
Advances in the Role of Dark Septate Endophytes in the Plant Resistance to Abiotic and Biotic Stresses

 , ,
, ,
Abstract
:1. Introduction
2. Nutrient Uptake and Plant Growth Promotion
3. Abiotic Stress
4. Biotic Stress: Biological Control
4.1. Mechanisms of Action
4.2. Biological Control Assays
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Sherwood, M.; Carroll, G. Fungal succession on needles and young twigs of old-growth Douglas fir. Mycologia 1974, 66, 499–506. [Google Scholar] [CrossRef]
- Carroll, G. Fungal endophytes in stems and leaves–from latent pathogen to mutualistic symbiont. Ecology 1988, 69, 2–9. [Google Scholar] [CrossRef]
- Stone, J.K.; Polishook, J.D.; White, J.R.J. Endophytic fungi. In Biodiversity of Fungi: Inventory and Monitoring Methods; Mueller, G., Bills, G.F., Foster, M.S., Eds.; Elsevier: Burlington, VT, USA, 2004; pp. 241–270. [Google Scholar]
- Rodriguez, R.J.; White, J.F., Jr.; Arnold, A.E.; Redman, R.S. Fungal endophytes: Diversity and functional roles. N. Phytol. 2009, 182, 314–330. [Google Scholar] [CrossRef] [PubMed]
- Arnold, A.E.; Lutzoni, F. Diversity and host range of foliar fungal endophytes: Are tropical leaves biodiversity hotspots? Ecology 2007, 88, 541–549. [Google Scholar] [CrossRef]
- Jumpponen, A.; Trappe, J.M. Dark septate endophytes: A review of facultative biotrophic root-colonizing fungi. N. Phytol. 1998, 140, 295–310. [Google Scholar] [CrossRef]
- Han, L.; Zuo, Y.; He, X.; Hou, Y.; Li, M.; Li, B. Plant identity and soil variables shift the colonisation and species composition of dark septate endophytes associated with medicinal plants in a northern farmland in China. Appl. Soil. Ecol. 2021, 167, 104042. [Google Scholar] [CrossRef]
- Mandyam, K.; Jumpponen, A. Seeking the elusive function of theroot-colonising dark septate endophytic fungi. Stud. Mycol. 2005, 53, 173–189. [Google Scholar] [CrossRef] [Green Version]
- Day, M.J.; Currah, R.S. Role of selected dark septate endophyte species and other hyphomycetes as saprobes on moss gametophytes. Botany 2011, 89, 349–359. [Google Scholar] [CrossRef]
- Schmidt, S.K.; Sobieniak-Wiseman, L.C.; Kageyama, S.A.; Halloy, S.R.P.; Schadt, C.W. Mycorrhizal and dark-septate fungi in plant roots above 4270 meters elevation in the Andes and Rocky Mountains. Arct. Antarct. Alp. Res. 2008, 40, 576–583. [Google Scholar] [CrossRef] [Green Version]
- Newsham, K.K. A meta-analysis of plant responses to dark septate root endophytes. N. Phytol. 2011, 190, 783–793. [Google Scholar] [CrossRef]
- Porras-Alfaro, A.; Herrera, J.; Sinsabaugh, R.L.; Odenbach, K.J.; Lowrey, T.; Natvig, D.O. Novel root fungal consortium associated with a dominant desert grass. Appl. Environ. Microbiol. 2008, 74, 2805–2813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knapp, D.G.; Pintye, A.; Kovács, G.M. The dark side is not fastidious–dark septate endophytic fungi of native and invasive plants of semiarid sandy areas. PLoS ONE 2012, 7, e32570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alberton, O.; Kuyper, T.W.; Summerbell, R.C. Dark septate root endophytic fungi increase growth of Scots pine seedlings under elevated CO2 through enhanced nitrogen use efficiency. Plant Soil 2010, 328, 459–470. [Google Scholar] [CrossRef] [Green Version]
- Sonjak, S.; Beguiristain, T.; Leyval, C.; Regvar, M. Temporal temperature gradient gel electrophoresis (TTGE) analysis of arbuscular mycorrhizal fungi associated with selected plants from saline and metal polluted environments. Plant Soil 2009, 314, 25–34. [Google Scholar] [CrossRef]
- Regvar, M.; Likar, M.; Piltaver, A.; Kugonič, N.; Smith, J.E. Fungal community structure under goat willows (Salix caprea L.) growing at metal polluted site: The potential of screening in a model phytostabilisation study. Plant Soil 2010, 330, 345–356. [Google Scholar] [CrossRef]
- Deram, A.; Languereau, F.; Haluwyn, C.V. Mycorrhizal and endophytic fungal colonization in Arrhenatherum elatius L. roots according to the soil contamination in heavy metals. Soil Sediment Contam. 2011, 20, 114–127. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, T.; Zhao, Z.W. Colonization characteristics and composition of dark septate endophytes (DSE) in a lead and zinc slag heap in Southwest China. Soil Sediment. Contam. 2013, 22, 532–545. [Google Scholar] [CrossRef]
- Knapp, D.G.; Kovács, G.M.; Zajta, E.; Groenewald, J.Z.; Crous, P.W. Dark septate endophytic pleosporalean genera from semiarid areas. Persoonia 2015, 35, 87–100. [Google Scholar] [CrossRef] [Green Version]
- Lugo, M.A.; Reinhart, K.O.; Menoyo, E.; Crespo, E.M.; Urcelay, C. Plant functional traits and phylogenetic relatedness explain variation in associations with root fungal endophytes in an extreme arid environment. Mycorrhiza 2015, 25, 85–95. [Google Scholar] [CrossRef]
- Dos Santos, S.G.; da Silva, P.R.A.; Garcia, A.C.; Zilli, J.É.; Berbara, R.L.L. Dark septate endophyte decreases stress on rice plants. Braz. J. Microbiol. 2017, 48, 333–341. [Google Scholar] [CrossRef]
