Human Umbilical Vein Endothelial Cells (HUVECs) Co-Culture with Osteogenic Cells: From Molecular Communication to Engineering Prevascularised Bone Grafts

 , , ,
, , ,  , , and
, , and
Abstract
:1. Introduction
2. Cell Sources
2.1. Human Umbilical Vein Endothelial Cells (HUVECs)
- Cell morphology (polygonal shape);
- The detection of Weibel-Palade bodies under electron microscopy;
- The presence of smooth muscle actomyosins;
- The expression of ABO blood group antigens.
2.2. Other Sources of Endothelial Cells
2.3. Mesenchymal Stem Cells
- Adherence to plastic in standard culture conditions;
- Specific surface antigen expression, i.e., the presence of CD73, CD90, CD105 antigens, and lack of haematopoietic markers, inter alia CD34, CD45;
- Multilineage differentiation, which implies the ability of cells to differentiate into osteoblasts, chondroblasts and adipocytes with appropriate stimuli in vitro.
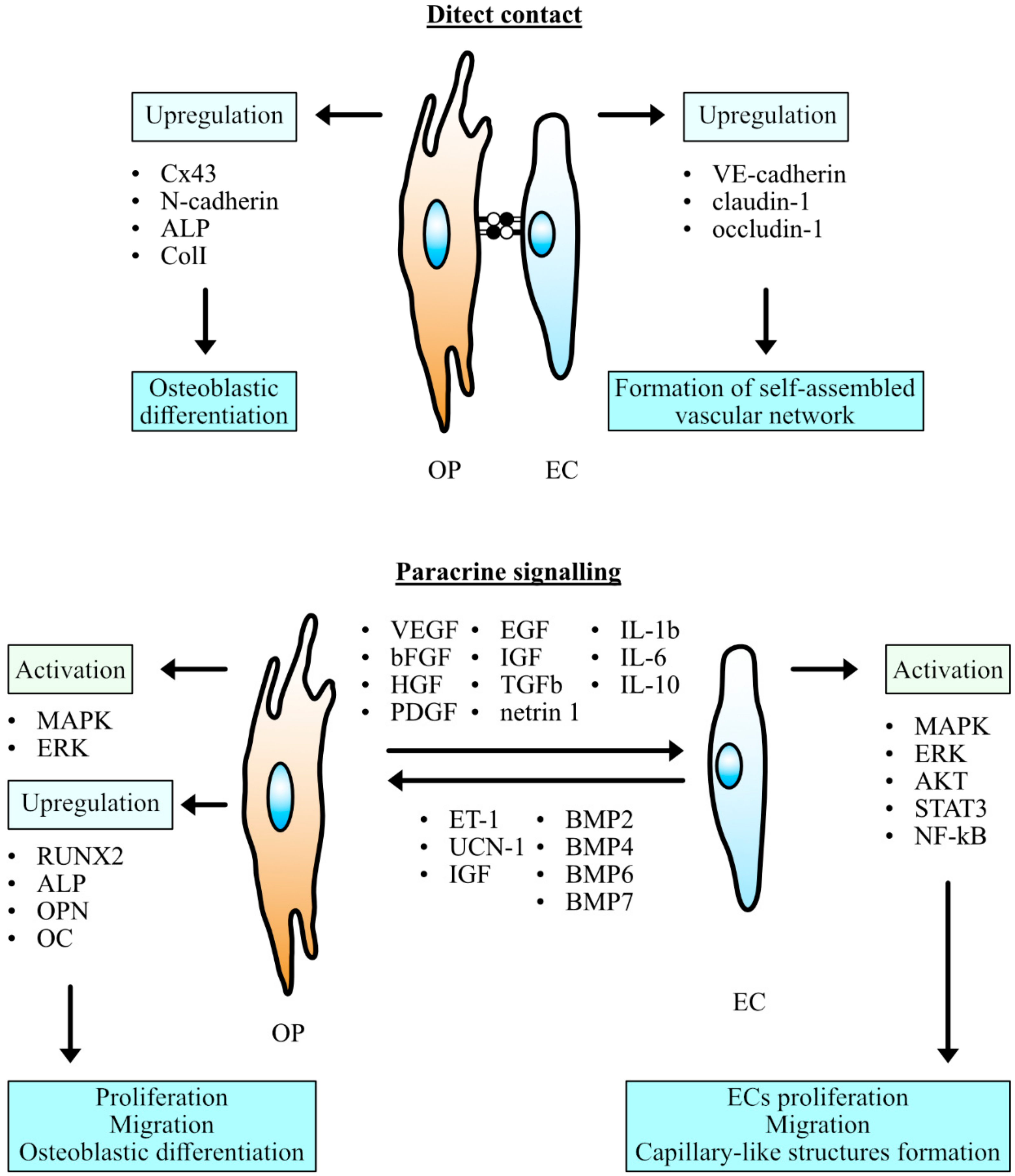
3. Cell Communication
3.1. Dynamics of Adherens Junctions in Co-Cultured Cells
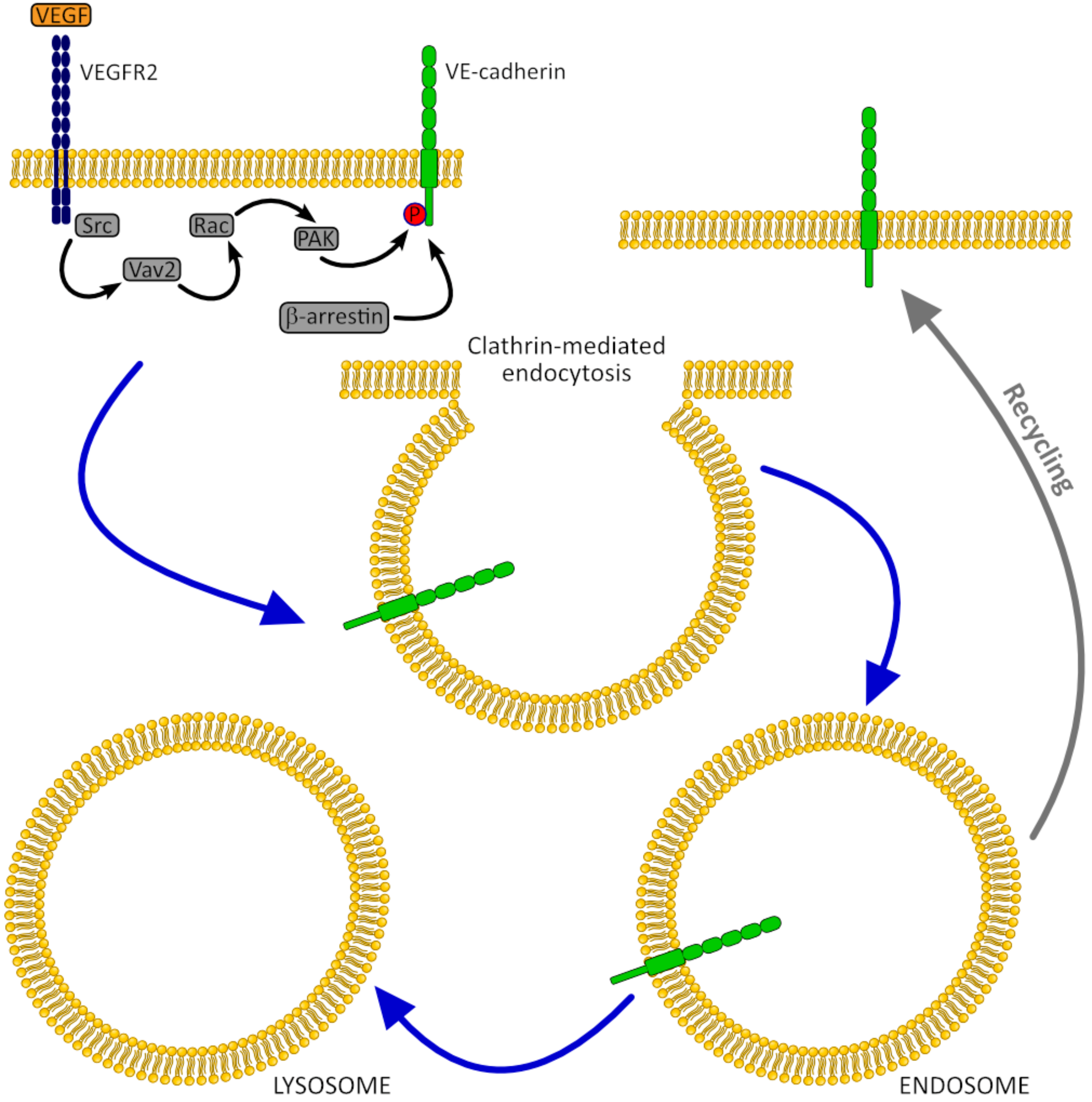
3.1.1. VE-Cadherin
3.1.2. N-Cadherin
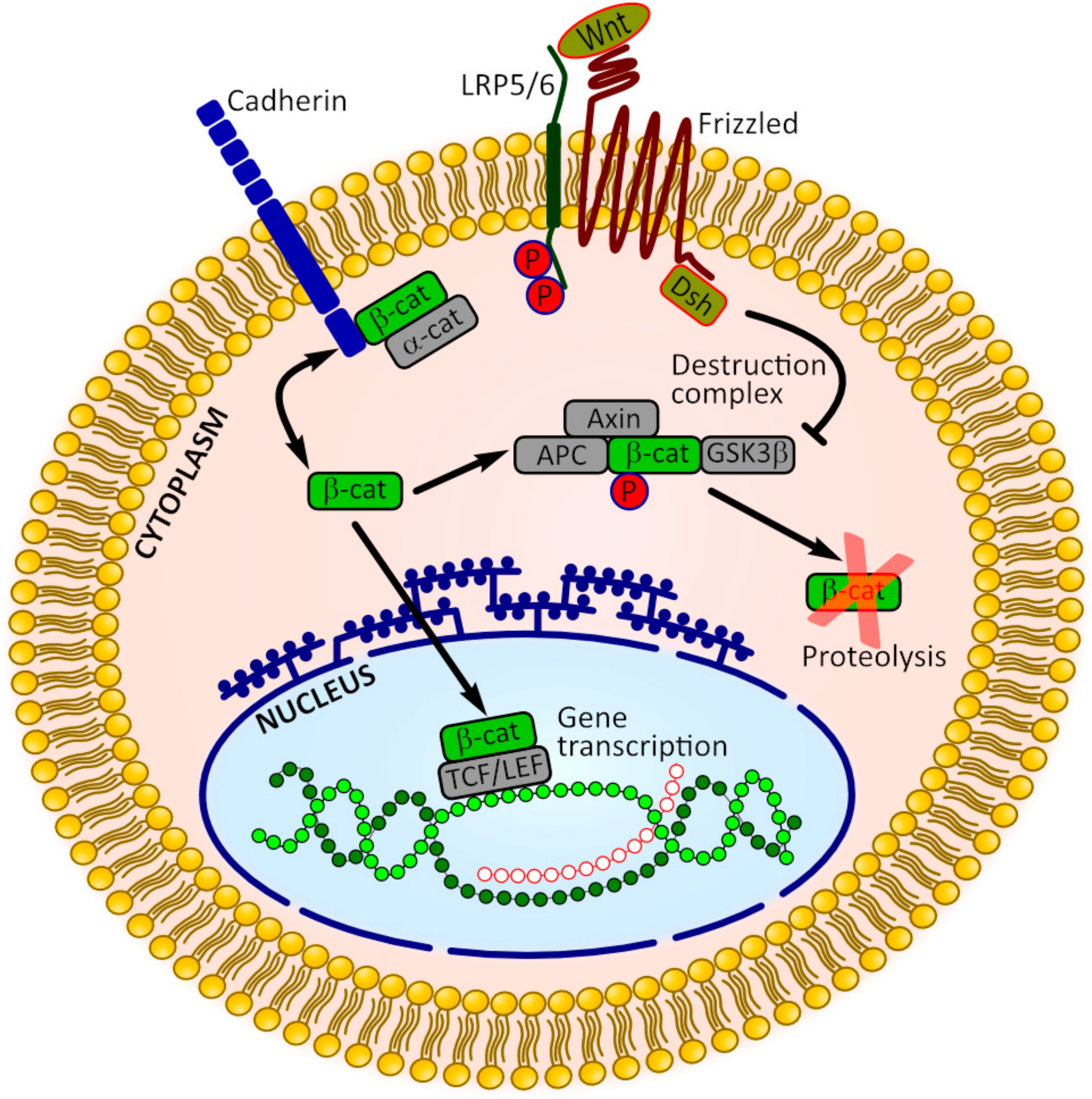
3.2. β-Catenin Signaling
3.3. Communication Through the Gap Junctions
3.4. Paracrine Action
3.5. Role of Extracellular Matrix in Cell-Cell Communication
4. Co-Culturing Systems and Optimization of Conditions
5. In Vivo Studies
6. Issues of Concern and Future Perspectives
Author Contributions
Conflicts of Interest
References
- Dimitriou, R.; Mataliotakis, G.I.; Angoules, A.G.; Kanakaris, N.K.; Giannoudis, P.V. Complications following autologous bone graft harvesting from the iliac crest and using the RIA: A systematic review. Injury 2011, 42, S3–S15. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.-C.; Chen, C.-Y.; Lin, K.-C.; Renn, J.-H.; Tarng, Y.-W.; Hsu, C.-J.; Chang, W.-N.; Yang, S.-W. Comparing morbidities of bone graft harvesting from the anterior iliac crest and proximal tibia: A retrospective study. J. Orthop. Surg. Res. 2018, 13, 115. [Google Scholar] [CrossRef] [PubMed]
- Myeroff, C.; Archdeacon, M. Autogenous Bone Graft: Donor Sites and Techniques. JBJS 2011, 93, 2227–2236. [Google Scholar] [CrossRef] [PubMed]
- Grabowski, G.; Robertson, R. Bone allograft with mesenchymal stem cells: A critical review of the literature. Hard Tissue 2012, 2. [Google Scholar] [CrossRef] [Green Version]
- Skovrlj, B.; Guzman, J.Z.; Al Maaieh, M.; Cho, S.K.; Iatridis, J.C.; Qureshi, S.A. Cellular bone matrices: Viable stem cell-containing bone graft substitutes. Spine J. 2014, 14, 2763–2772. [Google Scholar] [CrossRef] [PubMed]
- Grellier, M.; Bordenave, L.; Amédée, J. Cell-to-cell communication between osteogenic and endothelial lineages: Implications for tissue engineering. Trends Biotechnol. 2009, 27, 562–571. [Google Scholar] [CrossRef]
- Baiguera, S.; Ribatti, D. Endothelialization approaches for viable engineered tissues. Angiogenesis 2013, 16, 1–14. [Google Scholar] [CrossRef]
- Cenni, E.; Perut, F.; Baldini, N. In vitro models for the evaluation of angiogenic potential in bone engineering. Acta Pharmacol. Sin. 2011, 32, 21–30. [Google Scholar] [CrossRef]
- Unger, R.E.; Dohle, E.; Kirkpatrick, C.J. Improving vascularization of engineered bone through the generation of pro-angiogenic effects in co-culture systems. Adv. Drug Deliv. Rev. 2015, 94, 116–125. [Google Scholar] [CrossRef]
- Novosel, E.C.; Kleinhans, C.; Kluger, P.J. Vascularization is the key challenge in tissue engineering. Adv. Drug Deliv. Rev. 2011, 63, 300–311. [Google Scholar] [CrossRef]
- Rouwkema, J.; Rivron, N.C.; van Blitterswijk, C.A. Vascularization in tissue engineering. Trends Biotechnol. 2008, 26, 434–441. [Google Scholar] [CrossRef] [PubMed]
- Santos, M.I.; Reis, R.L. Vascularization in Bone Tissue Engineering: Physiology, Current Strategies, Major Hurdles and Future Challenges. Macromol. Biosci. 2010, 10, 12–27. [Google Scholar] [CrossRef] [PubMed]
- Lieberman, J.R.; Friedlaender, G.E. Bone Regeneration and Repair: Biology and Clinical Applications; Humana Press: Totowa, NJ, USA, 2005; ISBN 1592598633. [Google Scholar]
- Grosso, A.; Burger, M.G.; Lunger, A.; Schaefer, D.J.; Banfi, A.; Di Maggio, N. It Takes Two to Tango: Coupling of Angiogenesis and Osteogenesis for Bone Regeneration. Front. Bioeng. Biotechnol. 2017, 5, 68. [Google Scholar] [CrossRef] [PubMed]
