
An Engineered Thermostable Laccase with Great Ability to Decolorize and Detoxify Malachite Green

 ,
,
Abstract
:1. Introduction
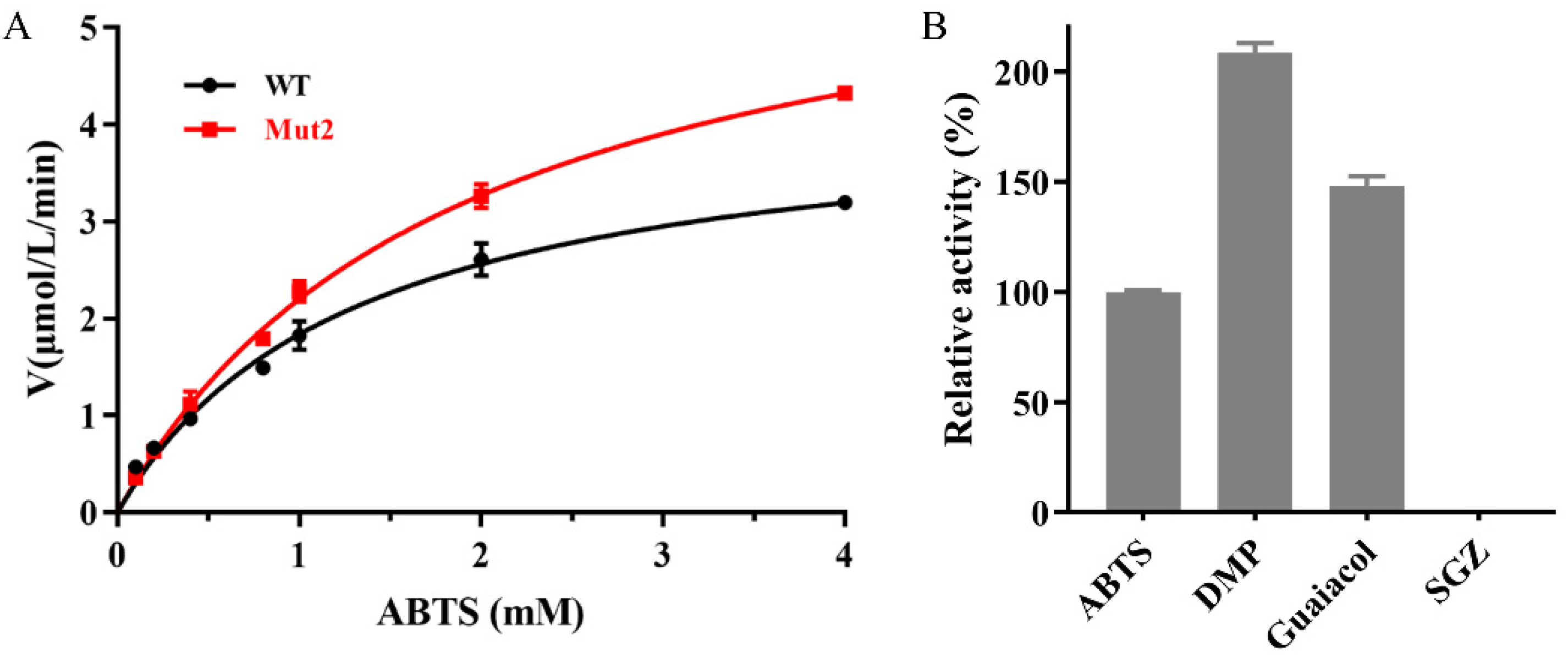
2. Results and Discussion
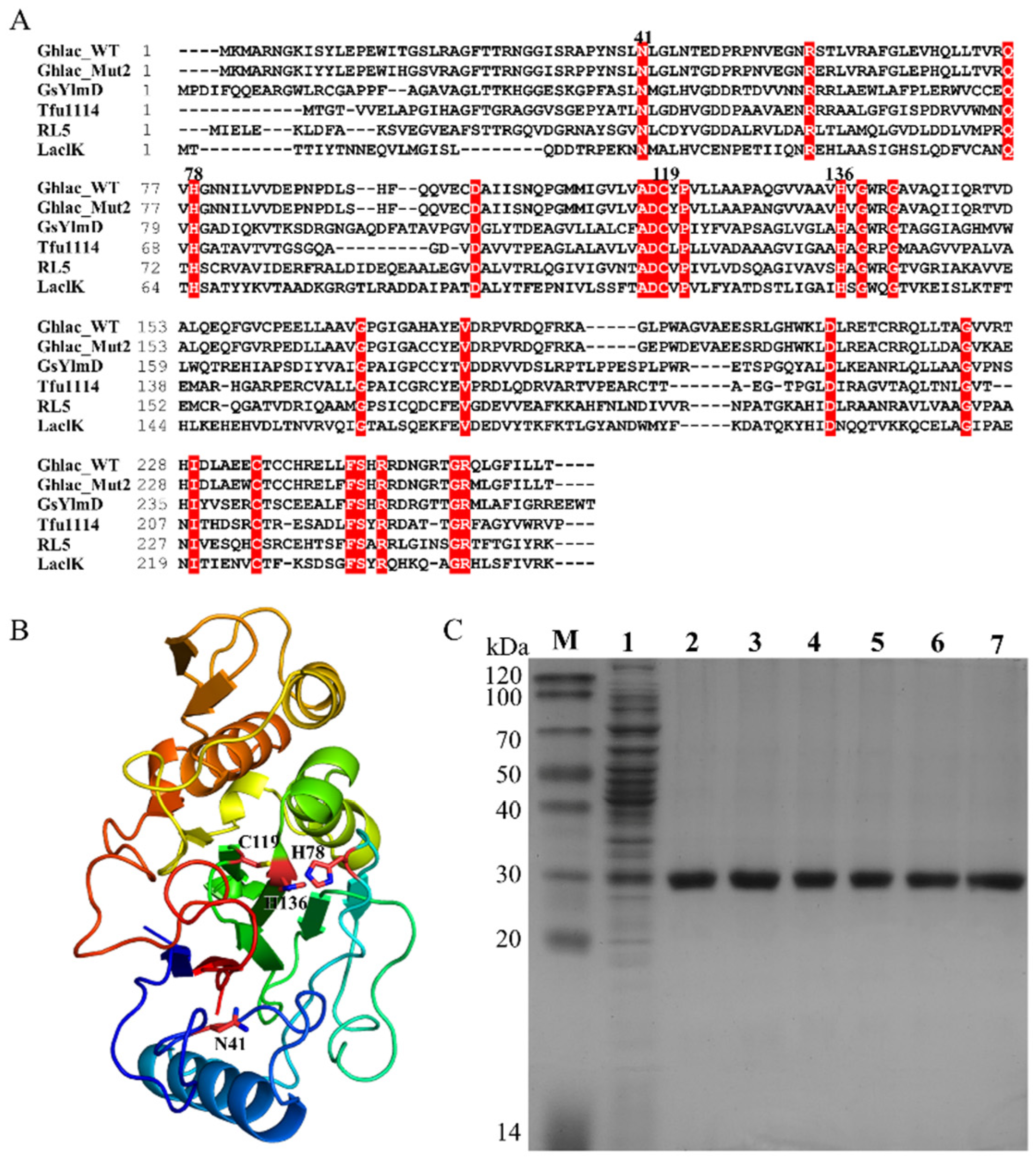
2.1. Sequence Analysis, Expression, Purification, and Mutation of Ghlac
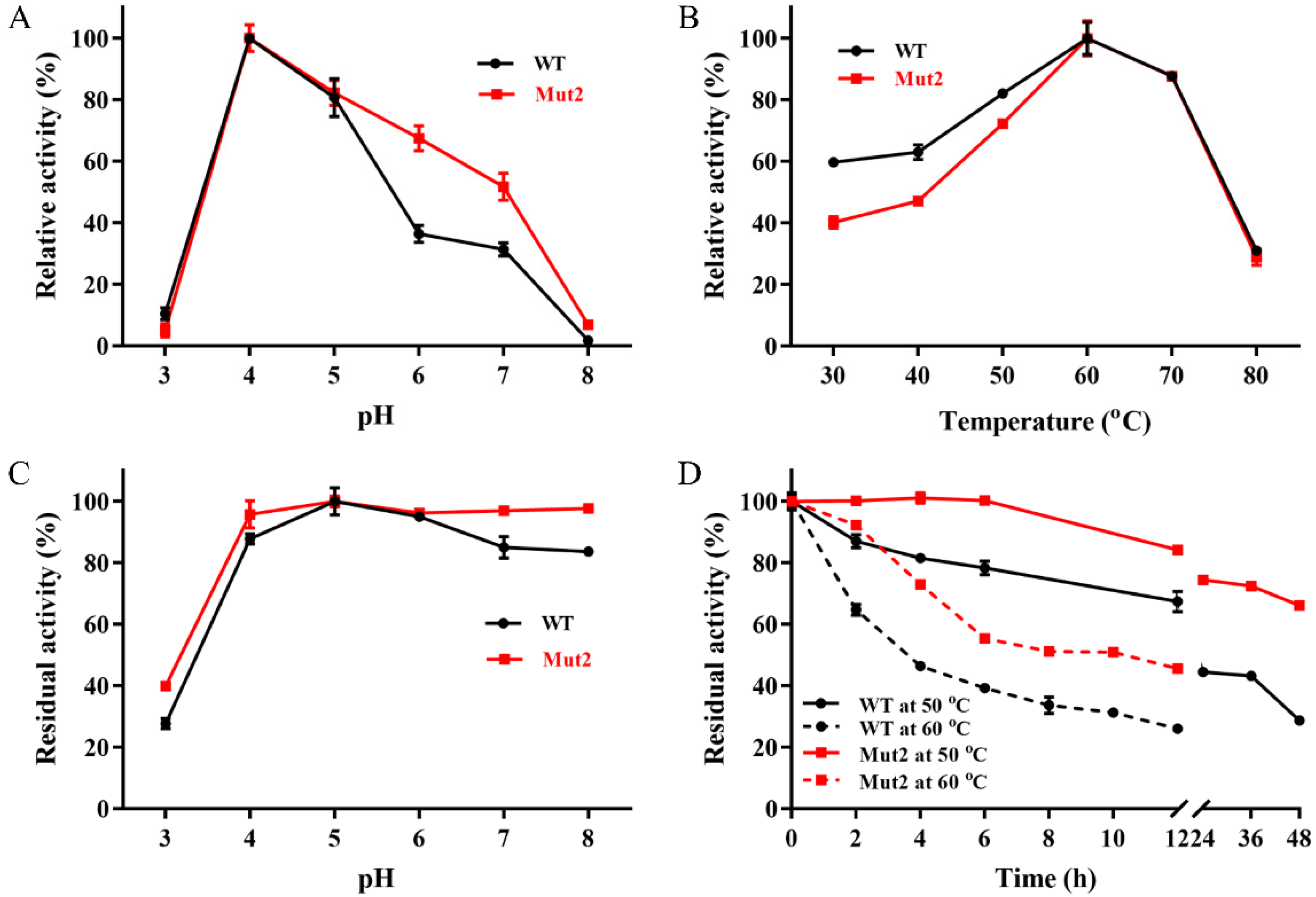
2.2. Effects of pH and Temperature on the Activity and Stability of Ghlac
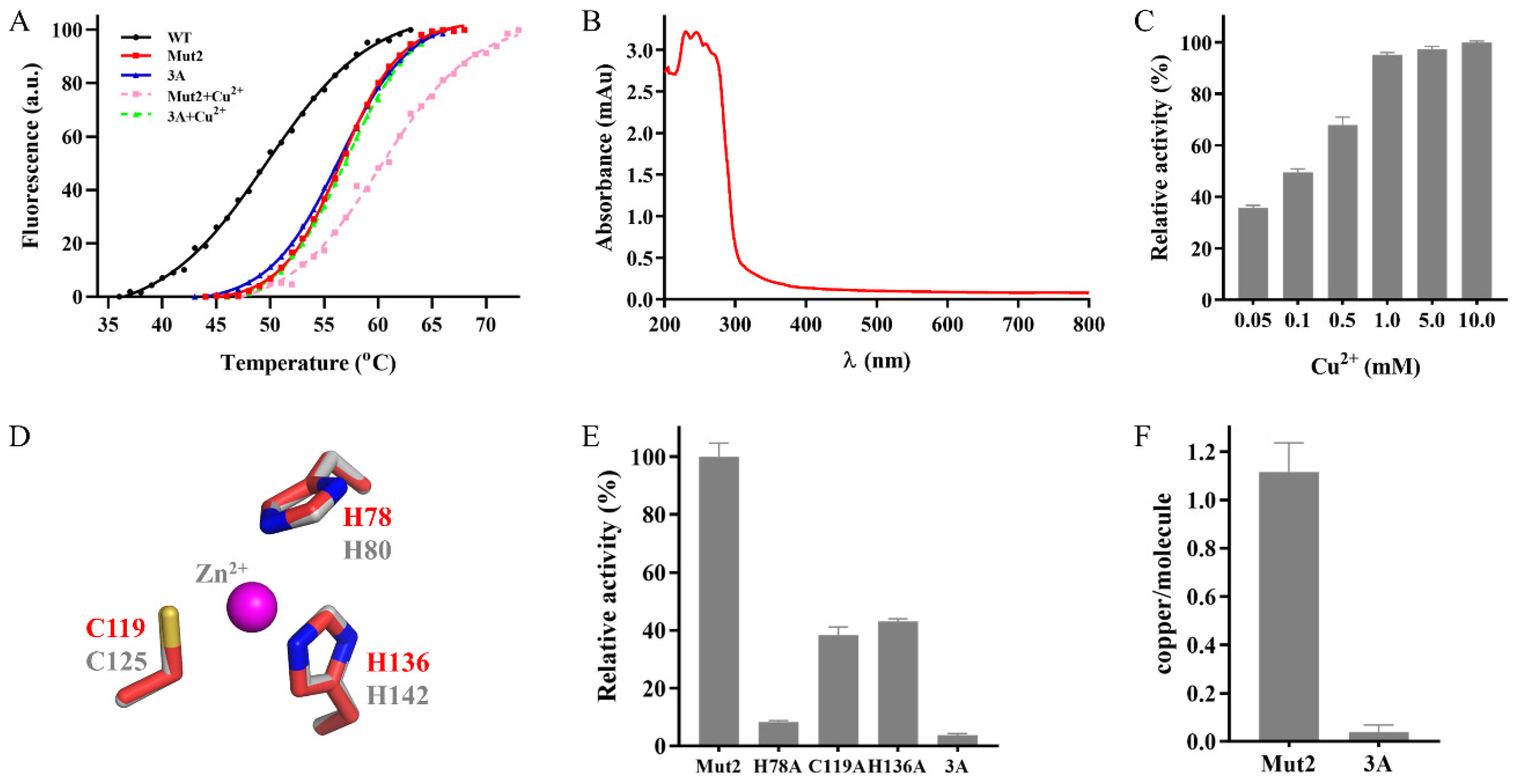
2.3. Identification of the Putative Copper Binding Site
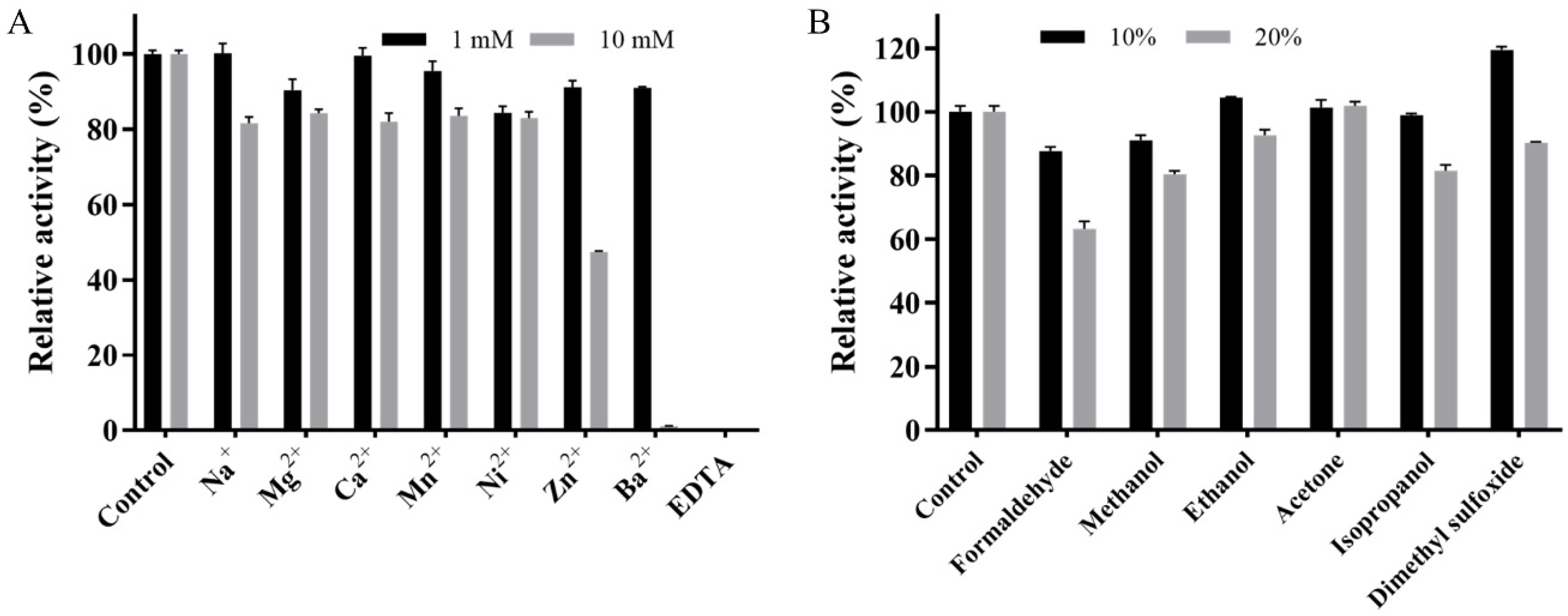
2.4. Effects of Metal Ions and Organic Solvents on the Activity of Mut2
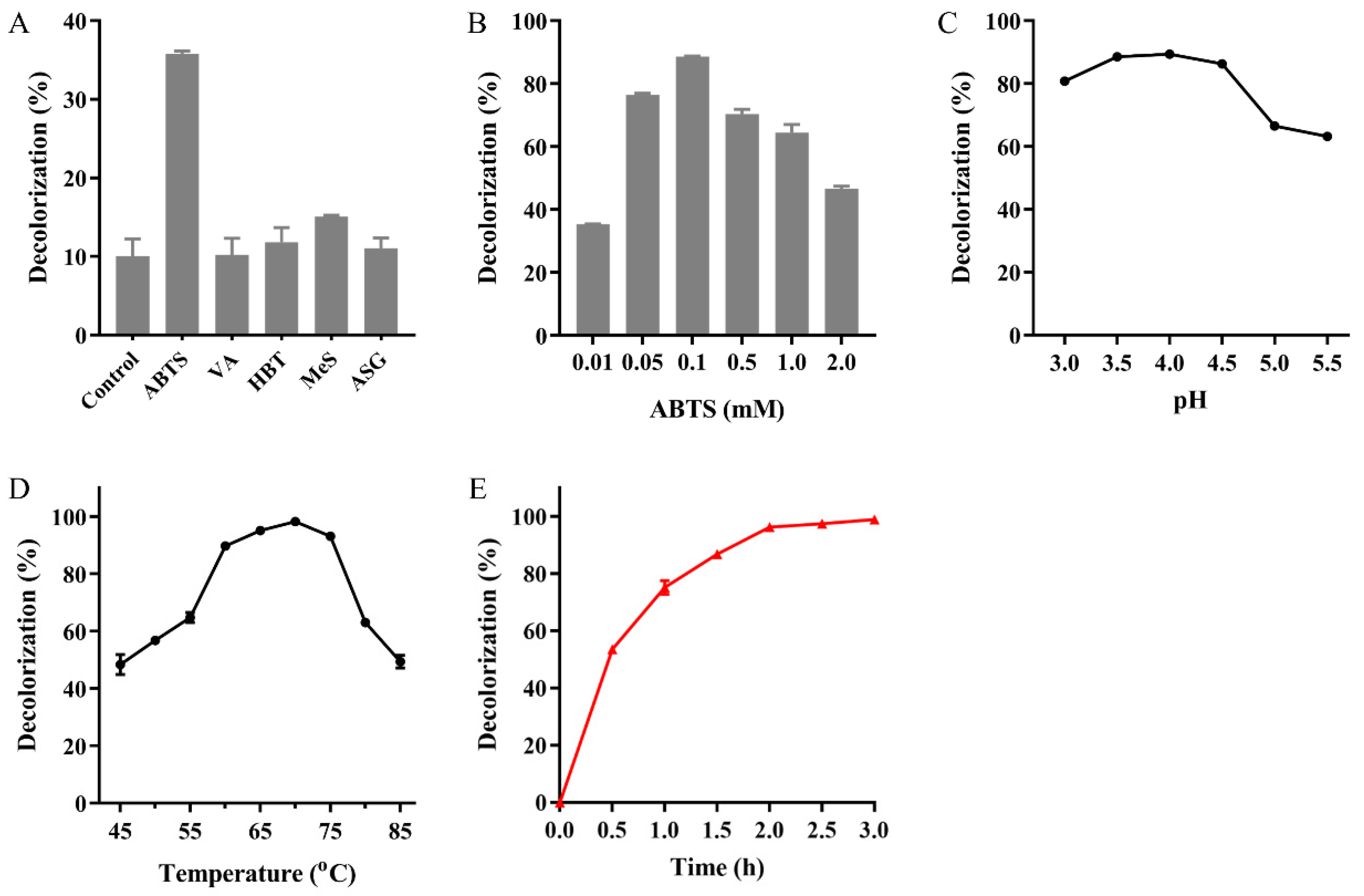
2.5. MG Decolorization Catalyzed by Mut2
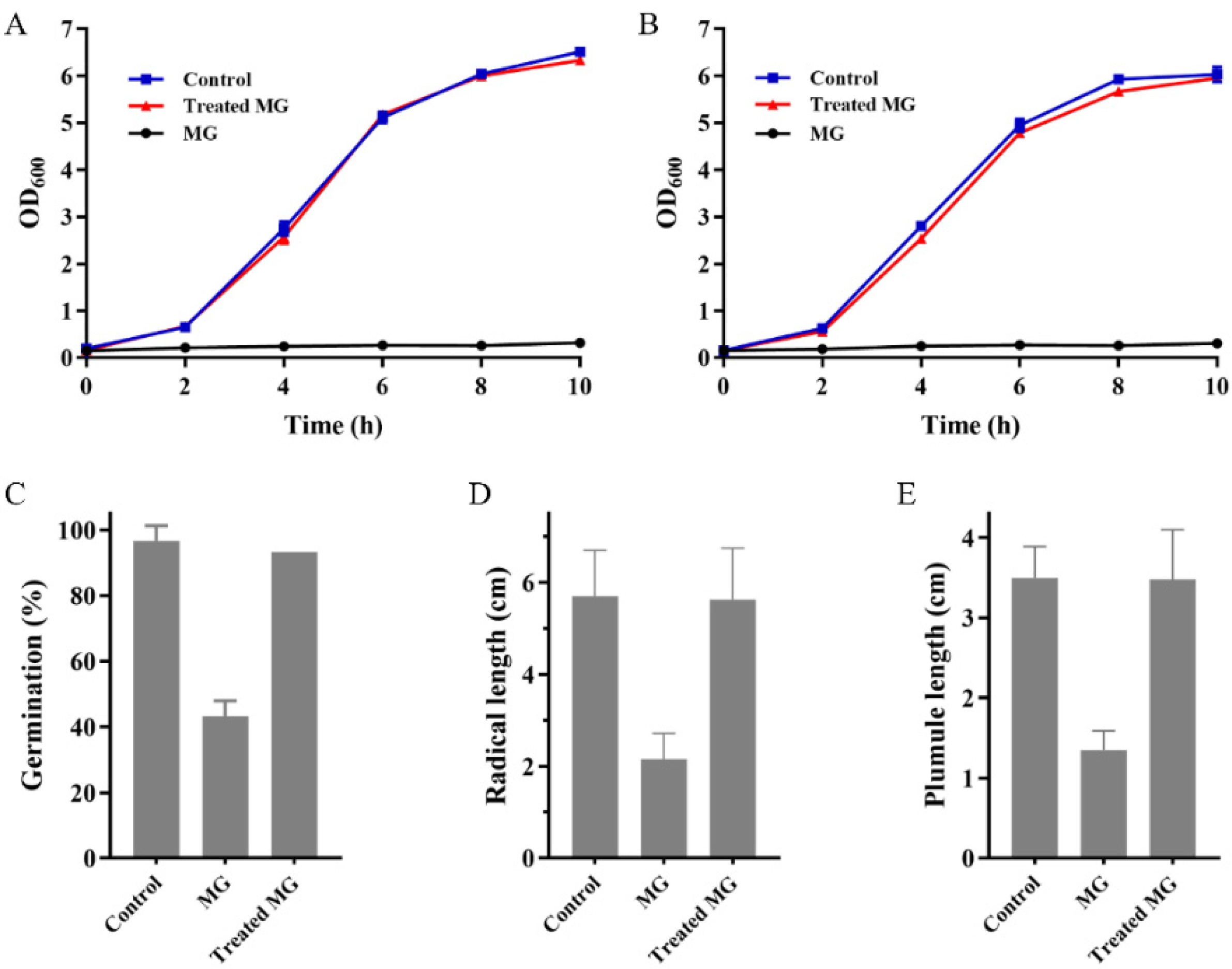
2.6. MG Detoxification Catalyzed by Mut2
3. Materials and Methods
3.1. Materials
3.2. Cloning, Expression, and Purification of Ghlac
3.3. Mutation Design Using PROSS and Site-Directed Mutagenesis
3.4. Enzyme Activity Assay
3.5. Effects of Temperature and pH on the Laccase Activity and Stability
3.6. Effects of Metal Ions and Organic Solvents on the Activity of Mut2
3.7. MG Decolorization Catalyzed by Mut2
3.8. Toxicity Tests
3.9. Sequence Analysis
3.10. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Janusz, G.; Pawlik, A.; Świderska-Burek, U.; Polak, J.; Sulej, J.; Jarosz-Wilkołazka, A.; Paszczyński, A. Laccase properties, physiological functions, and evolution. Int. J. Mol. Sci. 2020, 21, 966. [Google Scholar] [CrossRef] [Green Version]
- Chauhan, P.S.; Goradia, B.; Saxena, A. Bacterial laccase: Recent update on production, properties and industrial applications. 3 Biotech 2017, 7, 323. [Google Scholar] [CrossRef]
- Kumar, V.; Patel, S.K.S.; Gupta, R.K.; Otari, S.V.; Gao, H.; Lee, J.-K.; Zhang, L. Enhanced saccharification and fermentation of rice straw by reducing the concentration of phenolic compounds using an immobilized enzyme cocktail. Biotechnol. J. 2019, 14, 1800468. [Google Scholar] [CrossRef] [PubMed]
- Singh, G.; Arya, S.K. Utility of laccase in pulp and paper industry: A progressive step towards the green technology. Int. J. Biol. Macromol. 2019, 134, 1070–1084. [Google Scholar] [CrossRef] [PubMed]
