
Arginase 1 (Arg1) as an Up-Regulated Gene in COVID-19 Patients: A Promising Marker in COVID-19 Immunopathy

 ,
,  , ,
, ,  , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients and Samples
2.2. The RNA Extraction and Complementary DNA Synthesis
2.3. Real-Time PCR
2.4. Protein–Protein Interaction (PPI)
2.5. Statistical Analysis
3. Results
3.1. Population Study
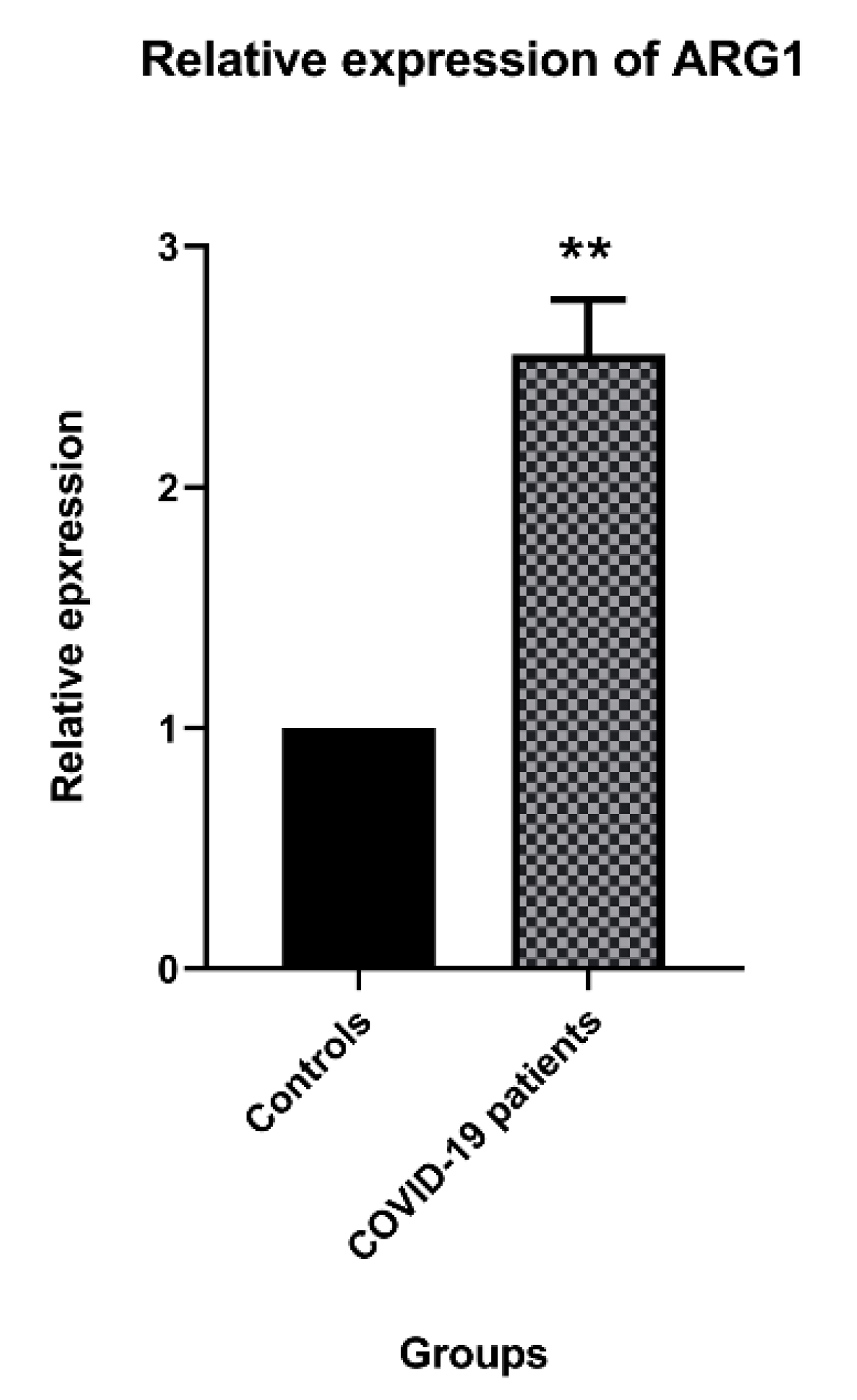
3.2. Arg1 Expression in the COVID-19 Patients
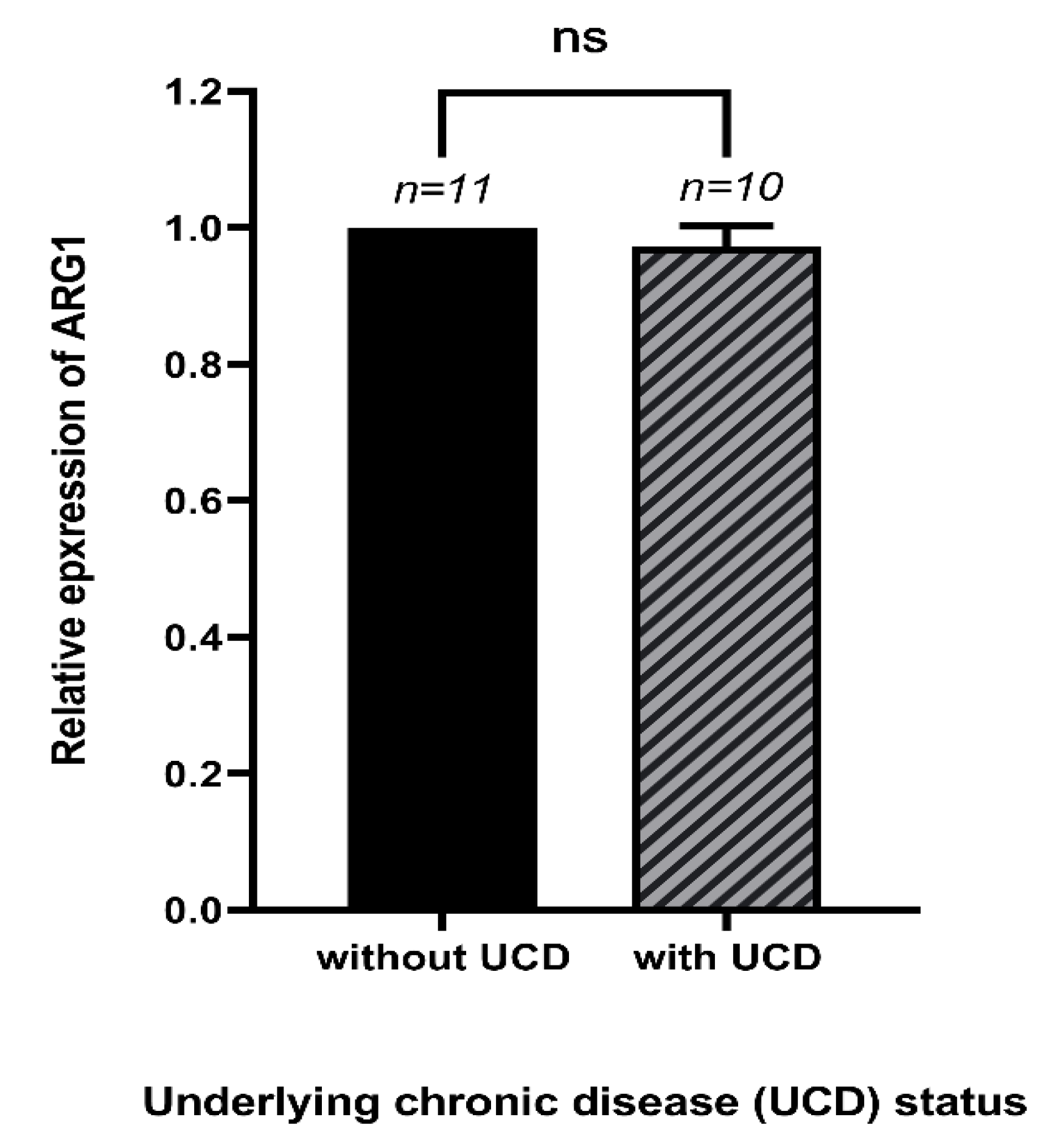
3.3. The Expression of Arg1 and Underlying Chronic Diseases of COVID-19 Patients
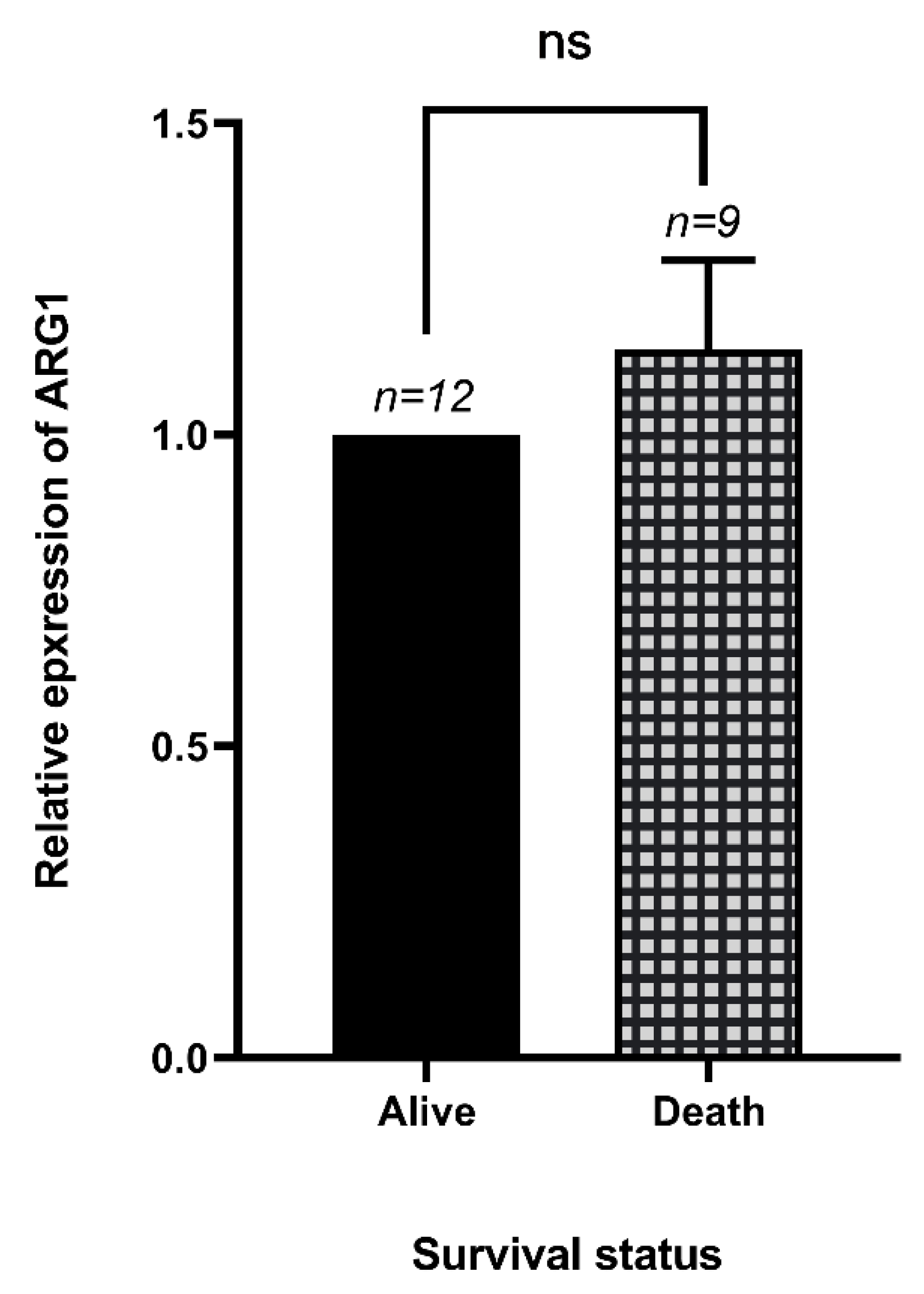
3.4. The Expression of Arg1 in Dead and Alive COVID-19 Patients
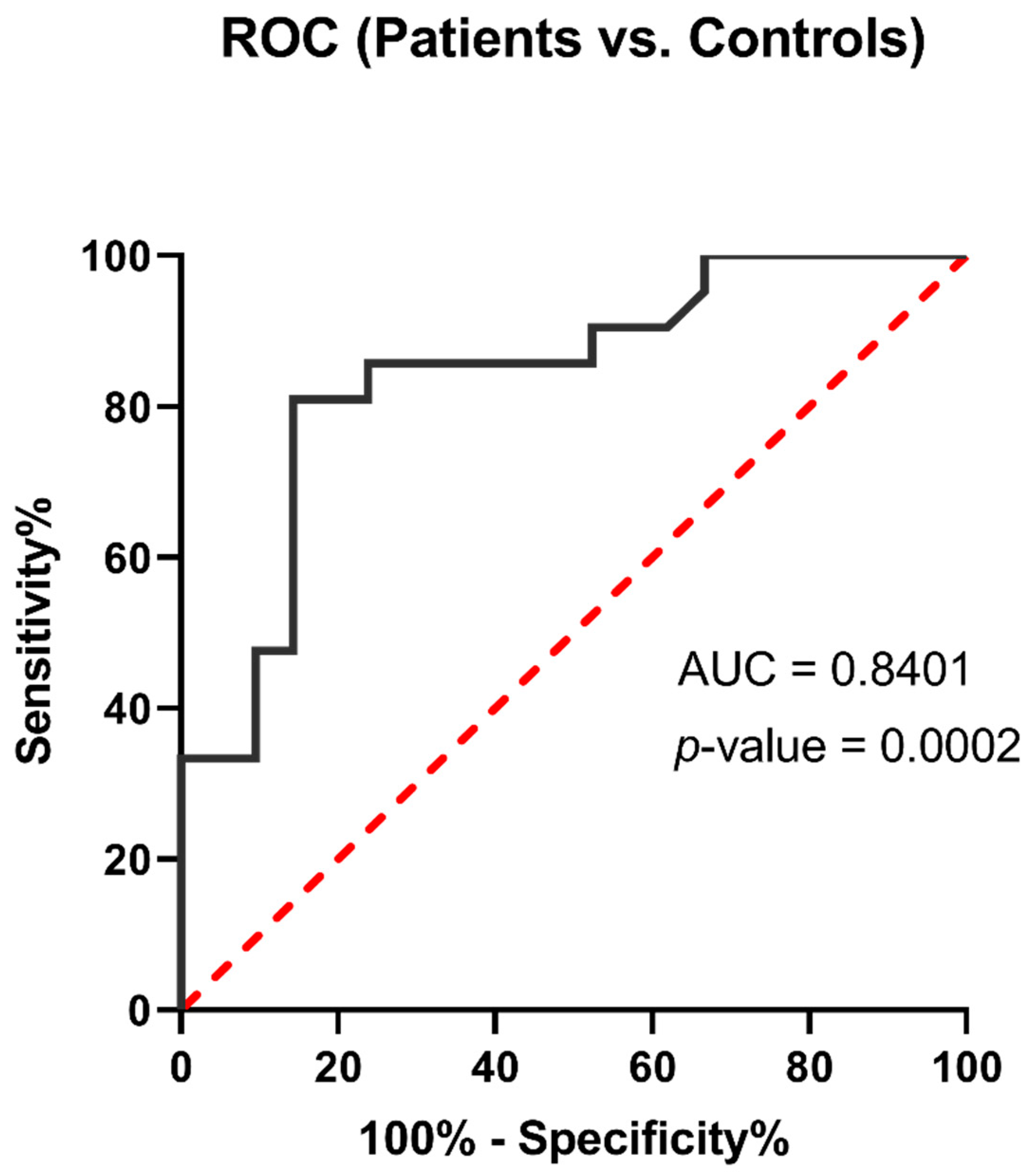
3.5. Arg1 Might Be a Promising Biomarker for COVID-19
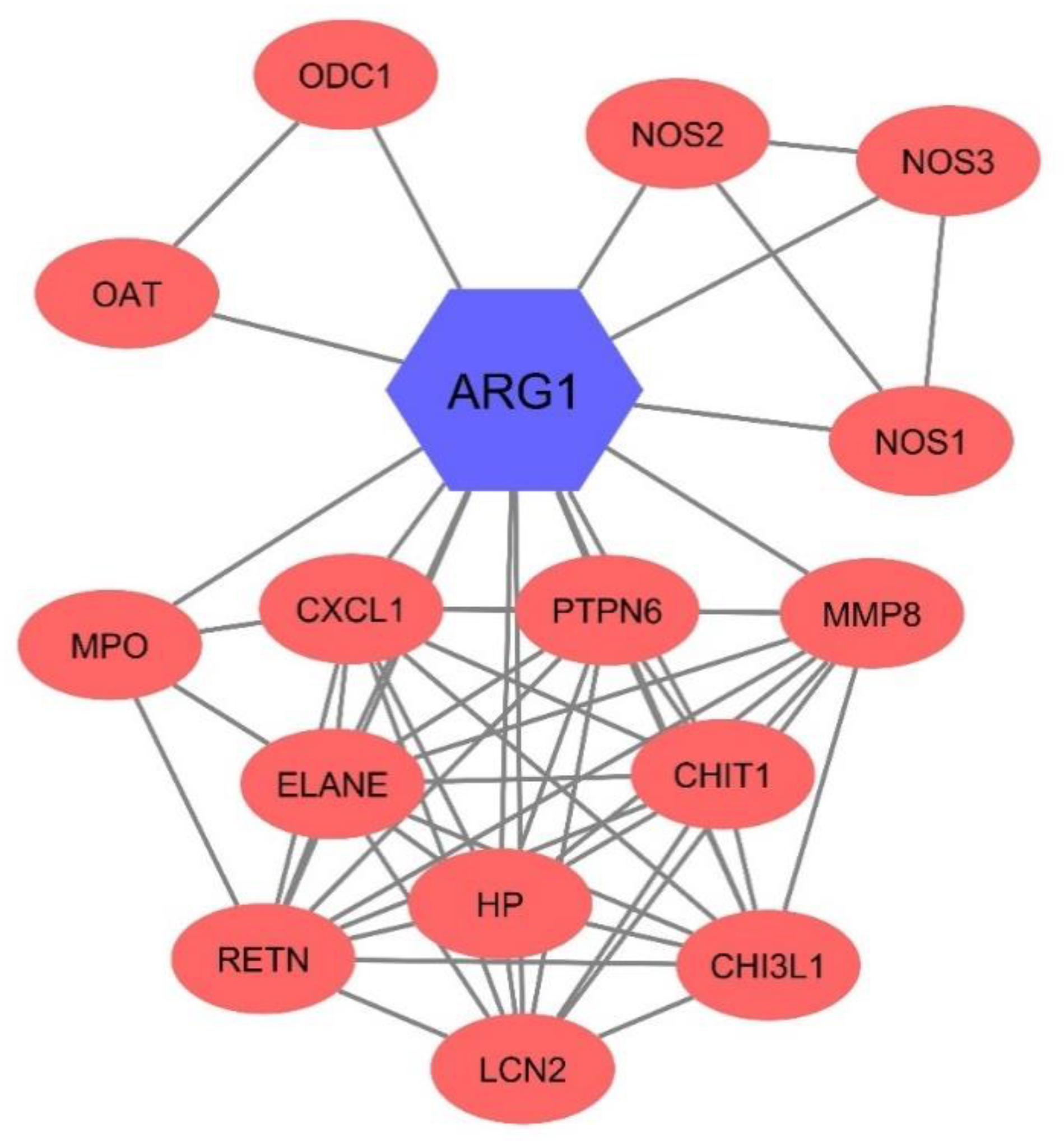
3.6. Arg1 Might Have Interacted with Other Enzymes in the Blood
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Maggi, E.; Canonica, G.W.; Moretta, L. COVID-19: Unanswered questions on immune response and pathogenesis. J. Allergy Clin. Immunol. 2020, 146, 18–22. [Google Scholar] [CrossRef]
- Ling, L.; Joynt, G.M.; Lipman, J.; Constantin, J.-M.; Joannes-Boyau, O. COVID-19: A critical care perspective informed by lessons learnt from other viral epidemics. Anaesth. Crit. Care Pain Med. 2020, 39, 163. [Google Scholar] [CrossRef]
- Calder, P.C. Nutrition, immunity and COVID-19. BMJ Nutr. Prev. Health 2020, 3, 74. [Google Scholar] [CrossRef] [PubMed]