- Correia, C.; Grayson, W.L.; Park, M.; Hutton, D.; Zhou, B.; Guo, X.E.; Niklason, L.; Sousa, R.A.; Reis, R.L.; Vunjak-Novakovic, G. In vitro model of vascularized bone: Synergizing vascular development and osteogenesis. PLoS ONE 2011, 6, e28352. [Google Scholar] [CrossRef]
- Ibrahim, M.; Richardson, M.K. Beyond organoids: In vitro vasculogenesis and angiogenesis using cells from mammals and zebrafish. Reprod. Toxicol. 2017, 73, 292–311. [Google Scholar] [CrossRef] [Green Version]
- Maruyama, Y. The human endothelial cell in tissue culture. Zeitschrift für Zellforschung und Mikroskopische Anatomie 1963, 60, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Nachman, R.L.; Jaffe, E.A. Endothelial cell culture: Beginnings of modern vascular biology. J. Clin. Investig. 2004, 114, 1037–1040. [Google Scholar] [CrossRef]
- Jaffe, E.A.; Nachman, R.L.; Becker, C.G.; Minick, C.R. Culture of Human Endothelial Cells Derived from Umbilical Veins. Identification by Morphologic and Immunologic Criteria. J. Clin. Investig. 1973, 52, 2745–2756. [Google Scholar] [CrossRef]
- Jaffe, E.A.; Hoyer, L.W.; Nachman, R.L. Synthesis of Antihemophilic Factor Antigen by Cultured Human Endothelial Cells. J. Clin. Investig. 1973, 52, 2757–2764. [Google Scholar] [CrossRef]
- WEIBEL, E.R. Fifty years of Weibel-Palade bodies: The discovery and early history of an enigmatic organelle of endothelial cells1. J. Thromb. Haemost. 2012, 10, 979–984. [Google Scholar] [CrossRef]
- Baudin, B.; Bruneel, A.; Bosselut, N.; Vaubourdolle, M. A protocol for isolation and culture of human umbilical vein endothelial cells. Nat. Protoc. 2007, 2, 481. [Google Scholar] [CrossRef] [PubMed]
- Jiménez, N.; Krouwer, V.J.D.; Post, J.A. A new, rapid and reproducible method to obtain high quality endothelium in vitro. Cytotechnology 2013, 65, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Davis, J.; Crampton, S.P.; Hughes, C.C.W. Isolation of Human Umbilical Vein Endothelial Cells (HUVEC). J. Vis. Exp. 2007, 3, e183. [Google Scholar] [CrossRef]
- Marin, V.; Kaplanski, G.; Grès, S.; Farnarier, C.; Bongrand, P. Endothelial cell culture: Protocol to obtain and cultivate human umbilical endothelial cells. J. Immunol. Methods 2001, 254, 183–190. [Google Scholar] [CrossRef]
- Caniuguir, A.; Krause, B.J.; Hernandez, C.; Uauy, R.; Casanello, P. Markers of early endothelial dysfunction in intrauterine growth restriction-derived human umbilical vein endothelial cells revealed by 2D-DIGE and mass spectrometry analyses. Placenta 2016, 41, 14–26. [Google Scholar] [CrossRef] [PubMed]
- Boerma, M.; Burton, G.R.; Wang, J.; Fink, L.M.; McGehee, R.E.; Hauer-Jensen, M. Comparative expression profiling in primary and immortalized endothelial cells: Changes in gene expression in response to hydroxy methylglutaryl-coenzyme A reductase inhibition. Blood Coagul. Fibrinolysis 2006, 17, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Patel, H.; Chen, J.; Das, K.C.; Kavdia, M. Hyperglycemia induces differential change in oxidative stress at gene expression and functional levels in HUVEC and HMVEC. Cardiovasc. Diabetol. 2013, 12, 142. [Google Scholar] [CrossRef] [PubMed]
- Walshe, T.E.; dela Paz, N.G.; D’Amore, P.A. The Role of Shear-Induced Transforming Growth Factor-β Signaling in the Endothelium. Arterioscler. Thromb. Vasc. Biol. 2013, 33, 2608–2617. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.; Jung, Y.; Kim, Y.; Jho, E.; Yoon, Y. LPS-induced inflammatory response is suppressed by Wnt inhibitors, Dickkopf-1 and LGK974. Sci. Rep. 2017, 7, 41612. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, J.G.; Wagner, F.; Martine, L.C.; Holzapfel, B.M.; Theodoropoulos, C.; Bas, O.; Savi, F.M.; Werner, C.; De-Juan-Pardo, E.M.; Hutmacher, D.W. Periosteum tissue engineering in an orthotopic in vivo platform. Biomaterials 2017, 121, 193–204. [Google Scholar] [CrossRef] [PubMed]
- Zhong, M.; Wei, D.; Yang, Y.; Sun, J.; Chen, X.; Guo, L.; Wei, Q.; Wan, Y.; Fan, H.; Zhang, X. Vascularization in Engineered Tissue Construct by Assembly of Cellular Patterned Micromodules and Degradable Microspheres. ACS Appl. Mater. Interfaces 2017, 9, 3524–3534. [Google Scholar] [CrossRef] [PubMed]
