The Effects of Glyphosate and Its Commercial Formulations to Marine Invertebrates: A Review



Department of Biology, University of Padova, 35121 Padova, Italy
*
Author to whom correspondence should be addressed.
J. Mar. Sci. Eng. 2020, 8(6), 399; https://doi.org/10.3390/jmse8060399
Submission received: 4 May 2020
/
Revised: 20 May 2020
/
Accepted: 30 May 2020
/
Published: 1 June 2020
(This article belongs to the Special Issue Effects of Emerging Contaminants to Marine Organisms: In Vitro and In Vivo Studies)
Abstract
:Glyphosate is the active ingredient of numerous commercial formulations of herbicides applied in different sectors, from agriculture to aquaculture. Due to its widespread use around the world, relatively high concentrations of glyphosate have been detected in soil and aquatic environments. The presence of glyphosate in aquatic ecosystems has aroused the attention of researchers because of its potential negative effects on living organisms, both animals and plants. In this context, this review intends to summarize results of studies aimed at evaluating the effects of glyphosate (both as active ingredient and component of commercial formulations) on marine invertebrates. Generally, data obtained in acute toxicity tests indicate that glyphosate and its commercial formulations are lethal at high concentrations (not environmentally realistic), whereas results of long-lasting experiments indicate that glyphosate can markedly affect biological responses of marine invertebrates. Consequently, more efforts should be addressed at evaluating chronic or sub-chronic effects of such substances to marine invertebrate species.
1. Introduction
Glyphosate, also known as N-(phosphonomethyl) glycine [CAS registry number 1071-83-6], is a broad-spectrum organophosphate herbicide with a non-selective, post emergence, and systemic activity (Table 1). It is absorbed by leaves and translocated through the phloem in all plant districts [1], but plants can also uptake the herbicide from roots and translocate it through the xylem [2]. Glyphosate affects the shikimate pathways inhibiting the 5-enolpyruvyl-shikimate-3-phosphate synthase (EPSPS), an enzyme which catalyzes the penultimate step in the shikimate pathway [3,4]. The inhibition lead to a decrease of the three essential aminoacids, tyrosine, phenylalanine, and tryptophan, as well as a possible decrease of second metabolites such as flavonoids, lignin, and phytoalexins [5,6]. EPSPS is an enzyme present in all plants, both herbaceous and arboreal, in fungi, algae, and some microorganisms such as Apicomplexa on which glyphosate can also act [7,8,9,10]. In addition, glyphosate, which has chelating properties, may affect the plant bioavailability of several elements, both reducing their uptake or mobilizing it [11,12,13], but these aspects are still debated [5,6]. Also, glyphosate can reduce the resistance of plants to pathogens [6,14]. Besides glyphosate mainly being used in agriculture with about 1.2 million km2 of crop land treated annually alone in the United States (U.S.) [15], it is also applied in non-agricultural sectors such as forestry, urban, and resident weed control practices, in the control of aquatic weeds and along railroads [15,16,17,18].
Glyphosate’s “history” starts in 1950 when the swiss chemist Henri Martin discovered it. However, the herbicide reached the market only in 1974 [19]. In almost 50 years, several glyphosate-based herbicides (GBHs) have been developed (e.g., Accord, Aquamaster, Glyfos, Roundup, Rodeo, Touchdown) and over than 750 GBHs are registered in the U.S. and around 500 in Australia [20,21]. GBHs contain a wide spectrum of glyphosate salts, such as isopropylamine, diammonium, monoammonium, potassium, trimethylsulfonium, and sesquisodium [22]. Adjuvants, usually surfactants, are also present to enhance the glyphosate absorption trough the cuticle of the leaves, as commonly performed for several herbicides [23]. Adjuvants commonly present in GBHs include polyethoxylated tallow amine (POEA), quaternary ammonium compounds, polyoxyethylene alkyl ether phosphates, and alkyl polyglucoside, but usually they are listed as generic surfactants by the manufactures [24,25].
In terms of production, glyphosate is the most worldwide selling herbicide [26] and GBHs are used in more than 130 countries on more than 100 crops [27]. In the U.S., for example, glyphosate is the most commonly used pesticide in agriculture since 2001 and the second most used active ingredient in the non-agricultural sectors since 2005 [28]. In the U.S. agricultural sector its usage increased 300-fold from 1974 to 2014, particularly after the introduction of genetically herbicide resistant crops in 1990s with over 113 thousand tons used in 2014 [29]. As for Asian region, the main users of glyphosate are China (which is also the main glyphosate producer) and India [30]. Interestingly, glyphosate import increased by 177% in South Africa [31]. Globally, the estimated use of glyphosate (both agricultural and non-agricultural) was 56.296 tons in 1994 and has risen to 825.804 tons in 2014 [29] with an expected increase to 740–920 thousand tons in 2025 [32].
1.1. Environmental Fate of Glyphosate
The half-life of glyphosate in soil is affected by mineral and organic composition, climate conditions, and microbial activity [33,34,35,36]. In soil, the half-life values are in the order of 1.7–197 days [37], 30–174 days [38], 143.3 days [39], and 16.9–151 days [40], but in some case glyphosate remain detectable even after several months [41]. As for freshwater ecosystems, Annett et al. [42] reported a half-life of 7–142 days. Other authors recorded half-life values for surface waters between 1.5–16 days [43,44,45,46], while Mallat and Barceló [47] observed a half-life of 60–770 hours for groundwater. Surprisingly, Pizzarro et al. [48] report a range of 31.5-33.5 weeks. In seawater, the glyphosate half-life was estimated to be 47 ± 7 days at 25 °C under light condition, 267 ± 21 days at the same temperature but in the dark and, 315 ± 29 days in the dark at 31 °C [49].
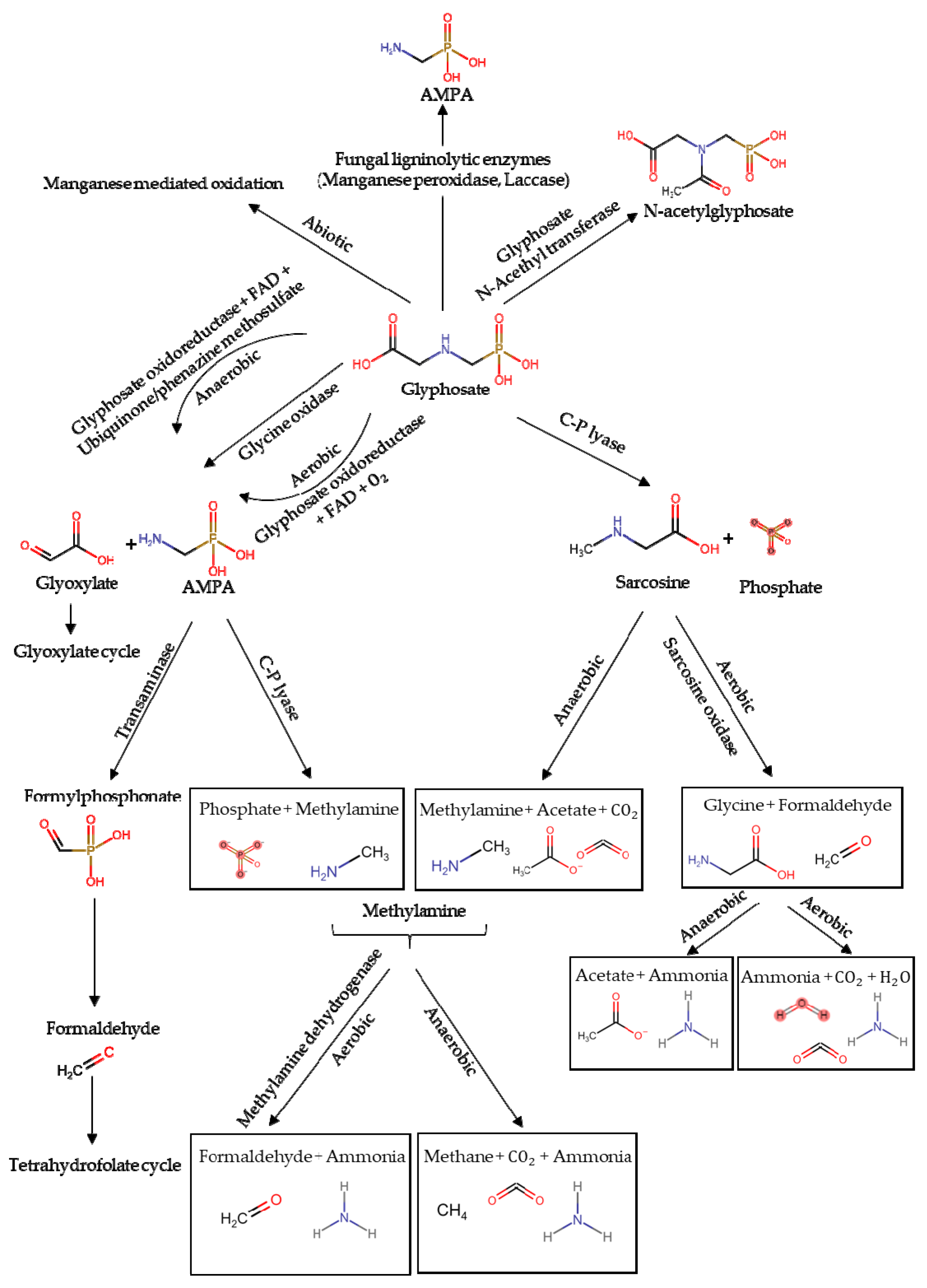
Environmental glyphosate degradation is mainly mediated by microbial activity [50], while the compound is more resistant to photolysis and chemical decomposition [38,47]. Moreover, an abiotic degradation pathway of glyphosate due to the manganese mineral birnessite, commonly present in soil, has also been proposed [51,52]. Bacteria and fungi degrade glyphosate through two main biochemical pathways [36,53,54]: C-P lyase pathway and AMPA pathway [6,38,55], allowing them to use the herbicide as a source of P, N and C [36,56] (Figure 1).
In the first degradation pathway glyphosate is oxidized in aerobic conditions by glyphosate oxidoreductase using FAD as cofactor and O2, or alternatively FAD and ubiquinone or phenazine methosulfate as electron acceptors in anaerobic conditions, making glyoxylate and its main metabolite aminomethylphosphonic acid (AMPA) [36]. Alternatively, also glycine oxidase degrades the herbicide to glyoxylate and AMPA [53]. Then, glyoxylate is used in the cellular glyoxylate cycle while AMPA can be excreted in the soil or even further biodegraded by C-P lyase to inorganic phosphate and methylamine. In aerobic conditions, methylamine is converted by methylamine dehydrogenase in ammonia and formaldehyde which is used in the tetrahydrofolate cycle. Methylamine can also be degraded anaerobically in ammonia, methane, and CO2 [38,54]. A different AMPA degradation pathway has been observed in which the compound is converted in formylphosphonate by a transaminase enzyme and then to formaldehyde [56,57]. In the second pathway the herbicide is degraded in sarcosine and inorganic phosphate by C-P lyase pathway trough several chemical reactions [58]. Sarcosine is then degraded by sarcosine oxidase in formaldehyde (see above for its degradation) and glycine which is commonly used in cellular cycles [57]. In aerobic conditions, glycine is then degraded in ammonia, water, and CO2, while in anerobic soil it is converted in acetate and ammonia. Alternately, sarcosine is converted in methylamine, acetate, and CO2 [54]. In a third pathway, glyphosate can be converted by glyphosate N-acetyltransferase in N-acetylglyphosate, but its further biodegradation remains unclear [53,59]. Finally, it has also been proposed that two fungi ligninolytic enzymes, laccase and manganese peroxidase, are able to degrade glyphosate in AMPA, but biochemical details remain unclear [60].