- Burke, H.; Freeman, A.; Cellura, D.; Stuart, B.L.; Brendish, N.J.; Poole, S.; Borca, F.; Phan, H.T.; Sheard, N.; Williams, S. Inflammatory phenotyping predicts clinical outcome in COVID-19. Respir. Res. 2020, 21, 1–9. [Google Scholar] [CrossRef]
- Tang, Y.; Liu, J.; Zhang, D.; Xu, Z.; Ji, J.; Wen, C. Cytokine storm in COVID-19: The current evidence and treatment strategies. Front. Immunol. 2020, 11, 1708. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Liu, S.; Liu, J.; Zhang, Z.; Wan, X.; Huang, B.; Chen, Y.; Zhang, Y. COVID-19: Immunopathogenesis and Immunotherapeutics. Signal Transduct. Target. Ther. 2020, 5, 128. [Google Scholar] [CrossRef]
- Dandekar, A.A.; Perlman, S. Immunopathogenesis of coronavirus infections: Implications for SARS. Nat. Rev. Immunol. 2005, 5, 917–927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosales, C. Neutrophil: A cell with many roles in inflammation or several cell types? Front. Physiol. 2018, 9, 113. [Google Scholar] [CrossRef] [PubMed]
- Haick, A.K.; Rzepka, J.P.; Brandon, E.; Balemba, O.B.; Miura, T.A. Neutrophils are needed for an effective immune response against pulmonary rat coronavirus infection, but also contribute to pathology. J. Gen. Virol. 2014, 95, 578. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Du, X.; Chen, J.; Jin, Y.; Peng, L.; Wang, H.H.; Luo, M.; Chen, L.; Zhao, Y. Neutrophil-to-lymphocyte ratio as an independent risk factor for mortality in hospitalized patients with COVID-19. J. Infect. 2020, 81, e6–e12. [Google Scholar] [CrossRef]
