Biomonitoring of the Application of Monoculture and Integrated Multi-Trophic Aquaculture (IMTA) Using Macrobenthic Structures at Tembelas Island, Kepulauan Riau Province, Indonesia


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Sediment Collection
2.3. Indices and Multivariate Analysis
3. Results
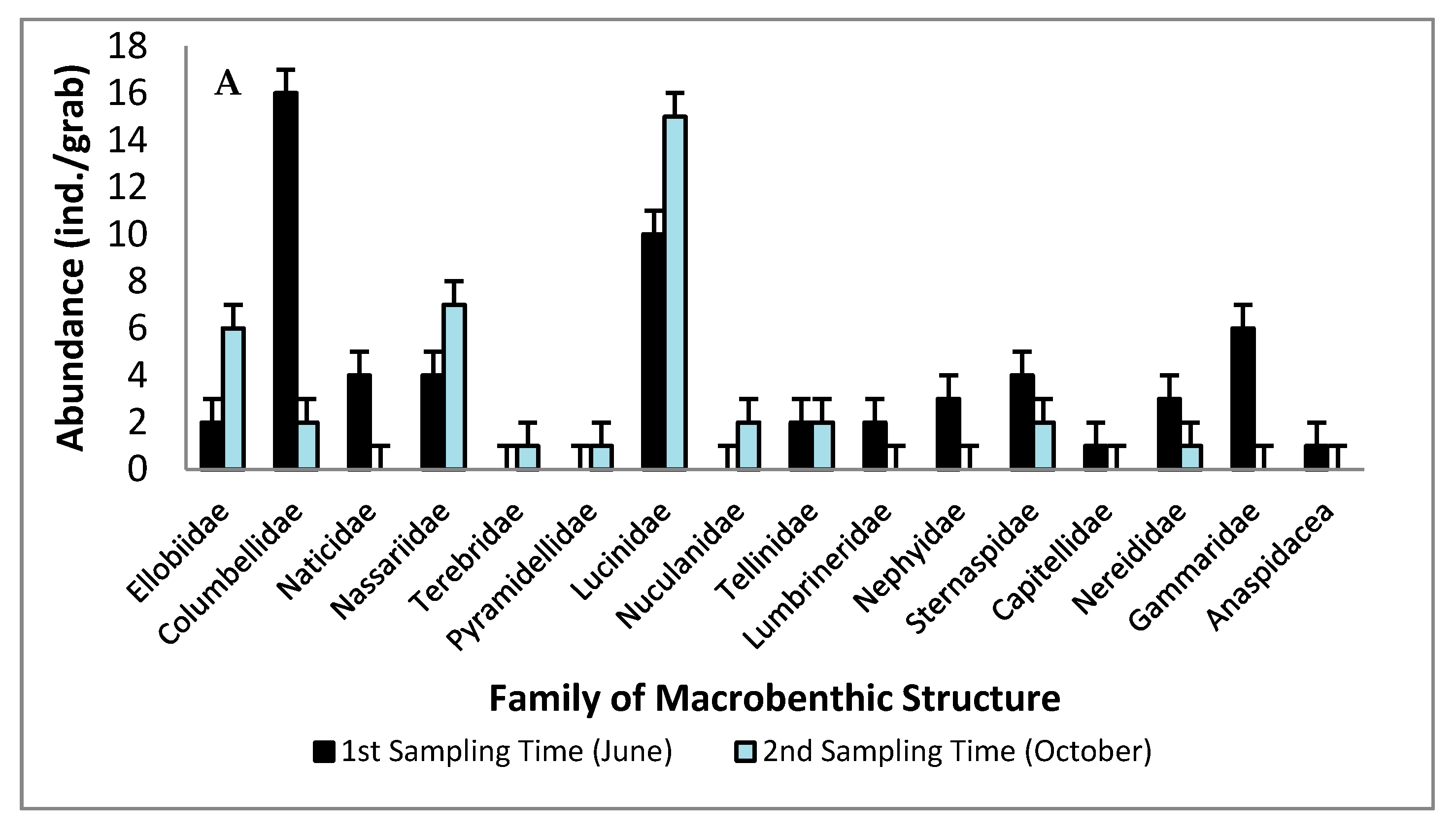
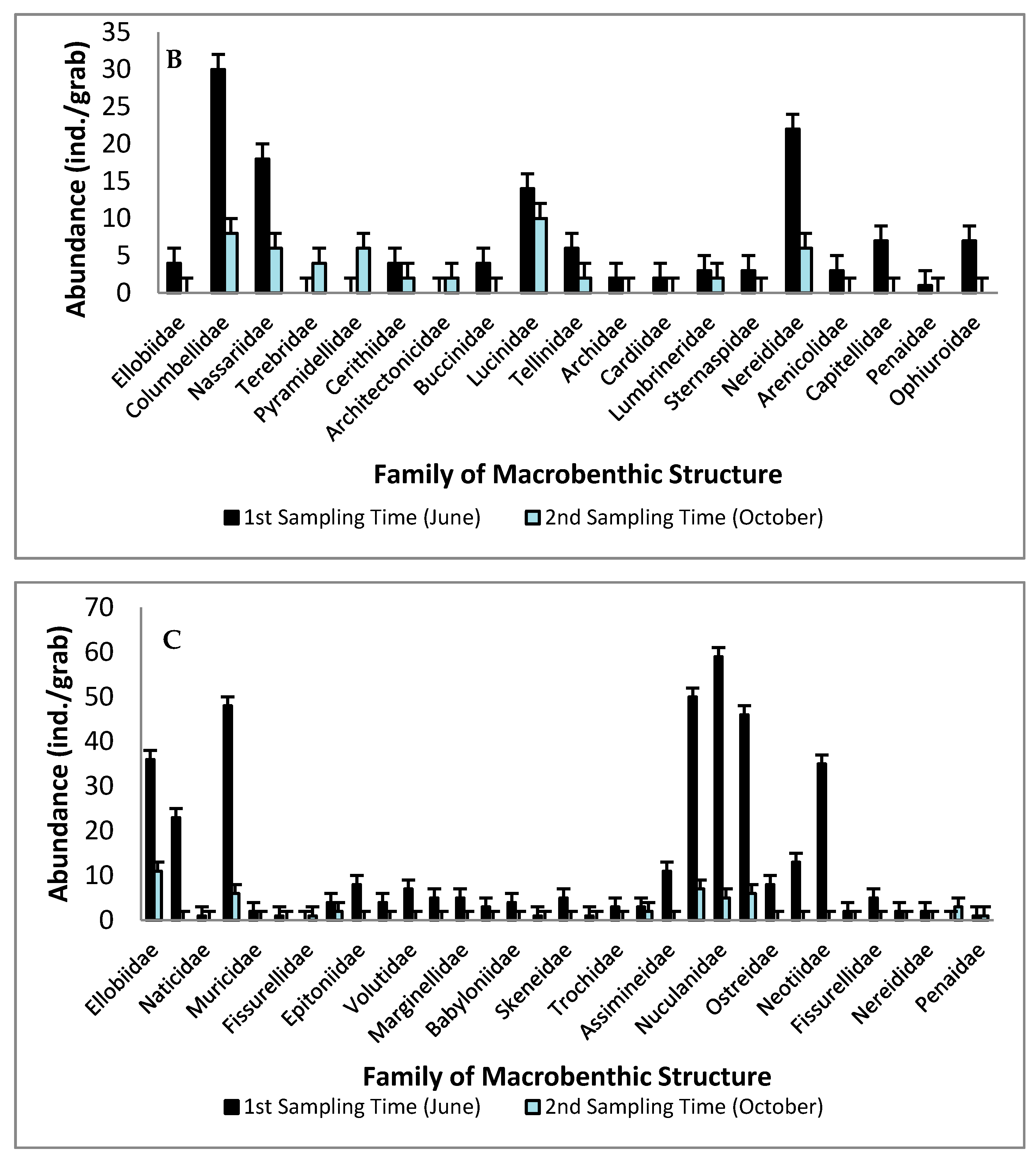
3.1. Macrobenthic Structure: Spatial and Temporal
3.2. The Dominant Taxa of Macrobenthic Assemblages
3.3. The Macrobenthic Assemblages Expressed in Indices
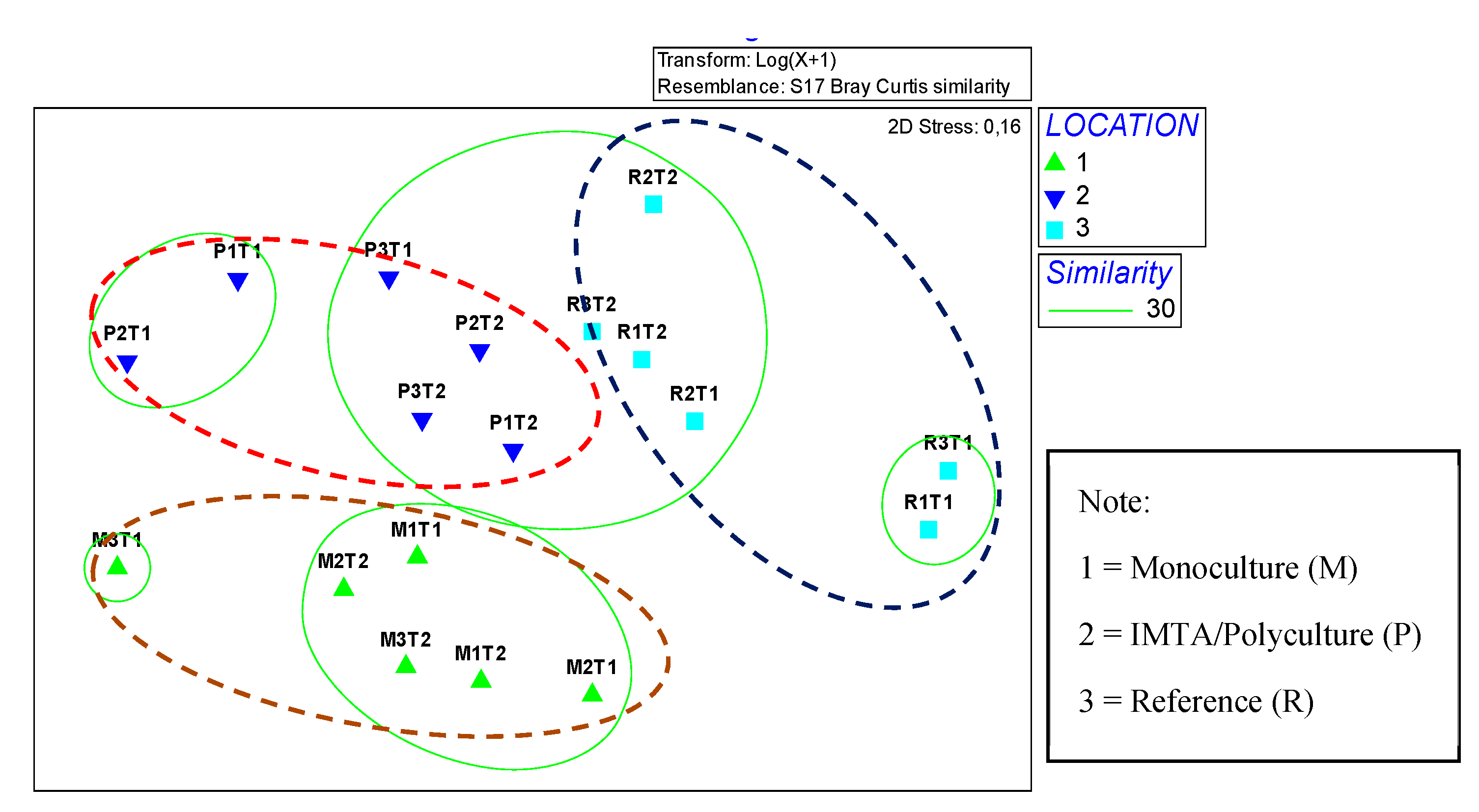