1.2. Occurrence of Glyphosate in Aquatic Environments
Despite the high affinity of glyphosate for soil particles and its consequent low mobility [61,62], it has been detected in a wide spectrum of water bodies (Table 2). Indeed, glyphosate can reach surface waters through either runoff and soil leaching or more rarely through a direct application into water (e.g., to control aquatic weeds) [15,36]. In groundwater, herbicide can occur due to karst phenomena, as reported in intensive agricultural areas, but it rarely reaches high levels in groundwater because it remains bind to soil particles [36]. Yang et al. [63], for example, reported that up to 14% of applied glyphosate can reach the water bodies due to runoff, while the European Food Safety Authority (EFSA) [64] has calculated a dissipation half-life (DT50) of 13.8–301 days in river.
Concentrations of glyphosate in water can vary along seasons and depends on rainfalls intensities [65,66,67,68] making it detectable not only in soil, but also in Wastewater Treatment Plants (WWTPs), agricultural areas surface water, groundwater, rivers and seawater [66,67,68,69,70,71,72,73,74,75,76,77,78,79,80,81,82,83,84,85,86,87,88,89,90,91,92,93,94,95,96,97,98,99,100,101,102,103,104,105,106,107,108,109,110,111,112,113,114,115,116]. According to Kemp et al. [73], who calculated that up to 2% of pesticide reach the marine environment, glyphosate has been observed in estuarine waters and seawaters [74,75,76]. In addition, glyphosate has also been detected in precipitations and phytotelmic water [69,72,77]. Surprisingly, despite its low vapor pressure, glyphosate has also been detected in atmosphere [78]. Overall, the environment concentrations are usually in order of few µg/L (see Table 2), much less than the worst scenario (up to 5.4 mg/L) [79]. However, high concentration values are detected in agricultural water bodies even up to hundreds of µg/L [66,80,81].
2. The Effects of Glyphosate to Marine Invertebrates
To our knowledge, most of the studies that have been performed to evaluate the toxic effects of glyphosate to non-target marine invertebrates are mainly related to mollusks (Table 3). However, in this review an attempt was also made to summarize the results obtained with other invertebrates.
2.1. Mollusks
Different species of marine mollusks have been used as model organisms to assess the effects of glyphosate and its commercial formulations on different levels of animal biological organization. For example, gametes and embryos from oysters (Crassostrea gigas) were used to assess the impact of glyphosate (as active ingredient) and Roundup® (commercial formulation) on population dynamics [117]. Three independent embryo-larval bioassays were conducted, but only one demonstrated that glyphosate (at 2.5 and 5 μg/L) can exert embryotoxic effects in term of increases of abnormal D-larvae percentage. Cumulative results of the three assays revealed a significant increase in the percentage of abnormal D-larvae only at 5 μg/L, whereas Roundup® did not affect significantly oyster embryos (as cumulative data from three independent bioassays). Neither glyphosate nor Roundup® exerted genotoxic effects on oyster spermatozoa.
In the same oyster species (C. gigas), the effects of long-term exposure (56 days) to glyphosate (0.1, 1, and 100 μg/L) were evaluated in juvenile oysters by means of different endpoints measured at both molecular and individual levels [118]. At the individual level, biomarkers (e.g., growth, condition index, sexual maturity, and tissue alterations) revealed a moderate effect of glyphosate. Neither mortality nor growth were detected during the study. Histological analyses revealed the absence of significant tissue alterations (atrophies of the wall of the digestive tubules and destructuration of the connective tissue), whereas a significant increase in hemocyte infiltration was detected between oysters at T0 and those at 56 days. Moderate effects of glyphosate were also recorded on oyster enzymatic activities, lipid peroxidation and expression of reference genes under the experimental conditions tested. In that study, the authors hypothesized that the low toxicity of glyphosate could be due to its chemical features, such as high-water solubility and moderate bioaccumulation potential [118].
The effects of different concentrations of the glyphosate-based herbicide Roundup Ready-To-Use Plus® (0.25, 1, 4, and 16 mg/L glyphosate) were assessed in the oyster Crassostrea virginica by means of sperm viability biomarkers [119]. Exposure for 20 min of C. virginica sperm to the herbicide did not alter significantly mitochondrial membrane potential in the sperm, as measured with MitoTracker Red CMXRos® [119]. Although cellular respiration did not decrease in sperm samples from the two bivalve species after Roundup exposure, the increase (not statistically significant for C. virginica) in the percentage of samples emitting high fluorescence intensity when exposed to 16 mg/L Roundup was probably due to plasma membrane damage, which in turn led to increased intracellular pH in the cytoplasm and polarization in the mitochondria [119].
A semi-quantitative multiplex RT-PCR (real-time polymerase chain reaction) method was used to evaluate mRNA expression of four different classes of glutathione S-transferases (GST)— pi, sigma, mu, and omega - in the Pacific oyster C. gigas exposed for 4 weeks to glyphosate (2 μg/L) [120]. Despite mu class of GST was not expressed in both gills and digestive gland from oysters, the expression of the remaining GSTs was markedly inhibited in gills, mainly after 30 days of exposure, while the expression generally increased in digestive gland [120]. In that study, the authors suggested that measurement of mRNA expression of pi and sigma class GST can be a useful biomarker of pesticide exposure in oysters. In the same oyster species, a suppression subtractive hybridization (SSH) method was used to reveal up and down-regulated genes following 30 days of exposure to glyphosate [121]. Identified genes from the SSH libraries resulted expressed differentially in gills and digestive gland from glyphosate-exposed oysters, compared to the control [121].
In the mussel Mytilus galloprovincialis, digestive gland transcriptional profiles were investigated through RNA-seq following exposure for 21 days to 10, 100, and 1000 μg/L [122]. A total of 111, 124, and 211 differentially regulated transcripts was found in mussels exposed to 10, 100, and 1000 μg/L, respectively. Five genes were shown to be differentially expressed at all glyphosate concentrations, including SERP2 (with a role in the protection of unfolded target proteins against degradation), GIMAP5 (an antiapoptotic protein) and MTMR14 (with a role in macroautophagy). At the functional level, that study revealed that several important biological functions, such as cell signaling, energy metabolism, and Ca2+ homeostasis were compromised, even at the lowest concentrations tested [122]. In the parallel study, we demonstrated that exposure for 7, 14, and 21 days to glyphosate (10, 100, and 1000 μg/L) affected significantly hemocyte parameters, such as total hemocyte count (THC, decreased at 7 and 14 days), hemocyte volume (increased at 7 and 21 days) hemolymph pH (increased after 14 days of exposure), hemocyte lysate lysozyme (reduced at 7, 14, and 21 days), and acid phosphatase (decreased at 7 days, increased at 14, and reduced again at 21 days) activities and cell membrane stability (lactate dehydrogenase activity increased in cell-free hemolymph of mussels exposed for 7 and 14 days) [123]. Conversely, exposure to glyphosate did not markedly alter antioxidant enzyme activities in both gills and digestive gland [123]. Overall, that study demonstrated that glyphosate affected mostly hemocyte functionality. Hemocyte parameters were also measured in the clam Ruditapes philippinarum to confirm the effects of glyphosate at the cellular level [124]. In that study, exposure for 7 days to glyphosate (10, 100, and 1000 μg/L) was shown to significantly reduce THC, whereas hemocyte diameter and volume increased significantly in treated clams. In addition, hemocyte proliferation and hemocyte lysate acid phosphatase activity increased significantly in glyphosate-exposed clams. That study demonstrated further that hemocytes (circulating cells that play a key role in immune defenses) are a target for glyphosate action in marine bivalves.
Although glyphosate is not considered an acetylcholinesterase (AChE) inhibitor, some in vitro and in vivo studies demonstrated that such compound may affect neurotransmission in mollusks. For example, pure glyphosate concentrations (from 0.075 to 15 mM) were shown to inhibit cholinesterase activity in vitro in a concentration-dependent manner in gills and muscle from the marine brown Perna perna, inhibition of enzyme activity reaching more than 50% even at the lowest concentrations tested (0.75–1 mM) [125]. In our in vivo study, exposure for 7, 14, and 21 days of M. galloprovincialis to glyphosate was shown to reduce significant AChE activity in gills, even if a non-linear response of enzyme activity was recorded. Indeed, AChE activity decreased at 7 days, increased at 14 days, and decreased after 21 days of exposure [123]. The increase in AChE activity in mussels exposed for 14 days was probably a response of mussels to face the decrease at 7 days, whereas the pronounced inhibition of AChE at 21 days indicated inability of mussels to cope with the herbicide.
At the organism level, the effects of the glyphosate-based herbicide Roundup Express® (REX) containing adjuvants, such as polyethoxylated tallow amines (POEAs), have been evaluated in C. gigas juveniles [126]. Exposure for 35 days to three concentrations (0.1, 1, and 100 μg/L) of REX did not cause significant effects on mortality rates, histological biomarkers, and condition index (CI), whereas shell length decreased significantly after 35 days of exposure to 0.1 and 100 μg/L, as well as the whole weight of oysters treated for 14 days with the lowest concentration tested. Interestingly, a delay in gametogenesis was recorded in oysters exposed for 35 days to 0.1 and 1 μg/L. As for digestive gland biomarkers, that study revealed no significant effects of REX on total protein content, catalase (CAT), and GST activities and lipid peroxidation. The authors suggested that biomarkers measured at the individual level, such as shell growth and reproduction, are more responsive to chronic exposure to herbicides [126].
A comparative in vitro study on the effects of glyphosate (as active ingredient) and REX (commercial formulation) has been performed on hemocytes from the abalone Haliotis tuberculata, a marine gastropod species [127]. Several concentrations (from 0.1 to 100,000 μg/L) of both the substances were tested and the effects on cell viability, phagocytosis, and lysosomal membrane stability were evaluated following exposure for 72 h of hemocytes. Cell viability was not affected by glyphosate, even at the highest concentration tested (100,000 μg/L), whereas exposure to REX induced a significant decrease in hemocyte viability at 40,000 μg/L, reaching a dramatic reduction (about 7%) at 100,000 μg/L. Similarly, glyphosate did not affect phagocytic activity of hemocytes, whereas it decreased significantly in hemocyte treated with REX (from 10,000 μg/L). As for lysosomal membrane stability, that study demonstrated that the lowest concentration of glyphosate (0.1 μg/L)—but not the two highest (10,000 and 100,000 μg/L)—affected significantly lysosomal stability, when compared to controls. An opposite response was observed after REX exposure, with a significant decrease in neutral red retention in lysosomes of hemocytes treated with the highest concentrations tested (from 10,000 μg/L) [127]. Overall, the study demonstrated that the commercial formulation can be more harmful than the active ingredient for hemocytes, probably due to the presence of adjuvants, such as POEAs, used to increase glyphosate efficacy towards plants.