- Guan, Z.-B.; Luo, Q.; Wang, H.-R.; Chen, Y.; Liao, X.-R. Bacterial laccases: Promising biological green tools for industrial applications. Cell. Mol. Life Sci. 2018, 75, 3569–3592. [Google Scholar] [CrossRef]
- Wiśniewska, K.M.; Twarda-Clapa, A.; Białkowska, A.M. Novel cold-adapted recombinant laccase KbLcc1 from Kabatiella bupleuri G3 IBMiP as a green catalyst in biotransformation. Int. J. Mol. Sci. 2021, 22, 9593. [Google Scholar] [CrossRef]
- Wikee, S.; Hatton, J.; Turbé-Doan, A.; Mathieu, Y.; Daou, M.; Lomascolo, A.; Kumar, A.; Lumyong, S.; Sciara, G.; Faulds, C.B.; et al. Characterization and dye decolorization potential of two laccases from the marine-derived fungus Pestalotiopsis sp. Int. J. Mol. Sci. 2019, 20, 1864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, S.K.S.; Gupta, R.K.; Kim, S.-Y.; Kim, I.-W.; Kalia, V.C.; Lee, J.-K. Rhus vernicifera laccase immobilization on magnetic nanoparticles to improve stability and its potential application in bisphenol A degradation. Indian J. Microbiol. 2021, 61, 45–54. [Google Scholar] [CrossRef]
- Agrawal, K.; Chaturvedi, V.; Verma, P. Fungal laccase discovered but yet undiscovered. Bioresour. Bioprocess. 2018, 5, 4. [Google Scholar] [CrossRef]
- Tonin, F.; Melis, R.; Cordes, A.; Sanchez-Amat, A.; Pollegioni, L.; Rosini, E. Comparison of different microbial laccases as tools for industrial uses. New Biotechnol. 2016, 33, 387–398. [Google Scholar] [CrossRef]
- Cañas, A.I.; Camarero, S. Laccases and their natural mediators: Biotechnological tools for sustainable eco-friendly processes. Biotechnol. Adv. 2010, 28, 694–705. [Google Scholar] [CrossRef]
- Gu, Y.; Yuan, L.; Jia, L.; Xue, P.; Yao, H. Recent developments of a co-immobilized laccase–mediator system: A review. RSC Adv. 2021, 11, 29498–29506. [Google Scholar] [CrossRef]
- Raval, N.P.; Shah, P.U.; Shah, N.K. Malachite green “a cationic dye” and its removal from aqueous solution by adsorption. Appl. Water Sci. 2017, 7, 3407–3445. [Google Scholar] [CrossRef] [Green Version]