- Barrow, J.R. Atypical morphology of dark septate fungal root endophytes of Bouteloua in arid southwestern USA rangelands. Mycorrhiza 2003, 13, 239–247. [Google Scholar] [CrossRef] [PubMed]
- McLellan, C.A.; Turbyville, T.J.; Wijeratne, E.K.; Kerschen, A.; Vierling, E.; Queitsch, C.; Gunatilaka, A.L. A rhizosphere fungus enhances Arabidopsis thermotolerance through production of an HSP90 inhibitor. Plant Physiol. 2007, 145, 174–182. [Google Scholar] [CrossRef] [PubMed]
- Ban, Y.; Tang, M.; Chen, H.; Xu, Z.; Zhang, H.; Yang, Y. The response of dark septate endophytes (DSE) to heavy metals in pure culture. PLoS ONE 2012, 7, e47968. [Google Scholar] [CrossRef]
- Zhan, F.; He, Y.; Zu, Y.; Li, T.; Zhao, Z. Characterization of melanin isolated from a dark septate endophyte (DSE), Exophiala pisciphila. World J. Microbiol. Biotechnol. 2011, 27, 2483–2489. [Google Scholar] [CrossRef]
- Gaber, D.A.; Berthelot, C.; Camehl, I.; Kovács, G.M.; Blaudez, D.; Franken, P. Salt Stress Tolerance of Dark Septate Endophytes Is Independent of Melanin Accumulation. Front. Microbiol. 2020, 11, 562931. [Google Scholar] [CrossRef] [PubMed]
- Berthelot, C.; Perrin, Y.; Leyval, C.; Blaudez, C. Melanization and ageing are not drawbacks for successful agro-transformation of dark septate endophytes. Fungal Biol. 2017, 121, 652–663. [Google Scholar] [CrossRef] [Green Version]
- Ohki, T.; Masuya, H.; Yonezawa, M.; Usuki, F.; Narisawa, K.; Hashiba, T. Colonization process of the root endophytic fungus Heteroconium chaetospira in roots of Chinese cabbage. Mycoscience 2002, 43, 191–194. [Google Scholar] [CrossRef]
- Currah, R.S.; Tsuneda, A.; Murakami, S. Morphology and ecology of Phialocephala fortinii in roots of Rhododendron brachycarpum. Can. J. Bot. 1993, 71, 1639–1644. [Google Scholar] [CrossRef]
- Addy, H.D.; Piercey, M.M.; Currah, R.S. Microfungal endophytes in roots. Can. J. Bot. 2005, 83, 1–13. [Google Scholar] [CrossRef]
- Grünig, C.R.; Queloz, V.; Sieber, T.N. Structure of Diversity in Dark Septate Endophytes: From Species to Genes. In Endophytes of Forest Trees: Biology and Applications, Forestry Sciences 80; Pirttila, A.M., Frank, A.C., Eds.; Springer Science Business Media B.V.: Berlin/Heidelberg, Germany, 2011; pp. 3–30. [Google Scholar]
- Grünig, C.R.; Queloz, V.; Sieber, T.N.; Holdenrieder, O. Dark septate endophytes (DSE) of the Phialocephala fortinii s.l.–Acephala applanata species complex in tree roots: Classification, population biology and ecology. Botany 2008, 86, 1355–1369. [Google Scholar] [CrossRef]
- Stoyke, G.; Currah, R.S. Endophytic fungi from the mycorrhizae of alpine ericoid plants. Can. J. Bot. 1991, 69, 347–352. [Google Scholar] [CrossRef]
- Addy, H.D.; Hambleton, S.; Currah, R.S. Distribution and molecular characterization of the root endophyte Phialocephala fortinii along an environmental gradient in the boreal forest of Alberta. Mycol. Res. 2000, 104, 1213–1221. [Google Scholar] [CrossRef]
- Stroheker, S.; Dubach, V.; Vögtli, I.; Sieber, T.N. Investigating Host Preference of Root Endophytes of Three European Tree Species, with a Focus on Members of the Phialocephala fortinii—Acephala applanata Species Complex (PAC). J. Fungi 2021, 7, 317. [Google Scholar] [CrossRef] [PubMed]
- Brenn, N.; Menkis, A.; Grünig, C.R.; Sieber, T.N.; Holdenrieder, O. Community structure of Phialocephala fortinii s. lat. in European tree nurseries, and assessment of the potential of the seedlings as dissemination vehicles. Mycol. Res. 2008, 112, 650–662. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.J.K.; Wilcox, H.E. New species of ectendomycorrhizal and pseudomycorrhizal fungi: Phialophora finlandia, Chloridium paucisporum, and Phialocephala fortinii. Mycologia 1985, 77, 951–958. [Google Scholar] [CrossRef]
- Maciá-Vicente, J.G.; Glynou, K.; Piepenbring, M. A new species of Exophiala associated with roots. Mycol. Prog. 2016, 15, 1. [Google Scholar] [CrossRef]
- Najafzadeh, M.J.; Dolatabadi, S.; Saradeghi Keisari, M.; Naseri, A.; Feng, P.; de Hoog, G.S. Detection and identification of opportunistic Exophiala species using the rolling circle amplification of ribo-somal internal transcribed spacers. J. Microbiol. Methods 2013, 94, 338–342. [Google Scholar] [CrossRef]
- Wang, Q.; Shen, S.K.; Zhang, A.L.; Wu, C.Y.; Wu, F.Q.; Zhang, X.J.; Wang, Y.H. Isolation and diversity analyses of endophytic fungi from Paris polyphylla var. Yunnanensis. Zhongguo Zhong Yao Za Zhi 2013, 38, 3838–3844. [Google Scholar]
- Piercey, M. Dark Septate Root Endophytic (DSE) Fungi in Canada: Geographic Distribution and Patterns of Variation. Master’s Thesis, Department of Biological Sciences, University of Alberta, Edmonton, AB, Canada, 2003. [Google Scholar]
- Summerbell, R.C. Microfungi associated with the mycorrhizal mantle of and adjacent microhabitats within the rhizosphere of black spruce in boreal Canada. Can. J. Bot. 1989, 67, 1085–1095. [Google Scholar] [CrossRef]