In another study, the effects of Roundup® 3plus on metabolic parameters of heart of the clam Ruditapes decussatus were evaluated by means of HRMAS NMR spectroscopy following exposure for 24 and 72 h to two doses of herbicide (0.2 and 1 g/L) [128]. As a result, exposure to the herbicide caused anaerobiosis and alterations of oxidative metabolism, as revealed by alteration in energy metabolism and metabolic biomarkers (e.g., alanine, succinate, acetate, and propionate). That study demonstrated usefulness of HRMAS NMR technique in investigating mechanisms of action of Roundup in marine bivalves.
Recently, potential estrogenic effects of a new glyphosate-based formulation, namely Roundup® Power 2.0, were evaluated in M. galloprovincialis [129]. Mussels were exposed for 7, 14, and 21 days to two concentrations of Roundup® Power 2.0, corresponding to 100 and 1000 µg/L of glyphosate. In that study, no significant alterations in vitellogenin (Vtg) gene expression were recorded in female gonads, whereas a significant decrease in alkali labile phosphate (ALP, an indirect method for Vtg measurement) levels was observed in female gonads exposed for 21 days and in males exposed for 7 days. Interestingly, ALP levels increased significantly in gonads from males exposed for 21 days. Results demonstrated that the glyphosate-based formulation can affect reproduction-related parameters in mussels.
2.2. Other Marine Invertebrates
To the best of our knowledge, information concerning the effects of glyphosate or its commercial formulations to other marine invertebrates are limited to few taxonomic groups (Table 4).
As for crustacea, attention has been paid to the acute toxicity of glyphosate on different life stages of organisms. For example, the acute toxicity of two glyphosate-based formulations, namely Roundup® Original and Glyphosate AKB 480 (AKB), on early life stages of Artemia salina have been investigated [130]. Exposure of nauplii for 48h to 5, 10, 25, 50, and 100 mg/L (corresponding to 1.8, 3.6, 9, 18, and 36 mg/L of glyphosate) to both glyphosate-based herbicides induced significant increases in mortality rate, even if Roundup® was more toxic than AKB, 48-h LC50 values being 14.19 mg of glyphosate acid equivalent/L and 37.53 mg/L of glyphosate acid equivalent, respectively. Conversely, 48-h LC50 values of 1.77 mg/L, 49.3 mg/L and 35.3 mg/L were recorded after 48 h of exposure of Acartia tonsa to Roundup®, glyphosate acid and isopropylamine salt of glyphosate [131]. In the blue crab Callinectes sapidus, the acute toxicity of Roundup® Pro on crab megalopae and J1–J4 stage juveniles was investigated following exposure for 24h [132]. The resulting LC50 values were 6279 μg/L for megalopae and 316,000 μg/L for juveniles, indicating that the post-larval stage is more sensitive than juveniles to Roundup® Pro. Overall, results of the three studies above [130,131,132] suggest that differences in LC50 values can be due partially at least to different compounds tested and the species used along their development stage (nauplii of A. salina, adults of A. Tonsa, megalopae and juveniles of C. sapidus). Exposure for 90 days during the pre-reproductive period (winter) of females of the estuarine crab Neohelice granulata to glyphosate (0.02, 0.2, and 1 mg/L) caused a decrease in body weight at all the concentrations tested, whereas no alterations in gonadosomatic index and vitellogenic protein content of the ovary were recorded. However, a significantly increased percentage of reabsorbed vitellogenic oocytes was observed in crabs exposed to 1 mg/L, suggesting that glyphosate can affect both somatic and the ovarian growth in crabs [133]. In the same estuarine crab species, exposure of females for 90 days to Roundup Ultramax® (0.01 and 0.2 mg/L, acid equivalent) induced a significant increase in glycemia, while glycogen content in the muscle did not change significantly [134]. In addition, no significant effects of Roundup Ultramax® on gonadosomatic index were found, whereas a significantly higher percentage of reabsorbed vitellogenic oocyte and a significant decrease of vitellogenin content in the ovary were recorded at 0.2 mg/L. That study demonstrated that environmentally realistic concentrations of Roundup Ultramax® can affect significantly important reproduction-related parameters of crabs, such as ovary vitellogenin content and oocyte maturation [134]. Interestingly, the same group of researchers demonstrated that both glyphosate and its commercial formulation Roundup Ultramax® affected some important biological parameters in males of N. granulate [135]. Exposure for 30 days to 1 mg/L of both the compounds caused a significant decrease in weight and total proteins in muscle, and a significant increase in muscle glycogen content. Abnormal spermatophores—partially empty of spermatozoa—was observed following exposure to the two compounds. The authors concluded that 1-month exposure of crabs to glyphosate and its commercial formulation can affect the reproductive performance of male animals, by producing abnormal spermatophores and reducing sperm count [135].
Gametes and early life stages of echinoderms are widely considered as important targets to evaluate the effects of contaminants on the reproductive success of such invertebrates. In this regard, exposure of sperm from the sea urchin Lytechinus variegatus to different concentrations (0.25, 1, 4, and 16 mg/L) of Roundup Ready To-Use-Plus® did not cause significant alterations in mitochondrial membrane potential of sperm samples stained with MitoTracker. Sperm from only one animal treated with 16 mg/L showed a statistically significant variation in membrane integrity [119]. At concentrations ranging from 0 (control) to 200 μg/L, glyphosate did not affect significantly larval development of the sea urchin Paracentrotus lividus [136]. In addition, it was not possible to determine EC50 (median effective concentration) value, suggesting that glyphosate was not toxic to P. lividus larvae, at the concentrations tested at least [136].
In the ecotoxicological context, a growing concern is because the effects of contaminants can be exacerbated by variations in abiotic environmental factors, such as temperature. In this regard, Amid et al. [137] evaluated the combined effects of glyphosate and increased temperature to the tropical staghorn coral Acropora formosa. Branches of corals collected from both polluted and reference sites were exposed to different concentrations of glyphosate (0.12, 1.2, 6, and 12 mg/L) under two temperatures values, 28 °C ambient (ambient) and 31 °C (elevated temperature), and the effects on chlorophyll a content, zooxanthellae densities, and degree of bleaching were evaluated. Before exposure to increased temperature and glyphosate, corals from the reference site underwent bleaching, whereas those from the polluted site were more tolerant. Neither temperature nor glyphosate alone affected significantly coral pigmentation and chlorophyll a, whereas their interaction (temperature*glyphosate) had a significant effect on loss of color and chlorophyll a, mainly at the highest temperature and glyphosate levels. Results suggested that bleaching of corals can be promoted by different stressors occurring concomitantly.
Lastly, there is only one study in which a polychaete species was used to test glyphosate toxicity, to our knowledge at least. The species is the estuarine polychaeta Laeonereis acuta [138]. First, in that study the 96-h LC50 value for Roundup was determined, it resulting to be 8.19 mg/L. Animals were then exposed for 24 h and 96 h to 3.25 mg/L (corresponding to the no observed effect concentration (NOEC)) and 5.35 mg/L (LC10) and oxygen consumption of animals, as well as various biomarkers were measured in three body regions (anterior, middle and posterior) of polychaetes. Roundup did not cause significant alterations in oxygen consumption, whereas exposure induced a significant decrease in AChE activity in animals exposed to both Roundup concentrations. Significant reductions in reactive oxygen species (ROS) levels were recorded only in the posterior region of animals exposed for 24 h to Roundup concentrations, whereas a significant decrease in the antioxidant capacity against peroxyl radicals (ACAP) was observed in the three body regions of animals following exposure to the highest concentration of Roundup. No significant alterations in CAT, GST, and glutathione peroxidase (GPx) enzyme activities, along with reduced glutathione (GSH) levels, were recorded after exposure to Roundup. Conversely, superoxide dismutase (SOD) activity varied significantly depending on both animal regions and Roundup concentrations. Lastly, lipid peroxidation decreased significantly in all regions of animals exposed for 24 h to the highest concentration tested, whereas lipid damage generally increased in middle and posterior regions of polychaetes exposed for 96 h. Overall, results of that study demonstrated that Roundup can be seriously toxic to L. acuta, as alterations in ROS and ACAP levels, as well as in AChE activity demonstrated [138].
3. Conclusions and Perspectives
Although information concerning the levels of glyphosate in the marine environment is limited, the results of the studies that have been summarized in this review clearly indicate that this substance can cause undesirable effects on marine organisms, at different levels of biological organization. The growing interest in the potential risk posed by glyphosate to human health has also played an important role in the development of ecotoxicological studies aimed at understanding the negative effects of such substance to non-target organisms. Considering that the use of this herbicide is still permitted in many countries, further studies are necessary for a more in-depth assessment of the risk that glyphosate and its commercial formulations can pose to non-target marine organisms. In this context, it is important to highlight that data obtained in acute toxicity tests (few hours) indicate that glyphosate and its commercial formulations are generally lethal at high levels, that are not environmentally realistic. Conversely, information obtained following more prolonged exposure (several weeks) suggest that glyphosate can markedly affect biological responses of marine invertebrates. Consequently, efforts should be addressed at evaluating chronic or sub-chronic effects of such substances to other species of marine invertebrates.
