Triatoma rosai sp. nov. (Hemiptera, Triatominae): A New Species of Argentinian Chagas Disease Vector Described Based on Integrative Taxonomy

 ,
,  , , ,
, , ,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Morphological Identification and Description
2.3. Morphological Studies in Scanning Electron Microscopy
2.4. Morphological Studies in Optical Microscopy
2.5. Morphometric Studies
2.6. Molecular Phylogeny and Genetic Distance
2.7. Experimental Crosses
2.8. Distribution Map
3. Results
3.1. Systematics
Type Material
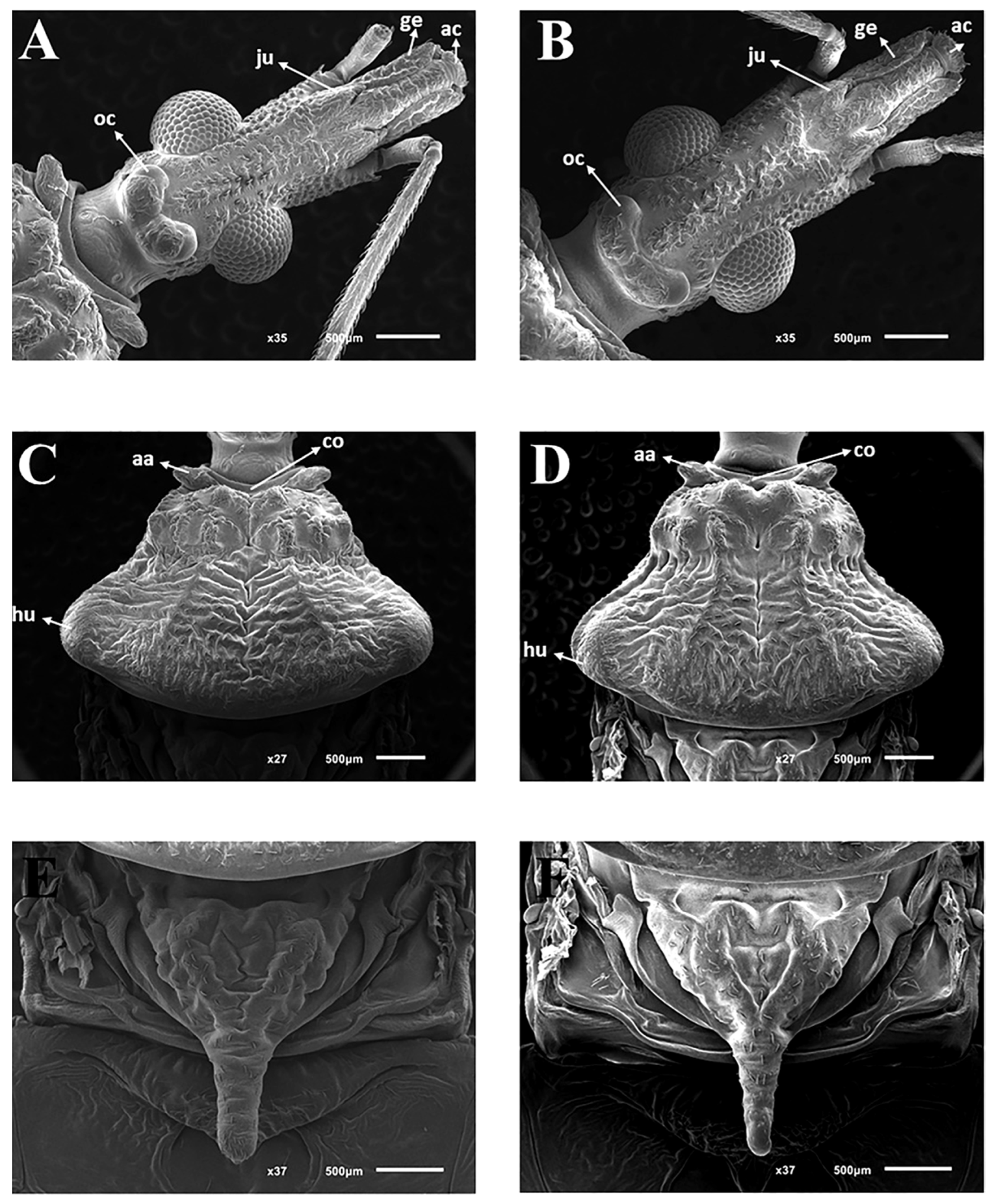
3.2. Description
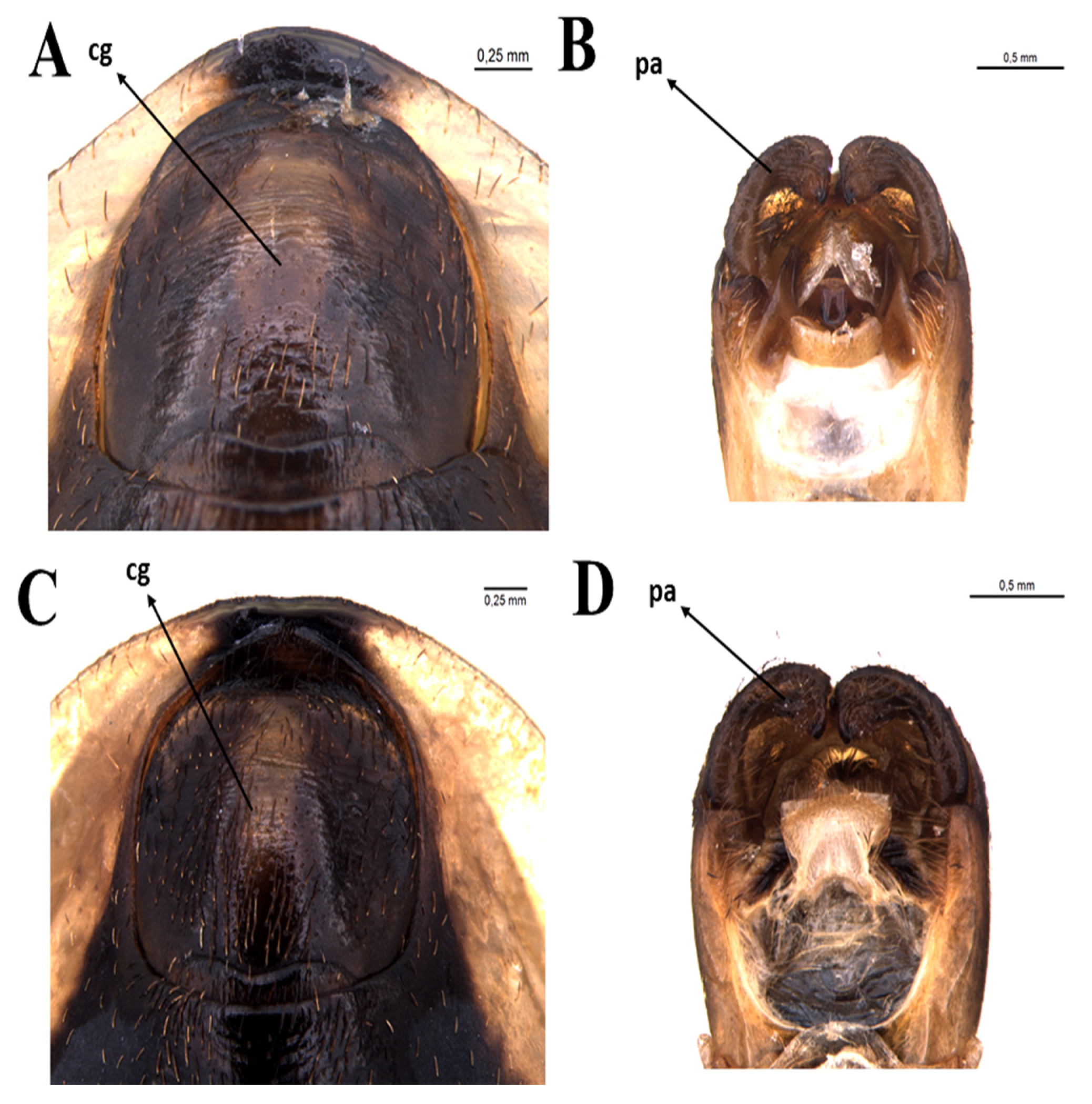
3.2.1. Male
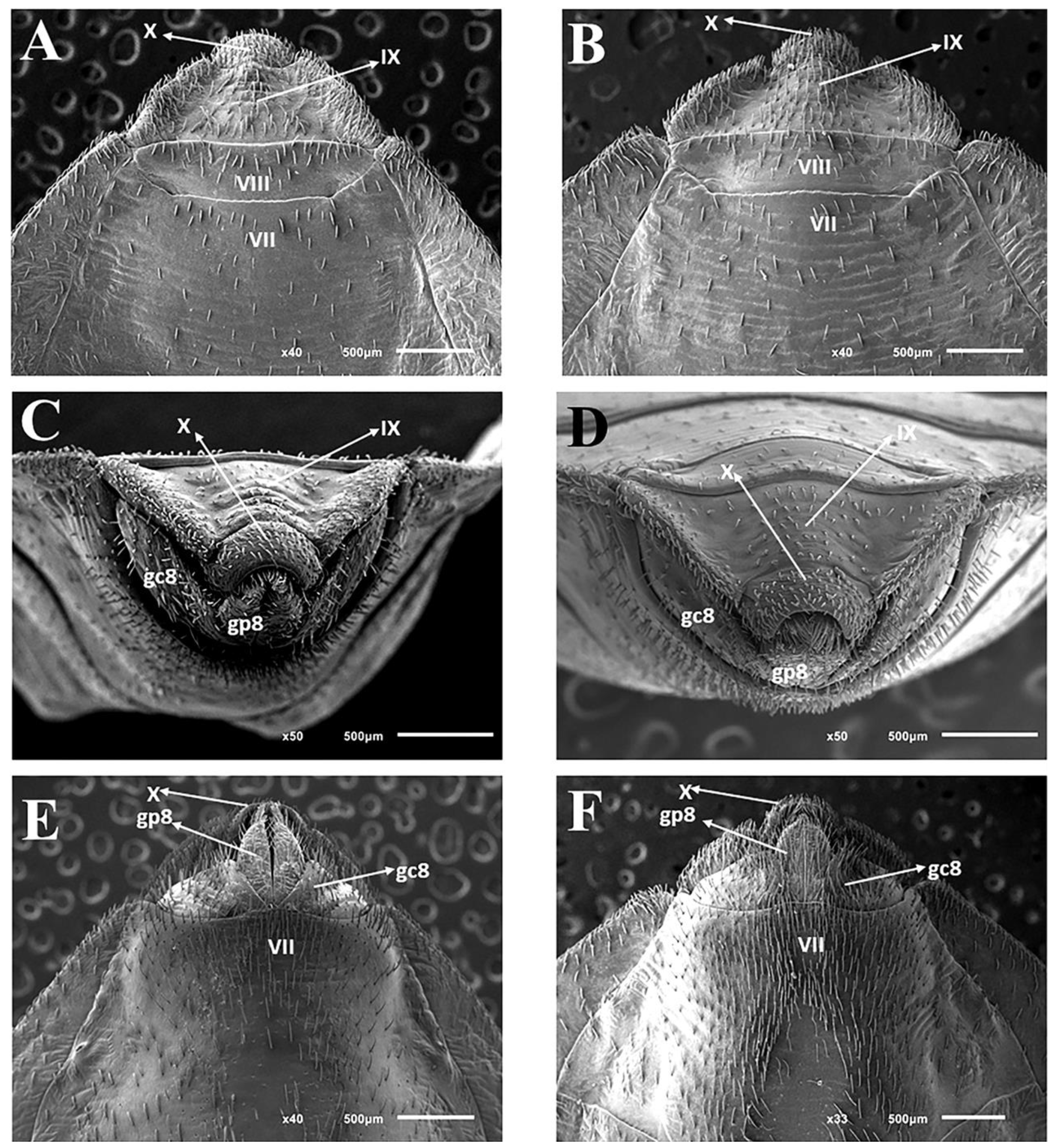
3.2.2. Female
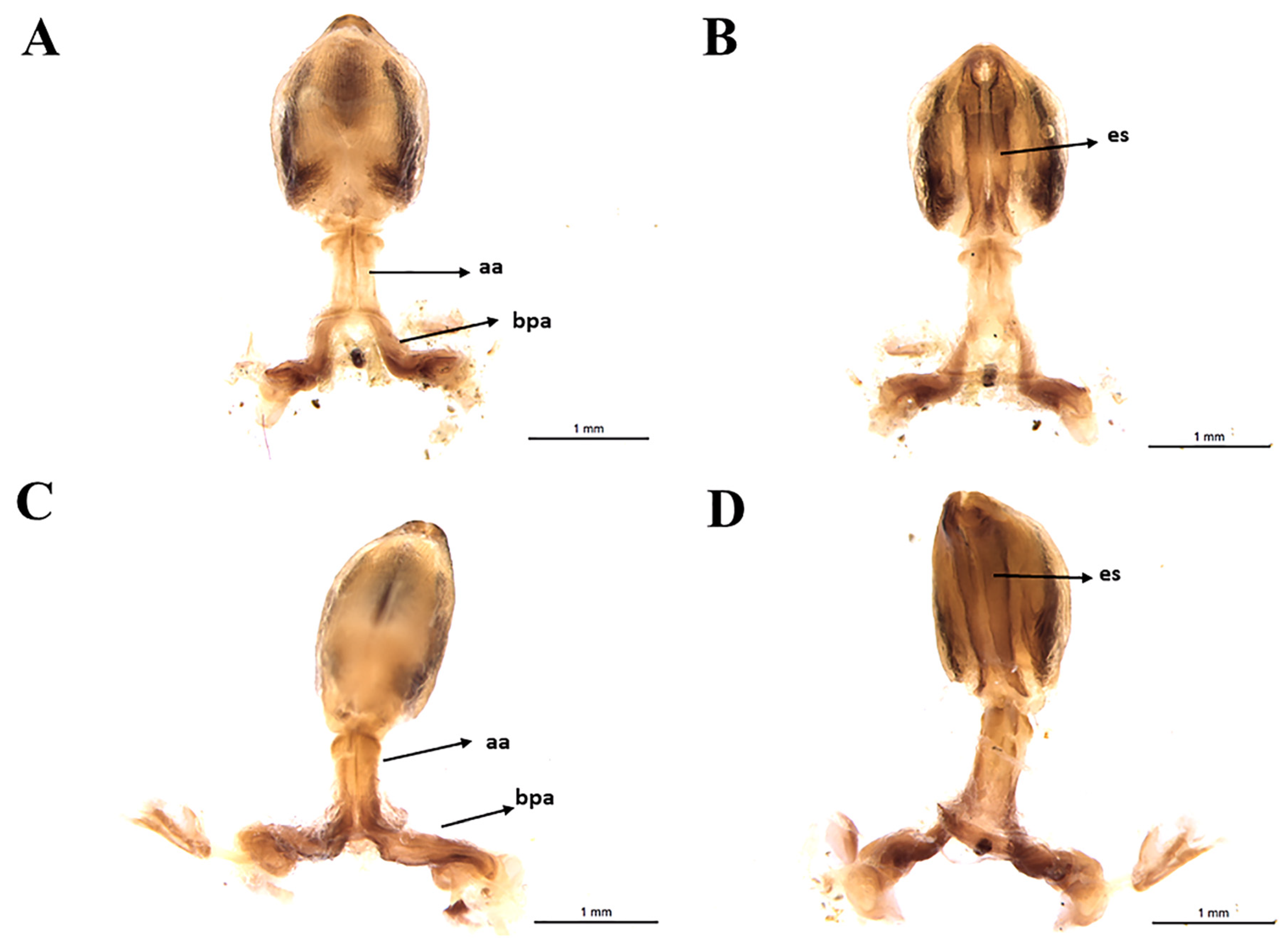
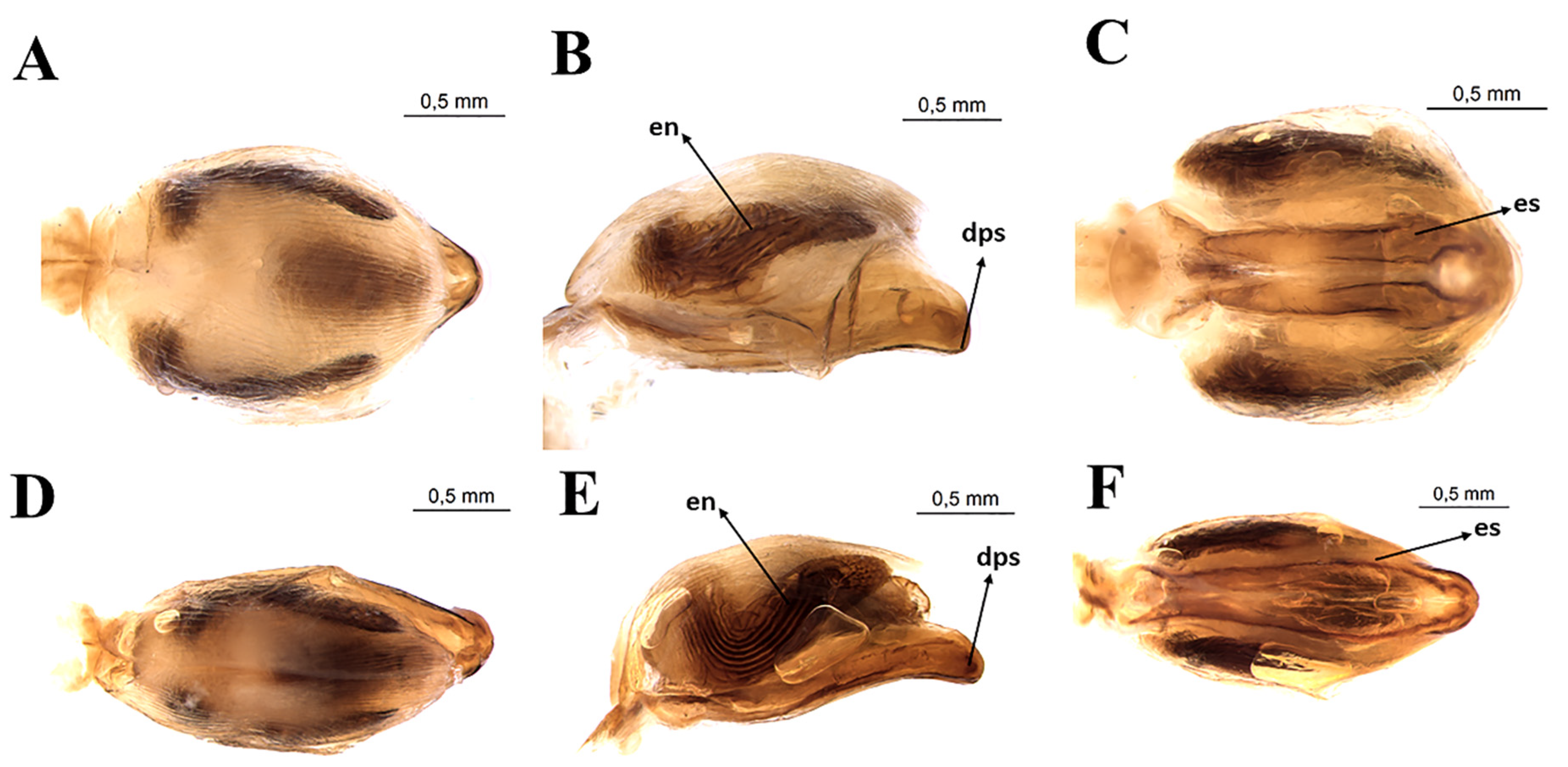
3.3. Female External Genitalia
3.4. Measurement Data
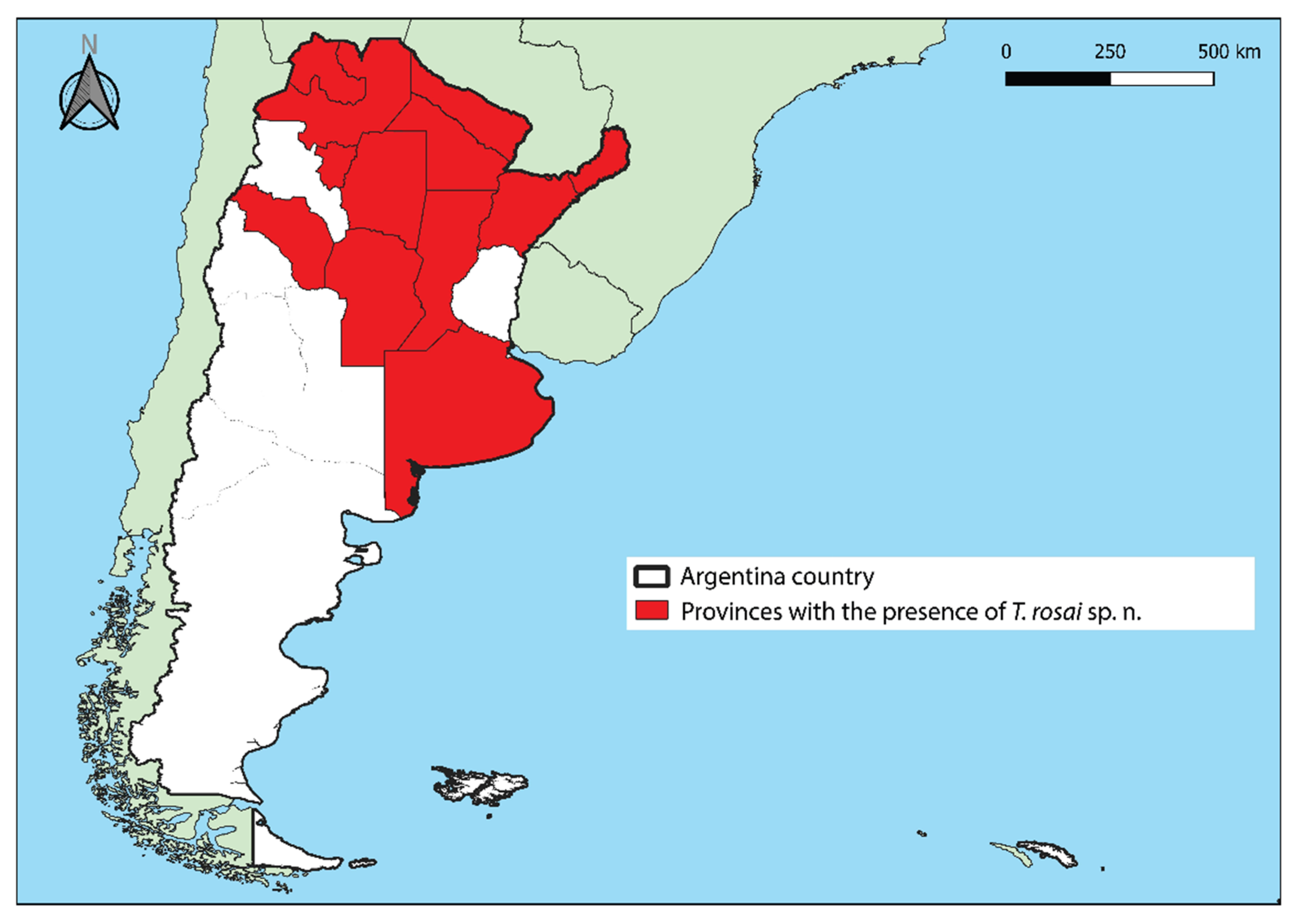
3.5. Distribution
3.6. Observation
3.7. Etymology
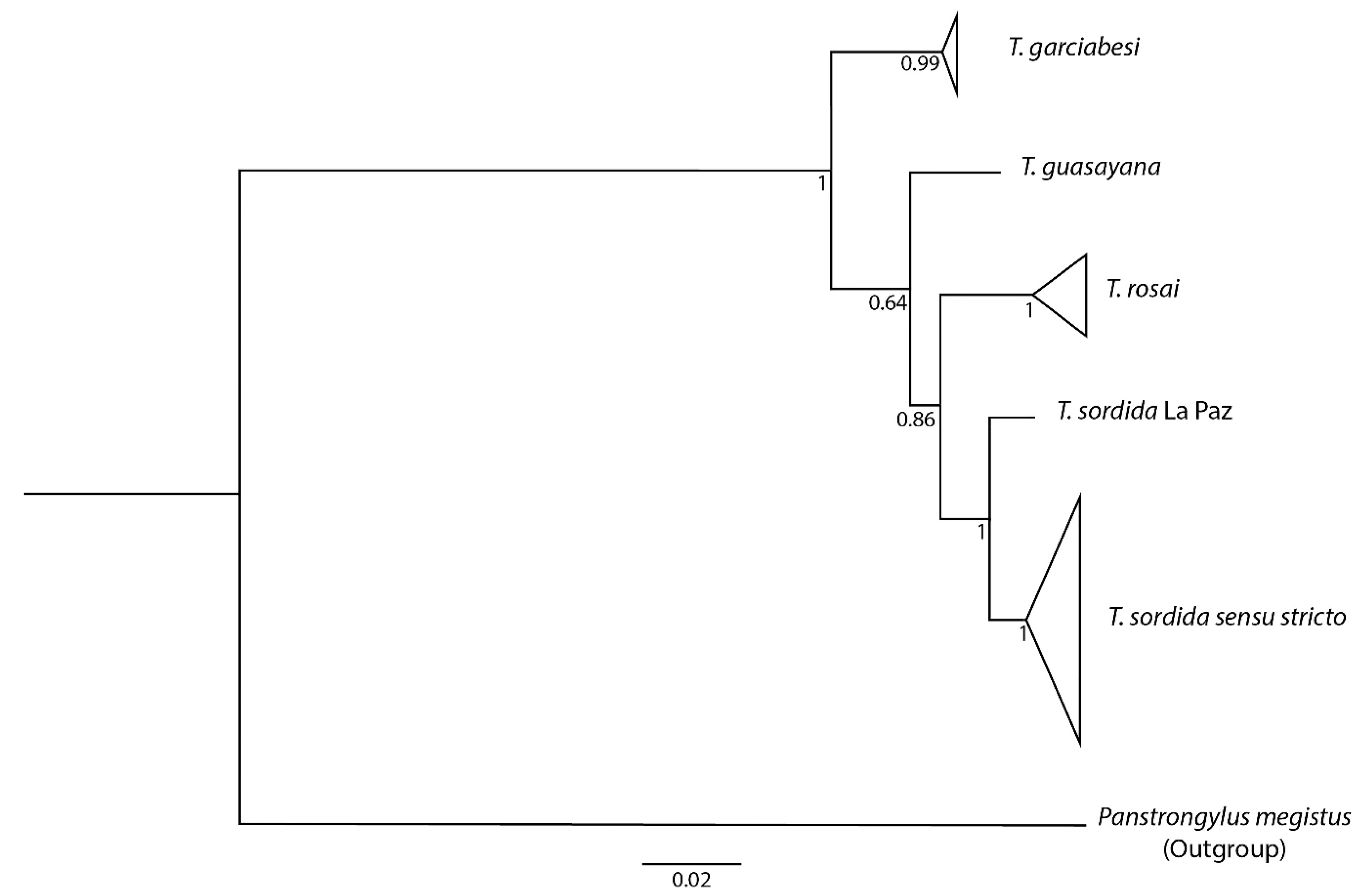
3.8. Phylogenetic Reconstruction and Genetic Distances
3.9. Experimental Crosses