- Bersini, S.; Gilardi, M.; Arrigoni, C.; Talò, G.; Zamai, M.; Zagra, L.; Caiolfa, V.; Moretti, M. Human in vitro 3D co-culture model to engineer vascularized bone-mimicking tissues combining computational tools and statistical experimental approach. Biomaterials 2016, 76, 157–172. [Google Scholar] [CrossRef] [PubMed]
- Smith, E.E.; Zhang, W.; Schiele, N.R.; Khademhosseini, A.; Kuo, C.K.; Yelick, P.C. Developing a biomimetic tooth bud model. J. Tissue Eng. Regen. Med. 2017, 11, 3326–3336. [Google Scholar] [CrossRef] [PubMed]
- Hauser, S.; Jung, F.; Pietzsch, J. Human Endothelial Cell Models in Biomaterial Research. Trends Biotechnol. 2017, 35, 265–277. [Google Scholar] [CrossRef] [PubMed]
- Mishra, R.; Roux, B.M.; Posukonis, M.; Bodamer, E.; Brey, E.M.; Fisher, J.P.; Dean, D. Effect of prevascularization on in vivo vascularization of poly(propylene fumarate)/fibrin scaffolds. Biomaterials 2016, 77, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Pirosa, A.; Gottardi, R.; Alexander, P.G.; Tuan, R.S. Engineering in-vitro stem cell-based vascularized bone models for drug screening and predictive toxicology. Stem Cell Res. Ther. 2018, 9, 112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laranjeira, M.S.; Fernandes, M.H.; Monteiro, F.J. Reciprocal induction of human dermal microvascular endothelial cells and human mesenchymal stem cells: Time-dependent profile in a co-culture system. Cell Prolif. 2012, 45, 320–334. [Google Scholar] [CrossRef]
- Unger, R.E.; Ghanaati, S.; Orth, C.; Sartoris, A.; Barbeck, M.; Halstenberg, S.; Motta, A.; Migliaresi, C.; Kirkpatrick, C.J. The rapid anastomosis between prevascularized networks on silk fibroin scaffolds generated in vitro with cocultures of human microvascular endothelial and osteoblast cells and the host vasculature. Biomaterials 2010, 31, 6959–6967. [Google Scholar] [CrossRef] [PubMed]
- Sandhu, K.; Njoroge, W.; Yang, Y.; Harper, A.; Butler, R. Endothelial Progenitor Cell Identification, Classification and Nomenclature: A Review. J. Blood Lymph 2018, 8. [Google Scholar]
- Peters, E.B. Endothelial Progenitor Cells for the Vascularization of Engineered Tissues. Tissue Eng. Part B Rev. 2018, 24, 1–24. [Google Scholar] [CrossRef]
- Sharma, R.R.; Pollock, K.; Hubel, A.; McKenna, D. Mesenchymal stem or stromal cells: A review of clinical applications and manufacturing practices. Transfusion 2014, 54, 1418–1437. [Google Scholar] [CrossRef] [PubMed]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.S.; Deans, R.J.; Keating, A.; Prockop, D.J.; Horwitz, E.M. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef] [PubMed]
- Fitzsimmons, R.E.B.; Mazurek, M.S.; Soos, A.; Simmons, C.A. Mesenchymal Stromal/Stem Cells in Regenerative Medicine and Tissue Engineering. Stem Cells Int. 2018, 2018. [Google Scholar] [CrossRef] [PubMed]
- Dufrane, D. Impact of Age on Human Adipose Stem Cells for Bone Tissue Engineering. Cell Transplant. 2017, 26, 1496–1504. [Google Scholar] [CrossRef] [PubMed]
- Baker, N.; Boyette, L.B.; Tuan, R.S. Characterization of bone marrow-derived mesenchymal stem cells in aging. Bone 2015, 70, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Kornicka, K.; Houston, J.; Marycz, K. Dysfunction of Mesenchymal Stem Cells Isolated from Metabolic Syndrome and Type 2 Diabetic Patients as Result of Oxidative Stress and Autophagy may Limit Their Potential Therapeutic Use. Stem Cell Rev. Rep. 2018, 14, 337–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caplan, A.I. Review: Mesenchymal Stem Cells: Cell–Based Reconstructive Therapy in Orthopedics. Tissue Eng. 2005, 11, 1198–1211. [Google Scholar] [CrossRef] [PubMed]
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The Hallmarks of Aging. Cell 2013, 153, 1194–1217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyette, L.; Tuan, R.; Boyette, L.B.; Tuan, R.S. Adult Stem Cells and Diseases of Aging. J. Clin. Med. 2014, 3, 88–134. [Google Scholar] [CrossRef] [Green Version]
- Borys-Wójcik, S.; Brązert, M.; Jankowski, M.; Ożegowska, K.; Chermuła, B.; Piotrowska-Kempisty, H.; Bukowska, D.; Antosik, P.; Pawelczyk, L.; Nowicki, M.; et al. Human Wharton’s jelly mesenchymal stem cells: Properties, isolation and clinical applications. J. Biol. Regul. Homeost. Agents 2019, 33, 119–123. [Google Scholar] [PubMed]