Author Contributions
V.M. designed and supervised the present manuscript. V.M. and J.F. wrote the manuscript. M.G.M. contributed to the interpretation of the results available in the literature. The final version of the manuscript has been approved by all authors. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Acknowledgments
The authors thank the three anonymous reviewers for their constructive comments, which helped us to improve the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Shaner, L.D. Role of translocation as a mechanism of resistance to glyphosate. Weed Sci. 2009, 57, 118–123. [Google Scholar] [CrossRef]
- Saunders, L.E.; Pezeshki, R. Glyphosate in runoff waters and in the root-zone: A review. Toxics 2015, 3, 462–480. [Google Scholar] [CrossRef] [Green Version]
- Steinrücken, H.; Amrhein, N. The herbicide glyphosate is a potent inhibitor of 5-enolpyruvylshikimic acid-3-phosphate synthase. Biochem. Biophys. Res. Commun. 1980, 94, 1207–1212. [Google Scholar] [CrossRef]
- Schönbrunn, E.; Eschenburg, S.; Shuttleworth, W.A.; Schloss, J.V.; Amrhein, N.; Evans, J.N.S.; Kabsch, W. Interaction of the herbicide glyphosate with its target enzyme 5-enolpyruvylshikimate 3-phosphate synthase in atomic detail. PNAS 2001, 98, 1376–1380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mertens, M.; Höss, S.; Neumann, G.; Afzal, J.; Reichenbecher, W. Glyphosate, a chelating agent-relevant for ecological risk assessment? Environ. Sci. Pollut. Res. 2018, 25, 5298–5317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duke, S.O.; Lydon, J.; Koskinen, W.C.; Moorman, T.B.; Chaney, R.L.; Hammerschmidt, R. Glyphosate effects on plant mineral nutrition, crop rhizosphere microbiota, and plant disease in glyphosate-resistant crops. J. Agric. Food. Chem. 2012, 60, 10375–10397. [Google Scholar] [CrossRef]
- Herrmann, K.M. The shikimate pathway as an entry to aromatic secondary metabolism. Plant Physiol. 1995, 107, 7–12. [Google Scholar] [CrossRef] [Green Version]
- Richards, T.A.; Dacks, J.B.; Campbell, S.A.; Blanchard, J.L.; Foster, P.G.; McLeod, R.; Roberts, C.W. Evolutionary origins of the eukaryotic shikimate pathway: Gene fusions, horizontal gene transfer, and endosymbiotic replacements. Eukaryot. Cell 2006, 5, 1517–1531. [Google Scholar] [CrossRef] [Green Version]
- Roberts, F.; Roberts, C.W.; Johnson, J.J.; Kyle, D.E.; Krell, T.; Coggins, J.R.; Coombs, G.H.; Milhous, W.K.; Tzipori, S.; Ferguson, D.J.; et al. Evidence for the shikimate pathway in apicomplexan parasites. Nature 1998, 393, 801–805. [Google Scholar] [CrossRef]
- Tohge, T.; Watanabe, M.; Hoefgen, R.; Fernie, A.R. Shikimate and phenylalanine biosynthesis in the green lineage. Front. Plant Sci. 2013, 4, 62. [Google Scholar] [CrossRef] [Green Version]
- Cakmak, I.; Yazici, A.; Tutus, Y.; Ozturk, L. Glyphosate reduced seed and leaf concentrations of calcium, manganese, magnesium, and iron in non-glyphosate resistant soybean. Eur. J. Agron. 2009, 31, 114–119. [Google Scholar] [CrossRef] [Green Version]
- Barrett, K.A.; McBride, M.B. Trace element mobilization in soils by glyphosate. Soil Sci. Soc. Am. J. 2006, 70, 1882–1888. [Google Scholar] [CrossRef]
- Eker, S.; Ozturk, L.; Yazici, A.; Erenoglu, B.; Romheld, V.; Cakmak, I. Foliar-applied glyphosate substantially reduced uptake and transport of iron and manganese in sunflower (Helianthus annuus L.) plant. J. Agric. Food Chem. 2006, 54, 10019–10025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johal, G.S.; Huber, D.M. Glyphosate effects on diseases of plants. Eur. J. Agron. 2009, 31, 144–152. [Google Scholar] [CrossRef]
- EPA, United States Environmental Protection Agency Washington, D.C., 20460. Glyphosate-Response to Comments Usage and Benefits–Final. 2019. Available online: https://www.epa.gov/sites/production/files/2019-04/documents/glyphosate-response-comments-usage-benefits-final.pdf (accessed on 1 May 2020).
- Rolando, C.A.; Baillie, B.R.; Thompson, D.G.; Little, K.M. The risks associated with glyphosate-based herbicide use in planted forests. Forests 2017, 8, 208. [Google Scholar] [CrossRef] [Green Version]
- Torstensson, L.; Börjesson, E.; Stenström, J. Efficacy and fate of glyphosate on Swedish railway embankments. Pest. Manag. Sci. 2005, 61, 881–886. [Google Scholar] [CrossRef]
- Tang, T.; Boënne, W.; Desmet, N.; Seuntjens, P.; Bronders, J.; van Griensven, A. Quantification and characterization of glyphosate use and loss in a residential area. Sci. Total Environ. 2015, 517, 207–214. [Google Scholar] [CrossRef]
- Duke, S.O.; Powles, S.B. Glyphosate: A once-in-a-century herbicide. Pest. Manag. Sci. 2008, 64, 319–325. [Google Scholar] [CrossRef]
- Valavanidis, A. Glyphosate, the Most Widely Used Herbicide. Health and Safety Issues. Why Scientists Differ in Their Evaluation of Its Adverse Health Effects. Scientific Reviews. 2018. Available online: www.chem-tox-ecotox.org/ScientificReviews (accessed on 1 May 2020).
- APVMA. Final Pesticide and Veterinary Medicines Product Sales 2016–2017; Commonwealth of Australia Gazette No APVMA 6; Australian Pesticides and Veterinary Medicines Authority: Canberra, Australia, 2018. Available online: https://apvma.gov.au/node/10756 (accessed on 26 May 2020).
- Travlos, I.; Cheimona, N.; Bilalis, D. Glyphosate Efficacy of Different Salt Formulations and Adjuvant Additives on Various Weeds. Agronomy 2017, 7, 60. [Google Scholar] [CrossRef] [Green Version]
- Hess, F.; Foy, C. Interaction of Surfactants with Plant Cuticles. Weed Technol. 2000, 14, 807–813. [Google Scholar] [CrossRef]
- Defarge, N.; Takács, E.; Lozano, V.L.; Mesnage, R.; Spiroux de Vendômois, J.; Séralini, G.E.; Székács, A. Co-formulants in glyphosate-based herbicides disrupt aromatase activity in human cells below toxic levels. Int. J. Environ. Res. Public Health 2016, 13, 264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mesnage, R.; Benbrook, C.; Antoniou, M.N. Insight into the confusion over surfactant co-formulants in glyphosate-based herbicides. Food Chem. Toxicol. 2019, 128, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Baylis, A.D. Why glyphosate is a global herbicide: Strengths, weaknesses and prospects. Pest. Manag. Sci. 2000, 56, 299–308. [Google Scholar] [CrossRef]
- Monsanto. Backgrounder History of Monsanto’s Glyphosate Herbicides. 2002. Available online: http://www.bingeiagung.com/files/back_history.pdf (accessed on 31 May 2020).
- Grube, A.H.; Donaldson, D.; Kiely, T.; Wu, L. Pesticides Industry Sales and Usage: 2006 and 2007 Market Estimates; Off. Pestic. Programs, US Environ. Prot. Agency: Washington, DC, USA, 2011. [Google Scholar]
- Benbrook, C.M. Trends in glyphosate herbicide use in the United States and globally. Environ. Sci. Eur. 2016, 28, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richmond, M.E. Glyphosate: A review of its global use, environmental impact, and potential health effects on humans and other species. J. Environ. Stud. Sci. 2018, 8, 416–434. [Google Scholar] [CrossRef]
- African Centre for Biosafety. Glyphosate in SA: Risky Pesticide at Large and Unregulated in Our Soil. 2015. Available online: https://www.acbio.org.za/en/glyphosate-sa-risky-pesticide-large-and-unregulated-our-soil-and-water (accessed on 31 May 2020).
- Maggi, F.; Tang, F.H.M.; la Cecilia, D.; McBratney, A. PEST-CHEMGRIDS, global gridded maps of the top 20 crop-specific pesticide application rates from 2015 to 2025. Sci. Data 2019, 6, 170. [Google Scholar] [CrossRef] [Green Version]
- Sørensen, S.R.; Schultz, A.; Jacobsen, O.S.; Aamand, J. Sorption, desorption and mineralisation of the herbicides glyphosate and MCPA in samples from two Danish soil and subsurface profiles. Environ. Pollut. 2006, 141, 184–194. [Google Scholar] [CrossRef]
- von Wirén-Lehr, S.; Komoßa, D.; Gläßgen, W.E.; Sandermann, H.; Scheunert, I., Jr. Mineralization of [14C] glyphosate and its plant-associated residues in arable soils originating from different farming systems. Pestic. Sci. 1997, 51, 436–442. [Google Scholar] [CrossRef]
- Helander, M.; Saloniemi, I.; Saikkonen, K. Glyphosate in northern ecosystems. Trends Plant Sci. 2012, 17, 56–574. [Google Scholar] [CrossRef]
- Borggaard, O.K.; Gimsing, A.L. Fate of glyphosate in soil and the possibility of leaching to ground and surface waters: A review. Pest. Manag. Sci. 2008, 64, 441–456. [Google Scholar] [CrossRef]
- Giesy, J.P.; Dobson, S.; Solomon, K.R. Ecotoxicological risk assessment for Roundup® herbicide. Rev. Environ. Toxicol. 2000, 167, 35–120. [Google Scholar]
- Singh, B.K.; Walker, A. Microbial degradation of organophosphorus compounds. FEMS Microbiol. Rev. 2006, 30, 428–471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- EFSA. Glyphosate renewal assessment report of 18 December 2013. Rapporteur Member State (RMS): Germany, Co-RMS: Slovakia. 2013. Available online: http://www.pan-germany.org/download/Glyphosat-Studie_Campact_PAN_korrigiert.pdf (accessed on 31 May 2020).