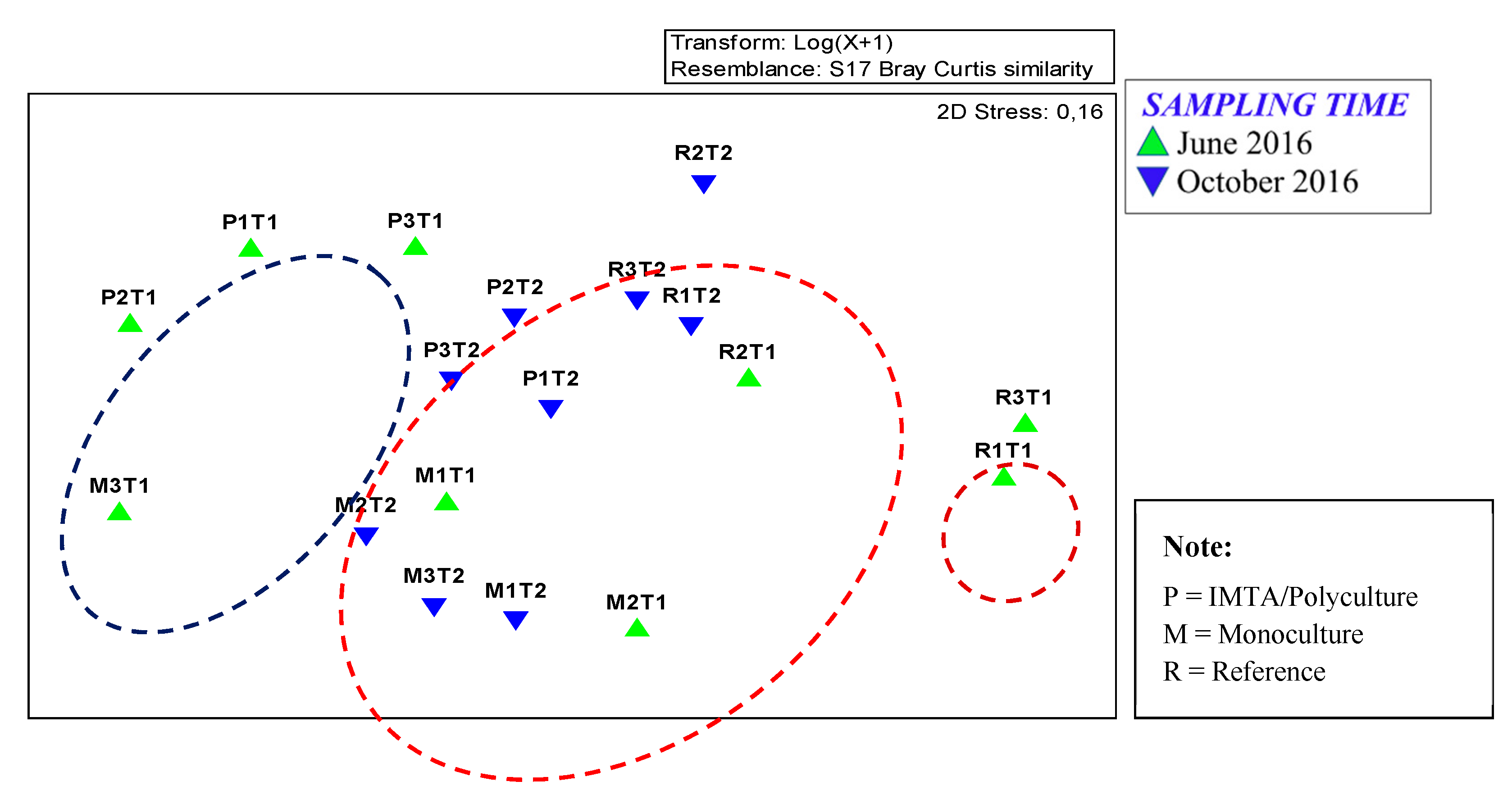
3.4. Macrobenthic Assemblages Expressed with the NMDS Graphical Method
4. Discussion
4.1. Expressing the Macrobenthic Assemblages with Indices
4.2. Abundance as an Indicator for Ecosystem Health
4.3. Examining the Macrobenthic Assemblages through the NMDS Graphical Method
4.4. On the IMTA Farming Practice
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Clarke, K.R.; Warwick, R.M. Change in Marine Communities an Approach to Statistical Analysis and Interpretation, 2nd ed.; PRIMER-E: Plymouth, UK, 2001; p. 172. [Google Scholar]