- Rutkovskiy, A.; Stensløkken, K.-O.; Vaage, I.J. Osteoblast Differentiation at a Glance. Med. Sci. Monit. Basic Res. 2016, 22, 95–106. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Liu, X.; Chen, Q.; Bao, C.; Zhao, L.; Zhu, Z.; Xu, H.H.K. Angiogenic and osteogenic regeneration in rats via calcium phosphate scaffold and endothelial cell co-culture with human bone marrow mesenchymal stem cells (MSCs), human umbilical cord MSCs, human induced pluripotent stem cell-derived MSCs and human embry. J. Tissue Eng. Regen. Med. 2018, 12, 191–203. [Google Scholar] [CrossRef] [PubMed]
- Guillotin, B.; Bareille, R.; Bourget, C.; Bordenave, L.; Amédée, J. Interaction between human umbilical vein endothelial cells and human osteoprogenitors triggers pleiotropic effect that may support osteoblastic function. Bone 2008, 42, 1080–1091. [Google Scholar] [CrossRef] [PubMed]
- Marie, P.J.; Haÿ, E. Cadherins and Wnt signalling: A functional link controlling bone formation. Bonekey Rep. 2013, 2, 330. [Google Scholar] [CrossRef] [PubMed]
- Liebner, S.; Cavallaro, U.; Dejana, E. The multiple languages of endothelial cell-to-cell communication. Arterioscler. Thromb. Vasc. Biol. 2006, 26, 1431–1438. [Google Scholar] [CrossRef] [PubMed]
- Giannotta, M.; Trani, M.; Dejana, E. VE-cadherin and endothelial adherens junctions: Active guardians of vascular integrity. Dev. Cell 2013, 26, 441–454. [Google Scholar] [CrossRef] [PubMed]
- Ladoux, B.; Nelson, W.J.; Yan, J.; Mège, R.M. The mechanotransduction machinery at work at adherens junctions. Integr. Biol. 2015, 7, 1109–1119. [Google Scholar] [CrossRef] [PubMed]
- Yamada, S.; Pokutta, S.; Drees, F.; Weis, W.I.; Nelson, W.J. Deconstructing the cadherin-catenin-actin complex. Cell 2005, 123, 889–901. [Google Scholar] [CrossRef] [PubMed]
- Drees, F.; Pokutta, S.; Yamada, S.; Nelson, W.J.; Weis, W.I. Alpha-catenin is a molecular switch that binds E-cadherin-beta-catenin and regulates actin-filament assembly. Cell 2005, 123, 903–915. [Google Scholar] [CrossRef]
- Carmeliet, P.; Lampugnani, M.-G.; Moons, L.; Breviario, F.; Compernolle, V.; Bono, F.; Balconi, G.; Spagnuolo, R.; Oosthuyse, B.; Dewerchin, M.; et al. Targeted Deficiency or Cytosolic Truncation of the VE-cadherin Gene in Mice Impairs VEGF-Mediated Endothelial Survival and Angiogenesis. Cell 1999, 98, 147–157. [Google Scholar] [CrossRef] [Green Version]
- Harris, E.S.; Nelson, W.J. VE-cadherin: At the front, center, and sides of endothelial cell organization and function. Curr. Opin. Cell Biol. 2010, 22, 651–658. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Daculsi, R.; Grellier, M.; Bareille, R.; Bourget, C.; Remy, M.; Amedee, J. The role of vascular actors in two dimensional dialogue of human bone marrow stromal cell and endothelial cell for inducing self-assembled network. PLoS ONE 2011, 6, e16767. [Google Scholar] [CrossRef] [PubMed]
- Hebda, J.K.; Leclair, H.M.; Azzi, S.; Roussel, C.; Scott, M.G.; Bidère, N.; Gavard, J. The C-terminus region of β-arrestin1 modulates VE-cadherin expression and endothelial cell permeability. Cell Commun. Signal. 2013, 11, 37. [Google Scholar] [CrossRef]
- Ebrahim, N.A.; Leach, L. Temporal studies into attachment, VE-cadherin perturbation, and paracellular migration of human umbilical mesenchymal stem cells across umbilical vein endothelial monolayers. Stem Cells Dev. 2015, 24, 426–436. [Google Scholar] [CrossRef] [PubMed]
- Pati, S.; Khakoo, A.Y.; Zhao, J.; Jimenez, F.; Gerber, M.H.; Harting, M.; Redell, J.B.; Grill, R.; Matsuo, Y.; Guha, S.; et al. Human mesenchymal stem cells inhibit vascular permeability by modulating vascular endothelial cadherin/β-catenin signaling. Stem Cells Dev. 2011, 20, 89–101. [Google Scholar] [CrossRef]
- Gavard, J. Endothelial permeability and VE-cadherin: A wacky comradeship. Cell Adh. Migr. 2014, 8, 158–164. [Google Scholar] [CrossRef] [PubMed]
- Gavard, J.; Gutkind, J.S. VEGF controls endothelial-cell permeability by promoting the β-arrestin-dependent endocytosis of VE-cadherin. Nat. Cell Biol. 2006, 8, 1223–1234. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Daculsi, R.; Grellier, M.; Bareille, R.; Bourget, C.; Amedee, J. Role of neural-cadherin in early osteoblastic differentiation of human bone marrow stromal cells cocultured with human umbilical vein endothelial cells. Am. J. Physiol. Physiol. 2010, 299, C422–C430. [Google Scholar] [CrossRef] [Green Version]
- Marie, P.J. Role of N-cadherin in bone formation. J. Cell. Physiol. 2002, 190, 297–305. [Google Scholar] [CrossRef]