- Kooravand, M.; Asadpour, S.; Haddadi, H.; Farhadian, S. An insight into the interaction between malachite green oxalate with human serum albumin: Molecular dynamic simulation and spectroscopic approaches. J. Hazard. Mater. 2021, 407, 124878. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Yang, X.; Lin, Y.; Ng, T.B.; Lin, J.; Ye, X. Laccase-catalyzed decolorization of malachite green: Performance optimization and degradation mechanism. PLoS ONE 2015, 10, e0127714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shanmugam, S.; Ulaganathan, P.; Swaminathan, K.; Sadhasivam, S.; Wu, Y.-R. Enhanced biodegradation and detoxification of malachite green by Trichoderma asperellum laccase: Degradation pathway and product analysis. Int. Biodeter. Biodegr. 2017, 125, 258–268. [Google Scholar] [CrossRef]
- Luo, Q.; Chen, Y.; Xia, J.; Wang, K.-Q.; Cai, Y.-J.; Liao, X.-R.; Guan, Z.-B. Functional expression enhancement of Bacillus pumilus CotA-laccase mutant WLF through site-directed mutagenesis. Enzyme Microb. Technol. 2018, 109, 11–19. [Google Scholar] [CrossRef]
- Sharma, N.; Leung, I.K.H. Characterization and optimisation of a novel laccase from Sulfitobacter indolifex for the decolourisation of organic dyes. Int. J. Biol. Macromol. 2021, 190, 574–584. [Google Scholar] [CrossRef] [PubMed]
- Espina, G.; Cáceres-Moreno, P.; Mejías-Navarrete, G.; Ji, M.; Sun, J.; Blamey, J.M. A novel and highly active recombinant spore-coat bacterial laccase, able to rapidly biodecolorize azo, triarylmethane and anthraquinonic dyestuffs. Int. J. Biol. Macromol. 2021, 170, 298–306. [Google Scholar] [CrossRef]
- Hossain, L.; Sarker, S.K.; Khan, M.S. Evaluation of present and future wastewater impacts of textile dyeing industries in Bangladesh. Environ. Dev. 2018, 26, 23–33. [Google Scholar] [CrossRef]
- Mojtabavi, S.; Khoshayand, M.R.; Fazeli, M.R.; Samadi, N.; Faramarzi, M.A. Combination of thermal and biological treatments for bio-removal and detoxification of some recalcitrant synthetic dyes by betaine-induced thermostabilized laccase. Environ. Technol. Inno. 2020, 20, 101046. [Google Scholar] [CrossRef]
