ± 0.28 | 0.83 ± 0.26 | 1.002 ± 0.2 | F = 1.5 p = 0.24 | 0.310 |
| FC | 0.852 ± 0.09 | 0.7 ± 0.21 | 0.95 ± 0.21 | 0.69 ± 0.1 | F = 3.9 p = 0.02 | 0.039 | ||
| Nicotinamide | 936 | Hipp | 4.05 ± 0.32 | 3.54 ± 0.57 | 4.01 ± 0.99 | 3.96 ± 0.45 | F = 0.96 p = 0.43 | 0.470 |
| FC | 4.13 ± 0.54 | 2.81 ± 0.53 | 3.26 ± 0.34 | 3.42 ± 0.42 | F = 9.5 p = 0.0003 | 0.002 | ||
| Phenylalanine | 994 | Hipp | 0.32 ± 0.06 | 0.25 ± 0.071 | 0.41 ± 0.24 | 0.33 ± 0.09 | F = 1.58 p = 0.22 | 0.310 |
| FC | 0.31 ± 0.12 | 0.17 ± 0.051 | 0.23 ± 0.04 | 0.23 ± 0.068 | F = 3.6 p = 0.03 | 0.050 | ||
| Alanine | 602 | Hipp | 0.14 ± 0.011 | 0.12 ± 0.027 | 0.17 ± 0.06 | 0.14 ± 0.017 | F = 2.4 p = 0.09 | 0.182 |
| FC | 0.11 ± 0.037 | 0.08 ± 0.018 | 0.14 ± 0.016 | 0.09 ± 0.02 | F = 7.3 p = 0.001 | 0.005 | ||
| Adenine | 190 | Hipp | 0.1 ± 0.01 | 0.09 ± 0.021 | 0.18 ± 0.054 | 0.1 ± 0.011 | F = 13.14 p < 0.001 | <0.001 |
| FC | 0.09 ± 0.012 | 0.07 ± 0.013 | 0.091 ± 0.012 | 0.08 ± 0.014 | F = 3.17 p = 0.043 | 0.066 | ||
| Serine | 617 | Hipp | 0.35 ± 0.057 | 0.28 ± 0.067 | 0.44 ± 0.15 | 0.35 ± 0.064 | F = 3.4 p = 0.033 | 0.110 |
| FC | 0.31 ± 0.06 | 0.23 ± 0.022 | 0.32 ± 0.05 | 0.24 ± 0.025 | F = 8.5 p = 0.0005 | 0.003 | ||
| Glyceraldehyde | 751 | Hipp | 0.027 ± 0.019 | 0.015 ± 0.005 | 0.016 ± 0.015 | 0.028 ± 0.015 | F = 1.5 p = 0.24 | 0.310 |
| FC | 0.015 ± 0.009 | 0.003 ± 0.002 | 0.003 ± 0.002 | 0.008 ± 0.005 | F = 7.34 p = 0.001 | 0.005 | ||
| Cytidine | 6175 | Hipp | 1.78 ± 0.34 | 1.34 ± 0.35 | 1.91 ± 0.64 | 1.58 ± 0.26 | F = 2.42 p = 0.091 | 0.182 |
| FC | 1.16 ± 0.29 | 0.92 ± 0.41 | 1.01 ± 0.18 | 0.89 ± 0.12 | F = 1.4 p = 0.27 | 0.282 | ||
| Acetylcarnitine | 7045767 | Hipp | 0.94 ± 0.21 | 0.83 ± 0.19 | 0.99 ± 0.32 | 1.1 ± 0.25 | F = 1.5 p = 0.24 | 0.310 |
| FC | 0.53 ± 0.17 | 0.39 ± 0.15 | 0.39 ± 0.12 | 0.5 ± 0.07 | F = 2 p = 0.14 | 0.170 | ||
| Carnitine | 288 | Hipp | 2.21 ± 0.29 | 1.8 ± 0.37 | 0.66 ± 0.16 | 2.13 ± 0.52 | F = 27.53 p < 0.001 | <0.001 |
| FC | 1.12 ± 0.18 | 0.83 ± 0.33 | 0.92 ± 0.27 | 0.94 ± 0.19 | F = 1.6 p = 0.22 | 0.250 | ||
| Xanthine | 1188 | Hipp | 0.15 ± 0.06 | 0.09 ± 0.061 | 0.09 ± 0.023 | 0.14 ± 0.047 | F = 2.5 p = 0.08 | 0.182 |
| FC | 0.02 ± 0.009 | 0.043 ± 0.03 | 0.038 ± 0.027 | 0.024 ± 0.006 | F = 2 p = 0.14 | 0.170 | ||
| l-glutation oxydazed | 65359 | Hipp | 0.2 ± 0.05 | 0.11 ± 0.052 | 0.16 ± 0.098 | 0.14 ± 0.018 | F = 2.7 p = 0.07 | 0.182 |