- Nuaima, R.H.; Ashrafi, S.; Maier, W.; Heuer, H. Fungi isolated from cysts of the beet cyst nematode parasitized its eggs and counterbalanced root damages. J. Pest Sci. 2021, 94, 563–572. [Google Scholar] [CrossRef]
- Gene, J.; Verdejo-Lucas, S.; Stchigel, A.M.; Sorribas, F.J.; Guarro, J. Microbial parasites associated with Tylenchulus semipenetrans in citrus orchards of Catalonia, Spain. Biocontrol Sci. Technol. 2005, 15, 721–731. [Google Scholar] [CrossRef]
- Harrington, T.C.; McNew, D.L. Phylogenetic analysis places the Phialophora-like anamorph genus Cadophora in the Helotiales. Mycotaxon 2003, 87, 141–151. [Google Scholar]
- Ellis, M.B. More Dematiaceous Hyphomycetes; Commonwealth Mycological Institute: Kew, UK, 1976. [Google Scholar]
- Currah, R.S.; Sigler, L.; Hambleton, S. New records and new taxa of fungi from the mycorrhizae of terrestrial orchids of Alberta Canada. Can. J. Bot. 1987, 65, 2473–2482. [Google Scholar] [CrossRef]
- Caldwell, B.A.; Jumpponen, A.; Trappe, J.M. Utilization of major detrital substrates by dark-septate root endophytes. Mycologia 2000, 92, 230–232. [Google Scholar] [CrossRef] [Green Version]
- Della Monica, I.F.; Saparrat, M.C.N.; Godeas, A.M.; Scervino, J.M. The co-existence between DSE and AMF symbionts affects plant P pools through P mineralization and solubilization processes. Fungal Ecol. 2015, 17, 10–17. [Google Scholar] [CrossRef]
- Vergara, C.; Araujo, K.E.C.; Alves, L.S.; de Souza, S.R.; Santos, L.A.; Santa-Catarina, C.; da Silva, K.; Pereira, G.M.D.; Xavier, G.R.; Zilli, J.E. Contribution of dark septate fungi to the nutrient uptake and growth of rice plants. Braz. J. Microbiol. 2018, 49, 67–78. [Google Scholar] [CrossRef] [PubMed]
- Vergara, C.; Araujo, K.E.C.; Sperandio, M.V.L.; Santos, L.A.; Urquiaga, S.; Zilli, J.E. Dark septate endophytic fungi increase the activity of proton pumps, efficiency of 15N recovery from ammonium sulphate, N content, and micronutrient levels in rice plants. Braz. J. Microbiol. 2019, 50, 825–838. [Google Scholar] [CrossRef]
- Xu, R.; Li, T.; Shen, M.; Yang, Z.L.; Zhao, Z.W. Evidence for a Dark Septate Endophyte (Exophiala Pisciphila, H93) Enhancing Phosphorus Absorption by Maize Seedlings. Plant Soil 2020, 452, 249–266. [Google Scholar] [CrossRef]
- Yakti, W.; Kovács, G.M.; Vági, P.; Franken, P. Impact of dark septate endophytes on tomato growth and nutrient uptake. Plant Ecol. Divers. 2018, 11, 637–648. [Google Scholar] [CrossRef] [Green Version]
- Wu, F.L.; Qu, D.H.; Tian, W.; Wang, M.Y.; Chen, F.Y.; Li, K.K.; Sun, Y.D.; Su, Y.H.; Yang, L.N.; Su, H.Y.; et al. Transcriptome analysis for understanding the mechanism of dark septate endophyte S16 in promoting the growth and nitrate uptake of sweet Cherry. J. Integr. Agric. 2021, 20, 1819–1831. [Google Scholar] [CrossRef]
- Haselwandter, K.; Read, D.J. Fungal associations of roots of dominant and sub-dominant plants in high-alpine vegetation systems with special reference to mycorrhiza. Oecologia 1980, 45, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.Q.; Lv, Y.L.; Meng, Z.X. The promoting role of an isolate of dark-septate fungus on its host plant Saussurea involucrata Kar et Kir. Mycorrhiza 2010, 20, 127–135. [Google Scholar] [CrossRef]
- Li, X.; He, X.L.; Hou, L.F.; Ren, Y.; Wang, S.J.; Su, F. Dark septate endophytes isolated from a xerophyte plant promote the growth of Ammopiptanthus mongolicus under drought condition. Sci. Rep. 2018, 8, 7896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuo, Y.; Su, F.; He, X.; Li, M. Colonization by dark septate endophytes improves the growth of Hedysarum scoparium under multiple inoculum levels. Symbiosis 2020, 82, 201–214. [Google Scholar] [CrossRef]
- Surono; Narisawa, K. The cellulolytic activity and symbiotic potential of dark septate endophytic fungus Phialocephala fortinii to promote non-mycorrhizal plants growth. IOP Conf. Ser. Earth Environ. Sci. 2021, 648, 012165. [Google Scholar] [CrossRef]
- Lin, L.C.; Lin, C.Y.; Lin, W.R.; Tung, Y.T.; Wu, J.H. Effects of ericoid Mycorrhizal fungi or dark septate endophytic fungi on the secondary metabolite of Rhododendron pseudochrysanthum (R. Morii) seedlings. Appl. Ecol. Environ. Res. 2021, 19, 1221–1232. [Google Scholar] [CrossRef]
- Hesse, U.; Schoberlein, W.; Wittenmayer, L.F.; Orster, K.; Warnstorff, K.; Diepenbrock, W.; Merbach, W. Effects of Neotyphodium endophytes on growth, reproduction and drought: Tress tolerance of three Lolium perenne L. genotypes. Grass Forage Sci. 2003, 58, 407–415. [Google Scholar] [CrossRef]
- Andrade-Linares, D.R.; Grosch, R.; Restrepo, S.; Krumbein, A.; Franken, P. Effects of dark septate endophytes on tomato plant performance. Mycorrhiza 2011, 21, 413–422. [Google Scholar] [CrossRef]
- Stroheker, S.; Dubach, V.; Queloz, V.; Sieber, T.N. Resilience of Phialocephala fortinii s.l. e Acephala applanata communities e effects of disturbance and strain introduction. Fungal Ecol. 2018, 31, 19–28. [Google Scholar] [CrossRef]
