
Shelterbelts Planted on Cultivated Fields Are Not Solutions for the Recovery of Former Forest-Related Herbaceous Vegetation


1
Institute of Advanced Studies, 9730 Kőszeg, Hungary
2
Institute of Environmental and Earth Sciences, University of Sopron, 9400 Sopron, Hungary
3
Institute of Botany, Natural Resources and Life Sciences, 1180 Vienna, Austria
*
Author to whom correspondence should be addressed.
Land 2021, 10(9), 930; https://doi.org/10.3390/land10090930
Submission received: 13 August 2021
/
Revised: 31 August 2021
/
Accepted: 1 September 2021
/
Published: 3 September 2021
(This article belongs to the Special Issue Land Degradation and Sustainable Land Management)
Abstract
:Establishing shelterbelts for field protection is one of the rediscovered agroforestry practices in Europe and Hungary. Several studies have focused on the effects of these plantations on agricultural production. Prior scholarship reveals that shelterbelts enhance the diversity of bird and insect communities but generally fail to consider herbaceous cover. Our study aimed to describe the herbaceous vegetation in shelterbelts of different origins, tree species composition, and land management. We investigated surveys in four agricultural landscapes of North West Hungary, where the intensity of the landscape transformation is different. The diversity and species composition of the herbaceous vegetation were analyzed, including plant sociology and forest affinity. Our results highlight the importance of landscape history in herbaceous flora. Shelterbelts planted on cultivated without an immediate connection to former woody vegetation soil are not appropriate for the appearance of forest-related herbaceous species, regardless of tree species composition or the extent of the shelterbelt. On the contrary, the remnants of former woody vegetation are refuges for those herbaceous species that are very slow at colonizing new plantations. These findings expose that protecting existing woody areas is an essential task of agricultural land management.
1. Introduction
Human activities in intensive land use systems often lead to soil degradation: erosion, unbalanced nutrient conditions, acidification, and the decline of soil functional diversity [1]. Living plant organisms can replace or increase the efficiency of technological solutions and at the same time favorably affect the quality of the environment, developing biologically active areas [2,3,4,5,6]. The widespread application of shelterbelts in Central Europe after the Second World War aimed to improve agricultural landscapes and production [7,8]. Later on, their biodiversity-enhancing effect was also regarded, mainly in terms of tree species diversity [9] and bird and insect communities [10,11,12,13]. Shelterbelts are ecological systems that can contain significant biodiversity. Several species within them are natural predators of pests, which can have a beneficial impact on pest control for agriculture [8,14,15]. Furthermore, by reducing wind speed, shelterbelts can significantly prevent the spread of some wind-carried pests and aphid-transmitted viruses [16]. By providing large quantities of pollen and nectar, shelterbelts positively affect the diversity of the pollinator fauna [17] and can play a significant role in domestic honey production [18].
The abundance and diversity of herbaceous plants within the shelterbelts can strongly influence the composition and richness of the food chain based on it and thus the overall biodiversity of the agroforestry system. Additionally, shelterbelts as extended linear structures have a corridor effect, improving connectivity between woody patches, increasing the area of tree stands, and offering edge habitats in the agricultural landscape [19]. These corridors increase the number of native plant species at single sites so that they are a crucial instrument for preserving biodiversity [20]. However, our knowledge of the herbaceous vegetation of shelterbelts is comparatively poor. Carlier–Moran [21] examined the understory layer of the hedgerows in Western Europe, and they found very few generalist species. Szarvas [8] also reflected that the herbaceous flora consists of the same widespread and disturbance-tolerant species in shelterbelts. On the other hand, the former forest or grassy vegetation can significantly impact the herbaceous layer of a tree plantation [22,23]. Research on soil biota showed that former agricultural management has a long-term effect on the afforested areas. However, in native tree plantations, positive processes occurred in the soil twenty years after afforestation [24].
The current study aimed to describe the herbaceous vegetation in shelterbelts of different origins in relation to tree species composition and land management. We hypothesized that under a tree plantation composed of native species, a more diverse herbaceous vegetation containing several forest-related species could develop. At the same time, the understory layer of non-native shelterbelts was supposed to be species-poor and dominated by the most abundant ruderal weeds. The results showed that the landscape history is a more significant predictor for herbaceous vegetation than the tree species composition of the shelterbelt.
2. Materials and Methods
2.1. Study Sites
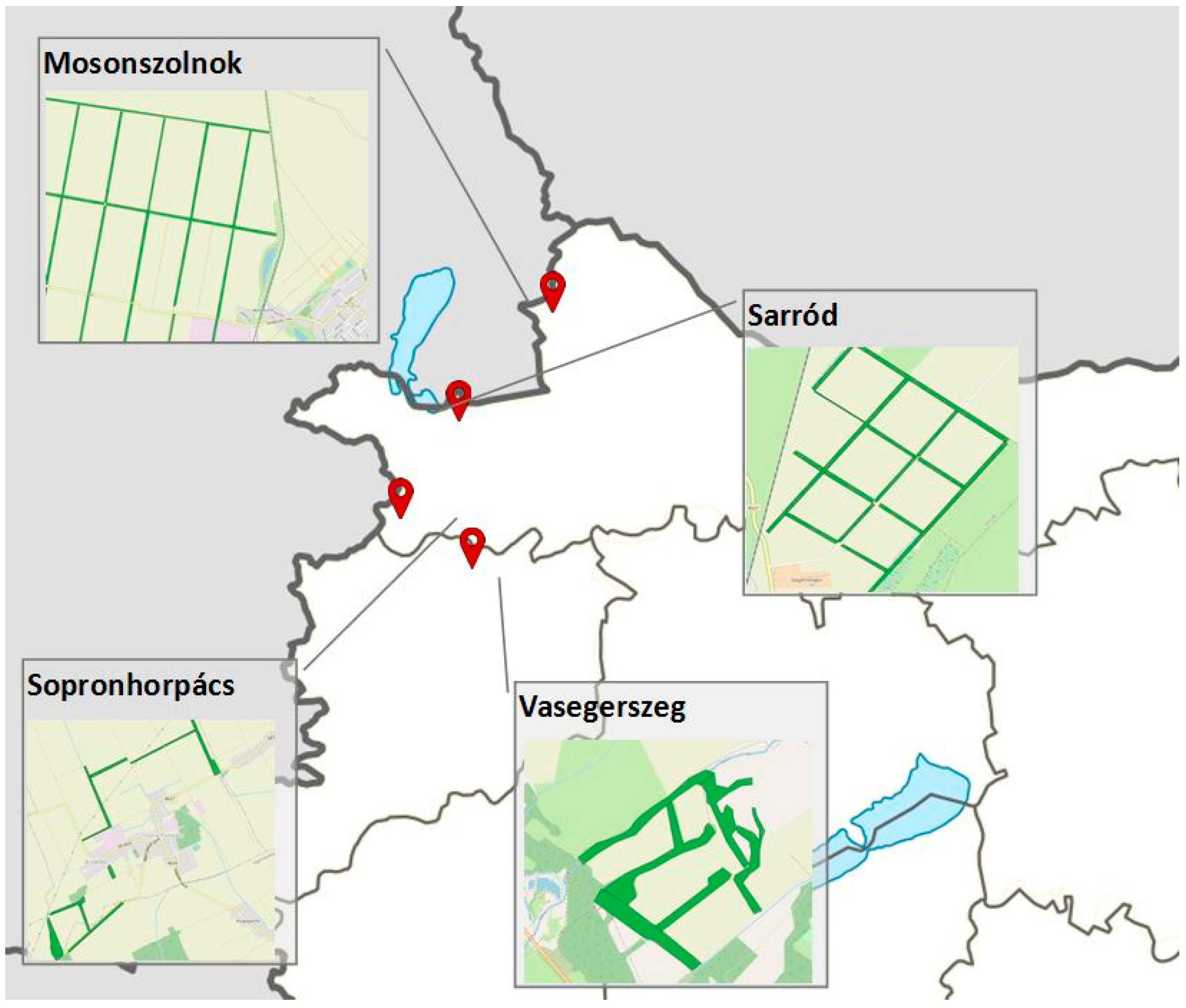
The surveys took place in North West Hungary in four agricultural landscapes segmented by shelterbelts of different tree species composition and origin, as shown in Figure 1. The GPS coordinates of the examined plots can be found in Appendix A.