| FC | 0.11 ± 0.03 | 0.081 ± 0.042 | 0.07 ± 0.023 | 0.06 ± 0.02 | F = 3.5 p = 0.032 | 0.053 | ||
| Uridine | 6029 | Hipp | 0.05 ± 0.01 | 0.04 ± 0.015 | 0.037 ± 0.012 | 0.05 ± 0.012 | F = 1.87 p = 0.16 | 0.270 |
| FC | 0.028 ± 0.005 | 0.022 ± 0.009 | 0.025 ± 0.008 | 0.02 ± 0.005 | F = 1.4 p = 0.28 | 0.282 | ||
| Ribose 5-phosphate | 77982 | Hipp | 0.07 ± 0.031 | 0.026 ± 0.013 | 0.029 ± 0.02 | 0.05 ± 0.012 | F = 6.5 p = 0.002 | 0.010 |
| FC | 0.027 ± 0.007 | 0.017 ± 0.011 | 0.014 ± 0.005 | 0.013 ± 0.009 | F = 4 p = 0.018 | 0.039 | ||
| Threonine | 205 | Hipp | 1.24 ± 0.21 | 1.04 ± 0.28 | 0.96 ± 0.32 | 1.14 ± 0.27 | F = 1.4 0.28 | 0.336 |
| FC | 1.39 ± 0.37 | 0.9 ± 0.132 | 0.98 ± 0.14 | 0.97 ± 0.21 | F = 6.4 0.002 | 0.008 | ||
| Glycine | 750 | Hipp | 0.0018 ± 0.0003 | 0.0015 ± 0.0005 | 0.0021 ± 0.001 | 0.0017 ± 0.0004 | F = 0.8 p = 0.49 | 0.510 |
| FC | 0.0012 ± 0.0004 | 0.001 ± 0.0003 | 0.0017 ± 0.0003 | 0.0011 ± 0.0002 | F = 4.4 p = 0.013 | 0.038 | ||
| Fructose 6-phosphate | 69507 | Hipp | 0.0085 ± 0.009 | 0.002 ± 0.0016 | 0.003 ± 0.003 | 0.0068 ± 0.004 | F = 2.17 p = 0.12 | 0.220 |
| FC | 0.0021 ± 0.0009 | 0.00078 ± 0.0002 | 0.00078 ± 0.0009 | 0.0024 ± 0.002 | F = 4.25 p = 0.015 | 0.039 | ||
| Citrulline | 833 | Hipp | 0.58 ± 0.14 | 0.65 ± 0.17 | 1.1 ± 0.4 | 0.645 ± 0.14 | F = 7 p = 0.002 | 0.008 |
| FC | 0.44 ± 0.14 | 0.33 ± 0.08 | 0.45 ± 0.087 | 0.335 ± 0.09 | F = 2.81 p = 0.06 | 0.088 | ||
| GABA | 119 | Hipp | 2.35 ± 0.52 | 1.15 ± 0.29 | 3.62 ± 0.98 | 1.5 ± 0.32 | F = 23.7 p < 0.001 | <0.001 |
| FC | 1.71 ± 0.4 | 0.74 ± 0.18 | 0.97 ± 0.22 | 1.41 ± 0.2 | F = 18.55 p < 0.001 | <0.001 | ||
| Tryptophan | 1148 | Hipp | 0.5 ± 0.11 | 0.42 ± 0.13 | 0.7 ± 0.42 | 0.47 ± 0.17 | F = 1.75 p = 0.18 | 0.280 |
| FC | 0.58 ± 0.16 | 0.31 ± 0.11 | 0.33 ± 0.05 | 0.37 ± 0.098 | F = 8.6 p = 0.0005 | 0.003 | ||
| Guanosine | 135398635 | Hipp | 0.005 ± 0.002 | 0.0037 ± 0.002 | 0.0082 ± 0.0045 | 0.005 ± 0.001 | F = 3.3 p = 0.04 | 0.120 |
| FC | 0.0013 ± 0.0005 | 0.004 ± 0.0048 | 0.003 ± 0.002 | 0.0011 ± 0.0005 | F = 2.2 p = 0.11 | 0.147 | ||
| AMP | 6083 | Hipp | 1.75 ± 0.57 | 1.98 ± 0.88 | 1.5 ± 0.8 | 1.88 ± 0.59 | F = 0.57 p = 0.64 | 0.641 |
| FC | 2.84 ± 0.46 | 1.61 ± 0.61 | 2.06 ± 0.77 | 2.4 ± 0.43 | F = 5.5 p = 0.005 | 0.015 | ||
| Arginine | 232 | Hipp | 0.21 ± 0.03 | 0.15 ± 0.05 | 0.15 ± 0.07 | 0.196 ± 0.045 | F = 2.5 p = 0.08 | 0.182 |