- Athanasios, D. COVID-19 Hyperinflammation: What about Neutrophils? mSphere 2020, 5. [Google Scholar]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef] [Green Version]
- Grzywa, T.M.; Sosnowska, A.; Matryba, P.; Rydzynska, Z.; Jasinski, M.; Nowis, D.; Golab, J. Myeloid cell-derived Arginase in Cancer immune response. Front. Immunol. 2020, 11, 938. [Google Scholar] [CrossRef]
- Munder, M.; Schneider, H.; Luckner, C.; Giese, T.; Langhans, C.-D.; Fuentes, J.M.; Kropf, P.; Mueller, I.; Kolb, A.; Modolell, M. Suppression of T-cell functions by human granulocyte arginase. Blood 2006, 108, 1627–1634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burrack, K.S.; Tan, J.J.; McCarthy, M.K.; Her, Z.; Berger, J.N.; Ng, L.F.; Morrison, T.E. Myeloid cell Arg1 inhibits control of arthritogenic alphavirus infection by suppressing antiviral T cells. PLoS Pathog. 2015, 11, e1005191. [Google Scholar] [CrossRef]
- Munder, M. Arginase: An emerging key player in the mammalian immune system. Br. J. Pharmacol. 2009, 158, 638–651. [Google Scholar] [CrossRef] [Green Version]
- Zea, A.H.; Rodriguez, P.C.; Atkins, M.B.; Hernandez, C.; Signoretti, S.; Zabaleta, J.; McDermott, D.; Quiceno, D.; Youmans, A.; O’Neill, A. Arginase-producing myeloid suppressor cells in renal cell carcinoma patients: A mechanism of tumor evasion. Cancer Res. 2005, 65, 3044–3048. [Google Scholar] [CrossRef] [Green Version]