- FAO. The State of World Fisheries and Aquaculture 2020. Sustainability in action; Food and Agriculture Organization of the United Nations: Rome, Italy, 2020. [Google Scholar]
- Putro, S.P.; Widowati, W.; Suhartana, S.; Muhammad, F. The Application of Integrated Multi Trophic Aquaculture Using Stratified Double Net Cage for Aquaculture Sustainability. Int. J. Sci. Eng. 2015, 9, 85–89. [Google Scholar]
- Henchion, M.; Hayes, M.; Mullen, A.M.; Fenelon, M.A.; Tiwari, B.K. Future Protein Supply and Demand: Strategies and Factors Influencing a Sustainable Equilibrium. Foods 2017, 6, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dahuri, R. Cetak Biru Pembangunan Kelautan dan Perikanan: Menuju Indonesia yang Maju, Adil-Makmur dan Berdaulat [Blueprint for Marine and Fisheries Development: Towards an Advanced, Just-Prosperous and Sovereign for Indonesia]; Roda Bahari: Bogor, Indonesia, 2012. [Google Scholar]
- Food and Agriculture Organization of the United Nations. Fishery and Aquaculture Country Profiles: The Republic of Indonesia. 2020. Available online: http://www.fao.org/fishery/facp/IDN/en (accessed on 30 October 2020).