- Bergström, L.; Börjesson, E.; Stenström, J. Laboratory and lysimeter studies of glyphosate and aminomethylphosphonic acid in a sand and a clay soil. J. Environ. Qual. 2011, 40, 98–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laitinen, P.; Rämö, S.; Nikunen, U.; Jauhiainen, L.; Siimes, K.; Turtola, E. Glyphosate and phosphorus leaching and residues in boreal sandy soil. Plant Soil 2009, 323, 267–283. [Google Scholar] [CrossRef]
- Annett, R.; Habibi, H.R.; Hontela, A. Impact of glyphosate and glyphosate-based herbicides on the freshwater environment. J. Appl. Toxicol. 2014, 34, 458–479. [Google Scholar] [CrossRef]
- Vera, M.S.; Lagomarsino, L.; Sylvester, M.; Pérez, G.L.; Rodríguez, P.; Mugni, H.; Sinistro, R.; Ferraro, M.; Bonetto, C.; Zagarese, H.; et al. New evidences of Roundup (glyphosate formulation) impact on the periphyton community and the water quality of freshwater ecosystems. Ecotoxicology 2010, 19, 710–721. [Google Scholar] [CrossRef]
- Vera, M.S.; Di Fiori, E.; Lagomarsino, L.; Sinistro, R.; Escaray, R.; Iummato, M.M.; Juárez, A.; Ríos de Molina, M.; Tell, G.; Pizarro, H. Direct and indirect effects of the glyphosate formulation Glifosato Atanor® on freshwater microbial communities. Ecotoxicology 2012, 21, 1805–1816. [Google Scholar] [CrossRef]
- Pérez, G.L.; Torremorell, A.; Mugni, H.; Rodríguez, P.; Solange Vera, M.; do Nascimento, M.; Allende, L.; Bustingorry, J.; Escaray, R.; Ferraro, M.; et al. Effects of the herbicide Roundup on freshwater microbial communities: A mesocosm study. Ecol. Appl. 2007, 17, 2310–2322. [Google Scholar] [CrossRef] [Green Version]
- Goldsborough, L.G.; Beck, A.E. Rapid dissipation of glyphosate in small forest ponds. Arch. Environ. Contam. Toxicol. 1989, 18, 537–544. [Google Scholar] [CrossRef]
- Mallat, E.; Barceló, D. Analysis and degradation study of glyphosate and of aminomethylphosphonic acid in natural waters by means of polymeric and ion-exchange solid-phase extraction columns followed by ion chromatography-post-column derivatization with fluorescence detection. J. Chromatogr. A. 1998, 823, 129–136. [Google Scholar] [CrossRef]
- Pizarro, H.; Vera, M.S.; Vinocur, A.; Perez, G.; Ferraro, M.; Menendez Helman, R.J.; Dos Santos Afonso, M. Glyphosate input modifies microbial community structure in clear and turbid freshwater systems. Environ. Sci. Pollut. Res. 2016, 23, 5143–5153. [Google Scholar] [CrossRef] [PubMed]
- Mercurio, P.; Flores, F.; Mueller, J.F.; Carter, S.; Negri, A.P. Glyphosate persistence in seawater. Mar. Pollut. Bull. 2014, 85, 385–390. [Google Scholar] [CrossRef] [PubMed]
- Kryuchkova, Y.V.; Burygin, G.L.; Gogoleva, N.E.; Gogolev, Y.V.; Chernyshova, M.P.; Makarov, O.E.; Fedorov, E.E.; Turkovskaya, O.V. Isolation and characterization of a glyphosate-degrading rhizosphere strain, Enterobacter cloacae K7. Microbiol. Res. 2014, 169, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Barrett, K.A.; McBride, M.B. Oxidative degradation of glyphosate and aminomethylphosphonate by manganese oxide. Environ. Sci. Technol. 2005, 39, 9223–9228. [Google Scholar] [CrossRef]
- Paudel, P.; Negusse, A.; Jaisi, D.P. Birnessite-catalyzed degradation of glyphosate: A mechanistic study aided by kinetics batch studies and NMR spectroscopy. Soil Sci. Soc. Am. J. 2015, 79, 815–825. [Google Scholar] [CrossRef]
- Pollegioni, L.; Schonbrunn, E.; Siehl, D. Molecular basis of glyphosate resistance-different approaches through protein engineering. FEBS J. 2011, 278, 2753–2766. [Google Scholar] [CrossRef] [Green Version]
- la Cecilia, D.; Maggi, F. Analysis of glyphosate degradation in a soil microcosm. Environ. Pollut. 2018, 233, 201–207. [Google Scholar] [CrossRef]
- Sviridov, A.V.; Shushkova, T.V.; Zelenkova, N.F.; Vinokurova, N.G.; Morgunov, I.G.; Ermakova, I.T.; Leontievsky, A.A. Distribution of glyphosate and methylphosphonate catabolism systems in soil bacteria Ochrobactrum anthropi and Achromobacter sp. Appl. Microbiol. Biotechnol. 2012, 93, 787–796. [Google Scholar] [CrossRef]
- Zhan, H.; Feng, Y.; Fan, X.; Chen, S. Recent advances in glyphosate biodegradation. Appl. Microbiol. Biotechnol. 2018, 102, 5033–5043. [Google Scholar] [CrossRef] [PubMed]
- Sviridov, A.V.; Shushkova, T.V.; Ermakova, I.T.; Ivanova, E.V.; Epiktetov, D.O.; Leont’evskii, A.A. Microbial Degradation of Glyphosate Herbicides (Review). Prikl. Biokhim. Mikrobiol. 2015, 51, 183–190. [Google Scholar]
- Hove-Jensen, B.; Zechel, D.L.; Jochimsen, B. Utilization of glyphosate as phosphate source: Biochemistry and genetics of bacterial carbon-phosphorus lyase. Microbiol. Mol. Biol. R. 2014, 78, 176–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shushkova, T.V.; Vinokurova, N.G.; Baskunov, B.P.; Zelenkova, N.F.; Sviridov, A.V.; Ermakova, I.T.; Leontievsky, A.A. Glyphosate acetylation as a specific trait of Achromobacter sp. Kg 16 physiology. Appl. Microbiol. Biotechnol. 2016, 100, 847–855. [Google Scholar] [CrossRef]
- Pizzul, L.; Castillo, M.D.P.; Stenström, J. Degradation of glyphosate and other pesticides by ligninolytic enzymes. Biodegradation 2009, 20, 751–759. [Google Scholar] [CrossRef] [PubMed]
- Sprankle, P.; Meggitt, W.F.; Penner, D. Adsorption, mobility, and microbial degradation of glyphosate in the soil. Weed Sci. 1975, 23, 229–234. [Google Scholar] [CrossRef]
- Vereecken, H. Mobility and leaching of glyphosate: A review. Pest. Manag. Sci. 2005, 61, 1139–1151. [Google Scholar] [CrossRef]
- Yang, X.; Wang, F.; Bento, C.; Xue, S.; Gai, L.; van Dam, R.; Mol, H.; Ritsema, C.J.; Geissen, V. Short-term transport of glyphosate with erosion in Chinese loess soil–a flume experiment. Sci. Total Environ. 2015, 512–513, 406–414. [Google Scholar] [CrossRef] [PubMed]
- EFSA. Conclusion on the peer review of the pesticide risk assessment of the active substance glyphosate: Peer review of the pesticide risk assessment of the active substance glyphosate. EFSA J. 2015, 13, 4302. [Google Scholar]
- Botta, F.; Lavison, G.; Couturier, G.; Alliot, F.; Moreau-Guigon, E.; Fauchon, N.; Guery, B.; Chevreuil, M.; Blanchoud, H. Transfer of glyphosate and its degradate AMPA to surface waters through urban sewerage systems. Chemosphere 2009, 77, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Peruzzo, P.J.; Porta, A.A.; Ronco, A.E. Levels of glyphosate in surface waters, sediments and soils associated with direct sowing soybean cultivation in north pampasic region of Argentina. Environ. Pollut. 2008, 156, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Mahler, B.J.; Van Metre, P.C.; Burley, T.E.; Loftin, K.A.; Meyer, M.T.; Nowell, L.H. Similarities and differences in occurrence and temporal fluctuations in glyphosate and atrazine in small Midwestern streams (USA) during the 2013 growing season. Sci. Total Environ. 2017, 579, 149–158. [Google Scholar] [CrossRef] [Green Version]
- Okada, E.; Allinson, M.; Barral, M.P.; Clarke, B.; Allinson, G. Glyphosate and aminomethylphosphonic acid (AMPA) are commonly found in urban streams and wetlands of Melbourne, Australia. Water Res. 2020, 168, 115–139. [Google Scholar] [CrossRef] [PubMed]
- Battaglin, W.A.; Meyer, M.T.; Kuivila, K.M.; Dietze, J.E. Glyphosate and its degradation product AMPA occur frequently and widely in U.S.; soils, surface water, groundwater, and precipitation. J. Am. Water Resour. Assoc. 2014, 50, 275–290. [Google Scholar] [CrossRef]
- Okada, E.; Pérez, D.; De Gerónimo, E.; Aparicio, V.; Massone, H.; Costa, J.L. Non-point source pollution of glyphosate and AMPA in a rural basin from the southeast Pampas, Argentina. Environ. Sci. Pollut. Res. 2018, 25, 15120–15132. [Google Scholar] [CrossRef] [PubMed]
- Hanke, I.; Singer, H.; Hollender, J. Ultratrace-level determination of glyphosate, aminomethylphosphonic acid and glufosinate in natural waters by solid-phase extraction followed by liquid chromatography-tandem mass spectrometry: Performance tuning of derivatization, enrichment and detection. Anal. Bioanal. Chem. 2008, 391, 2265–2276. [Google Scholar] [CrossRef] [Green Version]
- Scribner, E.A.; Battaglin, W.A.; Gilliom, R.J.; Meyer, M.T. Concentrations of Glyphosate, Its Degradation Product, Aminomethylphosphonic Acid, and Glufosinate in Ground- and Surface-Water, Rainfall, and Soil Samples Collected in the United States, 2001–2006; U.S. Geological Survey Scientific Investigations Report 2007–5122; U.S. Geological Survey: Reston, VA, USA, 2007; 111p. [Google Scholar]
- Kemp, W.M.; Twilley, R.R.; Stevenson, J.C.; Boynton, W.R.; Means, J.C. The decline of submerged vascular plants in upper Chesapeake Bay: Summary and results concerning possible causes. Mar. Technol. Soc. J. 1983, 2, 78–89. [Google Scholar]
- Burgeot, T.; Gagnaire, B.; Renault, T.; Haure, J.; Moraga, D.; David, E.; Boutet, I.; Sauriau, P.G.; Malet, N.; Bouchet, V.; et al. Oyster summer mortality risks associated with environmental stress. In Summer Mortality of Pacific Oyster Crassostrea Gigas; Samain, J.F., McCombie, H., Eds.; Editions Quæ: Versailles, France, 2007; pp. 107–151. [Google Scholar]
- Wang, S.; Liu, B.; Yuan, D.; Ma, J. A simple method for the determination of glyphosate and aminomethylphosphonic acid in seawater matrix with high performance liquid chromatography and fluorescence detection. Talanta 2016, 161, 700–706. [Google Scholar] [CrossRef]
- Skeff, W.; Neumann, C.; Schulz-Bull, D.E. Glyphosate and AMPA in the estuaries of the Baltic Sea method optimization and field study. Mar. Pollut. Bull. 2015, 100, 577–585. [Google Scholar] [CrossRef]
- Kaiser, K. Preliminary study of pesticide drift into the Maya Mountain protected areas of Belize. Bull. Environ. Contam. Toxicol. 2011, 86, 56–59. [Google Scholar] [CrossRef] [Green Version]
- Chang, F.C.; Simcik, M.F.; Capel, P.D. Occurrence and fate of the herbicide glyphosate and its degradate aminomethylphosphonic acid in the atmosphere. Environ. Toxicol. Chem. 2011, 30, 548–555. [Google Scholar] [CrossRef]
- Solomon, K.R.; Thompson, D.G. Ecological risk assessment for aquatic organisms from over-water uses of glyphosate. J. Toxicol. Env. Heal. B 2003, 6, 289–324. [Google Scholar] [CrossRef]
- Coupe, R.H.; Kalkhoff, S.J.; Capel, P.D.; Gregoire, C. Fate and transport of glyphosate and aminomethylphosphonic acid in surface waters of agricultural basins. Pest. Manag. Sci. 2012, 68, 16–30. [Google Scholar] [CrossRef] [PubMed]
- Avigliano, E.; Schenone, N.F. Human health risk assessment and environmental distribution of trace elements, glyphosate, fecal coliform and total coliform in Atlantic Rainforest mountain rivers (South America). Microchem. J. 2015, 122, 149–158. [Google Scholar] [CrossRef]