3.10. Taxonomic Notes
3.10.1. List of Synonymous
3.10.2. Distribution
3.10.3. Type Material
3.10.4. Comments
3.11. Literature Data
3.11.1. Genetics Differences
Cytogenetics Differences
Electrophoresis Pattern
Molecular Analyses
3.11.2. Morphological Differences
Cuticular Hydrocarbons Pattern
Geometric Morphometry
3.11.3. Life Cycle and Average Time of Development
Geographic Distribution
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rodriguez, J.B.; Falcone, B.N.; Szajnman, S.H. Detection and treatment of Trypanosoma cruzi : A patent review (2011–2015). Expert. Opin. Ther. Pat. 2016, 26, 993–1015. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Molina, J.A.; Molina, I. Chagas disease. Lancet 2018, 391, 82–94. [Google Scholar] [CrossRef]
- World Health Organization. Available online: https://www.who.int/news-room/fact-sheets/detail/chagas-disease-(american-trypanosomiasis) (accessed on 18 September 2020).
- Pan American Health Organization. Available online: https://www.paho.org/hq/index.php?option=com_content&view=article&id=13566&Itemid=40721&lang=pt (accessed on 18 September 2020).
- Dias, J.C.P.; Ramos, A.N., Jr.; Gontijo, E.D.; Luquetti, A.; Shikanai-Yasuda, M.A.; Coura, J.R.; Torres, R.M.; Melo, J.R.C.; Almeida, E.A.; Oliveira, W., Jr.; et al. 2nd Brazilian consensus on Chagas disease. Rev. Soc. Bras. Med. Trop. 2016, 49, 3–60. [Google Scholar] [CrossRef] [PubMed]
- Galvão, C. Taxonomia dos Vetores da Doença de Chagas da Forma à Molécula, quase três séculos de história. In Atualidades em Medicina Tropical no Brazil: Vetores; Oliveira, J., Alevi, K.C.C., Camargo, L.M.A., Meneguetti, D.U.O., Eds.; Stricto Sensu: Rio Branco, Brazil, 2020; pp. 9–37. [Google Scholar] [CrossRef]
- Chagas, C. Nova tripanozomiaze humana: Estudos sobre a morfolojia e o ciclo evolutivo do Schizotrypanum cruzi n. gen., n. sp., ajente etiolojico de nova entidade morbida do homem. Mem. Inst. Oswaldo Cruz 1909, 1, 159–218. [Google Scholar] [CrossRef] [Green Version]