- Likar, M.; Regvar, M. Isolates of dark septate endophytes reduce metal uptake and improve physiology of Salix caprea L. Plant Soil 2013, 370, 593–604. [Google Scholar] [CrossRef]
- Xu, R.B.; Li, T.; Cui, H.L.; Wang, J.L.; Yu, X.; Ding, Y.H.; Wang, C.J.; Yang, Z.L.; Zhao, Z.W. Diversity and characterization of Cd-tolerant dark septate endophytes (DSEs) associated with the roots of Nepal alder (Alnus nepalensis) in a metal mine tailing of southwest China. Appl. Soil Ecol. 2015, 93, 11–18. [Google Scholar] [CrossRef]
- Hou, L.; Yu, J.; Zhao, L.; He, X. Dark Septate Endophytes Improve the Growth and the Tolerance of Medicago sativa and Ammopiptanthus mongolicus under Cadmium Stress. Front. Microbiol. 2020, 10, 3061. [Google Scholar] [CrossRef]
- Yu, Y.; Teng, Z.; Mou, Z.; Lv, Y.; Li, T.; Chen, S.; Zhao, D.; Zhao, Z. Melatonin confers heavy metal-induced tolerance by alleviating oxidative stress and reducing the heavy metal accumulation in Exophiala pisciphila, a dark septate endophyte (DSE). BMC Microbiol. 2021, 21, 40. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.Q.; Liu, H.B.; Xie, Y.Y.; Zhang, Y.G.; Xu, Q.Q.; Mao, L.J.; Li, X.J.; Chen, J.C.; Lin, F.C.; Zhang, C.L. Effect of the dark septate endophytic fungus Acrocalymma vagum on heavy metal content in tobacco leaves. Symbiosis 2018, 74, 89–95. [Google Scholar] [CrossRef]
- He, C.; Wang, W.; Hou, J. Plant growth and soil microbial impacts of enhancing licorice with inoculating dark septate endophytes under drought stress. Front. Microbiol. 2019, 10, 2277. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wei, X. Dark septate endophyte improves the drought-stress resistance of Ormosia hosiei seedlings by altering leaf morphology and photosynthetic characteristics. Plant Ecol. 2021, 222, 761–771. [Google Scholar] [CrossRef]
- Boukhris, A.; Laffont-Schwob, I.; Rabier, J.; Salducci, M.D.; El Kadri, L.; Tonetto, A.; Tatoni, T.; Chaieb, M. Changes in mesophyll element distribution and phytometabolite contents involved in fluoride tolerance of the arid gypsum-tolerant plant species Atractylis serratuloides Sieber ex Cass. (Asteraceae). Environ. Sci. Pol. Res. 2015, 22, 7918–7929. [Google Scholar] [CrossRef] [PubMed]
- Chung, Y.A.; Sinsabaugh, R.L.; Kuske, C.R.; Reed, S.C.; Rudgers, J.A. Spatial variation in edaphic characteristics is a stronger control than nitrogen inputs in regulating soil microbial effects on a desert grass. J. Arid Environ. 2017, 142, 59–65. [Google Scholar] [CrossRef]
- Li, X.; He, X.L.; Zhou, Y.; Hou, Y.T.; Zuo, Y.L. Effects of dark septate endophytes on the performance of Hedysarum scoparium under water deficit stress. Front. Plant Sci. 2019, 10, 903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, C.; Zeng, Q.; Chen, Y.; Chen, C.; Wang, W.; Hou, J.; Li, X. Colonization by dark septate endophytes improves the growth and rhizosphere soil microbiome of licorice plants under different water treatments. Appl. Soil Ecol. 2021, 166, 103993. [Google Scholar] [CrossRef]
- He, C.; Wang, W.; Hou, J.; Li, X. Dark Septate Endophytes Isolated From Wild Licorice Roots Grown in the Desert Regions of Northwest China Enhance the Growth of Host Plants Under Water Deficit Stress. Front. Microbiol. 2021, 12, 522449. [Google Scholar] [CrossRef]
- Mateu, M.G.; Baldwin, A.H.; Maul, J.E.; Yarwood, S.A. Dark septate endophyte improves salt tolerance of native and invasive lineages of Phragmites australis. ISME J. 2020, 14, 1943–1954. [Google Scholar] [CrossRef] [PubMed]
- Hou, L.; Li, X.; He, X. Effect of dark septate endophytes on plant performance of Artemisia ordosica and associated soil microbial functional group abundance under salt stress. Appl. Soil Ecol. 2021, 165, 103998. [Google Scholar] [CrossRef]
- Tellenbach, C.; Sumarah, M.W.; Grunig, C.R.; Miller, J.D. Inhibition of Phytophthora species by secondary metabolites produced by the dark septate endophyte Phialocephala europaea. Fungal Ecol. 2013, 6, 12–18. [Google Scholar] [CrossRef]
- Deng, X.; Song, X.; Halifu, S.; Yu, W.; Song, R. Effects of Dark Septate Endophytes Strain A024 on Damping-off Biocontrol, Plant Growth, and the Rhizosphere Soil Environment of Pinus sylvestris var. mongolica Annual Seedlings. Plants 2020, 9, 913. [Google Scholar] [CrossRef]
- Narisawa, K.; Usuki, F.; Hashiba, T. Control of Verticillium yellows in Chinese cabbage by the dark septate endophytic fungus LtVB3. Phytopathology 2004, 94, 412–418. [Google Scholar] [CrossRef] [Green Version]
- Scervino, J.M.; Gottlieb, A.; Silvani, V.A.; Pergola, M.; Fernandez, L.; Godeas, A.M. Exudates of dark septate endophyte (DSE) modulate the development of the arbuscular mycorrhizal fungus (AMF) Gigaspora rosea. Soil Biol. Biochem. 2009, 41, 1753–1756. [Google Scholar] [CrossRef]
- Xiang, L.B.; Gong, S.J.; Yang, L.J.; Hao, J.J.; Xue, M.F.; Zeng, F.S. Biocontrol potential of endophytic fungi in medicinal plants from Wuhan Botanical Garden in China. Biol. Control 2016, 94, 47–55. [Google Scholar] [CrossRef]
- Jumpponen, A. Dark septate endophytes—are they mycorrhizal? Mycorrhiza 2001, 11, 207–211. [Google Scholar] [CrossRef]