- Beloqui, A.; Pita, M.; Polaina, J.; Martínez-Arias, A.; Golyshina, O.V.; Zumárraga, M.; Yakimov, M.M.; García-Arellano, H.; Alcalde, M.; Fernández, V.M. Novel polyphenol oxidase mined from a metagenome expression library of bovine rumen: Biochemical properties, structural analysis, and phylogenetic relationships. J. Biol. Chem. 2006, 281, 22933–22942. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.-Y.; Hsieh, Z.-S.; Cheepudom, J.; Yang, C.-H.; Meng, M. A 24.7-kDa copper-containing oxidase, secreted by Thermobifida fusca, significantly increasing the xylanase/cellulase-catalyzed hydrolysis of sugarcane bagasse. Appl. Microbiol. Biotechnol. 2013, 97, 8977–8986. [Google Scholar] [CrossRef]
- Guo, X.; Zhou, S.; Wang, Y.; Song, J.; Wang, H.; Kong, D.; Zhu, J.; Dong, W.; He, M.; Hu, G. Characterization of a highly thermostable and organic solvent-tolerant copper-containing polyphenol oxidase with dye-decolorizing ability from Kurthia huakuii LAM0618T. PLoS ONE 2016, 11, e0164810. [Google Scholar] [CrossRef]
- Davidi, L.; Moraïs, S.; Artzi, L.; Knop, D.; Hadar, Y.; Arfi, Y.; Bayer, E.A. Toward combined delignification and saccharification of wheat straw by a laccase-containing designer cellulosome. Proc. Natl. Acad. Sci. USA 2016, 113, 10854–10859. [Google Scholar] [CrossRef] [Green Version]
- Goldenzweig, A.; Goldsmith, M.; Hill, S.E.; Gertman, O.; Laurino, P.; Ashani, Y.; Dym, O.; Unger, T.; Albeck, S.; Prilusky, J.; et al. Automated structure- and sequence-based design of proteins for high bacterial expression and stability. Mol. Cell 2016, 63, 337–346. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Y.; Li, J.; Wu, P.; Ning, N.; Li, J.; Shen, Y.; Huang, Q.; Ni, J. An alkaline thermostable laccase from termite gut associated strain of Bacillus stratosphericus. Int. J. Biol. Macromol. 2021, 179, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Singh, G.; Capalash, N.; Goel, R.; Sharma, P.J.E.; Technology, M. A pH-stable laccase from alkali-tolerant γ-proteobacterium JB: Purification, characterization and indigo carmine degradation. Enzyme Microb. Technol. 2007, 41, 794–799. [Google Scholar] [CrossRef]
