
Esterase Activity of Serum Albumin Studied by 1H NMR Spectroscopy and Molecular Modelling

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
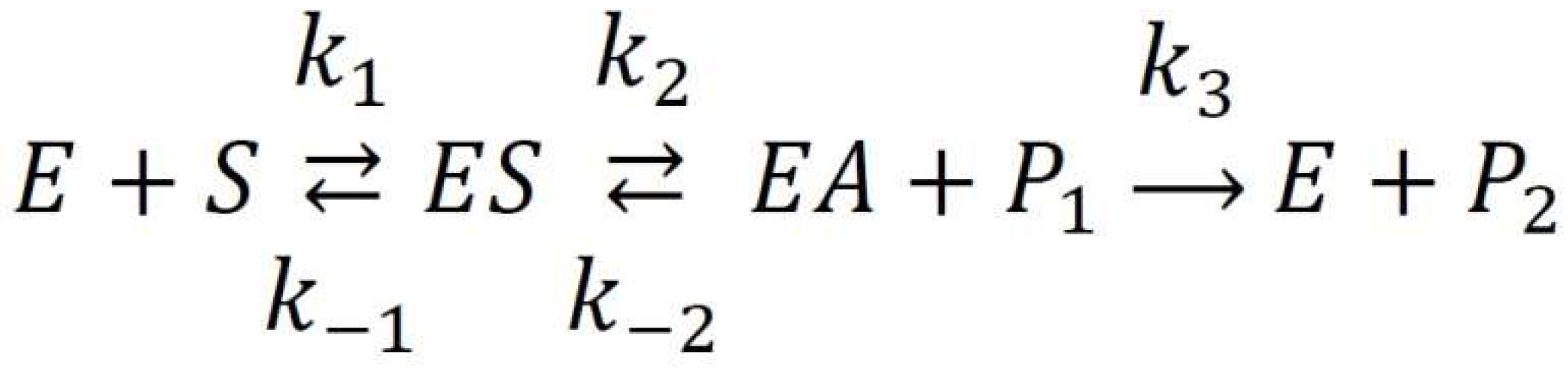{kind=link}
Abstract
:1. Introduction
2. Results
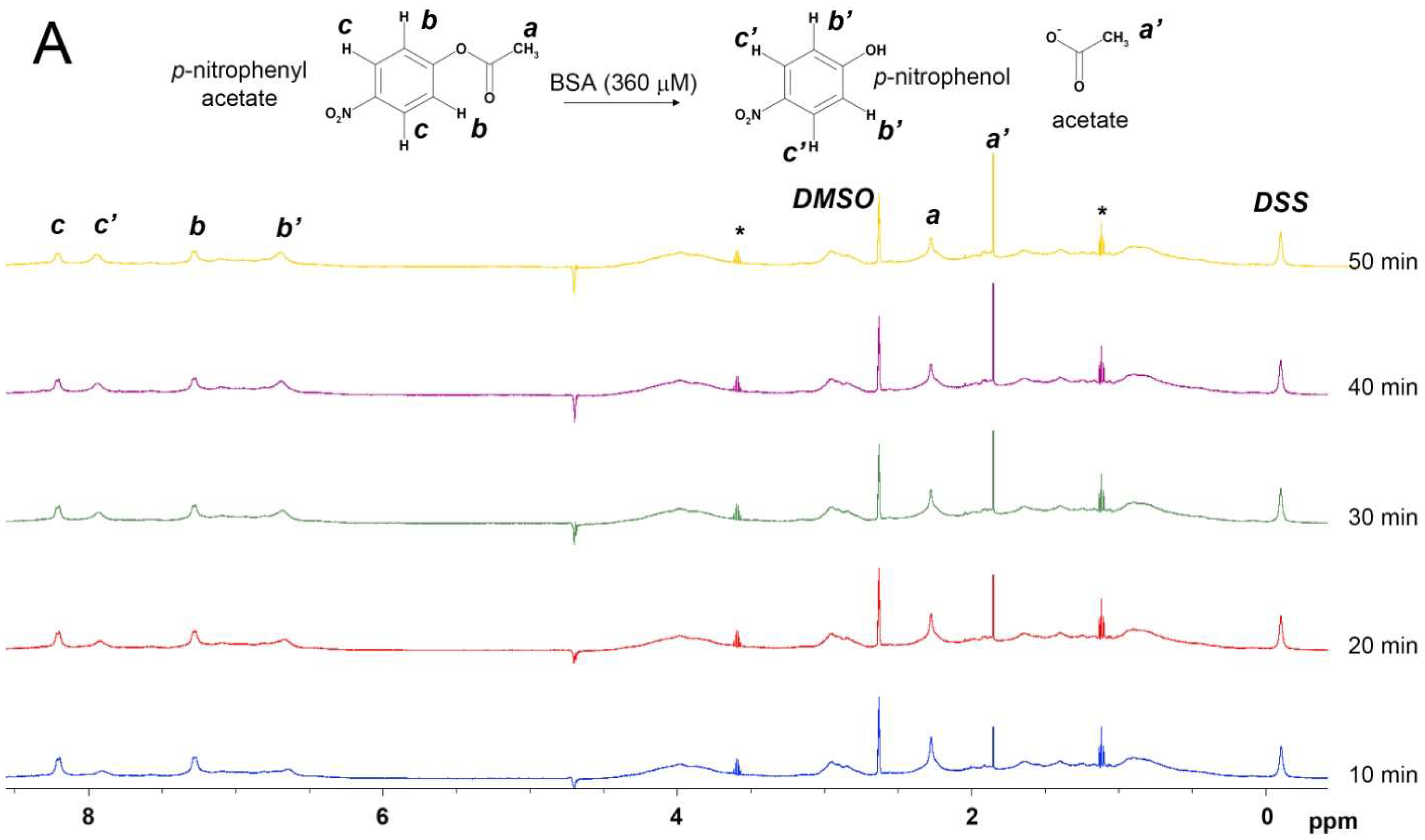
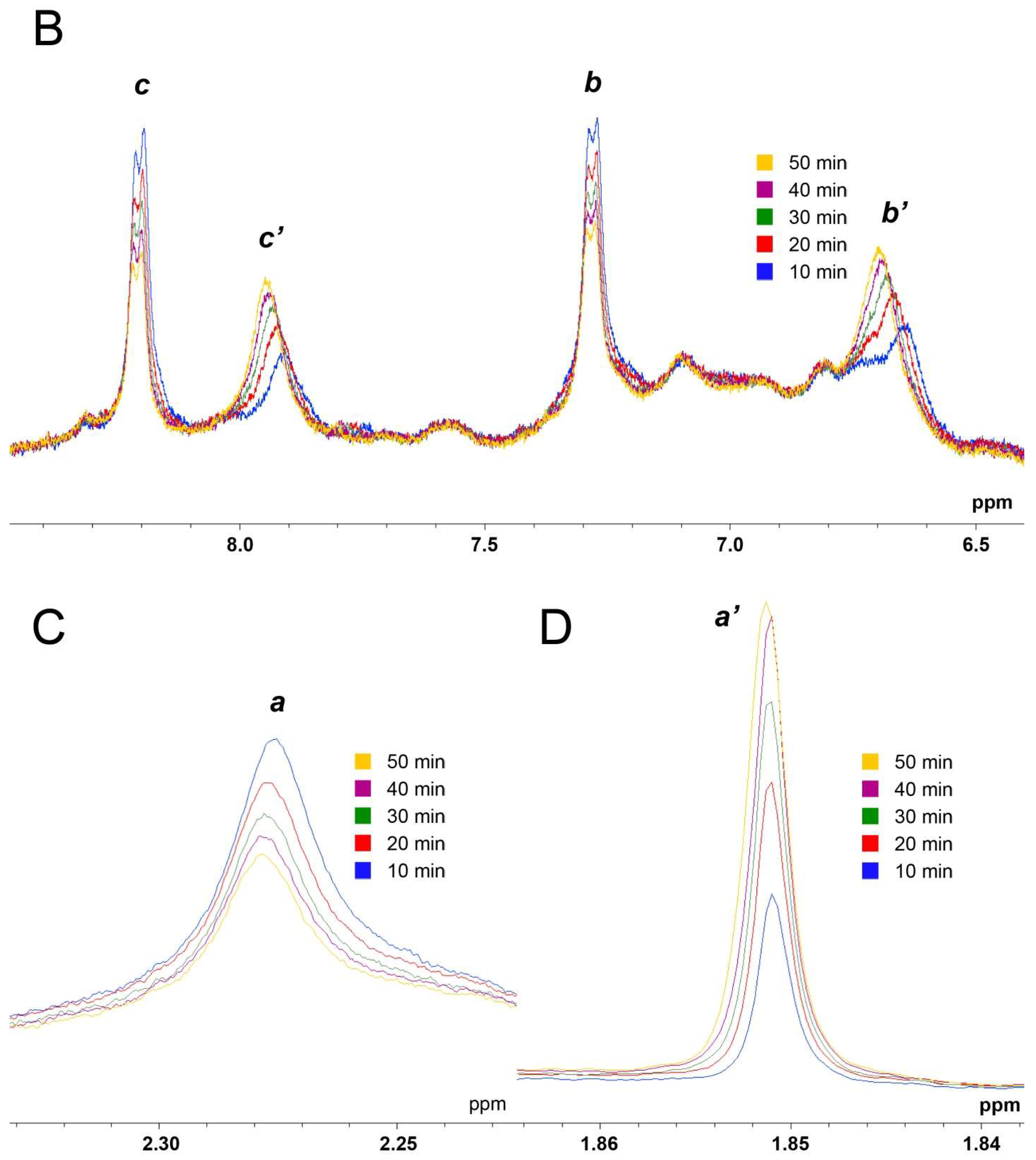
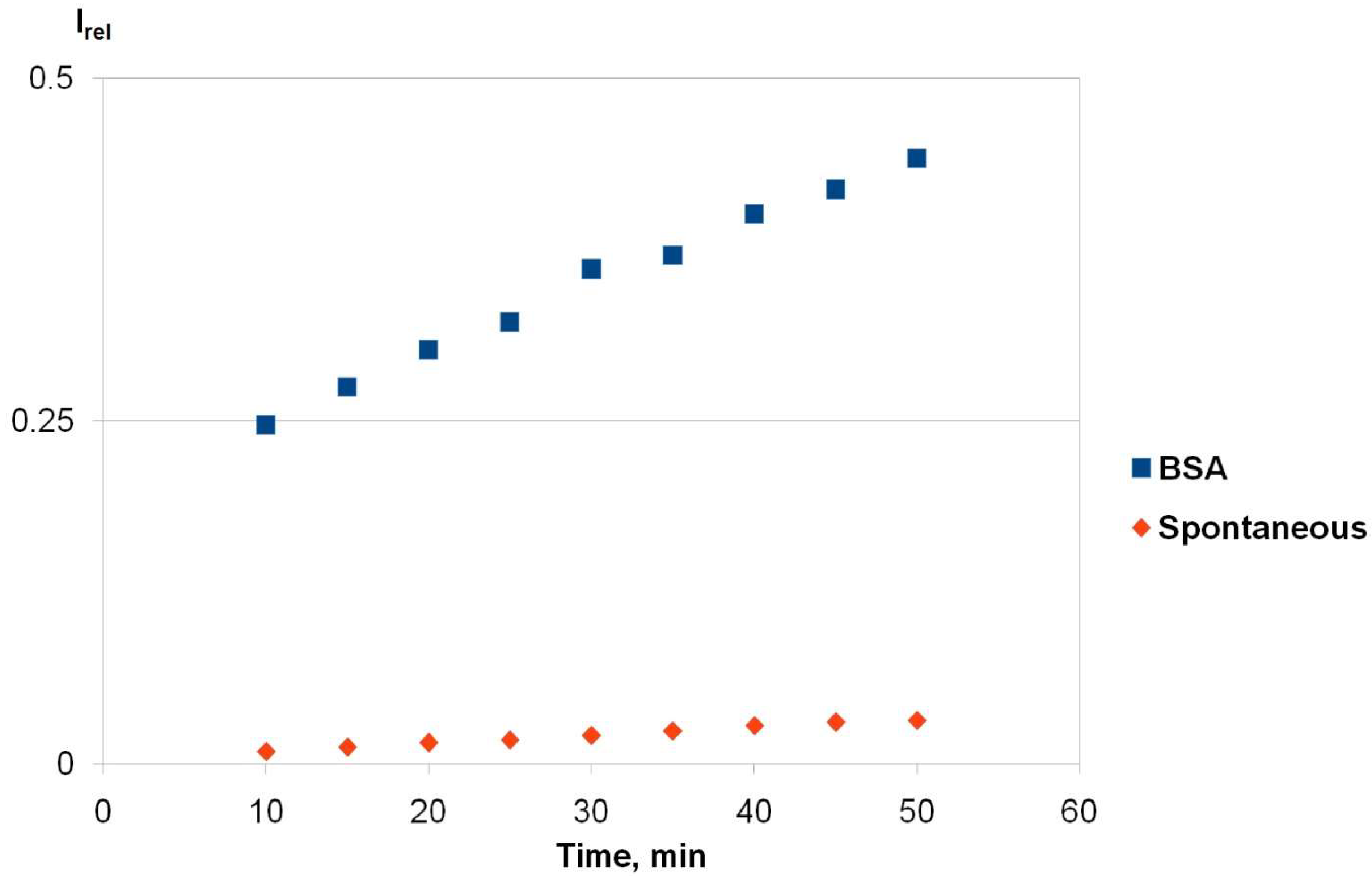
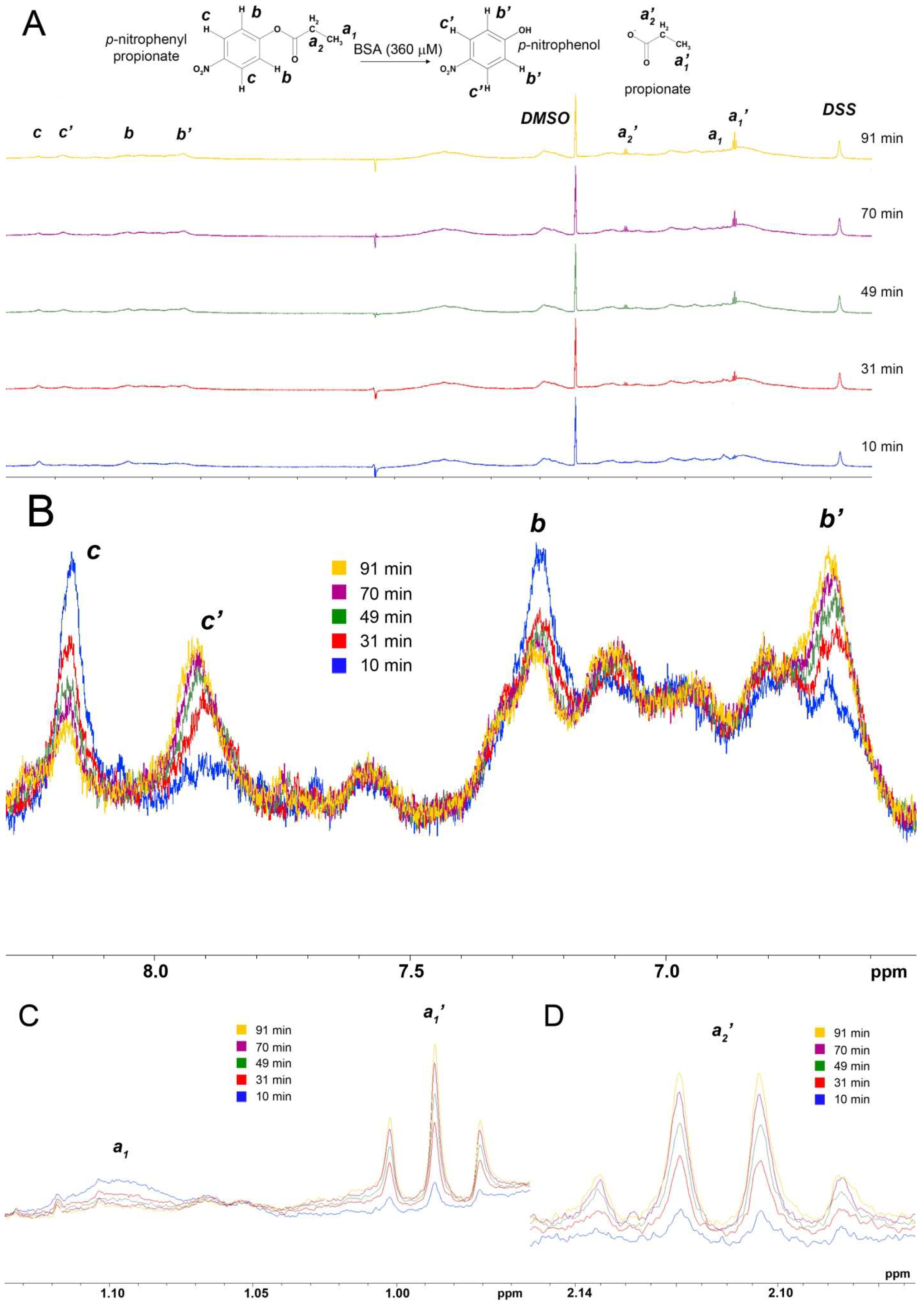
2.1. True Esterase Activity of Albumin towards NPA According to 1H NMR
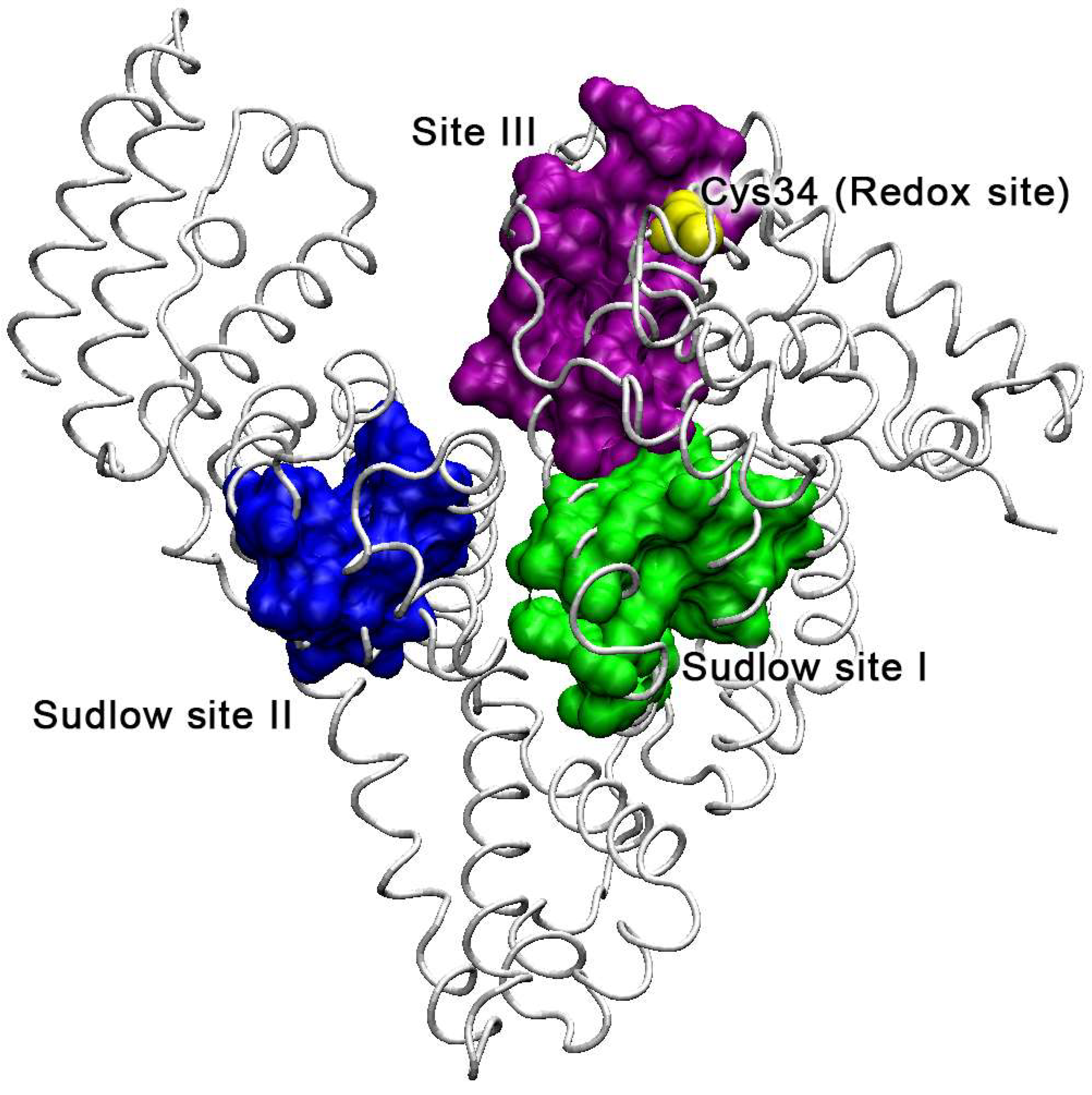
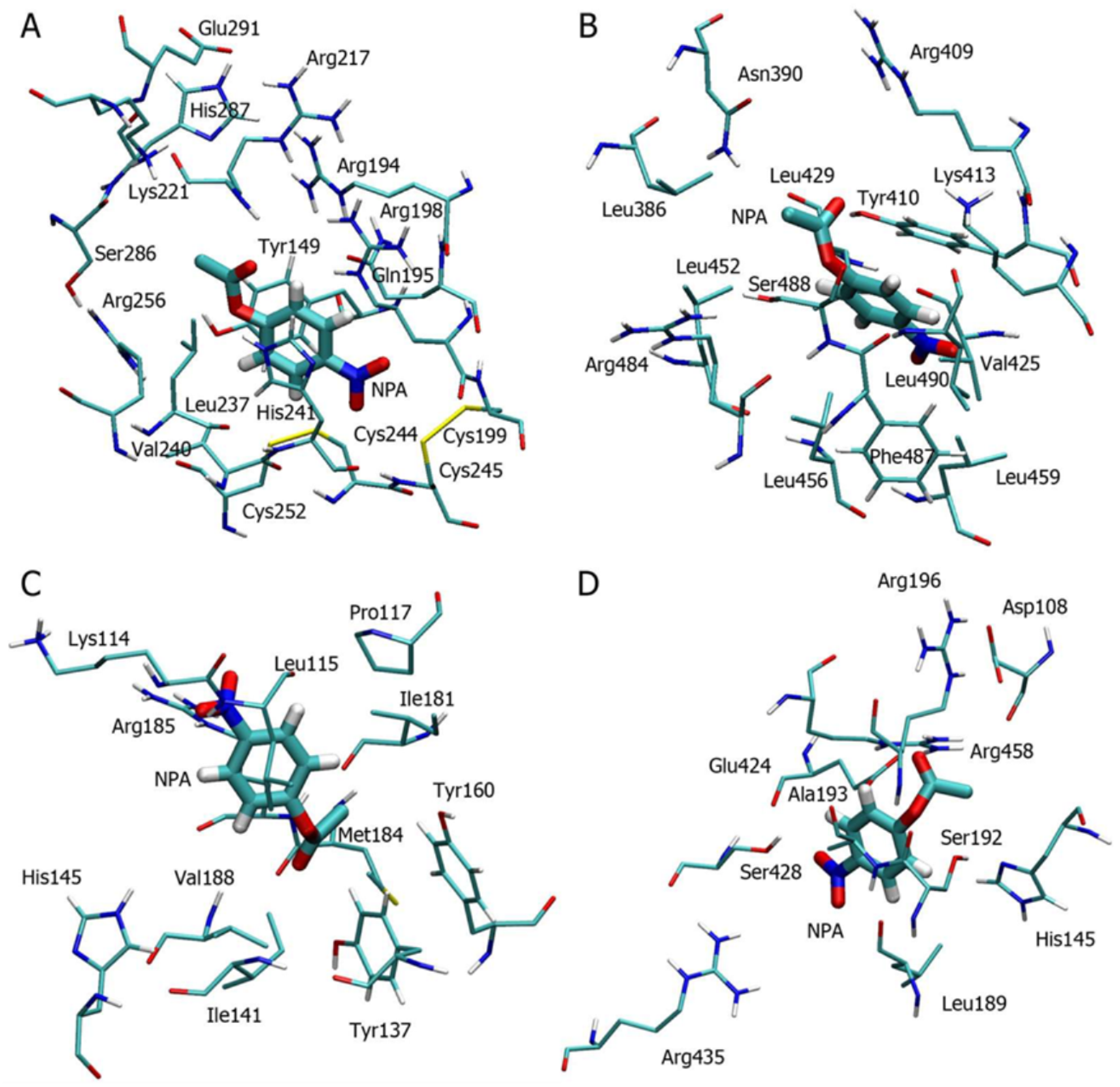
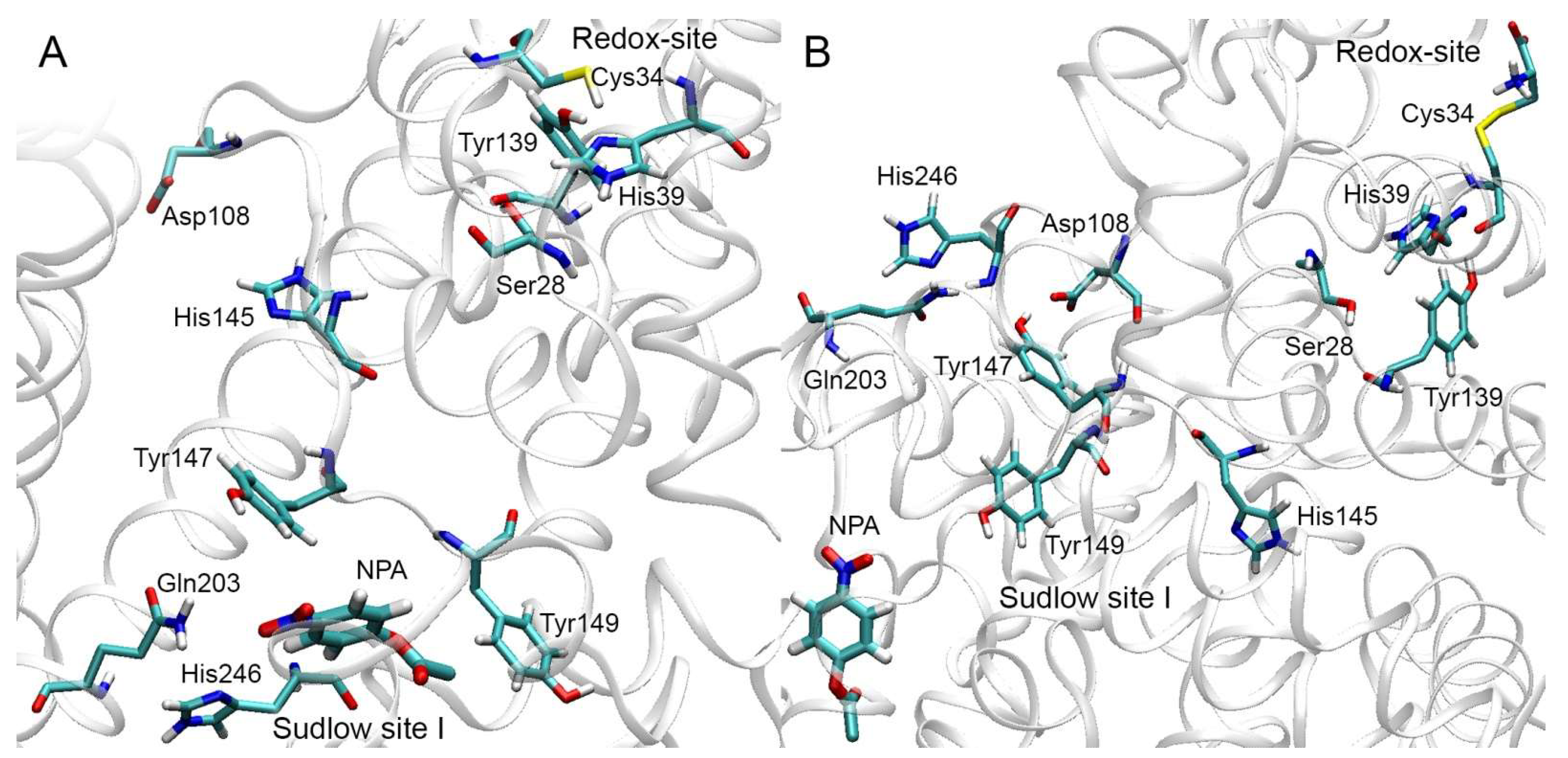
2.2. Analysis of Possible Sites of Albumin Esterase Activity
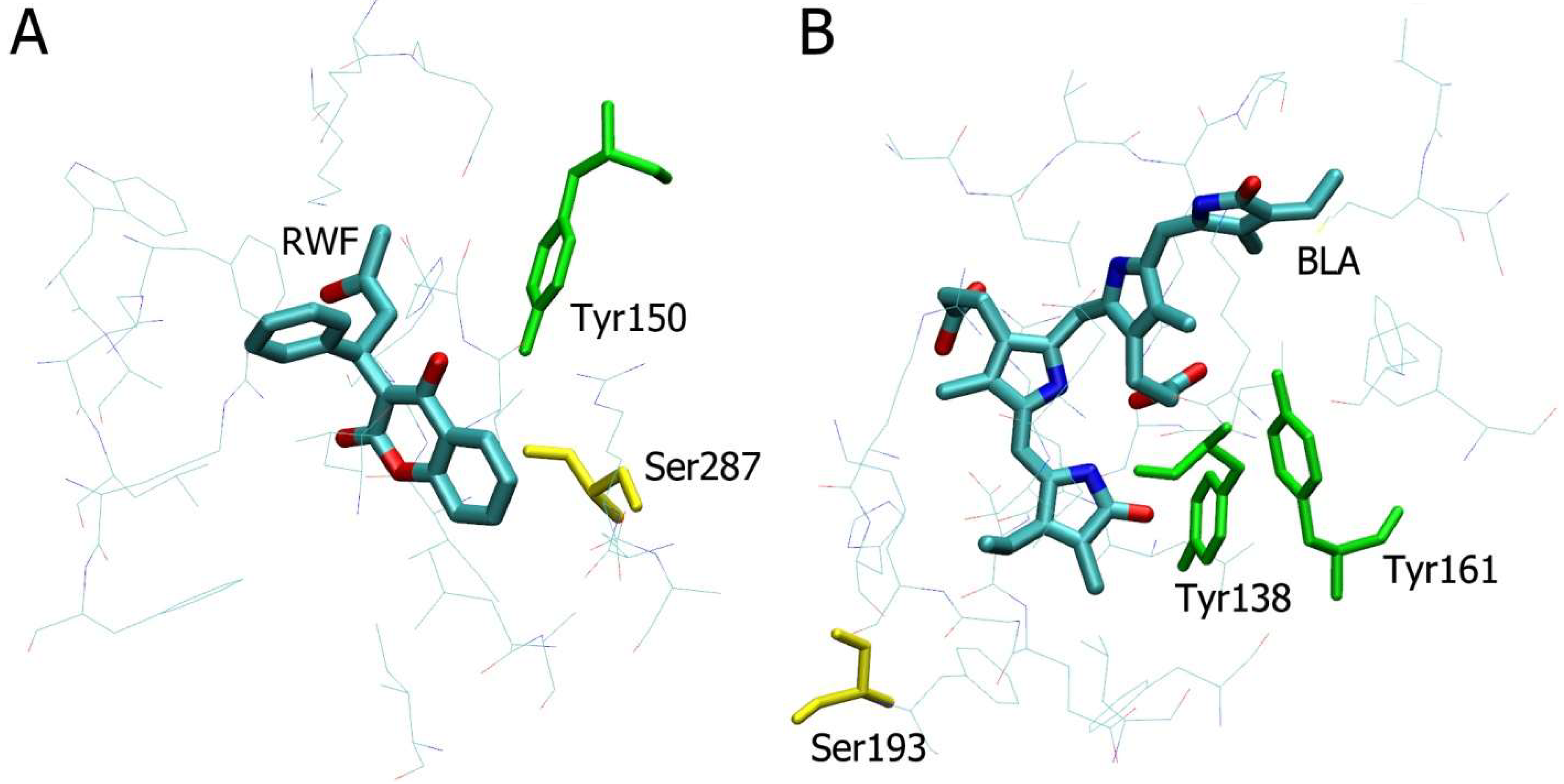
2.3. Molecular Docking of NPA into the Selected Binding Sites
2.4. Stability of NPA-BSA Complexes According to MD Simulation