- Schadt, C.W.; Mullen, R.B.; Schmidt, S.K. Isolation and phylogenetic identifi cation of a dark-septate fungus associated with the alpine plant Ranunculus adoneus. N. Phytol. 2001, 150, 747–755. [Google Scholar] [CrossRef]
- Kernaghan, G.; Patriquin, G. Host associations between fungal root endophytes and boreal trees. Microb. Ecol. 2011, 62, 460–473. [Google Scholar] [CrossRef]
- Mandyam, K.; Fox, C.; Jumpponen, A. Septate endophyte colonization and host responses of grasses and forbs native to a tallgrass prairie. Mycorrhiza 2012, 22, 109–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayerhofer, M.S.; Kernaghan, G.; Harper, K.A. The effects of fungal root endophytes on plant growth: A meta-analysis. Mycorrhiza 2013, 23, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Heredia-Acuña, C.; Alarcón, A.; Hernández-Cuevas, L.V.; Ferrera-Cerrato, R.; Almaraz-Suarez, J.J. Diversidad, Ecología e Importancia Potencial De Los Hongos Endófitos Septados Obscuros en México. Bot. Sci. 2014, 92, 321–333. [Google Scholar] [CrossRef] [Green Version]
- Usuki, F.; Narisawa, K. A mutualistic symbiosis between a dark septate endophytic fungus, Heteroconium chaetospira, and a nonmycorrhizal plant, Chinese cabbage. Mycologia 2007, 99, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Haselwandter, K.; Read, D.J. The significance of a root-fungus association in two Carex species of high-alpine plant communities. Oecologia 1982, 53, 352–354. [Google Scholar] [CrossRef] [PubMed]
- Diene, O.; Wang, W.; Narisawa, K. Pseudosigmoidea ibarakiensis sp. nov., a dark septate endophytic fungus from a cedar forest in Ibaraki, Japan. Microbes Environ. 2013, 28, 381–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahmoud, R.S.; Narisawa, K. A New Fungal Endophyte, Scolecobasidium humicola, promotes tomato growth under organic nitrogen conditions. PLoS ONE 2013, 8, e78746. [Google Scholar] [CrossRef]
- Richard, C.; Fortin, J.A.; Fortin, A. Protective effect of an ec-tomycorrhizal fungus against the root pathogen Mycelium radicis atrovirens. Can. J. For. Res. 1971, 1, 246–251. [Google Scholar] [CrossRef]
- Reininger, V.; Sieber, T.N. Mycorrhiza reduces adverse effects of dark septate endophytes (DSE) on growth of conifers. PLoS ONE 2012, 7, e42865. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.C.; Chen, S.J.; Li, K.T. Symbiotic fungi in nature Finnish peat moss promote vegetative growth in rabbiteye blueberry cuttings. Hortic. Environ. Biotechnol. 2021, 62, 191–198. [Google Scholar] [CrossRef]
- Guo, X.; Wan, Y.; Shakeel, M.; Wang, D.; Xiao, L. Effect of mycorrhizal fungi inoculation on bacterial diversity, community structure and fruit yield of blueberry. Rhizosphere 2021, 19, 100360. [Google Scholar] [CrossRef]
- Jumpponen, A.; Mattson, K.G.; Trappe, J.M. Mycorrhizal functioning of Phialocephala fortinii with Pinus contorta on glacier forefront soil: Interactions with soil nitrogen and organic matter. Mycorrhiza 1998, 7, 261–265. [Google Scholar] [CrossRef] [PubMed]
- Narisawa, K.; Kawamata, H.; Currah, R.S.; Hashiba, T. Suppression of Verticillium wilt in eggplant by some fungal root endophytes. Eur. J. Plant Pathol. 2002, 108, 103–109. [Google Scholar] [CrossRef]
- Vohník, M.; Albrechtova, J.; Vosatka, M. The inoculation with Oidiodendron maius and Phialocephala fortinii alters phosphorus and nitrogen uptake, foliar C:N ratio and root biomass distribution in Rhododendron cv. Azurro. Symbiosis 2005, 40, 87–96. [Google Scholar]
- Landolt, M.; Stroheker, S.; Queloz, V.; Gall, A.; Sieber, T.N. Does water availability influence the abundance of species of the Phialocephala fortinii s.l.-Acephala applanata complex (PAC) in roots of pubescent oak (Quercus pubescens) and Scots pine (Pinus sylvestris)? Fungal Ecol. 2020, 44, 100904. [Google Scholar] [CrossRef]
- Khan, A.L.; Hamayun, M.; Ahmad, N.; Waqas, M.; Kang, S.M.; Kim, Y.H.; Lee, I.J. Exophiala sp. LHL08 reprograms Cucumis sativus to higher growth under abiotic stresses. Physiol. Plant 2011, 143, 329–343. [Google Scholar] [CrossRef]
- Berthelot, C.; Chalot, M.; Leyval, C.; Blaudez, D. From darkness to light: Emergence of the mysterious dark septate endophytes in plant growth promotion and stress alleviation. In Endophytes for a Growing World; Hodkinson, T.R., Murphy, B.R., Saunders, M.J., Doohan, F.M., Eds.; Cambridge University Press: Cambridge, UK, 2019; pp. 143–164. [Google Scholar]
- Farias, G.C.; Nunes, K.G.; Soares, M.A.; de Siquiera, K.A.; Lima, W.C.; Neves, A.L.R.; de Lacerda, C.F.; Filho, E.G. Dark septate endophytic fungi mitigate the effects of salt stress on cowpea plants. Braz. J. Microbiol. 2020, 51, 243–253. [Google Scholar] [CrossRef]
- Baltruschat, H.; Fodor, J.; Harrach, B.D.; Niemczyk, E.; Barna, B.; Gullner, G.; Janeczko, A.; Kogel, K.H.; Schafer, P.; Schwarczinger, I.; et al. Salt tolerance of barley induced by the root endophyte Piriformospora indica is associated with a strong increase in antioxidants. N. Phytol. 2008, 180, 501–510. [Google Scholar] [CrossRef] [PubMed]