- Sampantamit, T.; Ho, L.; Lachat, C.; Sutummawong, N.; Sorgeloos, P.; Goethals, P.L. Aquaculture Production and Its Environmental Sustainability in Thailand: Challenges and Potential Solutions. Sustainability 2020, 12, 2010. [Google Scholar] [CrossRef] [Green Version]
- Tagliapietra, D.; Povilanskas, R.; Razinkovas-Baziukas, A.; Taminskas, J. Emerald Growth: A New Framework Concept for Managing Ecological Quality and Ecosystem Services of Transitional Waters. Water 2020, 12, 894. [Google Scholar] [CrossRef] [Green Version]
- Ape, F.; Manini, E.; Quero, G.M.; Luna, G.M.; Sarà, G.; Vecchio, P.; Brignoli, P.; Ansferri, S.; Mirto, S. Biostimulation of in situ microbial degradation processes in organically-enriched sediments mitigates the impact of aquaculture. Chemosphere 2019, 226, 715–725. [Google Scholar] [CrossRef] [PubMed]
- Lacoste, É.; Weise, A.M.; Lavoie, M.-F.; Archambault, P.; McKindsey, C.W. Changes in infaunal assemblage structure influence nutrient fluxes in sediment enriched by mussel biodeposition. Sci. Total. Environ. 2019, 692, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Putro, S.P. Environmental Quality Assessment of Fish Farming: Solutions toward Sustainable Aquaculture; Lambert Academic Publishing (LAP): Saarbrucken, Germany, 2010; p. 197. [Google Scholar]
- Clarke, K.R.; Gorley, R.N. PRIMER v6: User Manual/Tutorial; PRIMER-E: Plymouth, UK, 2006; p. 192. [Google Scholar]
- Tataranni, M.; Lardicci, C. Performance of some biotic indices in then real variable world: A case study at different spatial scales in North-Western Mediterranean Sea. Environ. Pollut. 2010, 158, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, R. Marine benthic faunal successional stages and related sedimentary activity. Sci. Mar. 2001, 65 (Suppl. 2), 107–119. [Google Scholar] [CrossRef] [Green Version]
- Pandey, V.; Thiruchitrambalam, G. Spatial and temporal variability of sandy intertidal macrobenthic communities and their relationship with environmental factors in a tropical island. Estuarine Coast. Shelf Sci. 2019, 224, 73–83. [Google Scholar] [CrossRef]
- Putro, S.P. Metode Sampling Penelitian Makrobentos dan Aplikasinya: Penentuan Tingkat Gangguan Lingkungan Akuakultur [Sampling Methods for Research on Macrobenthos and Its Applications: Assessment of Aquaculture Environmental Disturbance]; Graha Ilmu: Yogyakarta, Indonesia, 2014; p. 205. [Google Scholar]
- Putro, S.P.; Widowati, W.; Febria, I.J.; Muhammad, F.; Suhartana, S.; Suminto, S.; Sudaryono, A.; Koshio, S. Environmental assessment of polyculture farming practice based on macrobenthic assemblages: A study case at coastal area of Kaliwungu, Kendal (Central Java, Indonesia). J. Teknol. 2016, 78, 199–205. [Google Scholar]
- Dauvin, J.; Pezy, J.-P.; Baffreau, A.; Bachelet, Q.; Baux, N.; Méar, Y.; Murat, A.; Poizot, E. Effects of a salmon fish farm on benthic habitats in a high-energy hydrodynamic system: The case of the Rade de Cherbourg (English Channel). Aquaculture 2020, 518, 734832. [Google Scholar] [CrossRef]
- Barnes, R.S.K.; Hughes, R.N. An Introduction to Marine Ecology; Blackwell Scientific Publications: Boston, MA, USA, 1982. [Google Scholar]
- Morton, B.; Chan, K. Hunger rapidly overrides the risk of predation in the subtidal scavenger Nassarius siquijorensis (Gastropoda: Nassariidae): An energy budget and a comparison with the intertidal Nassarius festivus in Hong Kong. J. Exp. Mar. Biol. Ecol. 1999, 240, 213–228. [Google Scholar] [CrossRef]
- Gofas, S.; Bouchet, P. Nassarius Reticulatus: Mollusca Base. 2016. Available online: http://www.marinespecies.org/aphia.php?p=taxdetails&id=140513 (accessed on 27 August 2020).