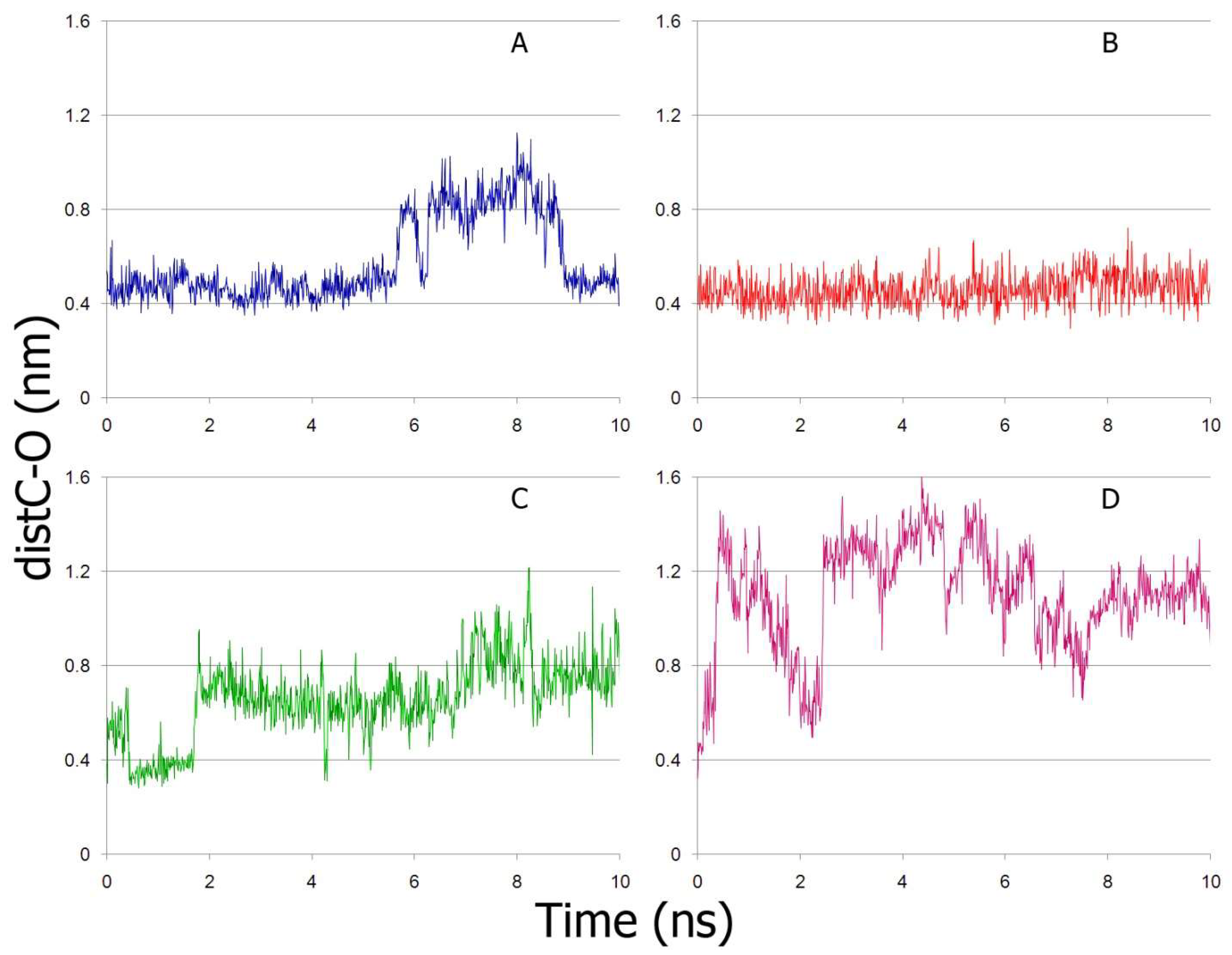
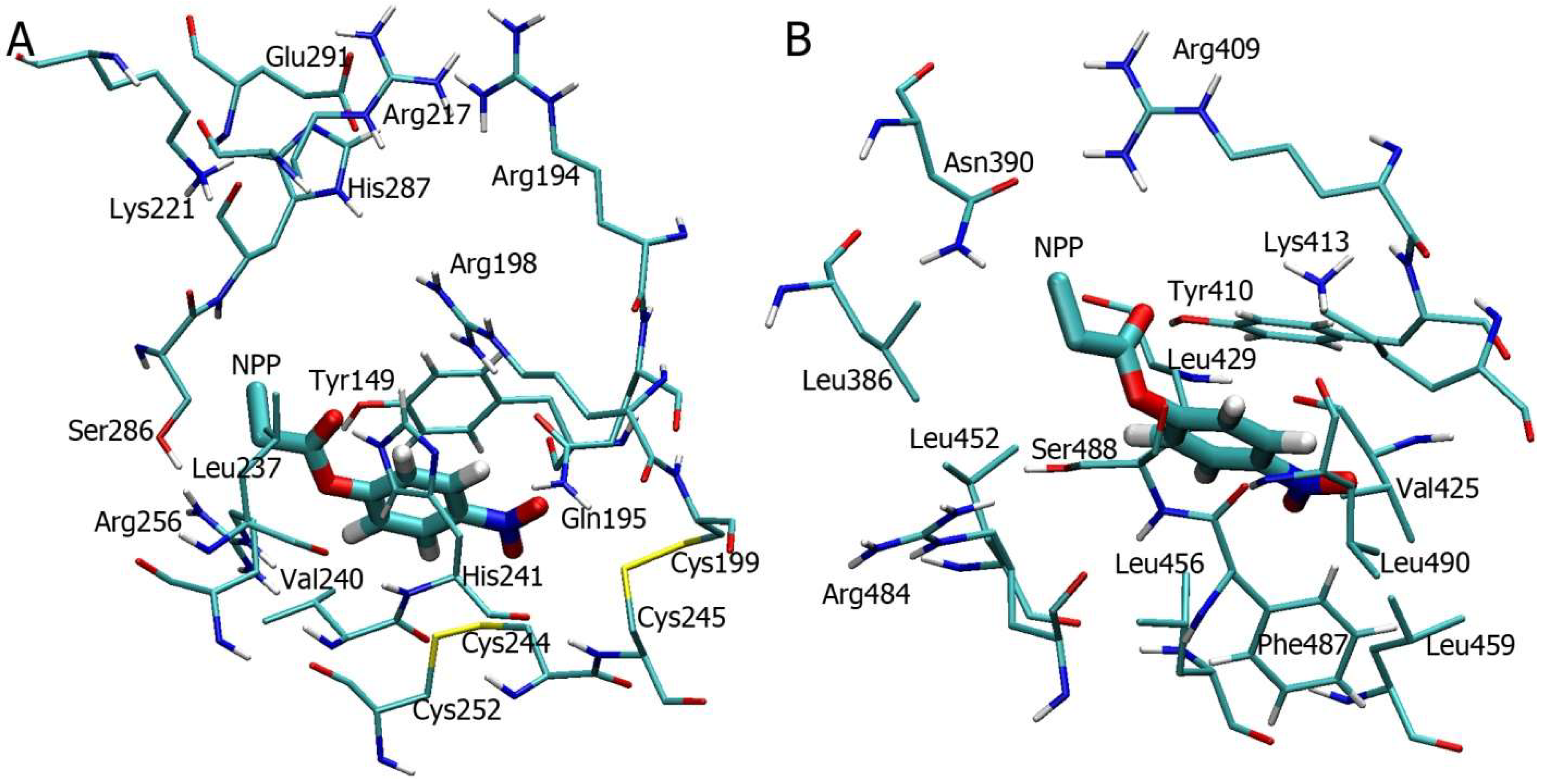
2.5. Interaction of NPA with Sudlow Site I and Sudlow Site II of BSA According to MD Simulation
2.6. Interaction of NPA with Sudlow Site I and Sudlow Site II of Oxidised BSA According to MD Simulation
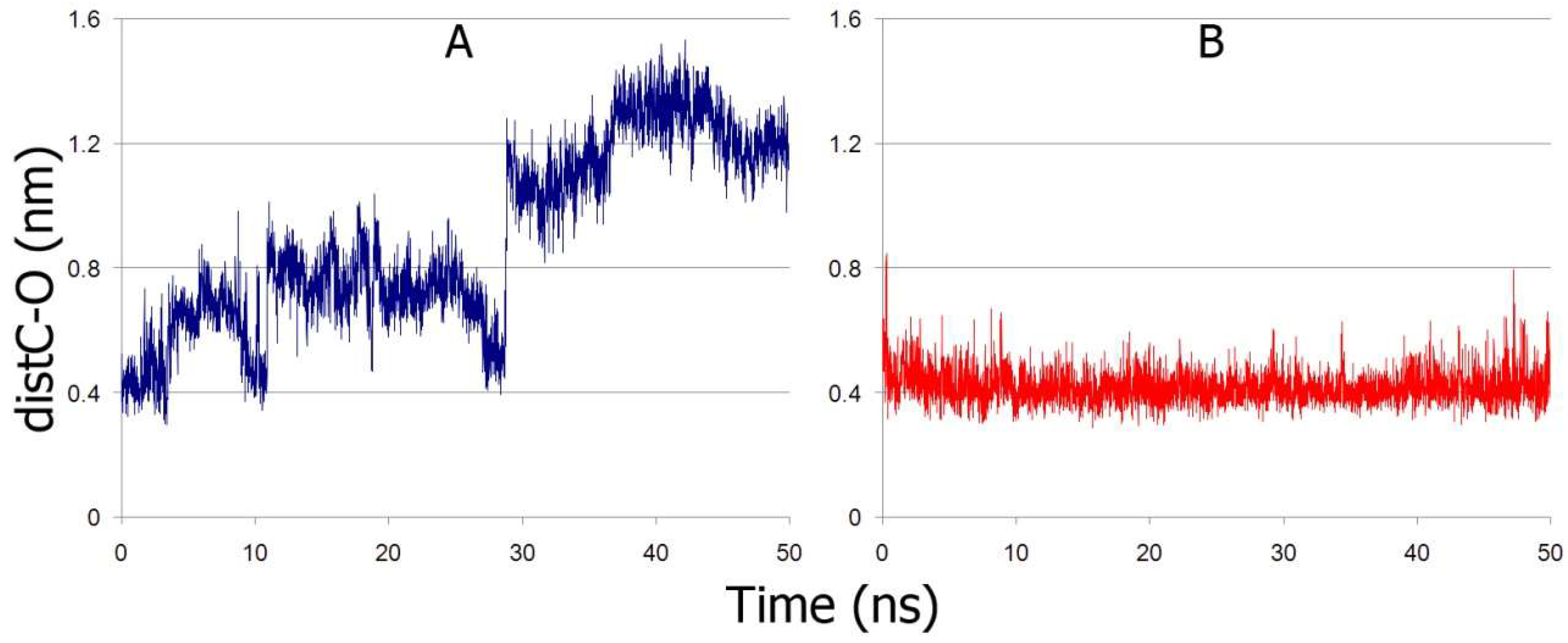
2.7. Interaction of BSA with p-Nitrophenyl Propionate
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. NMR Spectroscopy
4.3. Preparation of BSA and NPA Three-Dimensional Models
4.4. Molecular Docking
4.5. Molecular Dynamics Simulation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.N.; Bourne, P.E. The Protein Data Bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef] [Green Version]