- Lent, H.; Wygodzynsky, P. Revision of the Triatominae (Hemiptera–Reduviidae) and their significance as vectors of Chagas’ disease. Bull. Am. Mus. Nat. Hist. 1979, 163, 123–520. Available online: http://hdl.handle.net/2246/1282 (accessed on 18 September 2020).
- Gurgel-Gonçalves, R.; Ferreira, J.B.C.; Rosa, A.F.; Bar, M.E.; Galvão, C. Geometric morphometrics and ecological niche modelling for delimitation of near-sibling triatomine species. Med. Vet. Entomol. 2011, 25, 84–93. [Google Scholar] [CrossRef]
- Gurgel-Gonçalves, R.; Galvão, C.; Costa, J.; Peterson, A.T. Geographic distribution of Chagas disease vectors in Brazil based on ecological niche modeling. J. Trop. Med. 2012, 2012, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Galvão, C. Vetores da doença de Chagas no Brazil; Sociedade Brazileira de Zoologia: Curitiba, Brazil, 2014; p. 165. [Google Scholar] [CrossRef] [Green Version]
- Abalos, J.W.; Wygodzinsky, P. Las Triatominae Argentinas (Reduviidae, Hemiptera). Publ. Inst. Med. Reg. Univ. Nac. 1951, 601, 1–179. Available online: https://peerj.com/preprints/3006v1/Article_S1.pdf (accessed on 18 September 2020).