- Rochmady. Aspek Bioekologi Kerang Lumpur Anodontia edentula Linnaeus, 1758 (BIVALVIA: LUCINIDAE) di Perairan Pesisir Kabupaten Muna [Bioecological Aspect on Muddy Clam, Anodontia Edentula Linnaeus at Coastal Area of Muna Distict]; Program Pascasarjana Universitas Hasanuddin: Makassar, Indonesia, 2011. [Google Scholar]
- Bouchet, P.; Gofas, S.; le Renard, J. Tellina versicolor: Mollusca Base. 2018. Available online: http://marinespecies.org/aphia.php?p=taxdetails&id=157004 (accessed on 2 September 2020).
- Polytraits-Team. Polytraits: A Database on Biological Traits of Polychaetes. LifewatchGreece, Hellenic Centre for Marine Research. 2017. Available online: http://polytraits.lifewatchgreece.eu (accessed on 2 September 2020).
- Mendes, C.L.T.; Soares-Gomes, A. Macrobenthic community structure in a Brazilian chocked lagoon system under environmental stress. Zoology 2011, 28, 365–378. [Google Scholar] [CrossRef] [Green Version]
- Brito, R.M.; Pompolo, S.D.G.; Magalhães, M.F.M.; De Barros, E.G.; Sakamoto-Hojo, E.T. Cytogenetic Characterization of Two Partamona Species (Hymenoptera, Apinae, Meliponini) by Fluorochrome Staining and Localization of 18S rDNA Clusters by FISH. Cytology 2005, 70, 373–380. [Google Scholar] [CrossRef] [Green Version]
- Kristensen, E. Organic matter diagenesis at the oxic/anoxic interface in coastal marine sediments, with emphasis on the role of burrowing animals. Hydrobiology 2000, 426, 1–24. [Google Scholar] [CrossRef]
- Samaniego, L.G.S. Distribution of Soft-Bottom Polychaetes Assemblages at Different Scales in Shallow Waters of the Northern Mediterranean Spanish Coast. Ph.D. Thesis, The Center for Advance Studies of Blanes, Barcelona, Spain, 2012; pp. 52–60. [Google Scholar]
- Heip, C.H.R.; Herman, P.M.J.; Soetaert, K. Indices of diversity and evenness. Oceanis 1998, 24, 61–87. [Google Scholar]
- Jost, L. The Relation between Evenness and Diversity. Diversity 2010, 2, 207–232. [Google Scholar] [CrossRef]
- Carras, M.A.; Knowler, D.; Pearce, C.M.; Hamer, A.; Chopin, T.; Weaire, T. A discounted cash-flow analysis of salmon monoculture and Integrated Multi-Trophic Aquaculture in eastern Canada. Aquac. Econ. Manag. 2019, 1–21. [Google Scholar] [CrossRef]
- Troell, M.; Joyce, A.; Chopin, T.; Neori, A.; Buschmann, A.H.; Fang, J.-G. Ecological engineering in aquaculture—Potential for integrated multi-trophic aquaculture (IMTA) in marine offshore systems. Aquaculture 2009, 297, 1–9. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Class | Family | Genus | ABUNDANCE (ind./grab) | |||||
|---|---|---|---|---|---|---|---|---|---|