- Sekula, B.; Zielinski, K.; Bujacz, A. Crystallographic studies of the complexes of bovine and equine serum albumin with 3,5-diiodosalicylic acid. Int. J. Biol. Macromol. 2013, 60, 316–324. [Google Scholar] [CrossRef]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual molecular dynamics. J. Mol. Graph. 1996, 14, 28–33. [Google Scholar] [CrossRef]
- Goncharov, N.V.; Belinskaya, D.A.; Razygraev, A.V.; Ukolov, A.I. On the enzymatic activity of albumin. Russ. J. Bioorg. Chem. 2015, 41, 113–124. [Google Scholar] [CrossRef]
- Lockridge, O.; Xue, W.; Gaydess, A.; Grigoryan, H.; Ding, S.J.; Schopfer, L.M.; Hinrichs, S.H.; Masson, P. Pseudo-esterase activity of human albumin: Slow turnover on tyrosine 411 and stable acetylation of 82 residues including 59 lysines. J. Biol. Chem. 2008, 283, 22582–22590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belinskaia, D.A.; Goncharov, N.V. Theoretical and practical aspects of albumin esterase ctivity. Russ. J. Bioorg. Chem. 2020, 46, 287–298. [Google Scholar] [CrossRef]
- Casida, J.E.; Augustinsson, K.B. Reaction of plasma albumin with I-naphthyl N-methylcarbamate and certain other esters. Biochim. Biophys. Acta 1959, 36, 411–426. [Google Scholar] [CrossRef]
- Tildon, J.T.; Ogilvie, J.W. The esterase activity of bovine mercaptalbumin. The reaction of the protein with p-nitrophenyl acetate. J. Biol. Chem. 1972, 247, 1265–1271. [Google Scholar] [CrossRef]
- Zaidi, N.; Ajmal, M.R.; Rabbani, G.; Ahmad, E.; Khan, R.H. A comprehensive insight into binding of hippuric acid to human serum albumin: A study to uncover its impaired elimination through hemodialysis. PLoS ONE 2013, 8, e71422. [Google Scholar] [CrossRef] [Green Version]