- Fang, Z.-M.; Li, T.-L.; Chang, F.; Zhou, P.; Fang, W.; Hong, Y.-Z.; Zhang, X.-C.; Peng, H.; Xiao, Y.-Z. A new marine bacterial laccase with chloride-enhancing, alkaline-dependent activity and dye decolorization ability. Bioresour. Technol. 2012, 111, 36–41. [Google Scholar] [CrossRef]
- Yang, X.; Wu, Y.; Zhang, Y.; Yang, E.; Qu, Y.; Xu, H.; Chen, Y.; Irbis, C.; Yan, J. A thermo-active laccase isoenzyme from Trametes trogii and its potential for dye decolorization at high temperature. Front. Microbiol. 2020, 11, 241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Huang, L.; Guo, W.; Jia, L.; Fu, Y.; Gui, S.; Lu, F. Cloning, expression, and characterization of a thermostable and pH-stable laccase from Klebsiella pneumoniae and its application to dye decolorization. Process. Biochem. 2017, 53, 125–134. [Google Scholar] [CrossRef]
- Campeotto, I.; Goldenzweig, A.; Davey, J.; Barfod, L.; Marshall, J.M.; Silk, S.E.; Wright, K.E.; Draper, S.J.; Higgins, M.K.; Fleishman, S.J. One-step design of a stable variant of the malaria invasion protein RH5 for use as a vaccine immunogen. Proc. Natl. Acad. Sci. USA 2017, 114, 998–1002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kriegel, M.; Wiederanders, H.J.; Alkhashrom, S.; Eichler, J.; Muller, Y.A. A PROSS-designed extensively mutated estrogen receptor α variant displays enhanced thermal stability while retaining native allosteric regulation and structure. Sci. Rep. 2021, 11, 10509. [Google Scholar] [CrossRef]
- Berini, F.; Verce, M.; Ausec, L.; Rosini, E.; Tonin, F.; Pollegioni, L.; Mandić-Mulec, I. Isolation and characterization of a heterologously expressed bacterial laccase from the anaerobe Geobacter metallireducens. Appl. Microbiol. Biotechnol. 2018, 102, 2425–2439. [Google Scholar] [CrossRef]
- Mao, G.; Zhao, Y.; Kang, X.; Li, Z.; Zhang, Y.; Wang, X.; Sun, F.; Sankaran, K.; Zhang, X.C. Crystal structure of E. coli lipoprotein diacylglyceryl transferase. Nat. Commun. 2016, 7, 10198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forootanfar, H.; Faramarzi, M.A.; Shahverdi, A.R.; Yazdi, M.T. Purification and biochemical characterization of extracellular laccase from the ascomycete Paraconiothyrium variabile. Bioresour. Technol. 2011, 102, 1808–1814. [Google Scholar] [CrossRef]