| Sampling I | Sampling II | ||||||||
| I | II | III | I | II | III | ||||
| 1. | Gastropoda | Ellobiidae | Ellobium sp. | 2 | 4 | 36 | 6 | 0 | 11 |
| Columbellidae | Costoanachis sp. | 6 | 24 | 20 | 0 | 0 | 0 | ||
| Parvanachis sp. | 4 | 0 | 3 | 0 | 2 | 0 | |||
| Mitrella sp. | 4 | 0 | 0 | 0 | 4 | 0 | |||
| Pyrene sp. | 2 | 6 | 0 | 2 | 2 | 0 | |||
| Naticidae | Naticarius sp. | 4 | 0 | 1 | 0 | 0 | 0 | ||
| Nassariidae | Nassarius sp. | 4 | 18 | 48 | 7 | 6 | 6 | ||
| Terebridae | Terebra sp. | 0 | 0 | 0 | 1 | 4 | 0 | ||
| Pyramidellidae | Turbonilla sp. | 0 | 0 | 0 | 1 | 6 | 0 | ||
| Boonea sp. | 0 | 0 | 2 | 0 | 0 | 0 | |||
| Odostomia sp. | 0 | 0 | 2 | 0 | 0 | 0 | |||
| Cerithiidae | Clypeomorus sp. | 0 | 4 | 0 | 0 | 0 | 0 | ||
| Cerithium sp. | 0 | 0 | 0 | 0 | 2 | 0 | |||
| Architectonicidae | Architectonica sp. | 0 | 0 | 0 | 0 | 2 | 0 | ||
| Buccinidae | Neptunea sp. | 0 | 4 | 0 | 0 | 0 | 0 | ||
| Muricidae | Ocinebrina sp. | 0 | 0 | 2 | 0 | 0 | 0 | ||
| Ovulidae | Pellasimnia sp. | 0 | 0 | 1 | 0 | 0 | 0 | ||
| Fissurellidae | Tugali sp. | 0 | 0 | 0 | 0 | 0 | 1 | ||
| Mangeliidae | Agathotoma sp. | 0 | 0 | 2 | 0 | 0 | 0 | ||
| Pyrgocythara sp. | 0 | 0 | 2 | 0 | 0 | 2 | |||
| Epitoniidae | Epitonium sp. | 0 | 0 | 8 | 0 | 0 | 0 | ||
| Volutidae | Ericusa sp. | 0 | 0 | 7 | 0 | 0 | 0 | ||
| Muricidae | Murex sp. | 0 | 0 | 1 | 0 | 0 | 0 | ||
| Eupleura sp. | 0 | 0 | 2 | 0 | 0 | 0 | |||
| Ocinebrina sp. | 0 | 0 | 2 | 0 | 0 | 0 | |||
| Marginellidae | Prunum sp. | 0 | 0 | 5 | 0 | 0 | 0 | ||
| Turridae | Glyphoturris sp. | 0 | 0 | 3 | 0 | 0 | 0 | ||
| Babyloniidae | Babylonia sp. | 0 | 0 | 4 | 0 | 0 | 0 | ||
| Mitridae | Mitra sp. | 0 | 0 | 1 | 0 | 0 | 0 | ||
| Skeneidae | Leucorhynchia sp. | 0 | 0 | 5 | 0 | 0 | 0 | ||
| Turbinidae | Turbo sp. | 0 | 0 | 1 | 0 | 0 | 0 | ||
| Trochidae | Austrocochlea sp. | 0 | 0 | 2 | 0 | 0 | 0 | ||
| Clanculus sp. | 0 | 0 | 1 | 0 | 0 | 0 | |||
| Batillariidae | Batillaria sp. | 0 | 0 | 3 | 0 | 0 | 2 | ||
| Assimineidae | Assiminea sp. | 0 | 0 | 11 | 0 | 0 | 0 | ||
| 2. | Bivalvia | Lucinidae | Anodontia sp. | 10 | 14 | 14 | 15 | 10 | 7 |
| Lucina sp. | 0 | 0 | 36 | 0 | 0 | 0 | |||
| Nuculanidae | Nuculana sp. | 0 | 0 | 59 | 2 | 0 | 5 | ||
| Tellinidae | Tellina sp. | 2 | 6 | 0 | 2 | 2 | 0 | ||
| Archidae | Anadara sp. | 0 | 2 | 30 | 0 | 0 | 0 | ||
| Arca sp. | 0 | 0 | 16 | 0 | 0 | 6 | |||
| Cardiidae | Fulvia sp. | 0 | 2 | 0 | 0 | 0 | 0 | ||
| Ostreidae | Ostrea sp. | 0 | 0 | 8 | 0 | 0 | 0 | ||
| Nacellidae | Cellana sp. | 0 | 0 | 13 | 0 | 0 | 2 | ||
| Neotiidae | Arcopsis sp. | 0 | 0 | 35 | 0 | 0 | 0 | ||
| Crassatellidae | Crassatella sp. | 0 | 0 | 2 | 0 | 0 | 0 | ||