- Actis, A.S.; Traversa, O.C.; Carcavalho, R.U. Estudios taxonómicos sobre el genero Triatoma Laporte mediante la electrophoresis de la linfa. An. Esc. Nac. Cienc. Biol. 1964, 3, 97–106. [Google Scholar]
- Carcavallo, R.U.; Cichero, J.A.; Martínez, A.; Prosen, A.F.; Ronderos, R. Una nueva espécie del género Triatoma Laporte (Hemiptera, Reduviidae, Triatominae). Segundas Jorn. Entomo-Epidemiológicas Argent. 1967, 2, 43–48. [Google Scholar]
- Jurberg, J.; Galvão, C.; Lent, H.; Monteiro, F.; Lopes, M.C.; Panzera, F.; Pérez, R. Revalidação de Triatoma garciabesi Carcavallo, Cichero, Martínez, Prozen & Ronderos, 1967 (Hemiptera, Reduviidae). Entomol. Vectores 1998, 5, 107–122. Available online: http://refhub.elsevier.com/S0001-706X(20)30147-9/sbref0029 (accessed on 18 September 2020).
- Noireau, F.; Gutierrez, T.; Zegarra, M.; Flores, R.; Brenière, F.; Cardozo, L. Cryptic speciation in Triatoma sordida (Hemiptera: Reduviidae) from the Bolivian Chaco. Trop. Med. Int. Health 1998, 3, 364–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panzera, F.; Pita, S.; Nattero, J.; Panzera, Y.; Galvão, C.; Chavez, T. Cryptic speciation in the Triatoma sordida subcomplex (Hemiptera, Reduviidae) revealed by chromosomal markers. Parasit. Vectors 2015, 8, 495–504. [Google Scholar] [CrossRef] [Green Version]
- Panzera, F.; Hornos, S.; Pereira, J.; Cestau, R.; Canale, D.; Diotaiuti, L.; Dujardin, J.P.; Perez, R. Genetic variability and geographic differentiation among three species of triatomine bugs (Hemiptera-Reduviidae). Am. J. Trop. Med. Hyg. 1997, 57, 732–739. [Google Scholar] [CrossRef] [PubMed]
- Calderón-Fernández, G.M.; Juárez, M.P. The cuticular hydrocarbons of the Triatoma sordida species subcomplex (Hemiptera: Reduviidae). Mem. Inst. Oswaldo Cruz 2013, 108, 778–784. [Google Scholar] [CrossRef]
- Nattero, J.; Piccinali, R.M.; Lopes, C.M.; Hernandez, M.L.; Abrahan, L.; Lobbia, P.A.; Rodríguez, C.S.; Carbajal-de-la-Fuente, N.A. Morphometric variability among the species of the Sordida subcomplex (Hemiptera: Reduviidae: Triatominae): Evidence for differentiation across the distribution range of Triatoma Sordida. Parasit. Vectors 2017, 10, 412. [Google Scholar] [CrossRef] [Green Version]
- Rosa, J.A.; Rocha, C.S.; Gardim, S.; Pinto, M.C.; Mendonça, V.J.; Ferreira-Filho, J.C.R.; Carvalho, E.O.C.; Camargo, L.M.A.; Oliveira, J.; Nascimento, J.D.; et al. Description of Rhodnius montenegrensis n. sp. (Hemiptera, Reduviidae: Triatominae) from the state of Rondônia, Brazil. Zootaxa 2012, 3478, 62–76. [Google Scholar] [CrossRef] [Green Version]
- Monteiro, F.A.; Perez, R.; Panzera, F.; Dujardin, J.P.; Galvão, C.; Rocha, D.; Noireau, F.; Schofield, C.J.; Beard, C.B. Mitochondrial DNA variation of Triatoma infestans populations and its implication on the specific status of T. melanosoma. Mem. Inst. Oswaldo Cruz 1999, 94, 229–238. [Google Scholar] [CrossRef]
- Tartarotti, E.; Ceron, C.R. Ribosomal DNA ITS-1 intergenic spacer polymorphism in triatomines (Triatominae, Heteroptera). Biochem. Genet. 2005, 43, 365–373. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar] [CrossRef]
- Thompson, J.; Higgins, D.; Gibson, T. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Gouy, M.; Guindon, S.; Gascuel, O. SeaView version 4: A multiplatform graphical user interface for sequence alignment and phylogenetic tree building. Mol. Biol. Evol. 2010, 27, 221–224. [Google Scholar] [CrossRef] [Green Version]
- Maddison, W.P.; Maddison, D.R. Mesquite: A Modular System for Evolutionary Analysis. Version 3.61. 2019. Available online: http://www.mesquiteproject.org (accessed on 20 July 2020).
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. Available online: https://www.jstor.org/stable/2408678 (accessed on 18 September 2020). [CrossRef]
- Rambut, A. FigTree–Tree Figure Drawing Tool Version v.1.4.3; Institute of Evolutionary Biology, University of Edinburgh: Edinburgh. 2016. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 18 September 2020).
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarisation in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [Green Version]
- Mayr, E. Animal Species and Evolution; Harvard University Press: Cambridge, UK, 1963. [Google Scholar]
- Dobzhansky, T. Genetics of the Evolutionary Process; Columbia University Press: New York, NY, USA, 1970. [Google Scholar]
- Costa, J.; Almeida, C.E.; Dujardin, J.P.; Beard, C.B. Crossing Experiments Detect Genetic Incompatibility among Populations of Triatoma brasiliensis Neiva, 1911 (Heteroptera, Reduviidae, Triatominae). Mem. Inst. Oswaldo Cruz 2003, 98, 637–639. [Google Scholar] [CrossRef] [Green Version]
- Mendonça, V.J.; Alevi, K.C.C.; Pinotti, H.; Gurgel-Gonçalves, R.; Pita, S.; Guerra, A.L.; Panzera, F.; Araújo, R.F.; Azeredo-Oliveira, M.T.V.; Rosa, J.A. Revalidation of Triatoma bahiensis Sherlock & Serafim, 1967 (Hemiptera: Reduviidae) and phylogeny of the T. brasiliensis species complex. Zootaxa 2016, 4107, 239–254. [Google Scholar] [CrossRef] [PubMed]
- Dujardin, J.P.; Schofield, C.J.; Panzera, F. Los vectores de la enfermedad de Chagas; Académie Royale des Sciences D’Outre-Mer: Bruxelles, Belgium, 2002; 189p. [Google Scholar]
- Galvão, C.; Carcavallo, R.U.; Rocha, D.S.; Jurberg, J. A checklist of the current valid species of the subfamily Triatominae Jeannel, 1919 (Hemiptera, Reduviidae) and their geographical distribution, with nomenclatural and taxonomic notes. Zootaxa 2003, 202, 1–36. [Google Scholar] [CrossRef] [Green Version]
- QGIS Geographic Information System. Available online: http://qgis.org (accessed on 19 September 2020).
- Instituto Geográfico Nacional. Available online: https://www.ign.gob.ar/NuestrasActividades/InformacionGeoespacial/CapasSIG (accessed on 19 September 2020).