- Abdelaziz, M.E.; Kim, D.; Ali, S.; Fedoroff, N.V.; Al-Babili, S. The endophytic fungus Piriformospora indica enhances Arabidopsis thaliana growth and modulates Na+/K+ homeostasis under salt stress conditions. Plant Sci. 2017, 263, 107–115. [Google Scholar] [CrossRef] [Green Version]
- Gonçalves, D.R.; Pena, R.; Zotz, G.M.; Albach, D.C. Effects of fungal inoculation on the growth of Salicornia (Amaranthaceae) under different salinity conditions. Symbiosis 2021, 84, 195–208. [Google Scholar] [CrossRef]
- Tanha, S.R.; Ghasemnezhad, A.; Babaeizad, V.A. Study on the effect of endophyte fungus, Piriformospora indica, on the yield and phytochemical changes of globe artichoke (Cynara scolymus L.) leaves under water stress. Int. J. Adv. Biol. Biom. Res. 2014, 2, 1907–1921. [Google Scholar]
- Zhang, Q.; Gong, M.; Yuan, J.; Hou, Y.; Zhang, H.; Wang, Y.; Hou, X. Dark Septate Endophyte Improves Drought Tolerance in Sorghum. Int. J. Agric. Biol. 2017, 19, 53–60. [Google Scholar] [CrossRef]
- Panke-Buisse, K.; Cheng, L.; Gan, H.; Wickings, K.; Petrovic, M.; Kao-Kniffin, J. Root fungal endophytes and microbial extracellular enzyme activities show patterned responses in tall fescues under drought conditions. Agronomy 2020, 10, 1076. [Google Scholar] [CrossRef]
- Valli, P.P.S.; Muthukumar, T. Dark Septate Root Endophytic Fungus Nectria haematococca Improves Tomato Growth Under Water Limiting Conditions. Indian J. Microbiol. 2018, 58, 489–495. [Google Scholar] [CrossRef] [PubMed]
- Potisek, M.; Likar, M.; Vogel-Mikuš, K.; Arčon, I.; Grdadolnik, J.; Regvar, M. 1,8-dihydroxy naphthalene (DHN)-melanin confers tolerance to cadmium in isolates of melanised dark septate endophytes. Ecotoxicol. Environ. Saf. 2021, 222, 112493. [Google Scholar] [CrossRef] [PubMed]
- Ban, Y.; Xiao, Z.; Wu, C.; Lv, Y.; Meng, F.; Wang, J.; Xu, Z. The positive effects of inoculation using arbuscular mycorrhizal fungi and/or dark septate endophytes on the purification efficiency of CuO-nanoparticles-polluted wastewater in constructed wetland. J. Hazard. Mater. 2021, 416, 126095. [Google Scholar] [CrossRef] [PubMed]
- Shadmani, L.; Jamali, S.; Fatemi, A. Effects of root endophytic fungus, Microdochium bolleyi on cadmium uptake, translocation and tolerance by Hordeum vulgare L. Biologia 2021, 76, 711–719. [Google Scholar] [CrossRef]
- Su, Z.Z.; Dai, M.D.; Zhu, J.N.; Liu, X.H.; Li, L.; Zhu, X.M.; Wang, J.Y.; Yuan, Z.L.; Lin, F.C. Dark septate endophyte Falciphora oryzae-assisted alleviation of cadmium in rice. J. Hazard. Mater. 2021, 419, 126435. [Google Scholar] [CrossRef]
- De Beeck, M.O.; Lievens, B.; Busschaert, P.; Rineau, F.; Smits, M.; Vangronsveld, J.; Colpaert, J.V. Impact of metal pollution on fungal diversity and community structures. Environ. Microbiol. 2014, 17, 2035–2047. [Google Scholar] [CrossRef]
- Utmazian, M.; Schweiger, P.; Sommer, P.; Gorfer, M.; Strauss, J.; Wenzel, W.W. Influence of Cadophora finlandica and other microbial treatments on cadmium and zinc uptake in willows grown on polluted soil. Plant Soil Environ. 2007, 53, 158–166. [Google Scholar] [CrossRef] [Green Version]
- Berthelot, C.; Zegeye, A.; Chalot, M.; Kovács, G.M.; Franken, P.; Leyval, C.; Blaudez, C. Unravelling the Role of Melanin in Cd and Zn Tolerance and Accumulation of Three Dark Septate Endophytic Species. Microorganisms 2020, 8, 537. [Google Scholar] [CrossRef] [Green Version]
- Zhu, L.; Li, T.; Wang, C.; Zhang, X.; Xu, L.; Xu, R.; Zhao, Z. The effects of dark septate endophyte (DSE) inoculation on tomato seedlings under Zn and Cd stress. Environ. Sci. Pollut. Res. 2018, 25, 35232–35241. [Google Scholar] [CrossRef] [PubMed]
- Haruma, T.; Yamaji, K.; Masuya, H. Phialocephala fortinii increases aluminum tolerance in Miscanthus sinensis growing in acidic mine soil. Lett. Appl. Microbiol. 2021, 73, 300–307. [Google Scholar] [CrossRef] [PubMed]
- Shen, M.; Schneiderb, H.; Xua, R.; Caoc, G.; Zhanga, H.; Lia, T.; Zhaoa, Z. Dark septate endophyte enhances maize cadmium (Cd) tolerance by the remodeled host cell walls and the altered Cd subcellular distribution. Environ. Exp. Bot. 2020, 172, 104000. [Google Scholar] [CrossRef]
- Yung, L.; Blaudez, D.; Maurice, N.; Azou-Barré, A.; Sirguey, C. Dark septate endophytes isolated from non-hyperaccumulator plants can increase phytoextraction of Cd and Zn by the hyperaccumulator Noccaea caerulescens. Environ. Sci. Pollut. Res. 2021, 28, 16544–16557. [Google Scholar] [CrossRef] [PubMed]
- Ban, Y.; Xu, Z.; Yang, Y.; Zhang, H.; Chen, H.; Tang, M. Effect of Dark Septate Endophytic Fungus Gaeumannomyces cylindrosporus on Plant Growth, Photosynthesis and Pb Tolerance of Maize (Zea mays L.). Pedosphere 2017, 27, 283–292. [Google Scholar] [CrossRef]
- Berthelot, C.; Blaudez, D.; Beguiristain, T.; Chalot, M.; Leyval, C. Co-inoculation of Lolium perenne with Funneliformis mosseae and the dark septate endophyte Cadophora sp. in a trace element-polluted soil. Mycorrhiza 2018, 28, 301–314. [Google Scholar] [CrossRef] [Green Version]