- Sondhi, S.; Sharma, P.; Saini, S.; Puri, N.; Gupta, N. Purification and characterization of an extracellular, thermo-alkali-stable, metal tolerant laccase from Bacillus tequilensis SN4. PLoS ONE 2014, 9, e96951. [Google Scholar] [CrossRef] [Green Version]
- Sondhi, S.; Kaur, R.; Madan, J. Purification and characterization of a novel white highly thermo stable laccase from a novel Bacillus sp. MSK-01 having potential to be used as anticancer agent. Int. J. Biol. Macromol. 2021, 170, 232–238. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.-H.; Lin, M.-C.; Lee, C.-C.; Yu, S.-M.; Wang, A.H.J.; Ho, T.-H.D. Enhancement of laccase activity by pre-incubation with organic solvents. Sci. Rep. 2019, 9, 9754. [Google Scholar] [CrossRef]
- Yang, Q.; Zhang, M.; Zhang, M.; Wang, C.; Liu, Y.; Fan, X.; Li, H. Characterization of a novel, cold-adapted, and thermostable laccase-like enzyme with high tolerance for organic solvents and salt and potent dye decolorization ability, derived from a marine metagenomic library. Front. Microbiol. 2018, 9, 2998. [Google Scholar] [CrossRef] [Green Version]
- Olajuyigbe, F.M.; Fatokun, C.O. Biochemical characterization of an extremely stable pH-versatile laccase from Sporothrix carnis CPF-05. Int. J. Biol. Macromol. 2017, 94, 535–543. [Google Scholar] [CrossRef] [PubMed]
- Fang, Z.; Liu, X.; Chen, L.; Shen, Y.; Zhang, X.; Fang, W.; Wang, X.; Bao, X.; Xiao, Y. Identification of a laccase Glac15 from Ganoderma lucidum 77002 and its application in bioethanol production. Biotechnol. Biofuels 2015, 8, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Bendary, M.A.; Ezzat, S.M.; Ewais, E.A.; Al-Zalama, M.A. Optimization of spore laccase production by Bacillus amyloliquefaciens isolated from wastewater and its potential in green biodecolorization of synthetic textile dyes. Prep. Biochem. Biotech. 2021, 51, 16–27. [Google Scholar] [CrossRef] [PubMed]
- Anwer, H.; Mahmood, A.; Lee, J.; Kim, K.-H.; Park, J.-W.; Yip, A.C.K. Photocatalysts for degradation of dyes in industrial effluents: Opportunities and challenges. Nano Res. 2019, 12, 955–972. [Google Scholar] [CrossRef]