- Ramos, L.J.; Souza, J.L.; Souza, C.R.; Oliveira, J.; Rosa, J.A.; Camargo, L.M.A.; Cunha, R.M.; Meneguetti, D.U.O. First report of Triatoma sordida Stål, 1859 (Hemiptera, Reduviidae, Triatominae) in the State of Acre and Brazilian Western Amazon. Rev. Soc. Bras. Med. Trop. 2018, 51, 77–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, J.M.S.; Moreira, F.F.F.; Deckert, J.; Galvão, C. List of the type specimens of Triatominae (Hemiptera: Heteroptera: Reduviidae) in the Hemimetabola Collection of the Museum für Naturkunde, Berlin. Zootaxa 2020, 4809, 329–348. [Google Scholar] [CrossRef] [PubMed]
- Bardella, V.B.; Pita, S.; Vanzela, A.L.L.; Galvão, C.; Panzera, F. Heterochromatin base pair composition and diversification in holocentric chromosomes of kissing bugs (Hemiptera, Reduviidae). Mem. Inst. Oswaldo Cruz 2016, 111, 614–662. [Google Scholar] [CrossRef] [Green Version]
- García, B.A.; Moriyama, E.N.; Powell, J.R. Mitochondrial DNA sequences of triatomines (Hemiptera: Reduviidae): Phylogenetic relationships. J. Med. Entomol. 2001, 38, 675–683. [Google Scholar] [CrossRef]
- Belintani, T.; Oliveira, J.; Pinotti, H.; Silva, L.A.; Alevi, K.C.C.; Galvão, C.; Rosa, J.A. Phylogenetic and phenotypic relationships of the Triatoma sordida subcomplex (Hemiptera: Reduviidae: Triatominae). Acta Trop. 2020, 212, 105679. [Google Scholar] [CrossRef]
- Madeira, F.F.; Delgado, L.M.G.; Bittinelli, I.F.; Sartori, R.Q.; Oliveira, J.; Rosa, J.A.; Azeredo-Oliveira, M.T.V.; Alevi, K.C.C. Revisiting the genetic variability of the Brazilian peridomestic populations of the Chagas disease vector Triatoma sordida (Stål, 1859) (Hemiptera, Triatominae). Infect. Genet. Evol. 2020, 85, 104568. [Google Scholar] [CrossRef]
- Oscherov, E.B.; Damborsky, M.P.; Bar, M.E. Características biológicas de Triatoma sordida (Heteroptera, Reduviidae): Ciclo de vida. Rev. Soc. Eniomol. Argent. 1998, 57, 13–17. Available online: https://www.biotaxa.org/RSEA/article/download/33689/29858 (accessed on 18 September 2020).
- Souza, J.M.P.; Rodrigues, V.L.C.C.; Rocha e Silva, E.O. Triatoma sordida: Considerações sobre o tempo de vida das formas adultas e sobre a oviposição das fêmeas. Rev. Saúde Públ. 1978, 12, 291–296. [Google Scholar] [CrossRef] [Green Version]
- Pinto, C.F. Fatos curiosos sobre a biologia do Triatoma sordida (Nota prévia). Rev. Soc. Bras. Med. 1949, 6, 305. [Google Scholar]
- Schofield, C.J.; Lehane, M.J.; McEwan, P.; Catala, S.S.; Gorla, D.E. Dispersive flight by Triatoma sordida. Trans. R. Soc. Trop. Med. Hyg. 1991, 85, 676–678. [Google Scholar] [CrossRef]
- Bar, M.E.; Oscherov, E.B.; Damborsky, M.P.; Varela, M.E.; Mizdraji, G.; Porcel, E. Triatomismo del Departamento San Luis del Palmar de la Provincia de Corrientes, Argentina. Medicina 1992, 52, 193–201. [Google Scholar] [PubMed]
- Bar, M.E.; Oscherov, E.B.; Damborsky, M.P. Presencia de Triatoma sordida Stål, 1859 en ecótopos urbanos de la ciudad de Corrientes, Argentina. Rev. Saúde Públ. 1993, 27, 117–122. [Google Scholar] [CrossRef] [Green Version]
- Bar, M.E.; Oscherov, E.B.; Damborsky, M.P.; Porcel, E.A.; Alvarez, B.M. Population interactions between Triatoma infestans and Triatoma sordida. Rev. Saúde Públ. 1994, 28, 59–68. [Google Scholar] [CrossRef]
- Bar, M.E.; Oscherov, E.B.; Avalos, G.; Damborsky, M.P.; Alvarez, B.M. Infección natural de Triatoma sordida por Trypanosoma cruzi en ecótopos silvestres de la provincia de Corrientes, Argentina. In Resúmenes, 5ta. Jornada de Biología, Instituto de Investigaciones en Ciencias de la Salud; Universidad Nacional de Asunción: Asunción, Paraguay, 1996; pp. 32–33. [Google Scholar]
- Bar, M.E.; Damborsky, M.P.; Oscherov, E.B.; Alvarez, B.M.; Mizdraji, G.; Avalos, G. Infestación domiciliaria por triatominos y seroprevalencia humana en el Departamento Empedrado, Corrientes, Argentina. C. Saúde Pública 1997, 13, 305–312. [Google Scholar] [CrossRef]
- Bar, M.E.; Wisnievsky-Colli, C. Triatoma sordida Stål 1859 (Hemiptera, Reduviidae: Triatominae) in palms of northeastern Argentina. Mem. Inst. Oswaldo Cruz 2001, 96, 895–899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bar, M.E. Estructura y dinámica poblacional de Triatoma sordida Stål 1859 en una comunidad de Butia yatay en Argentina. Bol. Malariol. Salud Ambient. 2006, 46, 21–30. Available online: http://ve.scielo.org/scielo.php?script=sci_arttext&pid=S1690-46482006000100003&lng=es&nrm=iso (accessed on 18 September 2020).
- Gorla, D.E.; Jurberg, J.; Catala, S.S.; Schofield, C.J. Systematics of Triatoma sordida, T. guasayana and T. patagonica (Hemiptera, Reduviidae). Mem. Inst. Oswaldo Cruz 1993, 88, 379–385. [Google Scholar] [CrossRef]
- McEwen, P.K.; Lehane, M.J. Factors influencing flight initiation in the triatomine bug Triatoma sordida (Hemiptera: Reduviidae). Insect Sci. Appl. 1993, 14, 461–464. [Google Scholar] [CrossRef]
- Wisnivesky-Colli, C.; Gürtler, R.E.; Solarz, N.D.; Schweigmann, N.J.; Silvia, M.; Pietrokovsky, S.M.; Alberti, A.; Flo, J. Dispersive flight and house invasion by Triatoma guasayana and Triatoma sordida in Argentina. Mem. Inst. Oswaldo Cruz 1993, 88, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Wisnivesky-Colli, C.; Pietrokovsky, S.; Bottazzi, V.; Haedo, A.; Solarz, N.; Schweigmann, N.; Canale, D.; Reyes, O.R. Laboratory comparison of feeding success among Triatoma infestans, T. guasayana, and T. sordida (Hemiptera: Reduviidae). J. Med. Entomol. 1995, 32, 583–587. [Google Scholar] [CrossRef] [PubMed]
- Crocco, L.B.; Catalá, S.S. Feeding and defaecation patterns in Triatoma sordida. Mem. Inst. Oswaldo Cruz 1996, 91, 409–413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crocco, L.; Catalá, S. Host preferences of Triatoma sordida. Ann. Trop. Med. Parasitol. 1997, 91, 927–930. [Google Scholar] [CrossRef] [PubMed]
- Oscherov, E.B.; Damborsky, M.P.; Bar, M.E.; Porcel, E. Population statistics of Triatoma sordida Stäl 1859 (Hemiptera: Reduviidae) in experimental conditions. Rev. Saúde Públ. 1996, 30, 438–443. [Google Scholar] [CrossRef] [PubMed]
- Oscherov, E.B.; Damborsky, M.P.; Bar, M.E.; Avalos, G.; Alvarez, B.M.; Presman, H.; Wisnivesky-Colli, C. Interactions between Triatoma infestans and Triatoma sordida (Hemiptera: Reduviidae) in artificial ecotopes: Population growth and age structure. J. Med. Entomol. 2001, 38, 214–217. [Google Scholar] [CrossRef]
- Pietrokovsky, S.; Bottazzi, V.; Schweigmann, N.; Haedo, A.; Wisnivesky-Colli, C. Comparison of the blood meal size among Triatoma infestans, T. guasayana and T. sordida (Hemiptera: Reduviidae) of Argentina under laboratory conditions. Mem. Inst. Oswaldo Cruz 1996, 91, 241–242. [Google Scholar] [CrossRef] [Green Version]
- Schweigmann, N.J.; Pietrokovsky, S.; Conti, O.; Escosteguy, M.; Bottazzi, V.; Solarz, N.; Wisnivesky-Colli, C. Infection of Triatoma guasayana, Triatoma sordida and Triatoma infestans by Trypanosoma cruzi from a naturally infected opossum. Mem. Inst. Oswaldo Cruz 1997, 92, 151–152. [Google Scholar] [CrossRef] [Green Version]