- Guntur, A.R.; Rosen, C.J.; Naski, M.C. N-cadherin adherens junctions mediate osteogenesis through PI3K signaling. Bone 2012, 50, 54–62. [Google Scholar] [CrossRef] [Green Version]
- Fontana, F.; Hickman-Brecks, C.L.; Salazar, V.S.; Revollo, L.; Abou-Ezzi, G.; Grimston, S.K.; Jeong, S.Y.; Watkins, M.; Fortunato, M.; Alippe, Y.; et al. N-cadherin Regulation of Bone Growth and Homeostasis Is Osteolineage Stage-Specific. J. Bone Miner. Res. 2017, 32, 1332–1342. [Google Scholar] [CrossRef] [PubMed]
- Marie, P.J.; Haÿ, E.; Modrowski, D.; Revollo, L.; Mbalaviele, G.; Civitelli, R. Cadherin-mediated cell-cell adhesion and signaling in the skeleton. Calcif. Tissue Int. 2014, 94, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Nelson, J.; Nusse, R. Convergence of Wnt, ß-catenin, and cadherin pathways. Science 2004, 303, 1483–1487. [Google Scholar] [CrossRef] [PubMed]
- Stains, J.P.; Civitelli, R. Cell-cell interactions in regulating osteogenesis and osteoblast function. Birth Defects Res. Part C Embryo Today Rev. 2005, 75, 72–80. [Google Scholar] [CrossRef]
- Menge, T.; Gerber, M.; Wataha, K.; Reid, W.; Guha, S.; Cox, C.S.; Dash, P.; Reitz, M.S.; Khakoo, A.Y.; Pati, S.; et al. Human mesenchymal stem cells inhibit endothelial proliferation and angiogenesis via cell-cell contact through modulation of the VE-Cadherin/β-catenin signaling pathway. Stem Cells Dev. 2013, 22, 148–157. [Google Scholar] [CrossRef] [PubMed]
- Giepmans, B. Gap junctions and connexin-interacting proteins. Cardiovasc. Res. 2004, 62, 233–245. [Google Scholar] [CrossRef] [Green Version]
- Okamoto, T.; Kawamoto, E.; Takagi, Y.; Akita, N.; Hayashi, T.; Park, E.J.; Suzuki, K.; Shimaoka, M. Gap junction-mediated regulation of endothelial cellular stiffness. Sci. Rep. 2017, 7, 6134. [Google Scholar] [CrossRef]
- Chen, C.-H.; Mayo, J.N.; Gourdie, R.G.; Johnstone, S.R.; Isakson, B.E.; Bearden, S.E. The connexin 43/ZO-1 complex regulates cerebral endothelial F-actin architecture and migration. Am. J. Physiol. Cell Physiol. 2015, 309, C600–C607. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Yu, L.; Xu, S.; Qiu, F.; Fan, Y.; Fu, G. Down-regulation of connexin43 gap junction by serum deprivation in human endothelial cells was improved by (−)-Epigallocatechin gallate via ERK MAP kinase pathway. Biochem. Biophys. Res. Commun. 2011, 404, 217–222. [Google Scholar] [CrossRef]
- Schiller, P.C.; D’Ippolito, G.; Brambilla, R.; Roos, B.A.; Howard, G.A. Inhibition of gap-junctional communication induces the trans-differentiation of osteoblasts to an adipocytic phenotype in vitro. J. Biol. Chem. 2001, 276, 14133–14138. [Google Scholar] [CrossRef]
- Villars, F.; Guillotin, B.; Amédée, T.; Dutoya, S.; Bordenave, L.; Bareille, R.; Amédée, J. Effect of HUVEC on human osteoprogenitor cell differentiation needs heterotypic gap junction communication. Am. J. Physiol. Physiol. 2002, 282, C775–C785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Zhou, Z.; Saunders, M.M.; Donahue, H.J. Modulation of connexin43 alters expression of osteoblastic differentiation markers. Am. J. Physiol. Physiol. 2006, 290, C1248–C1255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guillotin, B.; Bourget, C.; Remy-Zolgadri, M.; Bareille, R.; Fernandez, P.; Conrad, V.; Amédée-Vilamitjana, J. Human Primary Endothelial Cells Stimulate Human Osteoprogenitor Cell Differentiation. Cell. Physiol. Biochem. 2004, 14, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Leszczynska, J.; Zyzynska-Granica, B.; Koziak, K.; Ruminski, S.; Lewandowska-Szumiel, M. Contribution of endothelial cells to human bone-derived cells expansion in coculture. Tissue Eng. Part A 2013, 19, 393–402. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.S.; Miura, M.; Demura, H.; Sato, K. Anabolic Effects of 1,25-Dihydroxyvitamin D 3 on Osteoblasts Are Enhanced by Vascular Endothelial Growth Factor Produced by Osteoblasts and by Growth Factors Produced by Endothelial Cells 1. Endocrinology 1997, 138, 2953–2962. [Google Scholar] [CrossRef] [PubMed]
- Steiner, D.; Lampert, F.; Stark, G.B.; Finkenzeller, G. Effects of endothelial cells on proliferation and survival of human mesenchymal stem cells and primary osteoblasts. J. Orthop. Res. 2012, 30, 1682–1689. [Google Scholar] [CrossRef] [PubMed]