The two southern study areas belong to the West-Hungarian Borderland, while the two northern ones to the Lesser Plain. An important common feature of the sample areas is the significant climatic drought caused by the frequent descending airflow (foehn) from the Eastern Alps, among others [25]. The climate of Mosonszolnok is moderately warm and dry. Vasegerszeg and Sopronhorpács belong to the moderately hot and moderately dry climate. Sarród represents the transition between these two climate types [25,26].
The bedrock for the soils are calcareous loess sediments in Mosonszolnok and Sarród and loamy alluvial sediments in Sopronhorpács and Vasegerszeg. The different soil physical characteristics and groundwater depths cause the divergences of the features of the study areas. The soil type in Mosonszolnok and Sarród is Chernic Phaeozem, while in Sopronhorpács and Vasegerszeg it is Mollic Gleysol.
Zonal oak forests are restricted to a few remnants in the higher elevation areas of the region [27], while they did not remain at all in the vicinity of the study areas. Secondary grassland vegetation and crop fields dominate the landscapes here.
In Mosonszolnok, agricultural cultivation determines the current appearance of the landscape; the few semi-natural habitats are highly fragmented. On the monocrop fields, mainly cereals are grown. Forest belts established for protecting the crop against wind damage and soil erosion separate the intensively managed fields. Three of the examined shelterbelts (MS1–MS3) were planted in agricultural areas, while the fourth (MS4) can be followed back to a chariot road on a historical map from the XVIII century (obtained at Mapire.eu). Two shelterbelts (MS1 and MS2) stands of black locust (Robinia pseudoacacia) and green ash (Fraxinus pennsylvanica) are quite similar in appearance; both of them are 15–18 m wide planted forest stands established in the 1950s. The two other belts (MS3 and MS4) are dominated by field maple (Acer campestre); their width is 20 m and 50 m, respectively.
Sarród study area is situated in the plain of the Fertő basin. At present, semi-natural forests, which could function as propagule sources, cannot be found in the vicinity of the study sites, and only planted shelterbelts represent woody vegetation. Most of the grassland habitats are also secondary, developed from former arable field fallows [28]. The examined 20 m wide Ulmus minor–Fraxinus pennsylvanica–Acer campestre shelterbelts are about 40–50 years old.
Sopronhorpács shelterbelt system belongs to the Répce plain, where the former valleys of the ancient river can be observed. Oak-hornbeam forest stands prove propagule sources, but the amount of planted pine and black locust forests is also high [28].
One of the examined shelterbelts (SH2) is an experimental plantation established in the 1950s for field protection on agricultural land [29]. SH3 was an already existing forest stand at that time. On the historical map of the XVIII century obtained from Mapire.eu, the area of SH1 is a chariot road. At present, a dirt road runs on the trail of it. All shelterbelts consist of mainly native tree species: Quercus robur, Q. cerris, and Acer campestre.
Vasegerszeg study area lies in the valley of the river Répce. Most of the former riparian Fraxino pannonicae-Ulmetum forests disappeared, but some relict stands still exist [28]. The shelterbelts and forest stands are such remnant forests, but the herbaceous vegetation is significantly transformed to a less characteristic condition in most of them.
2.2. Sampling
We designated three sampling quadrats with an extension of 25 m2 in each examined shelterbelts and recorded herbaceous plant species and their cover (%). The sampling was repeated in 3 different vegetation periods in 2019: early spring, summer, and autumn. In the case of each species, we considered the highest found value during the vegetation period and calculated the mean of the cover values of species calculated for each shelterbelt. Soil samples were taken from every plot to measure the following parameters: soil pH, soil organic matter content (SOM), available nitrogen, phosphorus, potassium content, and particle size distribution. The nomenclature of species follows the Hungarian Flora Database [30].
2.3. Diversity and Species Composition Analysis
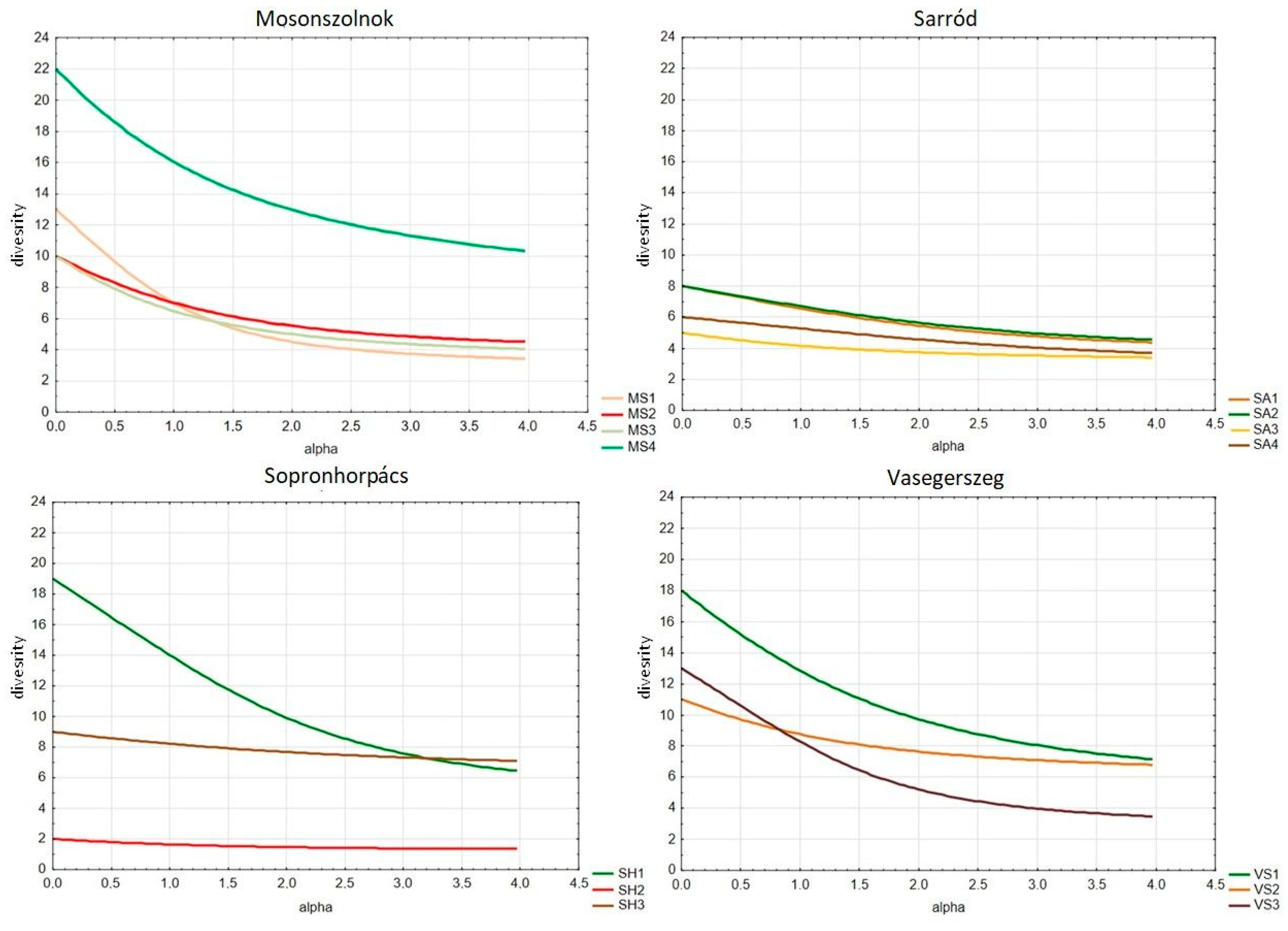
Rényi’s diversity profiles were used to compare the herb layers of the different habitats, calculated with PAST software. PAST uses the exponential of the so-called Renyi index, including the number of species, Shannon diversity, quadratic diversity, and Berger–Parker diversity. The value of the index depends upon the parameter “alpha”. For alpha = 0, this function gives the total species number. Alpha = 1 (in the limit) gives an index proportional to the Shannon index, while alpha = 2 gives an index that behaves like the Simpson index. With this method, the studied communities can be ranked in a partial order. A community is more diverse than another when its diversity profile is above or equal to the other community’s diversity profile [31].