| FC | 0.2 ± 0.05 | 0.15 ± 0.057 | 0.18 ± 0.033 | 0.174 ± 0.038 | F = 1.35 p = 0.28 | 0.282 | ||
| Aspartic acid | 424 | Hipp | 0.36 ± 0.05 | 0.25 ± 0.065 | 0.32 ± 0.16 | 0.35 ± 0.065 | F = 1.9 p = 0.15 | 0.260 |
| FC | 0.39 ± 0.09 | 0.26 ± 0.072 | 0.33 ± 0.05 | 0.31 ± 0.074 | F = 3.98 p = 0.02 | 0.039 | ||
| Glutaminic acid | 611 | Hipp | 4.89 ± 0.4 | 4.24 ± 1.01 | 4.24 ± 1.05 | 4.66 ± 0.93 | F = 0.94 p = 0.44 | 0.470 |
| FC | 4.76 ± 0.71 | 3.4 ± 1.23 | 3.83 ± 0.46 | 3.63 ± 0.45 | F = 4 p = 0.02 | 0.039 | ||
| Methionine | 876 | Hipp | 0.1 ± 0.017 | 0.06 ± 0.018 | 0.17 ± 0.1 | 0.088 ± 0.014 | F = 5.88 p = 0.004 | 0.015 |
| FC | 0.087 ± 0.02 | 0.038 ± 0.014 | 0.039 ± 0.011 | 0.062 ± 0.015 | F = 14 p < 0.001 | <0.001 | ||
| Ionosine | 135398641 | Hipp | 0.059 ± 0.05 | 0.04 ± 0.018 | 0.068 ± 0.027 | 0.046 ± 0.019 | F = 1.2 p = 0.32 | 0.380 |
| FC | 0.015 ± 0.007 | 0.025 ± 0.016 | 0.019 ± 0.014 | 0.008 ± 0.004 | F = 2.5 p = 0.08 | 0.112 | ||
| Hypoxanthine | 135398638 | Hipp | 0.17 ± 0.06 | 0.14 ± 0.035 | 0.067 ± 0.022 | 0.14 ± 0.018 | F = 9.4 p = 0.0003 | 0.002 |
| FC | 0.127 ± 0.02 | 0.11 ± 0.022 | 0.13 ± 0.045 | 0.103 ± 0.03 | F = 1.55 p = 0.23 | 0.254 |
| Brain Structure | Frontal Cortex | Hippocampus | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Group | Control | IN Clom | IP Clom | US | Control | IN Clom | IP Clom | US | |
| Control | SE | - | 0.15 | 0.339 | 0.128 | - | 0.171 | 0.207 | 0.181 |
| t | - | 4.262 | 1.501 | 0.181 | - | −1.471 | −3.218 | −0.939 | |
| df | - | 12 | 12 | 12 | - | 12 | 12 | 12 | |
| p | - | 0.001 * | 0.159 | 0.86 | - | 0.167 | 0.007 * | 0.366 | |
| IN Clom | SE | 0.15 | - | 0.361 | 0.169 | 0.171 | - | 0.121 | 0.072 |
| t | 4.262 | - | −0.364 | −3.645 | −1.471 | - | −3.432 | 1.129 | |
| df | 12 | - | 12 | 12 | 12 | - | 12 | 12 | |
| p | 0.001 * | - | 0.722 | 0.003 * | 0.167 | - | 0.005 * | 0.281 | |
| IP Clom | SE | 0.339 | 0.361 | - | 0.37 | 0.207 | 0.121 | - | 0.137 |
| t | 1.501 | −0.364 | - | −1.315 | −3.218 | −3.432 | - | 3.628 | |
| df | 12 | 12 | - | 12 | 12 | 12 | - | 12 | |
| p | 0.159 | 0.722 | - | 0.213 | 0.007 * | 0.005 * | - | 0.003 * | |
| US | SE | 0.128 | 0.169 | 0.37 | - | 0.181 | 0.072 | 0.137 | - |
| t | 0.181 | −3.645 | −1.315 | - | −0.939 | 1.129 | 3.628 | - | |
| df | 12 | 12 | 12 | - | 12 | 12 | 12 | - | |
| p | 0.86 | 0.003 * | 0.213 | - | 0.366 | 0.281 | 0.003 * | - | |