- Borges, L.E.; Bloise, E.; Dela Cruz, C.; Galleri, L.; Apa, R.; Petraglia, F.; Reis, F.M. Urocortin 1 expression and secretion by human umbilical vein endothelial cells: In vitro effects of interleukin 8, interferon γ, lipopolysaccharide, endothelin 1, prostaglandin F-2α, estradiol, progesterone and dexamethasone. Peptides 2015, 74, 64–69. [Google Scholar] [CrossRef]
- Edwards, S.S.; Zavala, G.; Prieto, C.P.; Elliott, M.; Martínez, S.; Egaña, J.T.; Bono, M.R.; Palma, V. Functional analysis reveals angiogenic potential of human mesenchymal stem cells from Wharton’s jelly in dermal regeneration. Angiogenesis 2014, 17, 851–866. [Google Scholar] [CrossRef] [PubMed]
- Shabbir, A.; Zisa, D.; Lin, H.; Mastri, M.; Roloff, G.; Suzuki, G.; Lee, T. Activation of host tissue trophic factors through JAK-STAT3 signaling: A mechanism of mesenchymal stem cell-mediated cardiac repair. Am. J. Physiol. Heart Circ. Physiol. 2010, 299, H1428–H1438. [Google Scholar] [CrossRef]
- Prockop, D.J. “Stemness” Does Not Explain the Repair of Many Tissues by Mesenchymal Stem/Multipotent Stromal Cells (MSCs). Clin. Pharmacol. Ther. 2007, 82, 241–243. [Google Scholar] [CrossRef]
- Jiang, W.; Tan, Y.; Cai, M.; Zhao, T.; Mao, F.; Zhang, X.; Xu, W.; Yan, Z.; Qian, H.; Yan, Y. Human Umbilical Cord MSC-Derived Exosomes Suppress the Development of CCl4-Induced Liver Injury through Antioxidant Effect. Stem Cells Int. 2018, 2018. [Google Scholar] [CrossRef] [PubMed]
- Gnecchi, M.; Zhang, Z.; Ni, A.; Dzau, V.J. Paracrine mechanisms in adult stem cell signaling and therapy. Circ. Res. 2008, 103, 1204–1219. [Google Scholar] [CrossRef] [PubMed]
- Park, K.-S.; Kim, Y.-S.; Kim, J.-H.; Choi, B.; Kim, S.-H.; Hyun-Kyung Tan, A.; Lee, M.-S.; Lee, M.-K.; Kwon, C.-H.; Joh, J.-W.; et al. Trophic Molecules Derived From Human Mesenchymal Stem Cells Enhance Survival, Function, and Angiogenesis of Isolated Islets After Transplantation. Transplantation 2010, 89, 509–517. [Google Scholar]
- Yu, B.; Zhang, X.; Li, X. Exosomes derived from mesenchymal stem cells. Int. J. Mol. Sci. 2014, 15, 4142–4157. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Tredget, E.E.; Wu, P.Y.G.; Wu, Y. Paracrine factors of mesenchymal stem cells recruit macrophages and endothelial lineage cells and enhance wound healing. PLoS ONE 2008, 3, e1886. [Google Scholar] [CrossRef]
- Prasadam, I.; Zhou, Y.; Du, Z.; Chen, J.; Crawford, R.; Xiao, Y. Osteocyte-induced angiogenesis via VEGF–MAPK-dependent pathways in endothelial cells. Mol. Cell. Biochem. 2014, 386, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Yang, S.; Zhang, Y.; Sun, Z.; Xu, W.; Ye, S. Co-culture of Mesenchymal Stem Cells with Umbilical Vein Endothelial Cells under Hypoxic Condition. J. Huazhong Univ. Sci. Technol. [Med. Sci.] 2012, 32, 137–180. [Google Scholar] [CrossRef]
- Prieto, C.P.; Ortiz, M.C.; Villanueva, A.; Villarroel, C.; Edwards, S.S.; Elliott, M.; Lattus, J.; Aedo, S.; Meza, D.; Lois, P.; et al. Netrin-1 acts as a non-canonical angiogenic factor produced by human Wharton’s jelly mesenchymal stem cells (WJ-MSC). Stem Cell Res. Ther. 2017, 8, 43. [Google Scholar] [CrossRef]
- Keshtkar, S.; Azarpira, N.; Ghahremani, M.H. Mesenchymal stem cell-derived extracellular vesicles: Novel frontiers in regenerative medicine. Stem Cell Res. Ther. 2018, 9, 63. [Google Scholar] [CrossRef]
- Rad, F.; Pourfathollah, A.A.; Yari, F.; Mohammadi, S.; Kheirandish, M. Microvesicles preparation from mesenchymal stem cells. Med. J. Islam. Repub. Iran. 2016, 30, 398. [Google Scholar]
- Shabbir, A.; Cox, A.; Rodriguez-Menocal, L.; Salgado, M.; Van Badiavas, E. Mesenchymal Stem Cell Exosomes Induce Proliferation and Migration of Normal and Chronic Wound Fibroblasts, and Enhance Angiogenesis In Vitro. Stem Cells Dev. 2015, 24, 1635–1647. [Google Scholar] [CrossRef] [PubMed]
- Gong, M.; Yu, B.; Wang, J.; Wang, Y.; Liu, M.; Paul, C.; Millard, R.W.; Xiao, D.-S.; Ashraf, M.; Xu, M. Mesenchymal stem cells release exosomes that transfer miRNAs to endothelial cells and promote angiogenesis. Oncotarget 2017, 8, 45200–45212. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Ma, Y.; Teng, R.; Guan, Q.; Lang, J.; Fang, J.; Long, H.; Tian, G.; Wu, Q. Transcriptional Profiling Reveals Crosstalk Between Mesenchymal Stem Cells and Endothelial Cells Promoting Prevascularization by Reciprocal Mechanisms. Stem Cells Dev. 2015, 24, 610–623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dreyfuss, J.L.; Regatieri, C.V.; Jarrouge, T.R.; Cavalheiro, R.P.; Sampaio, L.O.; Nader, H.B. Heparan sulfate proteoglycans: Structure, protein interactions and cell signaling. Ann. Braz. Acad. Sci. 2009, 81, 409–429. [Google Scholar] [CrossRef]