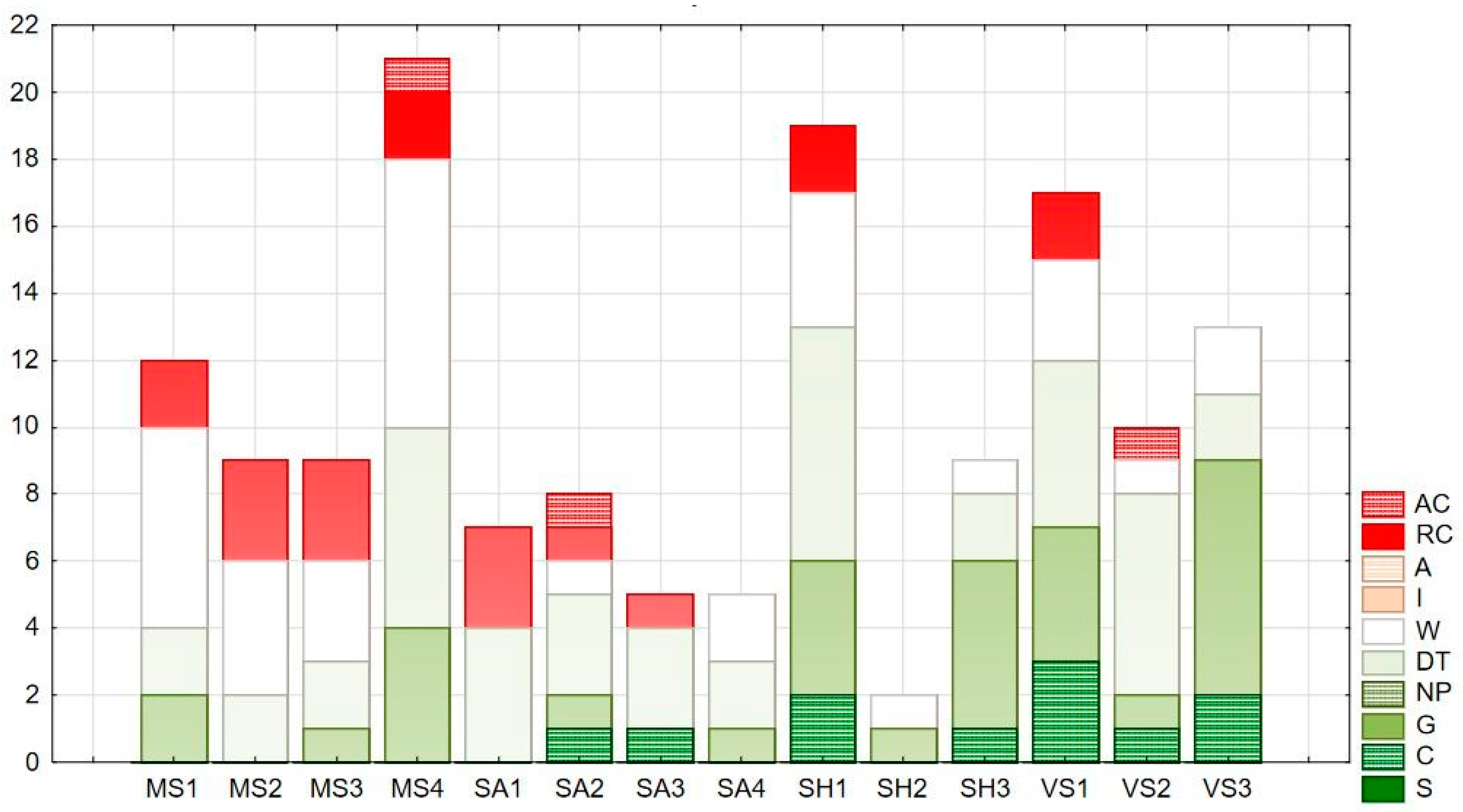
The naturalness values of the plant species are based on the social behavior type categories, derived from the CSR plant functional system [32], adapted to the Pannonian flora by Borhidi [33]. The role the species play in the communities and the way and naturalness with which they are linked to the habitat determine the SBT category of the species. The number and proportion of categories represented in a habitat prove information about the stability, the level of disturbance, or deviation from the natural state of the community. Borhidi’s SBT categories were obtained from the Hungarian Flora Database [30]. Stacked column charts display the proportion of species belonging to the different categories in a habitat, where the total species number of the examined habitat and the distribution of the species numbers by the SBT categories simultaneously can be examined. The abbreviations and naturalness values of SBT categories are introduced in Table 1.
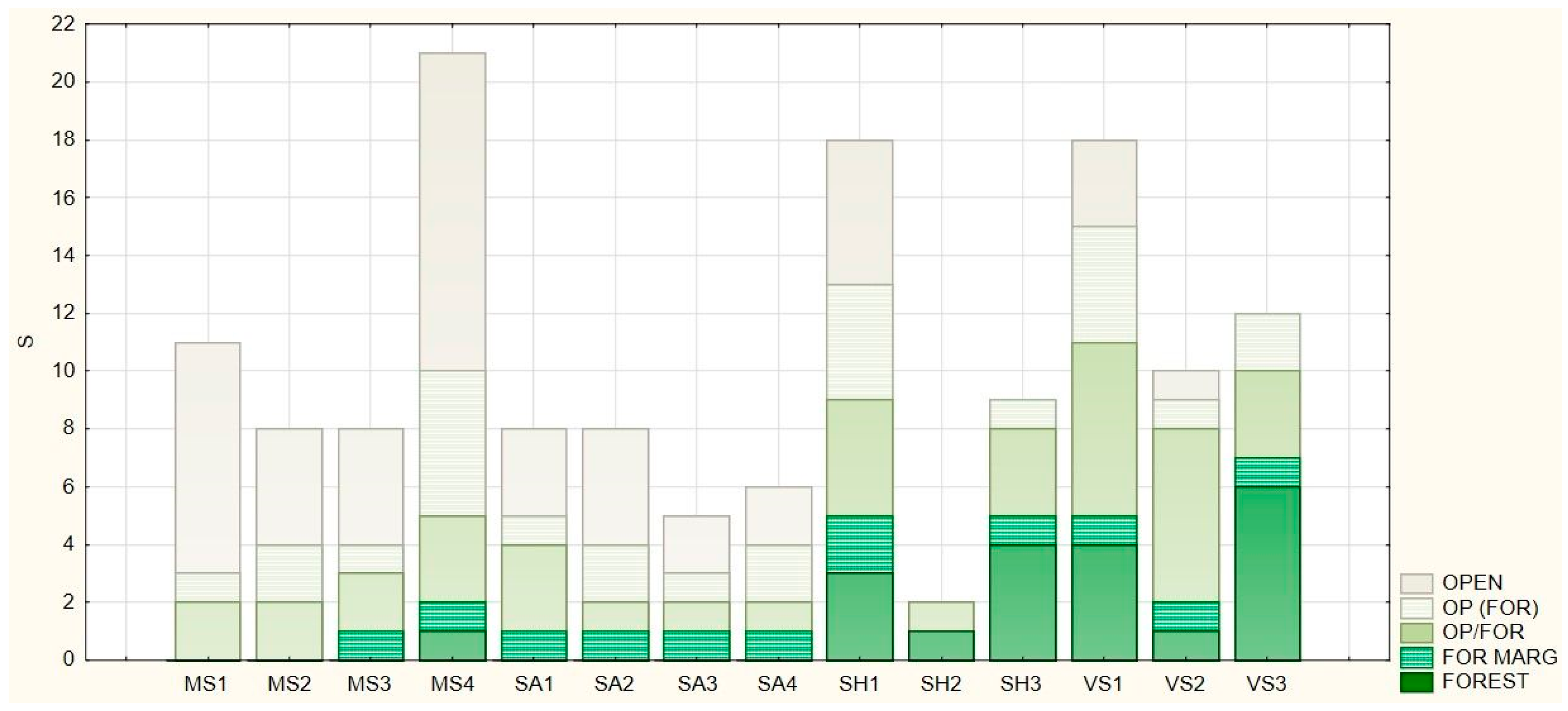
Forest affinity categories show the relation of the herbaceous species to forest stands. The data were obtained from the German plant database Floraweb (www.floraweb.de, accessed on 31 January 2021), which ranks species into five categories (Table 2).