- Pesce, J.T.; Ramalingam, T.R.; Mentink-Kane, M.M.; Wilson, M.S.; El Kasmi, K.C.; Smith, A.M.; Thompson, R.W.; Cheever, A.W.; Murray, P.J.; Wynn, T.A. Arginase-1–expressing macrophages suppress Th2 cytokine–driven inflammation and fibrosis. PLoS Pathog. 2009, 5, e1000371. [Google Scholar] [CrossRef] [Green Version]
- Caldwell, R.W.; Rodriguez, P.C.; Toque, H.A.; Narayanan, S.P.; Caldwell, R.B. Arginase: A multifaceted enzyme important in health and disease. Physiol. Rev. 2018, 98, 641–665. [Google Scholar] [CrossRef] [Green Version]
- Tatum, D.; Taghavi, S.; Houghton, A.; Stover, J.; Toraih, E.; Duchesne, J. Neutrophil-to-lymphocyte ratio and outcomes in Louisiana Covid-19 patients. Shock 2020, 54, 652–658. [Google Scholar] [CrossRef]
- Hemmat, N.; Derakhshani, A.; Bannazadeh Baghi, H.; Silvestris, N.; Baradaran, B.; De Summa, S. Neutrophils, Crucial, or Harmful Immune Cells Involved in Coronavirus Infection: A Bioinformatics Study. Front. Genet. 2020, 11, 641. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Doncheva, N.T.; Morris, J.H.; Gorodkin, J.; Jensen, L.J. Cytoscape StringApp: Network analysis and visualization of proteomics data. J. Proteome Res. 2018, 18, 623–632. [Google Scholar] [CrossRef] [PubMed]
- García, L.F. Immune response, inflammation, and the clinical spectrum of COVID-19. Front. Immunol. 2020, 11, 1441. [Google Scholar] [CrossRef]