- Page-McCaw, A.; Ewald, A.J.; Werb, Z. Matrix metalloproteinases and the regulation of tissue remodelling. Nat. Rev. Mol. Cell Biol. 2007, 8, 221–233. [Google Scholar] [CrossRef] [PubMed]
- Theocharis, A.D.; Skandalis, S.S.; Gialeli, C.; Karamanos, N.K. Extracellular matrix structure. Adv. Drug Deliv. Rev. 2016, 97, 4–27. [Google Scholar] [CrossRef] [PubMed]
- Barker, T.H. The role of ECM proteins and protein fragments in guiding cell behavior in regenerative medicine. Biomaterials 2011, 32, 4211–4214. [Google Scholar] [CrossRef] [PubMed]
- Villars, F.; Bordenave, L.; Bareille, R.; Amedee, J. Effect of human endothelial cells on Human Bone Marrow Stromal Cell phenotype: Role of VEGF? J. Cell. Biochem. 2000, 79, 672–685. [Google Scholar] [CrossRef]
- Kang, Y.; Kim, S.; Bishop, J.; Khademhosseini, A.; Yang, Y. The osteogenic differentiation of human bone marrow MSCs on HUVEC-derived ECM and β-TCP scaffold. Biomaterials 2012, 33, 6998–7007. [Google Scholar] [CrossRef]
- Palmieri, D.; Valli, M.; Viglio, S.; Ferrari, N.; Ledda, B.; Volta, C.; Manduca, P. Osteoblasts extracellular matrix induces vessel like structures through glycosylated collagen I. Exp. Cell Res. 2010, 316, 789–799. [Google Scholar] [CrossRef]
- Simunovic, F.; Steiner, D.; Pfeifer, D.; Stark, G.B.; Finkenzeller, G.; Lampert, F. Increased extracellular matrix and proangiogenic factor transcription in endothelial cells after cocultivation with primary human osteoblasts. J. Cell. Biochem. 2013, 114, 1584–1594. [Google Scholar] [CrossRef] [PubMed]
- Lampert, F.M.; Simunovic, F.; Finkenzeller, G.; Pfeifer, D.; Stark, G.B.; Winninger, O.; Steiner, D. Transcriptomic Changes in Osteoblasts Following Endothelial Cell-Cocultivation Suggest a Role of Extracellular Matrix in Cellular Interaction. J. Cell. Biochem. 2016, 117, 1869–1879. [Google Scholar] [CrossRef] [PubMed]
- Shah, A.R.; Wenke, J.C.; Agrawal, C.M. Manipulation of Human Primary Endothelial Cell and Osteoblast Coculture Ratios to Augment Vasculogenesis and Mineralization. Ann. Plast. Surg. 2016, 77, 122–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Moor, L.; Merovci, I.; Baetens, S.; Verstraeten, J.; Kowalska, P.; Krysko, D.V.; De Vos, W.H.; Declercq, H. High-throughput fabrication of vascularized spheroids for bioprinting. Biofabrication 2018, 10, 035009. [Google Scholar] [CrossRef]
- Ma, J.; van den Beucken, J.J.J.P.; Yang, F.; Both, S.K.; Cui, F.-Z.; Pan, J.; Jansen, J.A. Coculture of osteoblasts and endothelial cells: Optimization of culture medium and cell ratio. Tissue Eng. Part C Methods 2011, 17, 349–357. [Google Scholar] [CrossRef] [PubMed]
- Bok, J.-S.; Byun, S.-H.; Park, B.-W.; Kang, Y.-H.; Lee, S.-L.; Rho, G.-J.; Hwang, S.-C.; Woo, D.K.; Lee, H.-J.; Byun, J.-H. The Role of Human Umbilical Vein Endothelial Cells in Osteogenic Differentiation of Dental Follicle-Derived Stem Cells in In Vitro Co-cultures. Int. J. Med. Sci. 2018, 15, 1160–1170. [Google Scholar] [CrossRef]
- Klein, D.; Weisshardt, P.; Kleff, V.; Jastrow, H.; Jakob, H.G.; Ergün, S. Vascular wall-resident CD44+ multipotent stem cells give rise to pericytes and smooth muscle cells and contribute to new vessel maturation. PLoS ONE 2011, 6, e20540. [Google Scholar] [CrossRef]
- Koike, N.; Fukumura, D.; Gralla, O.; Au, P.; Schechner, J.S.; Jain, R.K. Creation of long-lasting blood vessels. Nature 2004, 428, 138–139. [Google Scholar] [CrossRef]
- Au, P.; Tam, J.; Fukumura, D.; Jain, R.K. Bone marrow-derived mesenchymal stem cells facilitate engineering of long-lasting functional vasculature. Blood 2008, 111, 4551–4558. [Google Scholar] [CrossRef]
- Ma, J.; Yang, F.; Both, S.K.; Prins, H.J.; Helder, M.N.; Pan, J.; Cui, F.Z.; Jansen, J.A.; Van Den Beucken, J.J. In vitro and in vivo angiogenic capacity of BM-MSCs/HUVECs and AT-MSCs/HUVECs cocultures. Biofabrication 2014, 6. [Google Scholar] [CrossRef]