- Castro Berman, M.; Marino, D.; Quiroga, M.V.; Zagarese, H. Occurrence and levels of glyphosate and AMPA in shallow lakes from the Pampean and Patagonian regions of Argentina. Chemosphere 2018, 200, 513–522. [Google Scholar] [CrossRef]
- Ruiz-Toledo, J.; Castro, R.; Rivero-Pérez, N.; Bello-Mendoza, R.; Sánchez, D. Occurrence of glyphosate in water bodies derived from intensive agriculture in a tropical region of southern Mexico. Bull. Environ. Contam. Toxicol. 2014, 93, 289–293. [Google Scholar] [CrossRef] [PubMed]
- Masiol, M.; Giannì, B.; Prete, M. Herbicides in river water across the northeastern Italy: Occurrence and spatial patterns of glyphosate, aminomethylphosphonic acid, and glufosinate ammonium. Environ. Sci. Pollut. Res. 2018, 25, 24368–24378. [Google Scholar] [CrossRef] [PubMed]
- Pérez, D.J.; Okada, E.; De Gerónimo, E.; Menone, M.L.; Aparicio, V.C.; Costa, J.L. Spatial and temporal trends and flow dynamics of glyphosate and other pesticides within an agricultural watershed in Argentina. Environ. Toxicol. Chem. 2017, 36, 3206–3216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daouk, S.; Copin, P.J.; Rossi, L.; Chèvre, N.; Pfeifer, H.R. Dynamics and environmental risk assessment of the herbicide glyphosate and its metabolite AMPA in a small vineyard river of the Lake Geneva catchment. Environ. Toxicol. Chem. 2013, 32, 2035–2044. [Google Scholar] [CrossRef]
- Huntscha, S.; Stravs, M.A.; Bühlmann, A.; Ahrens, C.H.; Frey, J.E.; Pomati, F.; Hollender, J.; Buerge, I.J.; Balmer, M.E.; Poiger, T. Seasonal dynamics of glyphosate and AMPA in Lake Greifensee: Rapid microbial degradation in the epilimnion during summer. Environ. Sci. Technol. 2018, 52, 4641–4649. [Google Scholar] [CrossRef]
- Rzymski, P.; Klimaszyk, P.; Kubacki, T.; Poniedziałek, B. The effect of glyphosate-based herbicide on aquatic organisms—A case study. Limnol. Rev. 2013, 4, 215–220. [Google Scholar] [CrossRef]
- John, J.; Liu, H. Glyphosate monitoring in water, foods, and urine reveals an association between urinary glyphosate and tea drinking: A pilot study. Int. J. Env. Health Eng. 2018, 7, 2. [Google Scholar]
- Poiger, T.; Buerge, I.J.; Bächli, A.; Müller, M.D.; Balmer, M.E. Occurrence of the herbicide glyphosate and its metabolite AMPA in surface waters in Switzerland determined with on-line solid phase extraction LC-MS/MS. Environ. Sci. Pollut. Res. 2017, 24, 1588–1596. [Google Scholar] [CrossRef] [PubMed]
- Tzaskos, D.F.; Marcovicz, C.; Dias, N.M.P.; Rosso, N.D. Development of sampling for quantification of glyphosate in natural waters. Cienc. Agrotec. 2012, 36, 399–405. [Google Scholar] [CrossRef] [Green Version]
- Popp, M.; Hann, S.; Mentler, A.; Fuerhacker, M.; Stingeder, G.; Koellensperger, G. Determination of glyphosate and AMPA in surface and waste water using high-performance ion chromatography coupled to inductively coupled plasma dynamic reaction cell mass spectrometry (HPIC–ICP–DRC–MS). Anal. Bioanal. Chem. 2008, 391, 695–699. [Google Scholar] [CrossRef] [PubMed]
- Montiel-León, J.M.; Munoz, G.; Vo Duy, S.; Do, D.T.; Vaudreuil, M.A.; Goeury, K.; Guillemette, F.; Amyot, M.; Sauvé, S. Widespread occurrence and spatial distribution of glyphosate, atrazine, and neonicotinoids pesticides in the St. Lawrence and tributary rivers. Environ. Pollut. 2019, 250, 29–39. [Google Scholar]
- Zhang, Q.; Zhou, H.; Li, Z.; Zhu, J.; Zhou, C.; Zhao, M. Effects of glyphosate at environmentally relevant concentrations on the growth of and microcystin production by Microcystis aeruginosa. Environ. Monit. Assess. 2016, 188, 632. [Google Scholar] [CrossRef]
- Aparicio, V.C.; De Gerónimo, E.; Marino, D.; Primost, J.; Carriquiriborde, P.; Costa, J.L. Environmental fate of glyphosate and aminomethylphosphonic acid in surface waters and soil of agricultural basins. Chemosphere 2013, 93, 1866–1873. [Google Scholar] [CrossRef]
- Giroux, I. Présence de pesticides dans l’eau au Québec: Portrait et tendances dans les zones de maïs et de soya - 2011 à 2014, Québec, ministère du Développement durable, de l’Environnement et de la Lutte contre les changements climatiques, Direction du suivi de l’état de l’environnement. ISBN 978-2-550-73603-5. Available online: http://www.mddelcc.gouv.qc.ca/eau/flrivlac/pesticides.htm (accessed on 24 May 2020).
- Ronco, A.E.; Marino, D.J.; Abelando, M.; Almada, P.; Apartin, C.D. Water quality of the main tributaries of the Paraná Basin: Glyphosate and AMPA in surface water and bottom sediments. Environ. Monit. Assess. 2016, 188, 458. [Google Scholar] [CrossRef]
- Byer, J.D.; Struger, J.; Klawunn, P.; Todd, A.; Sverko, E. Low cost monitoring of glyphosate in surface waters using the ELISA method: An evaluation. Environ. Sci. Technol. 2008, 42, 6052–6057. [Google Scholar] [CrossRef]
- ISPRA. Rapporto Nazionale Pesticidi Nelle Acque, Dati 2015–2016. 2018. Available online: http://www.confagricolturave.it/ambiente-archivio/694-rapporto-nazionale-pesticidi-nelle-acque-dati-2015-e-2016 (accessed on 31 May 2020).
- Glozier, N.E.; Struger, J.; Cessna, A.J.; Gledhill, M.; Rondeau, M.; Ernst, W.R.; Sekela, M.A.; Cagampan, S.J.; Sverko, E.; Murphy, C.; et al. Occurrence of glyphosate and acidic herbicides in select urban rivers and streams in Canada, 2007. Environ. Sci. Pollut. Res. 2012, 19, 821–834. [Google Scholar] [CrossRef]
- Freire, R.; Schneider, R.M.; de Freitas, F.H.; Bonifácio, C.M.; Tavares, C.R.G. Monitoring of toxic chemical in the basin of Maringá stream. Acta Sci. Technol. 2012, 34, 295–302. [Google Scholar] [CrossRef] [Green Version]
- Medalie, L.; Baker, N.T.; Shoda, M.E.; Stone, W.W.; Meyer, M.T.; Stets, E.G.; Wilson, M. Influence of land use and region on glyphosate and aminomethylphosphonic acid in streams in the USA. Sci. Total Environ. 2020, 707, 136008. [Google Scholar] [CrossRef] [PubMed]
- Mörtl, M.; Németh, G.; Juracsek, J.; Darvas, B.; Kamp, L.; Rubio, F.; Székács, A. Determination of glyphosate residues in Hungarian water samples by immunoassay. Microchem. J. 2013, 107, 143–151. [Google Scholar] [CrossRef]
- Struger, J.; Thompson, D.; Staznik, B.; Martin, P.; McDaniel, T.; Marvin, C. Occurrence of glyphosate in surface waters of Southern Ontario. Bull. Environ. Contam. Toxicol. 2008, 80, 378–384. [Google Scholar] [CrossRef] [PubMed]
- Bonansea, R.I.; Filippi, I.; Wunderlin, D.A.; Marino, D.; Amé, M.V. The fate of glyphosate and AMPA in a freshwater endorheic basin: An ecotoxicological risk assessment. Toxics 2017, 6, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tierney, K.B.; Sekela, M.A.; Cobbler, C.E.; Xhabija, B.; Gledhill, M.; Ananvoranich, S.; Zielinski, B.S. Evidence for behavioral preference toward environmental concentrations of urban-use herbicides in a model adult fish. Environ. Toxicol. Chem. 2011, 30, 2046–2054. [Google Scholar] [CrossRef] [PubMed]
- Kolpin, D.W.; Thurman, E.M.; Lee, E.A.; Meyer, M.T.; Furlong, E.T.; Glassmeyer, S.T. Urban contributions of glyphosate and its degradate AMPA to streams in the United States. Sci. Total Environ. 2006, 354, 191–197. [Google Scholar] [CrossRef]
- Battaglin, W.A.; Kolpin, D.W.; Scribner, E.A.; Kuivila, K.M.; Sandstrom, M.W. Glyphosate, other herbicides, and transformation products in midwestern streams, 2002. J. Am. Water. Resour. As. 2005, 41, 323–332. [Google Scholar] [CrossRef] [Green Version]
- Brüsch, W.; Rosenbom, A.E.; Badawi, N.; Gudmondsson, L.; von Platten-Hallermund, F.; Nielsen, C.B.; Plauborg, F.; Laier, T.; Olsen, P. The Danish Pesticide Leaching Assessment Programme; GEUS: København, Denmark, 2015; ISBN 978-87-7871-388-9. [Google Scholar]
- Ramirez, C.E.; Bellmund, S.; Gardinali, P.R. A simple method for routine monitoring of glyphosate and its main metabolite in surface waters using lyophilization and LC-FLD+MS/MS. Case study: Canals with influence on Biscayne National Park. Sci. Total Environ. 2014, 496, 389–401. [Google Scholar] [CrossRef]
- Trégouët, B.; Nirascou, F. Les pesticides dans les eaux, données 2003 et 2004. Les dossiers ifen 2006, 5, 1–40. [Google Scholar]
- Horth, H.; Blackmore, K. Survey of Glyphosate and AMPA in Groundwaters and Surface Waters in Europe; WRC Report No. UC8073.02; WRc plc: Swindon, UK, 2009. [Google Scholar]
- Sanchís, J.; Kantiani, L.; Llorca, M.; Rubio, F.; Ginebreda, A.; Fraile, J.; Garrido, T.; Farré, M. Determination of glyphosate in groundwater samples using an ultrasensitive immunoassay and confirmation by on-line solid-phase extraction followed by liquid chromatography coupled to tandem mass spectrometry. Anal. Bioanal. Chem. 2012, 402, 2335–2345. [Google Scholar] [CrossRef]
- Van Stempvoort, D.R.; Roy, J.W.; Brown, S.J.; Bickerton, G. Residues of the herbicide glyphosate in riparian groundwater in urban catchments. Chemosphere 2014, 95, 455–463. [Google Scholar] [CrossRef] [PubMed]
- Rendon-von Osten, J.; Dzul-Caamal, R. Glyphosate Residues in Groundwater, Drinking Water and Urine of Subsistence Farmers from Intensive Agriculture Localities: A Survey in Hopelchén, Campeche, Mexico. Int. J. Environ. Res. Public Health 2017, 14, 595. [Google Scholar] [CrossRef] [PubMed]
- Van Stempvoort, D.R.; Spoelstra, J.; Senger, N.D.; Brown, S.J.; Post, R.; Struger, J. Glyphosate residues in rural groundwater, Nottawasaga River Watershed, Ontario, Canada. Pest. Manag. Sci. 2016, 72, 1862–1872. [Google Scholar] [CrossRef] [PubMed]
- Akcha, F.; Spagnol, C.; Rouxel, J. Genotoxicity of diuron and glyphosate in oyster spermatozoa and embryos. Aquat. Toxicol. 2012, 106–107, 104–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mottier, A.; Séguin, A.; Devos, A.; Le Pabic, C.; Voiseux, C.; Lebel, J.M.; Serpentini, A.; Fievet, B.; Costil, K. Effects of subchronic exposure to glyphosate in juvenile oysters (Crassostrea gigas): From molecular to individual levels. Mar. Pollut. Bull. 2015, 95, 665–677. [Google Scholar] [CrossRef] [PubMed]
- Favret, K.P.; Lynn, J.W. Flow-cytometric analyses of viability biomarkers in pesticide-exposed sperm of three aquatic invertebrates. Arch. Environ. Contam. Toxicol. 2010, 58, 973–984. [Google Scholar] [CrossRef]
- Boutet, I.; Tanguy, A.; Moraga, D. Characterisation and expression of four mRNA sequences encoding glutathione S-transferases pi, mu, omega and sigma classes in the Pacific oyster Crassostrea gigas exposed to hydrocarbons and pesticides. Mar. Biol. 2004, 146, 53–64. [Google Scholar] [CrossRef] [Green Version]