In the database, the forest affinity of species is given for three different geographical zones: the Alpine region, the low mountain range, and northern German lowlands. We used the data from the first one for our analysis considering Pannonian basin to be in general closer to the alpine region than to the other two zones.
Redundancy analysis was performed with Canoco 5 to explore the relationships between herbaceous species composition and environmental descriptors. Besides soil parameters, the cover of different tree species and the width and the origin of the shelterbelt were taken into account. The species abundance matrix was transformed with the Hellinger method. Hellinger transformation is recommended as a basis for ordination as it allows one to explore the relationships between the species and the explanatory variables [34].
3. Results
3.1. Tree and Shrub Species Composition
The tree species composition is different in the examined shelterbelts of the four landscapes, and, in the case of Mosonszolnok (MS) and Sopronhorpács (SH), also within the tree stands. The reason for this diversity is conscious planning on the one hand and the landscape features, such as forest remnants, on the other hand. Table 3 shows the tree and shrub species and their cover values at the study sites.
Some common species dominate the shrub layer in most shelterbelts, for example, Ligustrum vulgare, Prunus spinosa, and Sambucus nigra (Table 4). Exotic species, such as Eleagnus angustifolia or Maclura pomifera, appear in the non-native shelterbelts of Mosonszolnok (MS1 and MS2), and the edge of the native stands in Sarród (SA1–SA4).
3.2. Herbaceous Diversity
According to the understory layer, 69 herbaceous species were detected in the 14 examined plots. Fifty-three of them were unique to one study area: 23 to Mosonszolnok, 7 to Sarród, 12 to Sopronhorpács, and 11 to Vasegerszeg. Only three species—Urtica dioica, Geum urbanum, and Galium aparine—appeared in all study areas. The species list and cover values of herbaceous plants can be found in Appendix A.
Significant differences can be observed among the diversity profiles of the examined shelterbelts, as shown in Figure 2. In Mosonszolnok, the herbaceous diversity of the three shelterbelts planted on an agricultural field (MS1, MS2, and MS3) is remarkably lower than that of one from the trail of the former chariot road (MS4). In Sarród, all of the four shelterbelts (SA1–SA4) stand on a former cultivated field, and all of them are poor in herbaceous species. In Sopronhorpács, the herbaceous diversity of the trial plantation (SH2) is extremely low due to the almost 100% cover of Hedera helix. According to herbaceous diversity, all of the shelterbelts of Vasegerszeg (VS1–VS3) represent intermediate values among the study sites.
3.3. Naturalness Value and Forest Affinity of the Herbaceous Vegetation
The distribution of species by social behavior type categories in Figure 3 shows that in most study sites, the majority of the herbaceous species are weeds (W) and disturbance tolerants (DT). In Mosonszolnok, the most frequent species of these categories are Arrhetatherum elatius, Ballota nigra, Galium aparine, and Lamium purpureum. In Sarród, this group is represented by Aegilops cylindrica and Geum urbanum, while in Sopronhorpács, Galium aparine and Urtica dioica are the most frequent ones. In Vasegerszeg, the proportion of these categories is the lowest; Geum urbanum and Lamium maculatum represent the disturbance-tolerant category.
The only species belonging to Competitors (C) in Sarród was Festuca rupicola, which might indicate the remnants of grassland vegetation. In Vasegerszeg, Ranunculus ficaria, Geranium sanguineum, and Corydalis cava represent this category. In the former chariot road of Sopronhorpács Vinca minor appears with low cover, while in Mosonszolnok, no natural competitors could be detected. Specialist species (S) are absent in all test sites. The proportion of generalists (G) is higher in MS4 and SH1 shelterbelts, both situated on the trail of the former chariot roads. This category is represented by Arum orientale, Viola odorata, and Polygonatum species, and in Vasegerszeg additionally by Anemone ranunculoides and Gagea lutea. The only adventive competitor (AC) present in Mosonszolnok and Sarród is Erigeron annuus, while the ruderal competitors (RC) Bromus sterilis and Convolvulus arvensis appear in most of the shelterbelts.
The proportion of forest-related herbaceous species also differed significantly between the examined shelterbelts, as shown in Figure 4. The shelterbelts planted on agricultural land do not contain any obligate forest species (FOR). This was true for Mosonszolnok (MS1–MS3), Sarród (SA1–SA4), and Soprohnorpács (SH2). Only a few forest-related species appear in the trail of the former chariot roads: Anemone ranunculoides in Mosonszolnok (MS4) and Brachypodium sylvaticum in Sopronhorpács (SH1). In the shelterbelts, which were former forest stands (SH3, VS1–VS3), more forest-related herbaceous species appear, among others Asarum europaeum, Convallaria majalis, and Corydalis cava.
3.4. Redundancy Analysis
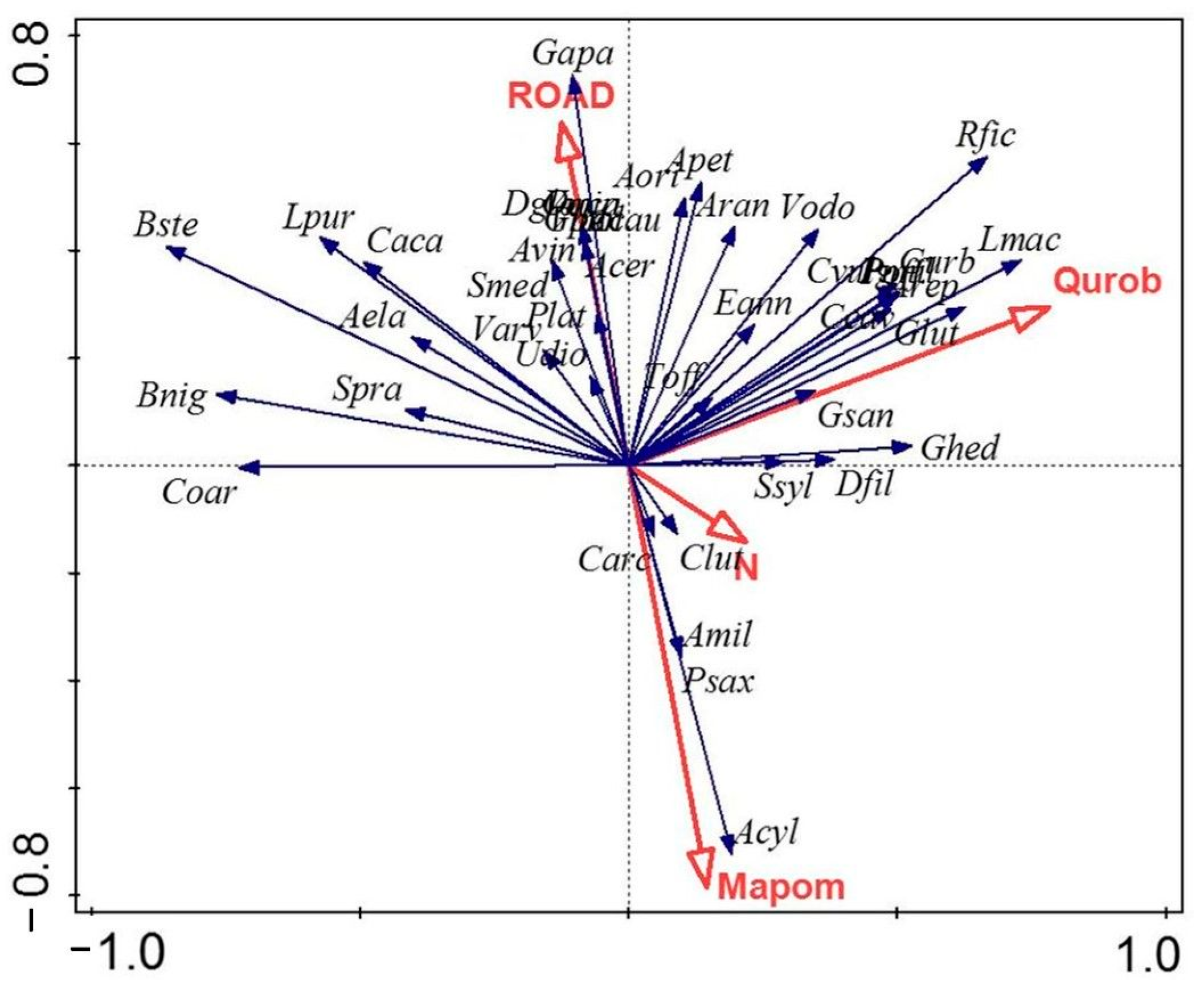
Twenty-five factors were taken into account in the redundancy analysis, including the cover of the different tree species, the measured soil parameters (pH; soil organic matter content (SOM); available nitrogen, phosphorus, and potassium content; and particle size distribution), and the extension and the origin of the shelterbelts. The forward selection showed that the most important factors explaining the species composition of the herbaceous level of shelterbelts are the dominant trees, especially Quercus robur (Qurob) and Maclura pomifera (Mapom). The origin of the shelterbelt was also significant (p < 0.05); formerly, it was a chariot road (ROAD). Soil and other parameters were insignificant; their effect is summarized in factor N, as shown in Figure 5. Th explanatory variables account for 54.64% of the variation.