- Sogorb, M.A.; Vilanova, E. Enzymes involved in the detoxification of organophosphorus, carbamate and pyrethroid insecticides through hydrolysis. Toxicol. Lett. 2002, 128, 215–228. [Google Scholar] [CrossRef]
- Sogorb, M.A.; García-Argüelles, S.; Carrera, V.; Vilanova, E. Serum albumin is as efficient as paraxonase in the detoxication of paraoxon at toxicologically relevant concentrations. Chem. Res. Toxicol. 2008, 21, 1524–1529. [Google Scholar] [CrossRef] [PubMed]
- Sogorb, M.A.; Vilanova, E. Serum albumins and detoxication of anti-cholinesterase agents. Chem. Biol. Interact. 2010, 187, 325–329. [Google Scholar] [CrossRef] [PubMed]
- Belinskaya, D.A.; Shmurak, V.I.; Prokofieva, D.S.; Goncharov, N.V. Investigation of soman binding with albumin by a molecular modeling method. Toksikol. Vestnik Toxicol. Bull. 2012, 116, 13–19. (In Russian) [Google Scholar]
- Belinskaia, D.A.; Shmurak, V.I.; Prokof’eva, D.S.; Goncharov, N.V. Serum albumin: Search for new sites of interaction with organophosphorus compounds by the example of soman. Russ. J. Bioorg. Chem. 2014, 40, 499–506. [Google Scholar] [CrossRef]
- Goncharov, N.V.; Terpilovskii, M.A.; Belinskaya, D.A.; Shmurak, V.I.; Avdonin, P.V. Comparative analysis of esterase and paraoxonase activities of different serum albumin species. J. Evol. Biochem. Physiol. 2017, 53, 271–281. [Google Scholar] [CrossRef]
- Schanda, P. Development and Application of Fast NMR Methods for the Study of Protein Structure and Dynamics. Biol. Phys. 2007. Available online: https://tel.archives-ouvertes.fr/tel-00181457/document (accessed on 22 September 2021).