- Rebagliati, P.; Papeschi, A.G.; Mola, L.M.; Pietrokovsky, S.; Gajate, P.; Botazzi, V.; Wisnivesky-Colli, C. Comparative meiotic studies in Triatoma sordida (Stål) and T. guasayana Wygodzinsky & Abalos (Reduviidae, Heteroptera). Mem. Inst. Oswaldo Cruz 1998, 93, 309–315. [Google Scholar] [CrossRef] [Green Version]
- Maffey, L.; Cardinal, M.V.; Ordonez-Krasnowski, P.C.; Lanati, L.A.; Lauricella, M.A.; Schijman, A.G.; Gürtler, R.E. Direct molecular identification of Trypanosoma cruzi discrete typing units in domestic and peridomestic Triatoma infestans, and Triatoma sordida from the Argentine Chaco. Parasitology 2012, 139, 1570–1579. [Google Scholar] [CrossRef] [Green Version]
- Macchiaverna, N.P.; Gaspe, M.S.; Enriquez, G.F.; Tomassone, L.; Gürtler, R.E.; Cardinal, M.V. Trypanosoma cruzi infection in Triatoma sordida before and after community-wide residual insecticide spraying in the Argentinean Chaco. Acta Trop. 2015, 143, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Pessoa, G.C.A.; Sousa, T.N.; Sonoda, I.V.; Diotaiuti, L. Assessing the mitochondrial DNA diversity of the chagas disease vector Triatoma sordida (Hemiptera: Reduviidae). Mem. Inst. Oswaldo Cruz 2016, 111, 322–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-Planes, L.I.; Gaspe, M.S.; Enriquez, G.F.; Gürtler, R.E. Habitat-specific occupancy and a metapopulation model of Triatoma sordida (Hemiptera: Reduviidae), a secondary vector of Chagas disease, in northeastern Argentina. J. Med. Entomol. 2018, 55, 370–381. [Google Scholar] [CrossRef] [PubMed]
- Pita, S.; Lorite, P.; Nattero, J.; Galvão, C.; Alevi, K.C.C.; Teves, S.C.; Azeredo-Oliveira, M.T.V.; Panzera, F. New arrangements taxon several species subcomplexes of Triatoma genus based on the chromosomal position of ribosomal genes (Hemiptera -Triatominae). Infect. Genet. Evol. 2016, 43, 225–231. [Google Scholar] [CrossRef] [Green Version]
- Carcavallo, R.U.; Jurberg, J.; Lent, H.; Noireau, F.; Galvão, C. Phylogeny of the Triatominae (Hemiptera Reduviidae). Proposals for taxonomic arrangements. Entomol. Vectores 2000, 7, 1–99. Available online: https://horizon.documentation.ird.fr/exl-doc/pleins_textes/divers12-11/010024157.pdf (accessed on 18 September 2020).
- Schofield, C.J.; Galvão, C. Classification, evolution and species groups within the Triatominae. Acta Trop. 2009, 110, 88–100. [Google Scholar] [CrossRef]
- Dorn, P.L.; Justi, S.A.; Dale, C.; Stevens, L.; Galvão, C.; Lima-Cordón, R.; Monroy, C. Description of Triatoma mopan sp. n. from a cave in Belize (Hemiptera, Reduviidae, Triatominae). Zookeys 2018, 775, 69–95. [Google Scholar] [CrossRef]
- Lima-Cordón, R.A.; Monroy, M.C.; Stevens, L.; Rodas, A.; Rodas, G.A.; Dorn, P.L.; Justi, A.S. Description of Triatoma huehuetenanguensis sp. n., a potential Chagas disease vector (Hemiptera, Reduviidae, Triatominae). ZooKeys 2019, 820, 51–70. [Google Scholar] [CrossRef]
- Schreiber, G.; Pinho, A.C.; Parentoni, R.; Salgado, A.A.; Godoi, T.L., Jr. Cytogenetics of Triatominae: II Spermatogenesis in hybrid between Triatoma infestans and Triatoma pseudomaculata (Hemiptera, Reduviidae). Rev. Bras. Biol. 1974, 31, 1–9. Available online: https://pubmed.ncbi.nlm.nih.gov/4618618/ (accessed on 18 September 2020).
- Pérez, R.; Hérnandez, M.; Quintero, O.; Scvortzoff, E.; Canale, D.; Méndez, L.; Cohanoff, C.; Martino, M.; Panzera, F. Cytogenetic analysis of experimental hybrids in species of Triatominae (Hemiptera-Reduviidae). Genetica 2005, 125, 261–270. [Google Scholar] [CrossRef]
- Martínez-Ibarra, J.A.; Grant-Guillén, Y.; Delgadillo-Aceves, I.N.; Zumaya-Estrada, F.A.; Rocha-Chávez, G.; Salazarschettino, P.M.; Alejandre-Aguilar, R.; Villalobos, G.; Nogueda-Torres, B. Biological and genetic aspects of crosses between phylogenetically close species of Mexican Triatomines (Hemiptera: Reduviidae). J. Med. Entomol. 2011, 48, 705–707. [Google Scholar] [CrossRef] [Green Version]
- Díaz, S.; Panzera, F.; Jaramillo-O, N.; Pérez, R.; Fernández, R.; Vallejo, G.; Saldaña, A.; Calzada, J.E.; Triana, O.; Gómez-Palácio, A. Genetic, Cytogenetic and Morphological Trends in the Evolution of the Rhodnius (Triatominae: Rhodniini) Trans-Andean Group. PLoS ONE 2014, 9, e87493. [Google Scholar] [CrossRef] [Green Version]
- Mendonça, V.J.; Alevi, K.C.C.; Medeiros, L.M.O.; Nascimento, J.D.; Azeredo-Oliveira, M.T.V.; Rosa, J.A. Cytogenetic and morphologic approaches of hybrids from experimental crosses between Triatoma lenti Sherlock & Serafim, 1967 and T. sherlocki Papa et al., 2002 (Hemiptera: Reduviidae). Infect. Genet. Evol. 2014, 26, 123–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campos-Soto, R.; Panzera, F.; Pita, S.; Lages, C.; Solari, A.; Botto-Mahan, C. Experimental crosses between Mepraia gajardoi and M. spinolai and hybrid chromosome analyses reveal the occurrence of several isolation mechanisms. Infect. Genet. Evol. 2016, 45, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Alevi, K.C.C.; Pinotti, H.; Araújo, R.F.; Azeredo-Oliveira, M.T.V.; Rosa, J.A.; Mendonça, V.J. Hybrid colapse confirm the specific status of Triatoma bahiensis Sherlock and Serafim, 1967 (Hemiptera, Triatominae). Am. J. Trop. Med. Hyg. 2018, 98, 475–477. [Google Scholar] [CrossRef] [PubMed]
- Villacís, A.G.; Dujardin, J.P.; Panzera, F.; Yumiseva, C.A.; Pita, S.; Santillán-Guayasamín, S.; Orozco, M.I.; Mosquera, K.D.; Grijalva, M.J. Chagas vectors Panstrongylus chinai (Del Ponte, 1929) and Panstrongylus howardi (Neiva, 1911): Chromatic forms or true species? Parasit. Vectors 2020, 13, 226. [Google Scholar] [CrossRef] [PubMed]
- Castro, G.B.; Machado, E.M.M.; Borges, E.C.; Lorosa, E.S.; Andrade, R.E.; Diotaiuti, L.; Azeredo, B.V.M. Trypanosoma cruzi Peridomiciliar Transmission by Triatoma sordida in the Municipality of Patis, Gerais State, Brazil. Mem. Inst. Oswaldo Cruz 1997, 92, 434. [Google Scholar] [CrossRef]
- Lorosa, E.S.; Andrade, R.E.; Santos, S.M.; Pereira, C.A. Estudo da infecção natural e da fonte alimentar do Triatoma sordida (Stal, 1859), (Hemiptera -Reduviidae) na região norte de Minas Gerais, Brazil, através da reação de precipitina. Entomol. Vectores 1998, 5, 13–22. [Google Scholar]
- Brenière, S.F.; Aliaga, C.; Waleckx, E.; Buitrago, R.; Salas, R.; Barnabé, C.; Tybayrenc, M.; Noireau, F. Genetic characterization of DTUs in wild Triatoma infestans from Bolivia: Predominance of TcI. PLoS Negl. Trop. Dis. 2012, 6, e1650. [Google Scholar] [CrossRef] [PubMed]
- Rossi, J.C.N.; Duarte, E.C.; Gurgel-Gonçalves, R. Factors associated with the occurrence of Triatoma sordida (Hemiptera: Reduviidae) in rural localities of Central-West Brazil. Mem. Inst. Oswaldo Cruz 2015, 110, 192–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pickenheyn, J.; Guimarães, R.B.; Lima, S.C.; Curto, S. Processo de urbanização da Doença de Chagas na Argentina e no Brazil. Hygeia 2009, 4, 58–69. Available online: http://www.seer.ufu.br/index.php/hygeia/article/view/16920 (accessed on 18 September 2020).