The high significance of Quercus robur and Maclura pomifera indicates the impact of the origin of the shelterbelt as common oak is characteristic of forest remnants, while Osage orange was commonly planted in shelterbelts of agricultural areas.
Woodland species, such as Gagea lutea and Corydalis cava, correlate positively with the remnant forests dominated by Quercus robur, whose factor explains 17.5% of the variation. Some species that can be found both in open and woody habitats, such as Lamium maculatum, Ranunculus ficaria, and the forest margin species Geum urbanum, are also characteristic of these shelterbelts.
Species of open habitats, like Aegilops cylindrica, Pimpinella saxifraga, and Achillea millefolium, correlate positively with Maclura pomifera, which represents recently established shelterbelts and explains 15.4% of the variation.
Shelterbelts, which can be followed back to former chariot roads in historical maps, form the third group. Many weeds and disturbance-tolerant species of open and woody habitats correlate positively with factor ROAD (13.2% variation explained), as among others Dactylis glomerata, Galium aparine, Anthriscus cerefolium, Onopordum acanthium, or Allium vineale.
All of the other factors, such as soil parameters, are not significant and explain only 8.6%.
4. Discussion
The understorey of the hedgerows in Western Europe was also considered species-poor [21]. Szarvas [8] found that the differences in the vegetation of tree rows and shelterbelts are given by tree and shrub species, not herbaceous plants. In most shelterbelts, the herbaceous flora consists of the same widespread and disturbance-tolerant species. However, the herbaceous community of hedgerows can vary depending on the hedge’s origin and history. Besides that, the land use characteristics of the adjacent habitats also affect the understorey layer. In the case of old hedgerows, a direct connection to pre-existing woodland vegetation can be presumed. These boundaries often retain an intact ancient woodland flora rich in species, characteristic of long-established woodland [35]. Wilson [22] also supports this idea, while species probably derived from ancient landscape features, such as Anemone nemorosa, Primula vulgaris, and Lamiastrum (Lamium) galeobdolon, are typically very slow to colonize new woodlands. In hedgerows, without woodland history, the herbaceous community includes generalist flora of forests, for example, Hedera helix, Geranium robertianum, Glechoma hederacea, Galium aparine, and grasslands, as well as Arrhenatherum elatius, Dactylis glomerata, and Anthriscus sylvestris. In tree stands planted on a previous meadow, grassy species appear in the herb layer [23]. In the most unfavorable case, the herbaceous vegetation of the hedgerow is affected by herbicides and fertilizers, where more competitive ruderals—Elymus repens, Poa trivialis, and Bromus sterilis—cover the soil. Some species such as Glechoma hederacea and Viola odorata are able to colonize disturbed forest areas and recent forest stands [36,37]. Epizoochorous species Geum urbanum and Urtica dioica are also considered typical quick colonizers [37,38].
In Vasegerszeg, all of the shelterbelts and the control forest are remnants of the former woodland that may cause the balanced diversity of the examined habitats. The majority of the forest-related species were found in the local species pool of the Répce floodplain, and many of them are considered indicators of ancient woodlands [35].
The reason for the lack of forest-related species in many shelterbelts is complex. These habitats represent various ecosystems along an intense usage and light gradient. Due to their comparatively small size and the edge effect in the whole area of the shelterbelt, species of open lands are likely to colonize them [39]. On the other hand, many woodland species are unable to colonize isolated woods [36] and recently established forests [38]. This can be caused by the dispersal characteristics and low competitive ability of woodland species. Ancient woodland indicators are mainly shade-tolerant, hemicryptophyte, and geophyte species and do not have persistent seed banks [37]. These species disappear from the soil seed bank in case of agricultural land use within a few decades [40]. The proportion of shade-tolerant species is lower in recent forests, even if they are adjacent to ancient woodlands [41]. The absence or presence of species with high forest affinity in the herbaceous layer of shelterbelts is an important marker of historical development and connectivity. Similar to our results, Van Den Berge et al. [42] also found that the forest-related herbaceous community is not related to the size of the woody area. On the contrary, the origin of the tree stand is a significant factor determining the number of forest species. In hedgerows, the structure is a less relevant agent in herbaceous diversity than the connectivity to woodlands [43]. The understory layer of young hedgerows plantations is affected significantly by the farming type; the naturalness of the herbaceous community is higher in hedgerows adjacent to organic farming compared to conventional areas [44].
5. Conclusions
The herbaceous layer of the examined shelterbelts showed a certain pattern according to the number of species and the naturalness value. This pattern is linked to the origin of the shelterbelt than tree species composition. Shelterbelts planted in agricultural fields are very important for many species, but they are not suitable for the appearance of forest-related herbaceous vegetation. The herbaceous flora of these shelterbelts is species-poor and contains mostly disturbance-tolerant (ruderal) plant species. Despite the allelopathic effects of black locust, the herbaceous layer is similar to the shelterbelts built by native trees.
Those tree stands, which have an immediate relation to a former woodland, do not contain a high number of species, but the naturalness value of these species is relatively high as they preserve several species of the former forest vegetation.
Shelterbelts standing on the trail of former chariot roads cannot be ordered to one of the abovementioned groups. Both of them possess a relatively diverse herb layer with a few forest-related species and many disturbance tolerants and weeds.
The reasons for the low species diversity of herbaceous vegetation in shelterbelts are complex. On the one hand, dispersal limitation and the negative effects of fragmentation and the agricultural surrounding can be mentioned.
Nevertheless, the protection of existing shelterbelts, particularly ancient forest fragments, is of high importance to preserve at least a small portion of the herbaceous vegetation of the former woodlands. These woody islands may significantly increase the biodiversity of the intensively managed agricultural fields. Once the area of the recent shelterbelt was a part of an arable field, the forest-related herbaceous vegetation cannot appear for a long time.
Author Contributions
N.S.: conceptualization, investigation, methodology, writing, review and editing. I.B.: conceptualization and investigation. A.V.: Supervision. L.R.: methodology, supervision, review and editing. All authors have read and agreed to the published version of the manuscript.
Funding
This project received funding from the European Union’s Horizon 2020 Research and Innovation Programme, under Grant Agreement No. 818346 (Sino-EU Soil Observatory for Intelligent Land Use Management).
Data Availability Statement
All data used in the study can be found in the Appendix A.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table A1.
GPS coordinates of the quadrats.
| Mosonszolnok | ||||
| SHELT I | SHELT II | SHELT III | SHELT IV | |
| A | 47.85429, 17.14683 | 47.86501, 17.12457 | 47.86219, 17.19912 | 47.82253, 17.2079 |
| B | 47.85525, 17.14708 | 47.86484, 17.1254 | 47.86277, 17.20043 | 47.82196, 17.20875 |
| C | 47.85653, 17.14742 | 47.86467, 17.12718 | 47.86324, 17.20153 | 47.82114, 17.20992 |
| Sarród | ||||
| SHELT I | SHELT II | SHELT III | SHELT IV | |
| A | 47.65314, 16.87423 | 47.65546, 16.87111 | 47.65484, 16.87037 | 47.65232, 16.87603 |
| B | 47.65392, 16.87264 | 47.6558, 16.87156 | 47.65463, 16.87015 | 47.65223, 16.87625 |
| C | 47.65438, 16.87161 | 47.65665, 16.87261 | 47.65414, 16.86954 | 47.65197, 16.87679 |
| Sopronhorpács | ||||
| SHELT I | SHELT II | SHELT III | ||
| A | 47.493, 16.74852 | 47.47178, 16.72377 | 47.46881, 16.71774 | |
| B | 47.49838, 16.74481 | 47.4732, 16.72307 | 47.46984, 16.71738 | |
| C | 47.50114, 16.74311 | 47.4745, 16.72259 | 47.47036, 16.71707 | |
| Vasegerszeg | ||||
| SHELT I | SHELT II | SHELT III | ||
| A | 47.37375, 16.89984 | 47.3728, 16.90045 | 47.37294, 16.89763 | |
| B | 47.3736, 16.89995 | 47.37279, 16.90053 | 47.37259, 16.89771 | |
| C | 47.37346, 16.9 | 47.37262, 16.90068 | 47.37243, 16.89771 | |
Table A2.