- Pirraco, R.P.; Iwata, T.; Yoshida, T.; Marques, A.P.; Yamato, M.; Reis, R.L.; Okano, T. Endothelial cells enhance the in vivo bone-forming ability of osteogenic cell sheets. Lab. Investig. 2014, 94, 663–673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aird, W.C. Phenotypic heterogeneity of the endothelium: I. Structure, function, and mechanisms. Circ. Res. 2007, 100, 158–173. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Wang, D.-A. In Situ Organ-Specific Vascularization in Tissue Engineering. Trends Biotechnol. 2018, 36, 834–849. [Google Scholar] [CrossRef] [PubMed]
- Stenman, J.M.; Rajagopal, J.; Carroll, T.J.; Ishibashi, M.; McMahon, J.; McMahon, A.P. Canonical Wnt signaling regulates organ-specific assembly and differentiation of CNS vasculature. Science 2008, 322, 1247–1250. [Google Scholar] [CrossRef] [PubMed]
- Oddo, A.; Peng, B.; Tong, Z.; Wei, Y.; Tong, W.Y.; Thissen, H.; Voelcker, N.H. Advances in Microfluidic Blood–Brain Barrier (BBB) Models. Trends Biotechnol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Adriani, G.; Ma, D.; Pavesi, A.; Kamm, R.D.; Goh, E.L.K. A 3D neurovascular microfluidic model consisting of neurons, astrocytes and cerebral endothelial cells as a blood–brain barrier. Lab. Chip 2017, 17, 448–459. [Google Scholar] [CrossRef] [PubMed]
- Lafage-Proust, M.-H.; Roche, B.; Langer, M.; Cleret, D.; Vanden Bossche, A.; Olivier, T.; Vico, L. Assessment of bone vascularization and its role in bone remodeling. Bonekey Rep. 2015, 4, 662. [Google Scholar] [CrossRef]
- Richardson, M.R.; Lai, X.; Witzmann, F.A.; Yoder, M.C. Venous and arterial endothelial proteomics: Mining for markers and mechanisms of endothelial diversity. Expert Rev. Proteom. 2010, 7, 823–831. [Google Scholar] [CrossRef] [PubMed]
- Spencer, J.A.; Ferraro, F.; Roussakis, E.; Klein, A.; Wu, J.; Runnels, J.M.; Zaher, W.; Mortensen, L.J.; Alt, C.; Turcotte, R.; et al. Direct measurement of local oxygen concentration in the bone marrow of live animals. Nature 2014, 508, 269–273. [Google Scholar] [CrossRef] [Green Version]
- Vali, P.; Lakshminrusimha, S. The Fetus Can Teach Us: Oxygen and the Pulmonary Vasculature. Children 2017, 4, 67. [Google Scholar] [CrossRef]
- Murray, M.; Michelle, L. Antepartal and Intrapartal Fetal Monitoring; Springer: Berlin/Heidelberg, Germany, 2007; ISBN 0826104835. [Google Scholar]
- Sen, C.K.; Semenza, G.L. Oxygen Sensing; Elsevier Academic Press: Cambridge, MA, USA, 2004; ISBN 0080497195. [Google Scholar]
- Addis, R.; Campesi, I.; Fois, M.; Capobianco, G.; Dessole, S.; Fenu, G.; Montella, A.; Cattaneo, M.G.; Vicentini, L.M.; Franconi, F. Human umbilical endothelial cells (HUVECs) have a sex: Characterisation of the phenotype of male and female cells. Biol. Sex. Differ. 2014, 5, 18. [Google Scholar] [CrossRef] [PubMed]
- Aird, W.C. Endothelium and Allotransplantation. Transplantation 2006, 82, S6–S8. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Pros | Cons |
|---|---|
| Non-invasive harvesting method from “medical waste” | Site-specific phenotype |
| Can be easily isolated in high numbers | Immunogenicity |
| A large number of published studies = comparable results | Impossibility of autotransplantation in adult patients |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kocherova, I.; Bryja, A.; Mozdziak, P.; Angelova Volponi, A.; Dyszkiewicz-Konwińska, M.; Piotrowska-Kempisty, H.; Antosik, P.; Bukowska, D.; Bruska, M.; Iżycki, D.; et al. Human Umbilical Vein Endothelial Cells (HUVECs) Co-Culture with Osteogenic Cells: From Molecular Communication to Engineering Prevascularised Bone Grafts. J. Clin. Med. 2019, 8, 1602. https://doi.org/10.3390/jcm8101602
Kocherova I, Bryja A, Mozdziak P, Angelova Volponi A, Dyszkiewicz-Konwińska M, Piotrowska-Kempisty H, Antosik P, Bukowska D, Bruska M, Iżycki D, et al. Human Umbilical Vein Endothelial Cells (HUVECs) Co-Culture with Osteogenic Cells: From Molecular Communication to Engineering Prevascularised Bone Grafts. Journal of Clinical Medicine. 2019; 8(10):1602. https://doi.org/10.3390/jcm8101602
Chicago/Turabian StyleKocherova, Ievgeniia, Artur Bryja, Paul Mozdziak, Ana Angelova Volponi, Marta Dyszkiewicz-Konwińska, Hanna Piotrowska-Kempisty, Paweł Antosik, Dorota Bukowska, Małgorzata Bruska, Dariusz Iżycki, and et al. 2019. "Human Umbilical Vein Endothelial Cells (HUVECs) Co-Culture with Osteogenic Cells: From Molecular Communication to Engineering Prevascularised Bone Grafts" Journal of Clinical Medicine 8, no. 10: 1602. https://doi.org/10.3390/jcm8101602