- Bhatia, D.; Sharma, N.R.; Singh, J.; Kanwar, R.S. Biological methods for textile dye removal from wastewater: A review. Crit. Rev. Env. Sci. Tec 2017, 47, 1836–1876. [Google Scholar] [CrossRef]
- Wang, H.; Huang, L.; Li, Y.; Ma, J.; Wang, S.; Zhang, Y.; Ge, X.; Wang, N.; Lu, F.; Liu, Y. Characterization and application of a novel laccase derived from Bacillus amyloliquefaciens. Int. J. Biol. Macromol. 2020, 150, 982–990. [Google Scholar] [CrossRef]
- Joshi, M.; Mukherjee, A.K.; Thakur, B.D. Development of a new styrene copolymer membrane for recycling of polyester fibre dyeing effluent. J. Membr. Sci. 2001, 189, 23–40. [Google Scholar] [CrossRef]
- Li, C.; Zhang, Z.; Li, Y.; Cao, J. Study on dyeing wastewater treatment at high temperature by MBBR and the thermotolerant mechanism based on its microbial analysis. Process. Biochem. 2015, 50, 1934–1941. [Google Scholar] [CrossRef]
- Ma, X.; Liu, L.; Li, Q.; Liu, Y.; Yi, L.; Ma, L.; Zhai, C. High-level expression of a bacterial laccase, CueO from Escherichia coli K12 in Pichia pastoris GS115 and its application on the decolorization of synthetic dyes. Enzyme Microb. Technol. 2017, 103, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Mao, G.; Fan, H.; Song, A.; Zhang, Y.-H.P.; Chen, H. Biochemical properties of GH94 cellodextrin phosphorylase THA_1941 from a thermophilic eubacterium Thermosipho africanus TCF52B with cellobiose phosphorylase activity. Sci. Rep. 2017, 7, 4849. [Google Scholar] [CrossRef]
- Yang, X.; Zheng, J.; Lu, Y.; Jia, R. Degradation and detoxification of the triphenylmethane dye malachite green catalyzed by crude manganese peroxidase from Irpex lacteus F17. Environ. Sci. Pollut. Res. 2016, 23, 9585–9597. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.S.; Al-Tohamy, R.; Xie, R.; El-Sheekh, M.M.; Sun, J. Construction of a new lipase- and xylanase-producing oleaginous yeast consortium capable of reactive azo dye degradation and detoxification. Bioresour. Technol. 2020, 313, 123631. [Google Scholar] [CrossRef] [PubMed]