| Substance | Q1 | Q3 | Polarity | |
|---|---|---|---|---|
| l-Glutathione oxidized | 613.008 | 354.9 | 484 | + |
| Adenine | 135.93 | 119 | 92 | + |
| Adenosinmonophosphat | 347.971 | 136 | 119 | + |
| Alanine | 89.988 | 44.1 | N/A | + |
| Arginine | 175.074 | 70 | 116 | + |
| Aspartic acid | 133.921 | 74 | 88 | + |
| γ-Aminobutyric acid | 104.076 | 87 | 69 | + |
| Hypoxanthine | 136.97 | 119 | 94 | + |
| Glyceraldehyde 3-phosphate | 168.748 | 97 | 78.9 | − |
| Glycine | 75.985 | 30 | 48 | + |
| Guanosine | 281.834 | 150 | 133 | − |
| Inosine | 266.762 | 134.9 | 134.5 | − |
| Carnitine | 162.011 | 103 | 60.1 | + |
| Xanthine | 150.833 | 107.9 | 42 | − |
| Methionine | 150.006 | 104 | 133 | + |
| Nicotinamide | 122.99 | 80 | 78 | + |
| O-Acetyl-l-carnitine | 204.085 | 85 | 145.1 | + |
| Proline | 116.03 | 70.1 | N/A | + |
| Ribose 5-phosphate | 228.69 | 96.9 | 78.9 | − |
| Serine | 105.982 | 60 | 42 | + |
| Threonine | 119.969 | 74 | 102 | + |
| Tryptophan | 205.01 | 188.1 | 145.9 | + |
| Uridine | 242.799 | 199.8 | 110 | − |
| Phenylalanine | 166.011 | 119.9 | 102.9 | + |
| Citidine | 244.016 | 112 | 95 | + |
| Citrulline | 176.021 | 158.9 | 113.1 | + |
| Cotinine-d3 | 180.04 | 85 | 101 | + |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abramova, O.; Zorkina, Y.; Syunyakov, T.; Zubkov, E.; Ushakova, V.; Silantyev, A.; Soloveva, K.; Gurina, O.; Majouga, A.; Morozova, A.; et al. Brain Metabolic Profile after Intranasal vs. Intraperitoneal Clomipramine Treatment in Rats with Ultrasound Model of Depression. Int. J. Mol. Sci. 2021, 22, 9598. https://doi.org/10.3390/ijms22179598
Abramova O, Zorkina Y, Syunyakov T, Zubkov E, Ushakova V, Silantyev A, Soloveva K, Gurina O, Majouga A, Morozova A, et al. Brain Metabolic Profile after Intranasal vs. Intraperitoneal Clomipramine Treatment in Rats with Ultrasound Model of Depression. International Journal of Molecular Sciences. 2021; 22(17):9598. https://doi.org/10.3390/ijms22179598
Chicago/Turabian StyleAbramova, Olga, Yana Zorkina, Timur Syunyakov, Eugene Zubkov, Valeria Ushakova, Artemiy Silantyev, Kristina Soloveva, Olga Gurina, Alexander Majouga, Anna Morozova, and et al. 2021. "Brain Metabolic Profile after Intranasal vs. Intraperitoneal Clomipramine Treatment in Rats with Ultrasound Model of Depression" International Journal of Molecular Sciences 22, no. 17: 9598. https://doi.org/10.3390/ijms22179598