- Arthanari, H.; Takeuchi, K.; Dubey, A.; Wagner, G. Emerging solution NMR methods to illuminate the structural and dynamic properties of proteins. Curr. Opin. Struct. Biol. 2019, 58, 294–304. [Google Scholar] [CrossRef]
- Denmark, S.E.; Williams, B.J.; Eklov, B.M.; Pham, S.M.; Beutner, G.L. Design, validation, and implementation of a rapid-injection NMR system. J. Org. Chem. 2010, 75, 5558–5572. [Google Scholar] [CrossRef]
- Jensen, P.R.; Matos, M.R.A.; Sonnenschein, N.; Meier, S. Combined in-cell NMR and simulation approach to probe redox-dependent pathway control. Anal. Chem. 2019, 91, 5395–5402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mine, M.; Matsumoto, N.; Mizuguchi, H.; Takayanagi, T. Kinetic analysis of an enzymatic hydrolysis of p-nitrophenyl acetate with carboxylesterase by pressure-assisted capillary electrophoresis/dynamic frontal analysis. Anal. Methods 2020, 12, 5846–5851. [Google Scholar] [CrossRef]
- Lockridge, O.; Quinn, D.M. Esterases. In Comprehensive Toxicology, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 2010; Volume 4, pp. 243–273. [Google Scholar] [CrossRef]
- Rudakova, E.V.; Boltneva, N.P.; Makhaeva, G.F. Comparative analysis of esterase activities of human, mouse, and rat blood. Bull. Exp. Biol. Med. 2011, 152, 73–75. [Google Scholar] [CrossRef]
- Ghuman, J.; Zunszain, P.A.; Petitpas, I.; Bhattacharya, A.A.; Otagiri, M.; Curry, S. Structural basis of the drug-binding specificity of human serum albumin. J. Mol. Biol. 2005, 353, 38–52. [Google Scholar] [CrossRef] [PubMed]
- Zunszain, P.A.; Ghuman, J.; McDonagh, A.F.; Curry, S. Crystallographic analysis of human serum albumin complexed with 4Z,15E-bilirubin-IXalpha. J. Mol. Biol. 2008, 381, 394–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petitpas, I.; Grüne, T.; Bhattacharya, A.A.; Curry, S. Crystal structures of human serum albumin complexed with monounsaturated and polyunsaturated fatty acids. J. Mol. Biol. 2001, 314, 955–960. [Google Scholar] [CrossRef] [PubMed]
- Goncharov, N.V.; Belinskaia, D.A.; Shmurak, V.I.; Terpilowski, M.A.; Jenkins, R.O.; Avdonin, P.V. Serum albumin binding and esterase cctivity: Mechanistic interactions with organophosphates. Molecules 2017, 22, 1201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belinskaia, D.A.; Terpilovskii, M.A.; Batalova, A.A.; Goncharov, N.V. Effect of Cys34 oxidation state of albumin on its interaction with paraoxon according to molecular modeling data. Russ. J. Bioorg. Chem. 2019, 45, 535–544. [Google Scholar] [CrossRef]
- Sakurai, Y.; Ma, S.F.; Watanabe, H.; Yamaotsu, N.; Hirono, S.; Kurono, Y.; Kragh-Hansen, U.; Otagiri, M. Esterase-like activity of serum albumin: Characterization of its structural chemistry using p-nitrophenyl esters as substrates. Pharm. Res. 2004, 21, 285–292. [Google Scholar] [CrossRef]
- Grigoryan, H.; Li, H.; Iavarone, A.T.; Williams, E.R.; Rappaport, S.M. Cys34 adducts of reactive oxygen species in human serum albumin. Chem. Res. Toxicol. 2012, 25, 1633–1642. [Google Scholar] [CrossRef] [Green Version]
- Oettl, K.; Stauber, R.E. Physiological and pathological changes in the redox state of human serum albumin critically influence its binding properties. Br. J. Pharmacol. 2007, 151, 580–590. [Google Scholar] [CrossRef]
- Nagumo, K.; Tanaka, M.; Chuang, V.T.; Setoyama, H.; Watanabe, H.; Yamada, N.; Kubota, K.; Tanaka, M.; Matsushita, K.; Yoshida, A.; et al. Cys34-cysteinylated human serum albumin is a sensitive plasma marker in oxidative stress-related chronic diseases. PLoS ONE 2014, 9, 85216. [Google Scholar] [CrossRef]
- Suzuki, Y.; Suda, K.; Matsuyama, Y.; Era, S.; Soejima, A. Close Relationship between redox Ssate of human serum albumin and serum cysteine levels in non-diabetic CKD patients with various degrees of renal function. Clin. Nephrol. 2014, 82, 320–325. [Google Scholar] [CrossRef]
- Spanidis, Y.; Priftis, A.; Stagos, D.; Stravodimos, G.A.; Leonidas, D.D.; Spandidos, D.A.; Tsatsakis, A.M.; Kouretas, D. Oxidation of human serum albumin exhibits inter-individual variability after an ultra-marathon mountain race. Exp. Ther. Med. 2017, 13, 2382–2390. [Google Scholar] [CrossRef] [Green Version]
- Bar-Or, D.; Bar-Or, R.; Rael, L.T.; Gardner, D.K.; Slone, D.S.; Craun, M.L. Heterogeneity and oxidation status of commercial human albumin preparations in clinical use. Crit. Care Med. 2005, 33, 1638–1641. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, T.; Terada, T.; Arikawa, H.; Kizaki, K.; Terawaki, H.; Imai, H.; Itoh, Y.; Era, S. Quantitation of oxidative modifications of commercial human albumin for clinical use. Biol. Pharm. Bull. 2016, 39, 401–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakae, H.; Tomida, K.; Kikuya, Y.; Okuyama, M.; Igarashi, T. Comparison of quality of human serum albumin preparations in two pharmaceutical products. Acute Med. Surg. 2017, 4, 251–254. [Google Scholar] [CrossRef] [Green Version]
- Yasuhara, T.; Nokihara, K. Quantitative determination of biological sulfhydryl groups by postcolumn derivatization and elucidation of microheterogeneity of serum albumins. Anal. Chem. 1998, 70, 3505–3509. [Google Scholar] [CrossRef] [PubMed]
- Anraku, M.; Yamasaki, K.; Maruyama, T.; Kragh-Hansen, U.; Otagiri, M. Effect of oxidative stress on the structure and function of human serum albumin. Pharm. Res. 2001, 18, 632–639. [Google Scholar] [CrossRef]
- Sakurama, K.; Nishi, K.; Chuang, V.T.G.; Hashimoto, M.; Yamasaki, K.; Otagiri, M. Effects of oxidation of human serum albumin on the binding of aripiprazole. Biol. Pharm. Bull. 2020, 43, 1023–1026. [Google Scholar] [CrossRef]
- Buller, A.R.; Townsend, C.A. Intrinsic evolutionary constraints on protease structure, enzyme acylation, and the identity of the catalytic triad. Proc. Natl. Acad. Sci. USA 2013, 110, E653–E661. [Google Scholar] [CrossRef] [Green Version]
- Stryer, L.; Berg, J.M.; Tymoczko, J.L. Biochemistry, 6th ed.; W.H.Freeman & Co: New York, NY, USA, 2006. [Google Scholar]