- Lotfinejad, P.; Asadzadeh, Z.; Najjary, S.; Somi, M.H.; Hajiasgharzadeh, K.; Mokhtarzadeh, A.; Derakhshani, A.; Roshani, E.; Baradaran, B. COVID-19 Infection: Concise Review Based on the Immunological Perspective. Immunol. Investig. 2020, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Shi, L.; Wang, Y.; Zhang, J.; Huang, L.; Zhang, C.; Liu, S.; Zhao, P.; Liu, H.; Zhu, L. Pathological findings of COVID-19 associated with acute respiratory distress syndrome. Lancet Respir. Med. 2020, 8, 420–422. [Google Scholar] [CrossRef]
- Liu, J.; Li, S.; Liu, J.; Liang, B.; Wang, X.; Wang, H.; Li, W.; Tong, Q.; Yi, J.; Zhao, L. Longitudinal characteristics of lymphocyte responses and cytokine profiles in the peripheral blood of SARS-CoV-2 infected patients. EBioMedicine 2020, 55, 102763. [Google Scholar] [CrossRef]
- Mez, J.; Daneshvar, D.H.; Kiernan, P.T.; Abdolmohammadi, B.; Alvarez, V.E.; Huber, B.R.; Alosco, M.L.; Solomon, T.M.; Nowinski, C.J.; McHale, L. Clinicopathological evaluation of chronic traumatic encephalopathy in players of American football. JAMA 2017, 318, 360–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Tan, Y.; Ling, Y.; Lu, G.; Liu, F.; Yi, Z.; Jia, X.; Wu, M.; Shi, B.; Xu, S. Viral and host factors related to the clinical outcome of COVID-19. Nature 2020, 583, 437–440. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Zhao, Z.-J.; Zhu, X.-Q.; Ren, Q.-S.; Nie, F.-F.; Gao, J.-M.; Gao, X.-J.; Yang, T.-B.; Zhou, W.-L.; Shen, J.