Measured soil parameters in the examined shelterbelts.
| Plot | pH (KCl) | Viscosity | CaCO3 % | SOM % | NH4+ + NO3− N (mg/kg) | AL P (mg P2O5/kg) | AL K (mg K2O/kg) |
|---|---|---|---|---|---|---|---|
| MS1 | 7.11 ± 0.06 | 47.33 ± 1.25 | 5.99 ± 0.90 | 2.10 ± 0.22 | 8.90 ± 0.96 | 132.33 ± 14.34 | 513.00 ± 42.81 |
| MS2 | 6.98 ± 0.08 | 52.00 ± 1.41 | 3.33 ± 0.80 | 2.10 ± 0.16 | 9.70 ± 0.37 | 125.33 ± 7.72 | 615.67 ± 64.79 |
| MS3 | 6.88 ± 0.07 | 57.67 ± 2.87 | 5.99 ± 0.54 | 1.97 ± 0.17 | 10.07 ± 0.29 | 133.67 ± 5.31 | 513.67 ± 46.33 |
| MS4 | 7.19 ± 0.04 | 59.00 ± 4.90 | 7.99 ± 0.77 | 2.17 ± 0.17 | 9.47 ± 0.42 | 618.67 ± 24.96 | 747.67 ± 37.04 |
| SA1 | 7.29 ± 0.03 | 61.00 ± 6.68 | 22.25 ± 4.16 | 5.32 ± 0.15 | 8.06 ± 4.88 | 193.90 ± 11.49 | 532.00 ± 130.43 |
| SA2 | 7.26 ± 0.04 | 64.00 ± 7.87 | 26.22 ± 1.21 | 5.36 ± 0.03 | 2.32 ± 1.09 | 273.25 ± 163.27 | 769.33 ± 131.11 |
| SA3 | 7.22 ± 0.06 | 77.67 ± 12.26 | 16.15 ± 2.82 | 7.27 ± 1.40 | 6.85 ± 2.40 | 229.20 ± 58.24 | 880.33 ± 286.15 |
| SA4 | 7.45 ± 0.08 | 50.00 ± 4.90 | 13.74 ± 0.16 | 4.66 ± 0.40 | 3.52 ± 0.43 | 198.07 ± 15.34 | 543.33 ± 24.31 |
| SH1 | 6.83 ± 0.07 | 68.00 ± 2.16 | 0.68 ± 0.00 | 5.36 ± 0.08 | 3.66 ± 0.92 | 34.41 ± 13.20 | 384.00 ± 6.98 |
| SH2 | 6.48 ± 0.17 | 62.00 ± 1.41 | <0.10 | 4.86 ± 0.15 | 8.34 ± 4.42 | 81.57 ± 61.85 | 502.67 ± 64.22 |
| SH3 | 5.06 ± 0.54 | 52.33 ± 4.19 | <0.10 | 4.67 ± 0.69 | 27.12 ± 31.97 | 688.26 ± 927.21 | 589.67 ± 486.93 |
| VS1 | 6.23 ± 0.37 | 79.33 ± 7.54 | <0.10 | 5.31 ± 0.06 | 8.30 ± 1.05 | 268.19 ± 153.00 | 357.00 ± 99.03 |
| VS2 | 5.62 ± 0.65 | 73.33 ± 8.38 | <0.10 | 4.94 ± 0.58 | 14.43 ± 6.79 | 178.08 ± 106.25 | 399.00 ± 111.72 |
| VS3 | 5.86 ± 0.05 | 70.67 ± 11.12 | <0.10 | 5.32 ± 0.02 | 12.22 ± 1.14 | 208.04 ± 52.15 | 368.67 ± 21.70 |
Table A3.
Herbaceous species list and cover (%).
| Species/Cover (%) | MS1 | MS2 | MS3 | MS4 | SA1 | SA2 | SA3 | SA4 | SH1 | SH2 | SH3 | VS1 | VS2 | VS3 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Achillea millefolium | 0 | 0 | 0 | 0 | 0 | 3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Aegilops cylindrica | 0 | 0 | 0 | 0 | 0 | 10 | 13 | 7 | 0 | 0 | 0 | 0 | 0 | 0 |
| Ajuga reptans | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 17 | 0 |
| Alliaria petiolata | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 7 | 0 | 0 | 3 | 0 | 0 |
| Allium vineale | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 13 | 0 | 0 | 0 | 0 | 0 |
| Amaranthus retroflexus | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 0 | 0 |
| Anagallis arvensis | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Anemone ranunculoides | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 |
| Anthriscus caucalis | 0 | 0 | 0 | 20 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Anthriscus cerefolium | 0 | 0 | 0 | 10 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Arrhenatherum elatius | 0 | 33 | 0 | 7 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Arum orientale | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 12 | 0 | 0 | 0 | 0 | 3 |
| Ballota nigra | 20 | 13 | 30 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Brachypodium sylvaticum | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 0 | 0 | 0 | 0 | 0 |
| Bromus sterilis | 30 | 33 | 40 | 7 | 2 | 0 | 0 | 0 | 7 | 0 | 0 | 0 | 0 | 0 |
| Carduus acanthoides | 3 | 8 | 0 | 13 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Carex hirta | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 7 | 0 | 0 | 0 | 0 | 0 |
| Chenopodium album | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Circaea lutetiana | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 7 | 0 | 0 | 0 |
| Cirsium arvense | 0 | 0 | 0 | 3 | 0 | 0 | 8 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Cirsium vulgare | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 7 | 0 | 3 |
| Convolvulus arvensis | 2 | 2 | 7 | 0 | 3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Corydalis cava | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 12 | 0 | 5 |
| Crepis setosa | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 0 |
| Dactylis glomerata | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 7 | 0 | 0 | 0 | 0 | 0 |
| Daucus carota subsp. carota | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Dryopteris filix-mas | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 7 | 0 | 3 | 0 |
| Elymus repens | 0 | 7 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Erigeron annuus | 0 | 0 | 0 | 10 | 0 | 3 | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 0 |
| Euphorbia virgata | 3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Falcaria vulgaris | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Festuca rupicola | 0 | 0 | 0 | 0 | 0 | 3 | 3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Gagea lutea | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 5 |
| Galium aparine | 5 | 13 | 3 | 33 | 0 | 0 | 0 | 2 | 10 | 12 | 7 | 7 | 8 | 3 |
| Galium mollugo | 0 | 0 | 0 | 7 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Geranium robertianum | 0 | 0 | 0 | 0 | 3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Geranium sanguineum | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 3 | 0 | 0 |
| Geum urbanum | 0 | 0 | 17 | 12 | 3 | 7 | 10 | 3 | 15 | 0 | 3 | 12 | 17 | 8 |
| Glechoma hederacea | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 13 | 0 |
| Hypericum perforatum | 0 | 0 | 0 | 7 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Lamium maculatum | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 3 | 7 |
| Lamium purpureum | 2 | 5 | 7 | 23 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Onopordum acanthium | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 7 | 0 | 0 | 0 | 0 | 0 |
| Ornithogalum umbellatum | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 0 |
| Pimpinella saxifraga | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Poa pratensis | 0 | 0 | 0 | 3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Poa trivialis | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 7 | 0 | 0 | 0 | 0 | 0 |
| Polygonatum latifolium | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 50 | 3 | 5 | 0 | 0 | 0 |
| Polygonatum multiflorum | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 13 | 0 | 7 |
| Potentilla reptans | 0 | 0 | 0 | 3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Pulmonaria officinalis | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 0 | 2 |
| Ranunculus ficaria | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 10 | 0 | 0 | 33 | 15 | 33 |
| Salvia pratensis | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Setaria viridis | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 7 | 0 | 0 |
| Silene latifolia subsp. alba | 3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Solanum dulcamara | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 7 | 0 | 0 | 0 | 0 | 0 |