- Berthelot, C.; Leyval, C.; Foulon, J.; Chalot, M.; Blaudez, D. Plant growth promotion, metabolite production and metal tolerance of dark septate endophytes isolated from metal-polluted poplar phytomanagement sites. FEMS Microbiol. Ecol. 2016, 92, fiw144. [Google Scholar] [CrossRef] [Green Version]
- Suelgaray, F.J.U.; Beltramo, D.M.A.; Lavado, R.S.; Chiocchio, V.M. Dark septate endophytes (DSE): Potential bioremedial promoters of oil derivatives. Int. J. Phytoremediation 2021, 23, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Gehring, C.; Sevanto, S.; Patterson, A.; Ulrich, D.E.M.; Kuske, C.R. Ectomycorrhizal and Dark Septate Fungal Associations of Pinyon Pine Are Differentially Affected by Experimental Drought and Warming. Front. Plant Sci. 2020, 11, 582574. [Google Scholar] [CrossRef] [PubMed]
- Berthelot, C.; Leyva, C.; Chalot, M.; Blaudez, D. Interactions between dark septate endophytes, ectomycorrhizal fungi, and root pathogens in vitro. FEMS Microbiol. Lett. 2019, 366, fnz158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morita, S.; Azuma, M.; Aoba, T.; Satou, H.; Narisawa, K.; Hashiba, T. Induced systemic resistance of Chinese cabbage to bacterial leaf spot and Alternaria leaf spot by the root endophytic fungus, Heteroconium chaetospira. J. Gen. Plant Pathol. 2003, 69, 71–75. [Google Scholar] [CrossRef]
- Bartholdy, B.A.; Berreck, M.; Haselwandter, K. Hydroxamate siderophore synthesis by Phialocephala fortinii, a typical dark septate fungal root endophyte. BioMetals 2001, 14, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Su, Z.; Mao, L.J.; Li, N.; Feng, X.X.; Yuan, Z.L.; Wang, L.W.; Lin, F.C.; Zhang, C.L. Evidence for Biotrophic Lifestyle and Biocontrol Potential of Dark Septate Endophyte Harpophora oryzae to Rice Blast Disease. PLoS ONE 2013, 8, e61332. [Google Scholar] [CrossRef] [Green Version]
- Khastini, R.O.; Ogawara, T.; Sato, Y.; Narisawa, K. Control of Fusarium wilt in melon by the fungal endophyte, Cadophora sp. Eur. J. Plant Pathol. 2014, 139, 339–348. [Google Scholar] [CrossRef]
- Yakti, W.; Kovacs, G.M.; Franken, P. Differential interaction of the dark septate endophyte Cadophora sp. and fungal pathogens in vitro and in planta. FEMS Microbiol. Ecol. 2019, 95, 164. [Google Scholar] [CrossRef]
- Harsonowati, W.; Marian, M.; Surono; Narisawa, K. The Effectiveness of a Dark Septate Endophytic Fungus, Cladophialophora chaetospira SK51, to Mitigate Strawberry Fusarium Wilt Disease and With Growth Promotion Activities. Front. Microbiol. 2020, 11, 585. [Google Scholar] [CrossRef] [Green Version]
- Chu, H.; Wang, C.; Li, Z.; Wang, H.; Xiao, Y.; Chen, J.; Tang, M. The dark septate endophytes and ectomycorrhizal fungi effect on Pinus tabulaeformis Carr. Seedling growth and their potential effects to pine wilt disease resistance. Forests 2019, 10, 140. [Google Scholar] [CrossRef] [Green Version]
- Chu, H.; Wang, H.; Zhang, Y.; Li, Z.; Wang, C.; Dai, D.; Tang, M. Inoculation With Ectomycorrhizal Fungi and Dark Septate Endophytes Contributes to the Resistance of Pinus spp. to Pine Wilt Disease. Front. Microbiol. 2021, 12, 687304. [Google Scholar] [CrossRef]
- Terhonen, E.; Kerio, S.; Sun, H.; Asiegbu, F.O. Endophytic fungi of Norway spruce roots in boreal pristine mire, drained peatland and mineral soil and their inhibitory effect on Heterobasidion parviporum in vitro. Fungal Ecol. 2014, 9, 17–26. [Google Scholar] [CrossRef]
- Terhonen, E.; Sipari, N.; Asiegbu, F.O. Inhibition of phytopathogens by fungal root endophytes of Norway spruce. Biol. Cont. 2016, 99, 53–63. [Google Scholar] [CrossRef]
- Martínez-Arias, C.; Plata, J.S.; Moncalvillo, S.O.; Gil, L.; Calcerrada, J.R.; Martín, J.A. Endophyte inoculation enhances Ulmus minor resistance to Dutch elm disease. Fungal Ecol. 2021, 50, 101024. [Google Scholar] [CrossRef]
- Narisawa, K.; Tokumasu, S.; Hashiba, T. Suppres-sion of clubroot formation in Chinese cabbage by the rootendophytic fungus, Heteroconium chaetospira. Plant Pathol. 1998, 47, 210–216. [Google Scholar] [CrossRef]
- Ohtaka, N.; Narisawa, K. Molecular characterization and endophytic nature of the root-associated fungus Meliniomyces variabilis (LtVB3). J. Gen. Plant Pathol. 2008, 74, 24–31. [Google Scholar] [CrossRef]
- Surono; Narisawa, K. The inhibitory role of dark septate endophytic fungus Phialocephala fortinii against Fusarium disease on the Asparagus officinalis growth in organic source conditions. Biol. Cont. 2018, 120, 159–167. [Google Scholar] [CrossRef]
- Khastini, R.O.; Ohta, H.; Narisawa, K. The Role of a Dark Septate Endophytic Fungus, Veronaeopsis simplex Y34, in Fusarium Disease Suppression in Chinese Cabbage. J. Microbiol. 2012, 50, 618–624. [Google Scholar] [CrossRef] [PubMed]
- Tellenbach, C.; Sieber, T.N. Do colonization by dark septate endophytes and elevated temperature affect pathogenicity of oomycetes? FEMS Microbiol. Ecol. 2012, 82, 157–168. [Google Scholar] [CrossRef] [Green Version]