- Sievers, F.; Higgins, D.G. Clustal Omega for making accurate alignments of many protein sequences. Protein Sci. 2018, 27, 135–145. [Google Scholar] [CrossRef] [Green Version]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeLano, W.L. Pymol: An open-source molecular graphics tool. CCP4 Newsl. Protein Crystallogr. 2002, 40, 82–92. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Laccase | Source | t1/2 | MG Decolorization | |||||
|---|---|---|---|---|---|---|---|---|
| 50 °C | 60 °C | MG (mg/L) | Mediator | Temperature (°C) | Time (h) | Decolorization Rate | ||
| Ghlac Mut2 | G. hydrogeniphilus | 80.6 h | 9.8 h | 100 | 0.1 mM ABTS | 70 | 3 | >99% |
| 60 | >90% | |||||||
| CotA WLF [17] | B. pumilus | 6.5 h | ND | 50 | 1 mM ASG | 37 | 10 | >95% |
| pLacSi [18] | S. indolifex | Unstable a | Unstable b | 50 | 1 mM ABTS | 30 | overnight | >80 |
| FNTL [19] | Bacillus sp. | ND | About 2.7 h | 50 | 2 mM ASG | 40 | 0.5 | >99% |
| LaclK [24] | K. huakuii | ND | Highly stable c | 9 | 0.1 mM ABTS | 60 | 1 | <40% |
| BaCotA [27] | B. stratosphericus | 2 h | 1 h | 100 | 0.01 mM ABTS | 60 | 3 | 82% |
| rLac [31] | K. pneumoniae | Stable d | 5 h | 100 | 70 | 1.5 | 90% | |
| rLAC [46] | B. amyloliquefaciens | Stable e | Stable f | 100 | 0.1 mM ABTS | 60 | 6 | 95% |
| CueO-p [49] | E. coli | Stable g | 80 | 0.1 mM ASG | 55 | 12 | 98.5% | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mao, G.; Wang, K.; Wang, F.; Li, H.; Zhang, H.; Xie, H.; Wang, Z.; Wang, F.; Song, A. An Engineered Thermostable Laccase with Great Ability to Decolorize and Detoxify Malachite Green. Int. J. Mol. Sci. 2021, 22, 11755. https://doi.org/10.3390/ijms222111755
Mao G, Wang K, Wang F, Li H, Zhang H, Xie H, Wang Z, Wang F, Song A. An Engineered Thermostable Laccase with Great Ability to Decolorize and Detoxify Malachite Green. International Journal of Molecular Sciences. 2021; 22(21):11755. https://doi.org/10.3390/ijms222111755
Chicago/Turabian StyleMao, Guotao, Kai Wang, Fangyuan Wang, Hao Li, Hongsen Zhang, Hui Xie, Zhimin Wang, Fengqin Wang, and Andong Song. 2021. "An Engineered Thermostable Laccase with Great Ability to Decolorize and Detoxify Malachite Green" International Journal of Molecular Sciences 22, no. 21: 11755. https://doi.org/10.3390/ijms222111755