- Zhang, Y.; Kua, J.; McCammon, J.A. Role of the catalytic triad and oxyanion hole in acetylcholinesterase catalysis: An ab initio QM/MM study. J. Am. Chem. Soc. 2002, 124, 10572–10577. [Google Scholar] [CrossRef]
- Hofer, P.; Fringeli, U.P. Acetylcholinesterase kinetics. Biophys. Struct. Mech. 1981, 8, 45–59. [Google Scholar] [CrossRef]
- Ekici, O.D.; Paetzel, M.; Dalbey, R.E. Unconventional serine proteases: Variations on the catalytic Ser/His/Asp triad configuration. Protein Sci. 2008, 17, 2023–2037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liyasova, M.S.; Schopfer, L.M.; Lockridge, O. Reaction of human albumin with aspirin in vitro: Mass spectrometric identification of acetylated lysines 199, 402, 519, and 545. Biochem. Pharmacol. 2010, 79, 784–791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phuangsawai, O.; Hannongbua, S.; Gleeson, M.P. Elucidating the origin of the esterase activity of human serum albumin using QM/MM calculations. J. Phys. Chem. B 2014, 118, 11886–11894. [Google Scholar] [CrossRef] [PubMed]
- Honma, K.; Nakamura, M.; Ishikawa, Y. Acetylsalicylate-human serum albumin interaction as studied by NMR spectroscopy--antigenicity-producing mechanism of acetylsalicylic acid. Mol. Immunol. 1991, 28, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Dahiya, V.; Anand, B.G.; Kar, K.; Pal, S. In vitro interaction of organophosphate metabolites with bovine serum albumin: A comparative 1H NMR, fluorescence and molecular docking analysis. Pestic. Biochem. Physiol. 2020, 163, 39–50. [Google Scholar] [CrossRef] [PubMed]
- Dahiya, V.; Anand, B.G.; Kar, K.; Pal, S. Analyzing organophosphate pesticide-serum albumin binding interaction: A combined STD NMR and molecular docking study. J. Biomol. Struct. Dyn. 2021, 39, 1865–1878. [Google Scholar] [CrossRef]
- Silva, D.; Cortez, C.M.; Cunha-Bastos, J.; Louro, S.R. Methyl parathion interaction with human and bovine serum albumin. Toxicol. Lett. 2004, 147, 53–61. [Google Scholar] [CrossRef]
- Zhao, H.; Bojko, B.; Liu, F.; Pawliszyn, J.; Peng, W.; Wang, X. Mechanism of interactions between organophosphorus insecticides and human serum albumin: Solid-phase microextraction, thermodynamics and computational approach. Chemosphere 2020, 253, 126698. [Google Scholar] [CrossRef]
- Lyagin, I.; Efremenko, E. Enzymes, Reacting with Organophosphorus Compounds as Detoxifiers: Diversity and Functions. Int. J. Mol. Sci. 2021, 22, 1761. [Google Scholar] [CrossRef]
- Pen, J.; Beintema, J.J. Nomenclature of esterases. Biochem. J. 1986, 240, 933. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Wang, S.; Huang, M. Structure and enzymatic activities of human serum albumin. Curr. Pharm. Des. 2015, 21, 1831–1836. [Google Scholar] [CrossRef] [PubMed]
- 4-Nitrophenol. Available online: http://www.biosite.dk/leksikon/4-nitrophenol.htm (accessed on 22 September 2021).
- Froimowitz, M. HyperChem: A software package for computational chemistry and molecular modeling. Biotechniques 1993, 14, 1010–1013. [Google Scholar] [PubMed]
- Ramachandran, S.; Kota, P.; Ding, F.; Dokholyan, N.V. Automated minimization of steric clashes in protein structures. Proteins 2011, 79, 261–270. [Google Scholar] [CrossRef] [Green Version]
- Abraham, M.J.; Murtola, T.; Schulz, R.; Páll, S.; Smith, J.C.; Hess, B.; Lindahl, E. GROMACS: High performance molecular simulations through multi-level parallelism from laptops to supercomputers. SoftwareX 2015, 1–2, 19–25. [Google Scholar] [CrossRef] [Green Version]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lyskov, S.; Chou, F.C.; Conchúir, S.Ó.; Der, B.S.; Drew, K.; Kuroda, D.; Xu, J.; Weitzner, B.D.; Renfrew, P.D.; Sripakdeevong, P.; et al. Serverification of molecular modeling applications: The Rosetta Online Server that Includes Everyone (ROSIE). PLoS ONE 2013, 8, e63906. [Google Scholar] [CrossRef] [Green Version]
- Berendsen, H.J.C.; Postma, J.P.M.; van Gunsteren, W.F.; Hermans, J. Interaction models for water in relation to protein hydration. In Intermolecular Forces. The Jerusalem Symposia on Quantum Chemistry and Biochemistry; Pullman, B., Ed.; Springer: Dordrecht, The Netherlands, 1981; Volume 14, pp. 331–342. [Google Scholar]
- Bussi, G.; Zykova-Timan, T.; Parrinello, M. Isothermal-isobaric molecular dynamics using stochastic velocity rescaling. J. Chem. Phys. 2009, 130, 074101. [Google Scholar] [CrossRef] [Green Version]
- Berendsen, H.J.C.; Postma, J.P.M.; di Nola, A.; van Gunsteren, W.F.; Haak, J.R. Molecular dynamics with coupling to an external bath. J. Chem. Phys. 1984, 81, 3684–3690. [Google Scholar] [CrossRef] [Green Version]
- Darden, T.; York, D.; Pedersen, L. Particle mesh Ewald: An N⋅log(N) method for Ewald sums in large systems. J. Chem. Phys. 1993, 3, 10089–10092. [Google Scholar] [CrossRef] [Green Version]
- Hess, B.; Bekker, H.; Berendsen, H.J.C.; Fraaije, J.G.E.M. LINCS: A linear constraint solver for molecular simulations. J. Comp. Chem. 1997, 18, 1463–1473. [Google Scholar] [CrossRef]













Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Belinskaia, D.A.; Voronina, P.A.; Vovk, M.A.; Shmurak, V.I.; Batalova, A.A.; Jenkins, R.O.; Goncharov, N.V. Esterase Activity of Serum Albumin Studied by 1H NMR Spectroscopy and Molecular Modelling. Int. J. Mol. Sci. 2021, 22, 10593. https://doi.org/10.3390/ijms221910593
Belinskaia DA, Voronina PA, Vovk MA, Shmurak VI, Batalova AA, Jenkins RO, Goncharov NV. Esterase Activity of Serum Albumin Studied by 1H NMR Spectroscopy and Molecular Modelling. International Journal of Molecular Sciences. 2021; 22(19):10593. https://doi.org/10.3390/ijms221910593
Chicago/Turabian StyleBelinskaia, Daria A., Polina A. Voronina, Mikhail A. Vovk, Vladimir I. Shmurak, Anastasia A. Batalova, Richard O. Jenkins, and Nikolay V. Goncharov. 2021. "Esterase Activity of Serum Albumin Studied by 1H NMR Spectroscopy and Molecular Modelling" International Journal of Molecular Sciences 22, no. 19: 10593. https://doi.org/10.3390/ijms221910593