-L. Differences in iNOS and arginase expression and activity in the macrophages of rats are responsible for the resistance against T. gondii infection. PLoS ONE 2012, 7, e35834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peranzoni, E.; Marigo, I.; Dolcetti, L.; Ugel, S.; Sonda, N.; Taschin, E.; Mantelli, B.; Bronte, V.; Zanovello, P. Role of arginine metabolism in immunity and immunopathology. Immunobiology 2008, 212, 795–812. [Google Scholar] [CrossRef]
- Santiago-Olivares, C.; Rivera-Toledo, E.; Gómez, B. Nitric oxide production is downregulated during respiratory syncytial virus persistence by constitutive expression of arginase 1. Arch. Virol. 2019, 164, 2231–2241. [Google Scholar] [CrossRef] [PubMed]
- Burrack, K.S.; Morrison, T.E. The role of myeloid cell activation and arginine metabolism in the pathogenesis of virus-induced diseases. Front. Immunol. 2014, 5, 428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoermer, K.A.; Burrack, A.; Oko, L.; Montgomery, S.A.; Borst, L.B.; Gill, R.G.; Morrison, T.E. Genetic ablation of arginase 1 in macrophages and neutrophils enhances clearance of an arthritogenic alphavirus. J. Immunol. 2012, 189, 4047–4059. [Google Scholar] [CrossRef] [PubMed]
- Cloots, R.H.; Sankaranarayanan, S.; Poynter, M.E.; Terwindt, E.; van Dijk, P.; Lamers, W.H.; Köhler, S.E. Arginase 1 deletion in myeloid cells affects the inflammatory response in allergic asthma, but not lung mechanics, in female mice. BMC Pulm. Med. 2017, 17, 158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rath, M.; Müller, I.; Kropf, P.; Closs, E.I.; Munder, M. Metabolism via arginase or nitric oxide synthase: Two competing arginine pathways in macrophages. Front. Immunol. 2014, 5, 532. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NO | Sex | Age | UCD | Common Symptoms | LRS | CVS | Other Symptoms | Status |