- Tanguy, A.; Boutet, I.; Laroche, J.; Moraga, D. Molecular identification and expression study of differentially regulated genes in the Pacific oyster Crassostrea gigas in response to pesticide exposure. FEBS J. 2005, 272, 390–403. [Google Scholar] [CrossRef]
- Milan, M.; Dalla Rovere, G.; Smits, M.; Ferraresso, S.; Pastore, P.; Marin, M.G.; Bogialli, S.; Patarnello, T.; Bargelloni, L.; Matozzo, V. Ecotoxicological effects of the herbicide glyphosate in non-target aquatic species: Transcriptional responses in the mussel Mytilus galloprovincialis. Environ. Pollut. 2018, 237, 442–451. [Google Scholar] [CrossRef]
- Matozzo, V.; Fabrello, J.; Masiero, L.; Ferraccioli, F.; Finos, L.; Pastore, P.; Di Gangi, I.M.; Bogialli, S. Ecotoxicological risk assessment for the herbicide glyphosate to non-target aquatic species: A case study with the mussel Mytilus galloprovincialis. Environ. Pollut. 2018, 233, 623–632. [Google Scholar] [CrossRef]
- Matozzo, V.; Zampieri, C.; Munari, M.; Marin, M.G. Glyphosate affects haemocyte parameters in the clam Ruditapes philippinarum. Mar. Environ. Res. 2019, 146, 66–70. [Google Scholar] [CrossRef] [PubMed]
- Sandrini, J.Z.; Rola, R.C.; Lopes, F.M.; Buffon, H.F.; Freitas, M.M.; Martins, C.d.M.G.; da Rosa, C.E. Effects of glyphosate on cholinesterase activity of the mussel Perna perna and the fish Danio rerio and Jenynsia multidentata: In vitro studies. Aquat. Toxicol. 2013, 130–131, 171–173. [Google Scholar] [CrossRef] [PubMed]
- Séguin, A.; Mottier, A.; Perron, C.; Lebel, J.M.; Serpentini, A.; Costil, K. Sub-lethal effects of a glyphosate-based commercial formulation and adjuvants on juvenile oysters (Crassostrea gigas) exposed for 35 days. Mar. Pollut. Bull. 2017, 117, 348–358. [Google Scholar] [CrossRef] [PubMed]
- Mottier, A.; Serpentini, A.; Dallas, L.; James, A.; Lebel, J.M.; Costil, K. In vitro effects of glyphosate-based herbicides and related adjuvants on primary culture of hemocytes from Haliotis tuberculata. Fish Shellfish Immunol. 2020, 100, 1–8. [Google Scholar] [CrossRef]
- Hanana, H.; Simon, G.; Kervarec, N.; Mohammadou, B.A.; Cérantola, S. HRMAS NMR as a tool to study metabolic responses in heart clam Ruditapes decussatus exposed to Roundup®. Talanta 2012, 97, 425–431. [Google Scholar] [CrossRef]
- Fabrello, J.; Grapputo, A.; Munari, M.; Marin, M.G.; Masiero, L.; Pacchioni, B.; Millino, C.; Matozzo, V. Molecular and biochemical responses of vitellogenin in the mussel Mytilus galloprovincialis exposed to the glyphosate-based herbicide Roundup® Power 2.0. Environ. Sci. Pollut. Res. 2020, in press. [Google Scholar] [CrossRef]
- de Brito Rodrigues, L.; de Oliveira, R.; Abe, F.R.; Brito, L.B.; Moura, D.S.; Valadares, M.C.; Grisolia, C.K.; de Oliveira, D.P.; de Oliveira, G.A.R. Ecotoxicological assessment of glyphosate-based herbicides: Effects on different organisms. Environ. Toxicol. Chem. 2017, 36, 1755–1763. [Google Scholar] [CrossRef]
- Tsui, M.T.; Chu, L.M. Aquatic toxicity of glyphosate-based formulations: Comparison between different organisms and the effects of environmental factors. Chemosphere 2003, 52, 1189–1197. [Google Scholar] [CrossRef]
- Osterberg, J.S.; Darnell, K.M.; Blickley, T.M.; Romano, J.A.; Rittschof, D. Acute toxicity and sub-lethal effects of common pesticides in post-larval and juvenile blue crabs, Callinectes sapidus. J. Exp. Mar. Bio. Ecol. 2012, 424–425, 5–14. [Google Scholar] [CrossRef]
- Avigliano, L.; Canosa, I.S.; Medesani, D.A.; Rodríguez, E. Effects of glyphosate on somatic and ovarian growth in the estuarine crab Neohelice granulata, during the pre-reproductive period. Water Air Soil Pollut. 2018, 229, 44. [Google Scholar] [CrossRef]
- Canosa, I.S.; Silveyra, G.R.; Avigliano, L.; Medesani, D.A.; Rodríguez, E.M. Ovarian growth impairment after chronic exposure to Roundup Ultramax® in the estuarine crab Neohelice granulata. Environ. Sci. Pollut. Res. 2018, 25, 1568–1575. [Google Scholar] [CrossRef] [PubMed]
- Canosa, I.S.; Zanitti, M.; Lonné, N.; Medesani, D.A.; López Greco, L.S.; Rodríguez, E.M. Imbalances in the male reproductive function of the estuarine crab Neohelice granulata, caused by glyphosate. Ecotoxicol. Environ. Saf. 2019, 182, 109405. [Google Scholar] [CrossRef] [PubMed]
- His, E.; Heyvang, I.; Geffard, O.; De Montaudouin, X. A comparison between oyster (Crassostrea gigas) and sea urchin (Paracentrotus lividus) larval bioassays for toxicological studies. Water Res. 1999, 33, 1706–1718. [Google Scholar] [CrossRef]
- Amid, C.; Olstedt, M.; Gunnarsson, J.S.; Le Lan, H.; Tran Thi Minh, H.; Van den Brink, P.J.; Hellström, M.; Tedengren, M. Additive effects of the herbicide glyphosate and elevated temperature on the branched coral Acropora formosa in Nha Trang, Vietnam. Environ. Sci. Pollut. Res. 2018, 25, 13360–13372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Melo Tarouco, F.; de Godoi, F.G.A.; Velasques, R.R.; da Silveira Guerreiro, A.; Geihs, M.A.; da Rosa, C.E. Effects of the herbicide Roundup on the polychaeta Laeonereis acuta: Cholinesterases and oxidative stress. Ecotoxicol. Environ. Saf. 2017, 135, 259–266. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Main degradation pathways of glyphosate. Oxygen, water molecules and hydrogen atoms were omitted to facilitate the visualization.
Figure 1.
Main degradation pathways of glyphosate. Oxygen, water molecules and hydrogen atoms were omitted to facilitate the visualization.


{kind=link}
Table 1.
Chemical and physical properties of glyphosate.
| IUPAC Name | N-(phosphonomethyl) Glycine |
|---|---|
| CAS Number | 1071-83-6 |
| Molecular Formula | C3H8NO5P |
| Chemical Structure |  |
| Molecular Weight | 169.07 g/mol |
| Physical state and Color | White crystalline powder |
| Melting Point | 189.5–230 °C |
| Density at 20 °C | 1.705 |
| Water Solubility at 25 °C | 1.2 g/100 mL |
| Octanol-water Coeff. (log Kow) | −3.40 to −1.0 |
| Vapor Pressure at 25 °C | 1.31 × 10-2 mPa |
Table 2.
Occurrence of glyphosate in water ecosystems. Data are reported as <LOD (limit of detection) up to the highest measured concentration.
Table 2.
Occurrence of glyphosate in water ecosystems. Data are reported as <LOD (limit of detection) up to the highest measured concentration.
| Glyphosate Concentrations | Water Environment, Country | Ref. |
|---|---|---|
| 0.10 to 0.70 mg/L | Surface waters, Argentina | [66] |
| <LOD-427 µg/L | Surface waters, USA | [69] |
| <LOD-430 μg/L | Surface waters, USA | [80] |
| <LOD-1600 μg/L | Surface waters, Argentina | [81] |
| <LOD-4.52 μg/L (2.11 * μg/L) | Surface waters, Argentina | [82] |
| <LOD-36.71 μg/L (3.02 * μg/L) | Surface waters, Mexico | [83] |
| <LOD-27.8 μg/L (1.68 ** μg/L) | Surface waters, USA | [67] |
| <0.1–427 μg/L | Surface waters, USA | [72] |
| <LOD-2.1 μg/L | Surface waters, Italy | [84] |
| <LOD-8.2 μg/L (0.4 * μg/L) | Surface waters, Argentina | [70] |
| LOD-2.9 μg/L (0.78 * μg/L) | Surface waters, Argentina | [85] |
| <LOD-4970 ng/L | Surface waters, Switzerland | [86] |
| <LOD-145 ng/L | Surface waters, Switzerland | [87] |
| 15–390 ng/L | Surface waters, Switzerland | [71] |
| <LOD-90 μg/L | Surface waters, Poland | [88] |
| <LOD-0.08 μg/L | Surface waters, USA | [89] |
| <LOD-2.1 μg/L (0.11 ** μg/L) | Surface waters, Switzerland | [90] |
| 1.258–1.550 mg/L | Surface waters, Brazil | [91] |
| < LOD-1.93 μg/L | Surface waters, Austria | [92] |
| <2–3000 ng/L (109 * ng/L; 26.9 ** ng/L) | Surface waters, Canada | [93] |
| 10 mg/L | Surface waters, China | [94] |
| <LOD-7.6 μg/L | Surface water, Argentina | [95] |
| <LOD-18 μg/L | Surface waters, Canada | [96] |
| <LOD-0.7 μg/L (0.6 * μg/L) | Surface water, Argentina | [97] |
| <LOD-12 μg/L | Surface waters, Canada | [98] |
| <LOD-0.08 μg/L | Surface waters, Italy | [99] |
| <LOD-11.8 μg/L (158.6 * ng/L; 19.8 ** ng/L) | Surface waters, Canada | [100] |
| <LOD-0.041 mg/L | Surface waters, Brazil | [101] |
| <LOD-8.1 μg/L (0.05 ** μg/L) | Surface waters, USA | [102] |
| <LOD-0.68 ng/mL | Surface water, Hungary | [103] |
| <LOD-40.8 μg/L | Surface water, Canada | [104] |
| <LOD-125 μg/L | Surface waters, Argentina | [105] |
| <LOD-455 ng/L | Surface water, Canada | [106] |
| <LOD-14.2 μg/L | Surface waters, Australia | [68] |
| <LOD-2.2 μg/L | Surface waters, USA | [107] |
| <LOD-8.7 μg/L | Surface waters, USA | [108] |
| <LOD-90 μg/L | Surface waters, France | [65] |
| <LOD-31 μg/L | Surface waters, Denmark | [109] |
| <LOD-59.9 μg/L | Surface waters, USA | [110] |
| <LOD-165 μg/L | Surface waters, France | [111] |
| <LOD-1.3 μg/L | Surface waters, Belgium | [112] |
| <LOD-0.46 μg/L | Surface waters, Finland | [112] |
| <LOD-50 μg/L | Surface waters, France | [112] |
| <LOD-4.7 μg/L | Surface waters, Germany | [112] |
| <LOD-1.8 μg/L | Surface waters, Ireland | [112] |
| <LOD-11 μg/L | Surface waters, Italy | [112] |
| <LOD-0.93 μg/L | Surface waters, Norway | [112] |
| <LOD-3.6 μg/L | Surface waters, Slovakia | [112] |
| <LOD-15.3 μg/L | Surface waters, Spain | [112] |
| <LOD-13 μg/L | Surface waters, Sweden | [112] |
| <LOD-8.8 μg/L | Surface waters, UK | [112] |
| <LOD-3.6 μg/L | Surface waters, Austria | [112] |
| <LOD-8.7 μg/L | Groundwaters, Denmark | [112] |
| <LOD-24 μg/L | Groundwaters, France | [112] |
| <LOD-1.2 μg/L | Groundwaters, Italy | [112] |
| <LOD-1.7 μg/L | Groundwaters, Sweden | [112] |
| <LOD-0.21 μg/L | Groundwaters, Switzerland | [112] |
| <LOD-4.7 μg/L | Groundwaters, Netherlands | [112] |
| <LOD-0.47 μg/L | Groundwaters, UK | [112] |
| <LOD-6.8 μg/L | Groundwaters, France | [111] |
| <LOD-0.67 μg/L | Groundwaters, Denmark | [109] |
| <LOD-0.98 ng/mL | Groundwaters, Hungary | [103] |
| <LOD-0.011 μg/L | Groundwaters, Italy | [99] |
| <LOD-2.56 μg/L (202 * ng/L) | Groundwaters, Spain | [113] |
| <LOD-0.025 μg/L | Groundwaters, Switzerland | [90] |
| <LOD-42 ng/L | Groundwater, Canada | [114] |
| <LOD-8.5 μg/L (0.4 * μg/L) | Groundwaters, Argentina | [70] |
| 0.44–1.41 μg/L | Groundwaters, Mexico | [115] |
| <0.1–4.7 μg/L | Groundwaters, USA | [72] |
| <LOD-663 ng/L | Groundwaters, Canada | [116] |
| <LOD-1690 ng/L (up to 665 * ng/L) | Sea water, Baltic Sea estuaries | [76] |
| 13–1377 μg/L | Sea waters, Western Pacific | [75] |
| <LOD-1.2 μg/L | Sea waters, French Atlantic coast | [74] |
| <0.1–2.5 μg/L (0.1–0.2 ** μg/L) | Precipitations, USA | [78] |
| 0.3–1.1 μg/L | Precipitations, USA | [72] |
| <LOD-135 ng/L | Precipitation, Canada Ontario | [116] |
| 0.2210–5 μg/L | Phytotelmic water, Belize | [77] |
When available, mean * and median ** concentrations are reported in parentheses.