| Fissurellidae | Diodora sp. | 0 | 0 | 4 | 0 | 0 | 0 | ||
| Emarginula sp. | 0 | 0 | 1 | 0 | 0 | 0 | |||
| 3. | Polychaeta | Lumbrineridae | Lumbrineris sp. | 2 | 3 | 0 | 0 | 2 | 0 |
| Nephtyidae | Nephtys sp. | 3 | 0 | 2 | 0 | 0 | 0 | ||
| Sternaspidae | Sternaspis sp. | 4 | 3 | 0 | 2 | 0 | 0 | ||
| Nereididae | Nereis sp. | 3 | 18 | 0 | 1 | 6 | 0 | ||
| Namanereis sp. | 0 | 4 | 2 | 0 | 0 | 0 | |||
| Arenicolidae | Arenicolides sp. | 0 | 3 | 0 | 0 | 0 | 0 | ||
| Capitellidae | Heteromastus sp. | 0 | 2 | 0 | 0 | 0 | 0 | ||
| Capitella sp. | 1 | 4 | 0 | 0 | 0 | 0 | |||
| Mediomastus sp. | 0 | 1 | 0 | 0 | 0 | 3 | |||
| 4. | Crustacea | Penaidae | Penaeus sp. | 0 | 1 | 1 | 0 | 0 | 1 |
| Gammaridae | Talorchestia sp. | 6 | 0 | 0 | 0 | 0 | 0 | ||
| Anaspidacea | Anaspides sp. | 1 | 0 | 0 | 0 | 0 | 0 | ||
| 5. | Ophiuroidea | Ophiuridae | Ophioplocus sp. | 0 | 7 | 0 | 0 | 0 | 0 |
| Total abundance (A) | 58 | 139 | 398 | 39 | 48 | 46 | |||
| Number of genera (s) | 15 | 19 | 40 | 9 | 12 | 11 | |||
| Diversity index (H’) | 2.53 | 2.63 | 2.94 | 1.78 | 2.31 | 2.15 | |||
| Evenness index (e) | 0.94 | 0.90 | 0.80 | 0.81 | 0.93 | 0.90 | |||
| Dominance index (C) | 0.09 | 0.09 | 0.03 | 0.23 | 0.11 | 0.14 | |||
| Sampling Location | Diversity Index (H’) | Pielou Index (J’) | Dominance Index (C) |
|---|---|---|---|
| IMTA I | 2.53 | 0.94 | 0.09 |
| IMTA II | 1.78 | 0.81 | 0.23 |
| Monoculture I | 2.63 | 0.90 | 0.09 |
| Monoculture II | 2.31 | 0.93 | 0.11 |
| Reference I | 2.94 | 0.80 | 0.03 |
| Reference II | 2.15 | 0.90 | 0.14 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Putro, S.P.; Sharani, J.; Widowati; Adhy, S.; Suryono. Biomonitoring of the Application of Monoculture and Integrated Multi-Trophic Aquaculture (IMTA) Using Macrobenthic Structures at Tembelas Island, Kepulauan Riau Province, Indonesia. J. Mar. Sci. Eng. 2020, 8, 942. https://doi.org/10.3390/jmse8110942
Putro SP, Sharani J, Widowati, Adhy S, Suryono. Biomonitoring of the Application of Monoculture and Integrated Multi-Trophic Aquaculture (IMTA) Using Macrobenthic Structures at Tembelas Island, Kepulauan Riau Province, Indonesia. Journal of Marine Science and Engineering. 2020; 8(11):942. https://doi.org/10.3390/jmse8110942
Chicago/Turabian StylePutro, Sapto P., Jeanny Sharani, Widowati, Satriyo Adhy, and Suryono. 2020. "Biomonitoring of the Application of Monoculture and Integrated Multi-Trophic Aquaculture (IMTA) Using Macrobenthic Structures at Tembelas Island, Kepulauan Riau Province, Indonesia" Journal of Marine Science and Engineering 8, no. 11: 942. https://doi.org/10.3390/jmse8110942