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Geographic Distribution | Marker | ||||
|---|---|---|---|---|---|---|
| COI | Cyt b | 28S | 16S | ITS-1 | ||
| T. sordida | Seabra, Bahia, Brazil | MH029692 | MH054940 | MG417085 | - | * |
| T. sordida | Posses, Goiás, Brazil | MH029695 | * | MH059640 | - | - |
| T. sordida | Monte Azul, Minas Gerais, Brazil | MH029693 | MH054941 | MG417086 | - | - |
| T. sordida | Pantanal, Mato Grosso do Sul, Brazil | MH029694 | KC249292 | KC249210 | KC249075 | - |
| T. sordida | Apolo, La Paz, Bolívia | MH029696 | MH054942 | MH059641 | - | * |
| T. rosai sp. nov. | San Miguel, Corrientes, Argentina | MH029697 | * | MG417088 | - | * |
| T. rosai sp. nov. | San Miguel, Corrientes, Argentina | KC249387 | KC249295 | KC249213 | KC249078 | - |
| T. garciabesi | Rivadavia, Salta, Argentina | MH029698 | MH054943 | MG417089 | - | - |
| T. garciabesi | Rivadavia, Salta, Argentina | KC249338 | KC249249 | KC249158 | KC249006 | - |
| T. guasayana | Santa Cruz, Bolívia | KC249343 | KC249252 | KC249162 | KC249011 | - |
| P. megistus | Nova Prata, Rio Grande do Sul, Brazil | KC249308 | KC249228 | KC249136 | KC248970 | AM949580 |
| Sex | Species | Structures | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Male | HL | IE | PHW | PRO | SCU | THO | ABD | TL | |
| T. rosai sp. nov. | 5.174 | 0.951 | 1.490 | 4.717 | 3.077 | 6.704 | 15.117 | 26.98 | |
| T. sordida | 4.857 | 0.966 | 1.381 | 4.107 | 2.816 | 6.093 | 14.084 | 25.02 | |
| Female | T. rosai sp. nov. | 5.055 | 1.003 | 1.486 | 4.429 | 3.215 | 6.531 | 16.418 | 27.99 |
| T. sordida | 5.208 | 1.066 | 1.510 | 4.221 | 3.214 | 6.459 | 16.167 | 27.81 | |
| Species | (1) | (2) | (3) | (4) | (5) | (6) | (7) | (8) | (9) |
|---|---|---|---|---|---|---|---|---|---|
| (1) Posse GO * | |||||||||
| (2) Seabra BA * | 0.007 | ||||||||
| (3) Monte Azul MG * | 0.007 | 0.000 | |||||||
| (4) Pantanal MS * | 0.007 | 0.000 | 0.000 | ||||||
| (5) T. rosai sp. nov. | 0.085 | 0.087 | 0.087 | 0.087 | |||||
| (6) T. garciabesi | 0.082 | 0.074 | 0.074 | 0.074 | 0.099 | ||||
| (7) T. garciabesi | 0.082 | 0.074 | 0.074 | 0.074 | 0.097 | 0.000 | |||
| (8) T. guasayana | 0.078 | 0.074 | 0.074 | 0.074 | 0.069 | 0.067 | 0.067 | ||
| (9) P. megistus | 0.245 | 0.234 | 0.234 | 0.234 | 0.259 | 0.240 | 0.241 | 0.244 |
| Experimental Crosses | Number of Eggs | Hatching Rate | Number of Nymphs | Mortality Rate |
|---|---|---|---|---|
| Interspecific | ||||
| ♀ T. sordida sensu stricto x T. rosai sp. nov. ♂ | 241 | 77% | 186 | 80% |
| ♀ T. rosai sp. nov. x T. sordida sensu stricto ♂ | 237 | 59% | 139 | 73% |
| Intraspecific | ||||
| ♀ T. sordida sensu stricto x T. sordida sensu stricto ♂ | 590 | 73% | 429 | 20% |
| ♀ T. rosai sp. nov. x T. rosai sp. nov. ♂ | 208 | 61% | 125 | 17% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alevi, K.C.C.; de Oliveira, J.; Garcia, A.C.C.; Cristal, D.C.; Delgado, L.M.G.; de Freitas Bittinelli, I.; dos Reis, Y.V.; Ravazi, A.; de Oliveira, A.B.B.; Galvão, C.; et al. Triatoma rosai sp. nov. (Hemiptera, Triatominae): A New Species of Argentinian Chagas Disease Vector Described Based on Integrative Taxonomy. Insects 2020, 11, 830. https://doi.org/10.3390/insects11120830
Alevi KCC, de Oliveira J, Garcia ACC, Cristal DC, Delgado LMG, de Freitas Bittinelli I, dos Reis YV, Ravazi A, de Oliveira ABB, Galvão C, et al. Triatoma rosai sp. nov. (Hemiptera, Triatominae): A New Species of Argentinian Chagas Disease Vector Described Based on Integrative Taxonomy. Insects. 2020; 11(12):830. https://doi.org/10.3390/insects11120830
Chicago/Turabian StyleAlevi, Kaio Cesar Chaboli, Jader de Oliveira, Ariane Cristina Caris Garcia, Daniel Cesaretto Cristal, Luiza Maria Grzyb Delgado, Isadora de Freitas Bittinelli, Yago Visinho dos Reis, Amanda Ravazi, Ana Beatriz Bortolozo de Oliveira, Cleber Galvão, and et al. 2020. "Triatoma rosai sp. nov. (Hemiptera, Triatominae): A New Species of Argentinian Chagas Disease Vector Described Based on Integrative Taxonomy" Insects 11, no. 12: 830. https://doi.org/10.3390/insects11120830