- Reininger, V.; Grünig, C.R.; Sieber, T.N. Host species and strain combination determine growth reduction of spruce and birch seedlings colonized by root-associated dark septate endophytes. Environ. Microbiol. 2012, 14, 1064–1076. [Google Scholar] [CrossRef]
- Hidayat, I. Dark Septate Endophytes and Their Role in Enhancing Plant Resistance to Abiotic and Biotic Stresses. In Plant Growth Promoting Rhizobacteria for Sustainable Stress Management; Sayyed, R., Arora, N., Reddy, M., Eds.; Springer: Singapore, 2019; Volume 12, pp. 35–63. [Google Scholar]
- Berek-Nagy, P.J.; Tóth, G.; Bősze, S.; Horváth, L.B.; Darcsi, A.; Csikos, S.; Knapp, G.; Kovács, G.M.; Boldizsár, I. The grass root endophytic fungus Flavomyces fulophazii: An abundant source of tetramic acid and chlorinated azaphilone derivative. Phytochemistry 2021, 190, 112851. [Google Scholar] [CrossRef]
- Lin, L.C.; Tan, Y.L.; Lin, W.R.; Ku, K.L.; Ho, S.T. The effect of dark septate endophytic fungi on Mahonia oiwakensis. Plants 2021, 10, 1723. [Google Scholar] [CrossRef] [PubMed]
- Spagnoletti, F.N.; Chiocchio, V.M. Tolerance of dark septate endophytes (DSE) to agrochemicals in vitro. Rev. Argent. Microbiol. 2020, 52, 43–49. [Google Scholar] [PubMed]



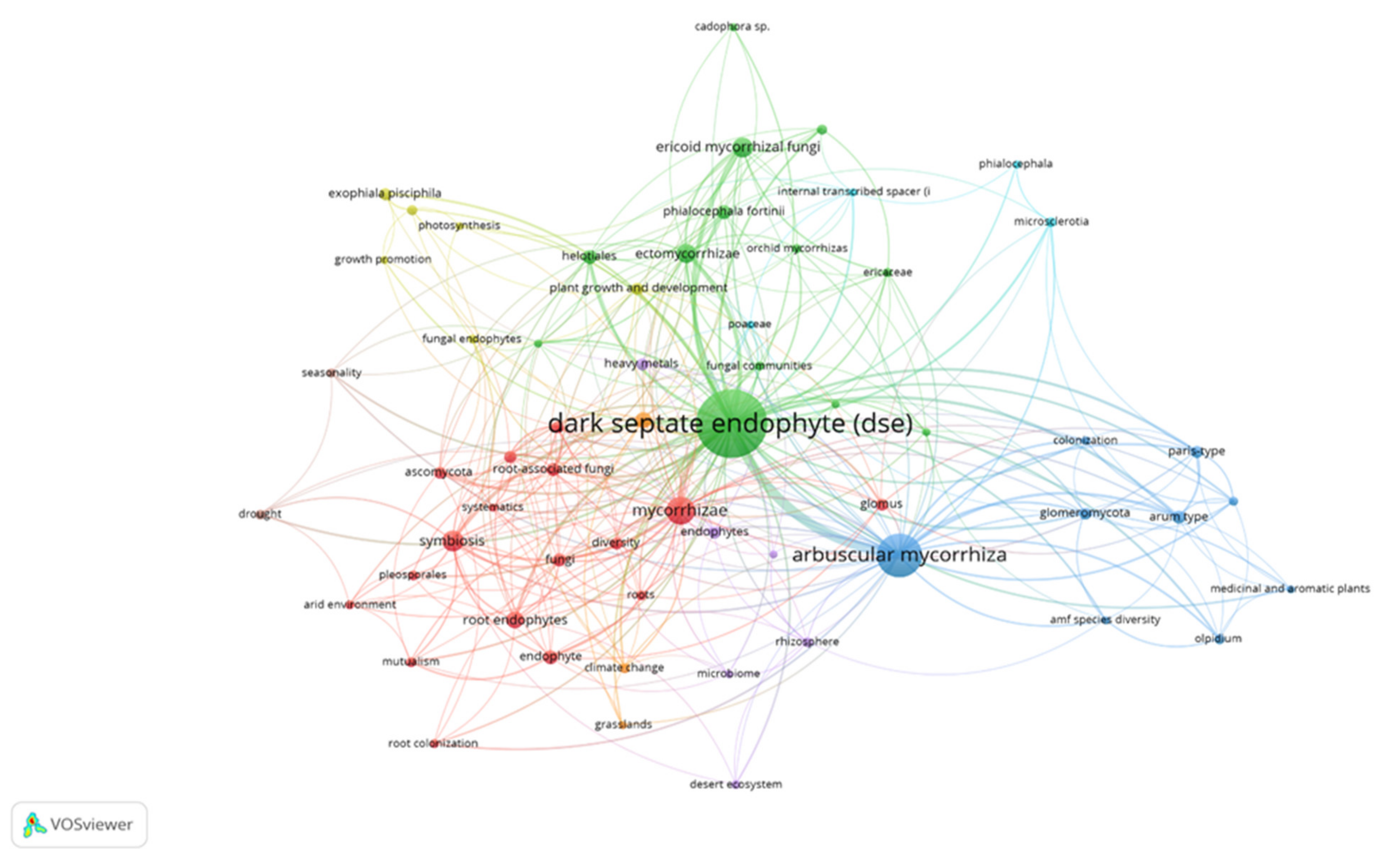{kind=link}
{kind=link}
| DSE | Phytopathogen | Host | References |
|---|---|---|---|
| Cadophora sp. | Fusarium oxysporum f. sp. meloni | Cucumis melo | [131] |
| Verticillium dahliae | In vitro/tomato | [132] | |
| Rhizoctonia solani | In vitro/tomato | [132] | |
| Pythium aphanidermatum | In vitro/tomato | [132] | |
| Heterobasidion annosum | In vitro | [127] | |
| Cladophialophora chaetospira | Fusarium oxysporum | Brassica pekinensis | [133] |
| Cladosporium cladosporioides | Bursaphelenchus xylophilus | Pinus tabulaeformis | [134,135] |
| Fragaria vesca | [133] | ||
| Cryptosporiopsis ericae | Heterobasidionparviporum | In vitro | [136] |
| Cryptosporiopsis sp. | Phytophtora pini, Heterobasidion parviporum | In vitro | [137] |
| Botrytis cinerea | |||
| Exophiala pisciphila | Fusarium oxysporum | Brassica pekinensis | [133] |
| Fusarium oxysporum f. sp. fragariae | Fragaria vesca | [134] | |
| Tylenchulus semipenetrans | Screening/in vitro | [44] | |
| Exophiala salmonis | Bursaphelenchus xylophilus | Pinus tabulaeformis | [134] |
| Exophiala sp. | Fusarium oxysporum | Brassica pekinensis | [133] |
| Fusarium oxysporum f. sp. fragariae | Fragaria vesca | [133] | |
| Heterodera schachti | Beta vulgaris | [43] | |
| Ophiostoma ulmi | Ulmus | [138] | |
| Gaeumannomyces cylindrosporus | Bursaphelenchus xylophilus | Pinus spp. | [135] |
| Bursaphelenchus xylophilus | Pinus tabulaeformis | [134] | |
| Harpophora oryzae | Magnaporthe oryzae | Rice | [130] |
| Heteroconium chaetospira | Verticillium longisporum, Plasmodiophora brassicae | Brassica pekinensis | [80,139] |
| Pseudomonas syringae pv. Macricola, A. brassicae | [128] | ||
| Leptodontidium orchidicola | Verticillium dahliae | Tomato | [62] |
| Leptodontidium sp | Pythium intermedium, Phytophthora citricola, H. annosum | In vitro | [127] |
| Meliniomyces variabilis | Verticillium longisporum | Brassica pekinensis | [80,140] |
| Paraphoma chrysanthemicola | Bursaphelenchus xylophilus | Pinus spp. | [135] |
| Pinus tabulaeformis | [134] | ||
| Phialocephala bamuru | Rhizoctonia solani | Pinus silvestris | [79] |
| Phialocephala sphareoides | Heterobasidionparviporum | In vitro/Picea abies | [134] |
| Phialocephala europaea | Phytophthora citrícola | In vitro | [127] |
| Phialocephala fortinii | Verticillium longisporum | Brassica pekinensis | [80] |
| Fusarium oxysporum f. sp. asparagi | Asparagus officinalis | [141] | |
| PAC | Heterobasidionparviporum | In vitro | [136] |
| Phialophora mustea | Phytophtora citricola, Pythium intermedium H. annosum | In vitro | [127] |
| Bursaphelenchus xylophilus | Pinus tabulaeformis | [134] | |
| Phialocephala subalpina | Phytophthora plurivora | In vitro | [132] |
| Elongisporangium undulatum | In vitro | [132] | |
| Veronaeopsis simplex | Fusarium oxysporum | In vitro | [142] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santos, M.; Cesanelli, I.; Diánez, F.; Sánchez-Montesinos, B.; Moreno-Gavíra, A. Advances in the Role of Dark Septate Endophytes in the Plant Resistance to Abiotic and Biotic Stresses. J. Fungi 2021, 7, 939. https://doi.org/10.3390/jof7110939
Santos M, Cesanelli I, Diánez F, Sánchez-Montesinos B, Moreno-Gavíra A. Advances in the Role of Dark Septate Endophytes in the Plant Resistance to Abiotic and Biotic Stresses. Journal of Fungi. 2021; 7(11):939. https://doi.org/10.3390/jof7110939
Chicago/Turabian StyleSantos, Mila, Ignacio Cesanelli, Fernando Diánez, Brenda Sánchez-Montesinos, and Alejandro Moreno-Gavíra. 2021. "Advances in the Role of Dark Septate Endophytes in the Plant Resistance to Abiotic and Biotic Stresses" Journal of Fungi 7, no. 11: 939. https://doi.org/10.3390/jof7110939