Table 3.
Effects of glyphosate (as active ingredient) and its commercial formulations in mollusks. Abbreviations: AChE (acetylcholinesterase); ALP (alkali labile phosphate); GST (glutathione S-transferases); HL (hemocyte lysate); REX: Roundup Express®; THC (total hemocyte count); Vtg (vitellogenin).
Table 3.
Effects of glyphosate (as active ingredient) and its commercial formulations in mollusks. Abbreviations: AChE (acetylcholinesterase); ALP (alkali labile phosphate); GST (glutathione S-transferases); HL (hemocyte lysate); REX: Roundup Express®; THC (total hemocyte count); Vtg (vitellogenin).
| Compound Tested | Concentrations (Exposure Type) | Species | Effects | Ref. |
|---|---|---|---|---|
| Glyphosate (active ingredient) Roundup (commercial formulation) | 0.5, 1, 1.5, 2.5, 5 µg/L (in vitro exposure) | Crassostrea gigas (gametes and embryos) | increases in the percentage of abnormal D-larvae no genotoxic effects on oyster spermatozoa | [117] |
| Glyphosate (acid, 97% purity) | 0.1, 1, 100 µg/L (in vivo exposure) | Crassostrea gigas (juveniles) | no mortality no growth no histological alterations moderate alterations of enzyme activities moderate alterations in gene expression | [118] |
| Roundup Ready-To-Use Plus® | 0.25, 1, 4, 16 mg/L (in vitro exposure) | Crassostrea virginica (sperm) | no significant alterations in mitochondrial membrane potential in the sperm | [119] |
| Glyphosate (active ingredient) | 2 µg/L (in vivo exposure) | Crassostrea gigas (adults) | gills: decreases in expression of GSTs digestive gland: increases in expression of GSTs | [120] |
| Glyphosate (active ingredient) | 2 µg/L (in vivo exposure) | Crassostrea gigas (adults) | differentially regulated gene expression in gills and digestive gland | [121] |
| Glyphosate (active ingredient) | 10, 100, 1000 µg/L (in vivo exposure) | Mytilus galloprovincialis (adults) | effects on the transcriptional regulation of genes involved in important cell functions | [122] |
| Glyphosate (active ingredient) | 10, 100, 1000 µg/L (in vivo exposure) | Mytilus galloprovincialis (adults) | alterations in hemocyte parameters no marked alteration in antioxidant enzyme activity in gills and digestive gland alterations in gill AChE activity | [123] |
| Glyphosate (active ingredient) | 10, 100, 1000 µg/L (in vivo exposure) | Ruditapes philippinarum (adults) | reductions in THC increases in hemocyte volume and diameter increases in hemocyte proliferation increase in HL acid phosphatase activity | [124] |
| Glyphosate (pure) | from 0.075 to 15 mM (in vitro exposure) | Perna perna (tissue from juveniles) | significant inhibition of cholinesterase activity in gills and muscle | [125] |
| Roundup Express® (REX) | 0.1, 1, 100 μg/L (in vivo exposure) | Crassostrea gigas (juveniles) | no mortality no effects on condition index delay in gametogenesis decreases in shell length slight reduction in whole weight moderate alterations of digestive gland enzyme activities | [126] |
| Glyphosate (active ingredient) REX (commercial formulation) | from 0.1 to 100,000 μg/L (in vitro exposure) | Haliotis tuberculata (hemocytes) | cell viability: significant reduction due to REX, but not to glyphosate phagocytosis: significant reduction due to REX, but not to glyphosate lysosomal stability: significant effects of low concentrations of glyphosate and high levels of REX | [127] |
| Roundup® 3plus | 0.2, 1 g/L (in vivo exposure) | Ruditapes decussatus (adults) | alteration of energy metabolism and metabolic biomarkers | [128] |
| Roundup® Power 2.0 | 100, 1000 μg/L (in vivo exposure) | Mytilus galloprovincialis (adults) | no significant alterations in vtg gene expression in female gonads reduction in ALP levels if female gonads at 21 days reduction in ALP levels if male gonads at 7 days and increases at 21 days | [129] |
Table 4.
Effects of glyphosate (as active ingredient) and its commercial formulations in other marine invertebrates. Abbreviations: AChE (acetylcholinesterase); LC50 (median lethal concentration); ROS (reactive oxygen species); SOD (superoxide dismutase); Vtg (vitellogenin).
Table 4.
Effects of glyphosate (as active ingredient) and its commercial formulations in other marine invertebrates. Abbreviations: AChE (acetylcholinesterase); LC50 (median lethal concentration); ROS (reactive oxygen species); SOD (superoxide dismutase); Vtg (vitellogenin).
| Compound Tested | Concentrations (Exposure Type) | Species Crustacea | Effects | Ref. |
|---|---|---|---|---|
| Roundup® Glyphosate AKB 480 | 5, 10, 25, 50, 100 mg/L, corresponding to 1.8, 3.6, 9, 18 and 36 mg/L of glyphosate (in vivo exposure) | Artemia salina (nauplii) | increased mortality rates, even if Roundup was more toxic than AKB | [130] |
| Glyphosate acid Isopropylamine salt of glyphosate Roundup® | 1 to 100 mg acid equivalent/L (in vivo exposure) | Acartia tonsa (adults) | LC50 determination | [131] |
| Roundup® Pro | 103 to 107 μg/L (in vivo exposure) | Callinectes sapidus (megalopae and juvenile stages) | LC50 determination | [132] |
| Glyphosate (active ingredient) | 0.02, 0.2, 1 mg/L (in vivo exposure) | Neohelice granulata (adult females) | decrease in body weight no effects on gonadosomatic index no effects on vitellogenic protein content of the ovary increased percentage of reabsorbed vitellogenic oocytes | [133] |
| Roundup Ultramax® | 0.01 and 0.2 mg/L asacid equivalent (in vivo exposure) | Neohelice granulata (adult females) | increases in glycemia no effects on muscle glycogen content no effects on gonadosomatic index increased percentage of reabsorbed vitellogenic oocytes decrease in Vtg levels in the ovary | [134] |
| Glyphosate (as-active ingredient) Roundup Ultramax® | 1 mg/L (in vivo exposure) | Neohelice granulata (adult males) | reductions in weight reductions in total protein levels in muscle increase in glycogen content in muscle increased percentage of abnormal spermatophores | [135] |
| Echinoderms | ||||
| Roundup Ready-To-Use Plus® | 0.25, 1, 4, 16 mg/L (in vitro exposure) | Lytechinus variegatus (sperm) | no effects on mitochondrial membrane potential of sperm | [119] |
| Glyphosate | 10, 25, 50, 100, 150, 200 μg/L (in vivo exposure) | Paracentrotus lividus (larvae) | no abnormal development of larvae | [136] |
| Corals | ||||
| Glyphosate + Increased temperature | 0.12, 1.2, 6, 12 mg/L (in vivo exposure) | Acropora formosa (adults) | significant effects of temperature*glyphosate interaction on coral pigmentation and chlorophyll a | [137] |
| Polychaetes | ||||
| Roundup | from 0.065 to 65 mg/L (in vivo exposure) | Laeonereis acuta (adults) | LC50 determination no significant change in oxygen consumption significant decrease in AChE activity significant decrease in ROS production after 24 h significant decrease in antioxidant capacity against peroxyl radicals after 24 h no significant alterations in antioxidant enzyme activities, except for SOD significant alterations in lipid peroxidation | [138] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Matozzo, V.; Fabrello, J.; Marin, M.G. The Effects of Glyphosate and Its Commercial Formulations to Marine Invertebrates: A Review. J. Mar. Sci. Eng. 2020, 8, 399. https://doi.org/10.3390/jmse8060399
AMA Style
Matozzo V, Fabrello J, Marin MG. The Effects of Glyphosate and Its Commercial Formulations to Marine Invertebrates: A Review. Journal of Marine Science and Engineering. 2020; 8(6):399. https://doi.org/10.3390/jmse8060399
Chicago/Turabian StyleMatozzo, Valerio, Jacopo Fabrello, and Maria Gabriella Marin. 2020. "The Effects of Glyphosate and Its Commercial Formulations to Marine Invertebrates: A Review" Journal of Marine Science and Engineering 8, no. 6: 399. https://doi.org/10.3390/jmse8060399
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.