| Stachys sylvatica | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 7 | 0 | 0 | 3 |
| Stellaria media | 2 | 5 | 10 | 20 | 7 | 0 | 0 | 3 | 0 | 0 | 0 | 7 | 10 | 0 |
| Symphytum officinale | 2 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Taraxacum officinale | 0 | 0 | 0 | 0 | 2 | 3 | 0 | 0 | 3 | 0 | 0 | 3 | 0 | 0 |
| Thlaspi perfoliatum | 0 | 0 | 0 | 3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Tripleurospermum inodorum | 3 | 0 | 0 | 3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Urtica dioica | 0 | 3 | 0 | 0 | 10 | 0 | 0 | 0 | 7 | 0 | 3 | 0 | 3 | 0 |
| Verbascum lychnitis | 0 | 0 | 0 | 7 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Verbena officinalis | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 0 | 0 | 0 | 0 | 0 |
| Veronica arvensis | 2 | 0 | 5 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 10 | 0 | 0 |
| Veronica persica | 0 | 0 | 0 | 20 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Vinca minor | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 7 | 0 | 0 | 0 | 0 | 0 |
| Viola odorata | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 23 | 0 | 3 | 23 | 0 | 3 |
References
- Tiwari, P.; Kumar, R.; Thakur, L.; Singh, R. Soil Treatment through Agroforestry: A Review. Indian J. Pure Appl. Biosci. 2017, 5, 24–30. [Google Scholar] [CrossRef]
- Feliciano, D.; Ledo, A.; Hillier, J.; Nayak, D.R. Which agroforestry options give the greatest soil and above ground carbon benefits in different world regions? Agric. Ecosyst. Environ. 2018, 254, 117–129. [Google Scholar] [CrossRef]
- Jezeer, R.E.; Santos, M.J.; Verweija, P.A.; Boot, R.G.A.; Cough, Y. Benefits for multiple ecosystem services in Peruvian coffee agroforestry systems without reducing yield. Ecosyst. Serv. 2019, 40, 101033. [Google Scholar] [CrossRef]
- Elagib, N.A.; Al-Saidi, M. Balancing the benefits from the water–energy–land–food nexus through agroforestry in the Sahel. Sci. Total Environ. 2020, 742, 140509. [Google Scholar] [CrossRef]
- Tschora, H.; Cherubini, F. Co-benefits and trade-offs of agroforestry for climate change mitigation and other sustainability goals in West Africa. Glob. Ecol. Conserv. 2020, 22, e00919. [Google Scholar] [CrossRef]
- Winkler, D.; Traser, G. Collembola Diversity in Agricultural Environments (Lajta Project, Western Hungary). In Proceedings of the International Scientific Conference on Sustainable Development and Ecological Footprint, Sopron, Hungary, 26–27 March 2012. [Google Scholar]
- Gál, J. A mezőgazdasági terméshozamok növekedése az erdősávok védelmében. Erdészettudományi Közlemények 1963, 1–2, 41–83. (In Hungarian) [Google Scholar]
- Szarvas, P. Mezővédő Erdősávok, Fasorok Jellemzése, Ökológiai Feltárása, Kihatásai. Characterization, Ecological Exploration and Effects of Field Protection Forest Strips and Tree Lines. Ph.D. Dissertation, University of Debrecen, Debrecen, Hungary, 2010. (In Hungarian). [Google Scholar]
- Updawatta, R.P.; Rankoth, L.; Jose, S. Agroforestry and Biodiversity. Sustainability 2019, 11, 2879. [Google Scholar] [CrossRef] [Green Version]
- Pierce, R.A., II; Farrand, D.T.; Kurtz, W.B. Projecting the bird community response resulting from the adoption of shelterbelt agroforestry practices in Eastern Nebraska. Agrofor. Syst. 2001, 53, 333–350. [Google Scholar] [CrossRef]
- Bonifacio, R.S.; Kinross, C.M.; Gurr, G.M.; Nicol, H. The effect of woody plant diversity and other stand and landscape factors on the diversity and abundance of birds using farm shelterbelts. Pac. Conserv. Biol. 2011, 17, 22–35. [Google Scholar] [CrossRef]
- Leles, B.; Xiao, X.; Pasion, B.O.; Nakamura, A.; Tomlinson, K.W. Does plant diversity increase top-down control of herbivorous insects in tropical forest? Oikos 2017, 126, 1142–1149. [Google Scholar] [CrossRef]
- Pardon, P.; Reheul, D.; Mertens, J.; Reubens, B.; De Frenne, P.; De Smedt, P.; Proesmans, W.; Van Vooren, L.; Verheyen, C. Gradients in abundance and diversity of ground dwelling arthropods as a function of distance to tree rows in temperate arable agroforestry systems. Agric. Ecosyst. Environ. 2019, 270–271, 114–128. [Google Scholar] [CrossRef]
- Todd, J.H.; Poulton, J.; Richards, K.; Malone, L.A. Effect of orchard management, neighbouring land-use and shelterbelt tree composition on the parasitism of pest leafroller (Lepidoptera: Tortricidae) larvae in kiwifruit orchard shelterbelts. Agric. Ecosyst. Environ. 2018, 260, 27–35. [Google Scholar] [CrossRef]
- Gontijo, L.M. Engineering natural enemy shelters to enhance conservation biological control in field crops. Biol. Control 2019, 130, 155–163. [Google Scholar] [CrossRef]
- Mize, C.W.; Brandle, J.R.; Schoeneberger, M.M.; Bentrup, G. Ecological Development and Function of Shelterbelts in Temperate North America. In Toward Agroforestry Design; Jose, S., Gordon, M., Eds.; Springer: Berlin/Heidelberg, Germany, 2008; Volume 4, pp. 27–54. [Google Scholar]
- Hass, A.L.; Liese, B.; Heong, K.L.; Settele, J.; Tscharntke, T.; Westphal, C. Plant-pollinator interactions and bee functional diversity are driven by agroforests in rice-dominated landscapes. Agric. Ecosyst. Environ. 2018, 253, 140–147. [Google Scholar] [CrossRef]
- Donkersley, P. Trees for bees. Agric. Ecosyst. Environ. 2019, 270–271, 79–83. [Google Scholar] [CrossRef]
- Damschen, E.I. Landscape corridors. In Encyclopedia of Biodiversity, 2nd ed.; Levin, S.A., Ed.; Academic Press: Cambridge, MA, USA, 2013; pp. 467–475. [Google Scholar] [CrossRef]
- Damschen, E.I.; Haddad, N.M.; Orrock, J.L.; Tewksbury, J.J.; Levey, D.J. Corridors Increase Plant Species Richness at Large Scales. Science 2006, 313, 1284–1286. [Google Scholar] [CrossRef] [Green Version]
- Carlier, J.; Moran, J. Hedgerow typology and condition analysis to inform greenway design in rural landscapes. J. Environ. Manag. 2019, 247, 790–803. [Google Scholar] [CrossRef]
- Wilson, P.J. Restoring pollinator communities and pollination services in hedgerows in intensively managed agricultural landscapes. In The Ecology of Hedgerows and Field Margins; Dover, J.W., Ed.; Routledge: New York, NY, USA, 2019; pp. 163–185. [Google Scholar]
- Milberg, P.; Bergman, K.; Jonason, D.; Karlsson, J.; Westerberg, L. Land-use history influence the vegetation in coniferous production forests in southern Sweden. For. Ecol. Manag. 2019, 440, 23–30. [Google Scholar] [CrossRef]
- Harta, I.; Simon, B.; Vinogradov, S.; Winkler, D. Collembola communities and soil conditions in forest plantations established in an intensively managed agricultural area. J. For. Res. 2020, 29, 1291–1297. [Google Scholar] [CrossRef]
- Bihari, Z.; Babolcsai, G.; Bartholdy, J.; Ferenczi, Z.; Gerhát-Kerényi, J.; Haszpra, L.; Homoki-Ijváry, K.; Kovács, T.; Lakatos, M.; Németh, Á.; et al. National Atlas of Hungary: Natural Environment; Kocsis, K., Ed.; MTA CSFK Geographical Institute: Budapest, Hungary, 2018; pp. 58–69. [Google Scholar]