|---|---|---|---|---|---|---|---|---|
| 1 | Female | 68 | Heart and kidney failure | Pyrexia and pharyngitis | - | - | - | Dead |
| 2 | Male | 80 | - | Cough | Dyspnea | - | - | Alive |
| 3 | Female | 89 | - | - | Dyspnea | - | - | Dead |
| 4 | Male | 61 | - | Pharyngitis | Dyspnea | - | Muscular pain and loss of appetite | Alive |
| 5 | Male | 40 | Heart failure | Pyrexia | - | - | - | Alive |
| 6 | Male | 69 | Hypertension | Cough | Dyspnea | - | Muscular pain and loss of appetite | Dead |
| 7 | Female | 56 | - | Pyrexia and cough | Dyspnea | - | Hemoptysis | Alive |
| 8 | Male | 38 | Hypertension and diabetes | Pyrexia and cough | Dyspnea | - | Hemoptysis | Dead |
| 9 | Female | 60 | Hypertension | Pyrexia | - | - | Hemoptysis | Alive |
| 10 | Female | 43 | Hypertension | Pyrexia | - | Chest pain | Hemoptysis | Alive |
| 11 | Female | 62 | - | Pharyngitis | Dyspnea | Chest pain | Arthralgia and loss of appetite | Dead |
| 12 | Female | 64 | Hypertension and diabetes | Cough | Dyspnea | Chest pain | Muscular pain, loss of appetite, arthralgia, headache, and vomit | Dead |
| 13 | Female | 34 | - | - | Dyspnea | - | Chills and loss of appetite | Alive |
| 14 | Female | 45 | - | Cough | Dyspnea | - | - | Alive |
| 15 | Male | 73 | Hypertension | Cough | Dyspnea | - | Muscular pain | Alive |
| 16 | Male | 56 | Lung disease | Cough and pharyngitis | Dyspnea | Chest pain | Loss of appetite | Dead |
| 17 | Male | 88 | Nervous and hypertension | - | - | - | - | Alive |
| 18 | Female | 64 | Diabetes and hypertension | Cough | Dyspnea | - | - | Dead |
| 19 | Male | 81 | - | Cough | Dyspnea | - | Loss of appetite | Dead |
| 20 | Female | 32 | - | Cough | Dyspnea | - | Arthralgia and diarrhea | Alive |
| 21 | Male | 32 | - | Pyrexia | Dyspnea | - | - | Alive |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Derakhshani, A.; Hemmat, N.; Asadzadeh, Z.; Ghaseminia, M.; Shadbad, M.A.; Jadideslam, G.; Silvestris, N.; Racanelli, V.; Baradaran, B. Arginase 1 (Arg1) as an Up-Regulated Gene in COVID-19 Patients: A Promising Marker in COVID-19 Immunopathy. J. Clin. Med. 2021, 10, 1051. https://doi.org/10.3390/jcm10051051
Derakhshani A, Hemmat N, Asadzadeh Z, Ghaseminia M, Shadbad MA, Jadideslam G, Silvestris N, Racanelli V, Baradaran B. Arginase 1 (Arg1) as an Up-Regulated Gene in COVID-19 Patients: A Promising Marker in COVID-19 Immunopathy. Journal of Clinical Medicine. 2021; 10(5):1051. https://doi.org/10.3390/jcm10051051
Chicago/Turabian StyleDerakhshani, Afshin, Nima Hemmat, Zahra Asadzadeh, Moslem Ghaseminia, Mahdi Abdoli Shadbad, Golamreza Jadideslam, Nicola Silvestris, Vito Racanelli, and Behzad Baradaran. 2021. "Arginase 1 (Arg1) as an Up-Regulated Gene in COVID-19 Patients: A Promising Marker in COVID-19 Immunopathy" Journal of Clinical Medicine 10, no. 5: 1051. https://doi.org/10.3390/jcm10051051