- Zepner, L.; Karrasch, P.; Wiemann, F.; Bernard, L. ClimateCharts.net—An interactive climate analysis web platform. Int. J. Digit. Earth 2020, 4, 338–356. [Google Scholar] [CrossRef]
- Simon, T. A Kisalföld természetes növénytakarója. Földrajzi Közlemények 1962, 86, 183–193. (In Hungarian) [Google Scholar]
- Dövényi, Z.; Ambrózy, P.; Juhász, Á.; Marosi, S.; Mezősi, G.; Michalkó, G.; Szalai, Z.; Tiner, T. Magyarország Kistájainak Katasztere (Inventory of Microregions in Hungary); MTA Földrajztudományi Kutatóintézet: Budapest, Hungary, 2010. (In Hungarian) [Google Scholar]
- Takács, V. A Sopronhorpácsi Mezővédő Erdősávrendszer Hasznosítási Lehetősége. Possibility of utilization of the Sopronhorpács shelterbelt system. Erdészeti Lapok 2004, 139, 127–130. (In Hungarian) [Google Scholar]
- Horváth, F.; Dobolyi, K.; Morschauer, T.; Lőkös, L.; Karas, L.; Szerdahelyi, T. Flóra Adatbázis [Flora Database]; MTA ÖBKI: Vácrátót, Hungary, 1995. (In Hungarian) [Google Scholar]
- Tóthmérész, B. Diversity; University of Debrecen: Debrecen, Hungary, 2013; p. 290. [Google Scholar]
- Grime, J.P. Plant Strategies, Vegetation Processes, and Ecosystem Properties; Wiley: New York, NY, USA, 1979. [Google Scholar]
- Borhidi, A. Social behaviour types, the naturalness and relative ecological indicator values of the higher plants in the Hungarian flora. Acta Bot Hung 1995, 39, 97–181. [Google Scholar]
- Legendre, P.; Gallagher, E.D. Ecologically meaningful transformations for ordination of species data. Oecologia 2001, 129, 271–280. [Google Scholar] [CrossRef]
- Rose, F. Indicators of ancient woodland. Br. Wildl. 1999, 10, 241–251. [Google Scholar]
- Dzwonko, Z.; Loster, S. Distribution of vascular plant species in small woodlands on the Western Carpathian foothills. Oikos 1989, 56, 77–86. [Google Scholar] [CrossRef]
- Hermy, A.; Honnay, O.; Firbank, L.; Grashof-Bokdam, C.; Lawesson, J.L. An ecological comparison between ancient and other forest plant species of Europe, and the implications for forest conservation. Biol. Conserv. 1999, 91, 9–22. [Google Scholar] [CrossRef]
- Wulf, M. Plant species as indicators of ancient woodland in northwestern Germany. J. Veg. Sci. 1997, 8, 635–642. [Google Scholar] [CrossRef]
- Reif, A.; Achtziger, R. Gebüsche, Hecken, Waldmäntel, Feldgehölze. In Handbuch Naturschutz und Landschaftspflege; Konold, W., Böcker, R., Hampicke, U., Eds.; Wiley: New York, NY, USA, 2014. [Google Scholar] [CrossRef]
- Sciama, D.; Augusto, L.; Dupouey, J.L.; Gonzalez, M.; Moares Domínguez, C. Floristic and ecological differences between recent and ancient forests growing on non-acidic soils. For. Ecol. Manag. 2009, 258, 600–608. [Google Scholar] [CrossRef]
- Peterken, G.F.; Game, M. Historical factors affecting the nuber and distribution of vascular plant species in the woodlands of Central Lincolnshire. J. Ecol. 1984, 72, 155–182. [Google Scholar] [CrossRef]
- Van Den Berge, S.; Baeten, L.; Vanhellemont, M.; Ampoorter, E.; Proesmans, W.; Earaerts, M.; Hermy, M.; Smagghe, G.; Vermeulen, I.; Verheyen, K. Species diversity, pollinator resource value and edibility potential of woody networks in the countryside in northern Belgium. Agric. Ecosyst. Environ. 2018, 259, 119–126. [Google Scholar] [CrossRef]
- Wehling, S.; Diekmann, M. Factors influencing the spatial distribution of forest plant species in hedgerows of North-western Germany. Biodivers. Conserv. 2008, 17, 2799–2813. [Google Scholar] [CrossRef]
- Aude, E.; Tybirk, K.; Bruus Pedersen, M. Vegetation diversity of conventional and organic hedgerows in Denmark. Agric. Ecosyst. Environ. 2003, 99, 135–147. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Position and optic of the four study areas in NW Hungary.

Figure 2.
Diversity profiles of herbaceous plants in the examined shelterbelts in Mosonszolnok, Sarród, Sopronhorpács, and Vasegerszeg.
Figure 2.
Diversity profiles of herbaceous plants in the examined shelterbelts in Mosonszolnok, Sarród, Sopronhorpács, and Vasegerszeg.

Figure 3.
Distribution of herbaceous species by social behavior type (SBT) categories.

Figure 4.
Distribution of herbaceous species by forest affinity categories.

Figure 5.
The redundancy analysis shows the impact of common oak (Qurob), Osage orange (Mapom), and former chariot roads (ROAD) on the herbaceous species composition of shelterbelts, while other factors (e.g., N) are not significant.
Figure 5.
The redundancy analysis shows the impact of common oak (Qurob), Osage orange (Mapom), and former chariot roads (ROAD) on the herbaceous species composition of shelterbelts, while other factors (e.g., N) are not significant.

Table 1.
Social behavior type categories and naturalness values [33].
Table 1.
Social behavior type categories and naturalness values [33].
| Habitat Type | Social Behavior Type | Abb. | Nat. Value |
|---|---|---|---|
| Natural habitats | Specialists | S | +6 |
| Competitors | C | +5 | |
| Generalists | G | +4 | |
| Natural pioneers | NP | +3 | |
| Disturbed, secondary, and artificial habitats | Disturbance-tolerant | DT | +2 |
| Weeds | W | +1 | |
| Introduced alien species | I | −1 | |
| Adventives | A | −1 | |
| Ruderal competitors | RC | −2 | |
| Aggressive alien species | AC | −3 |
Table 2.
Forest affinity categories and their abbreviations in the analysis.
| Forest Affinity Category | Abbreviation |
|---|---|
| Obligate forest species | FOREST |
| Species typical to forest edges and clearings | FOR MARG |
| Species that can be found both in woody habitats and open (non-woody) land | FOR/OP |
| Species appearing mainly in open land but also in forests | OP (FOR) |
| Species specific to open habitats | OPEN |
Table 3.
Tree species composition and cover values (%) in the examined shelterbelts.
| Layer/Species | MS1 | MS2 | MS3 | MS4 | SA1 | SA2 | SA3 | SA4 | SH1 | SH2 | SH3 | VS1 | VS2 | VS3 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Canopy layer cover | 70 | 80 | 80 | 90 | 90 | 80 | 90 | 95 | 90 | 95 | 80 | 80 | 90 | 80 |
| Acer campestre | 50 | 70 | 80 | 50 | 30 | 40 | ||||||||
| Acer platanoides | 5 | 5 | 10 | 30 | 30 | 20 | 10 | 80 | 10 | |||||
| Cerasus avium | 10 | |||||||||||||
| Fraxinus excelsior | 5 | 5 | ||||||||||||
| Fraxinus pennsylvanica | 60 | 20 | 10 | 20 | ||||||||||
| Gleditsia triacanthos | 5 | |||||||||||||
| Koelreuteria paniculata | 1 | |||||||||||||
| Larix decidua | 5 | |||||||||||||
| Maclura pomifera | 5 | 10 | 20 | 20 | 20 | |||||||||
| Pinus sylvestris | 30 | |||||||||||||
| Populus × canescens | 20 | 10 | ||||||||||||
| Quercus cerris | 10 | 10 | 10 | |||||||||||
| Quercus robur | 20 | 30 | 50 | 60 | 50 | |||||||||
| Quercus rubra | 10 | |||||||||||||
| Robinia pseudoacacia | 30 | 60 | 10 | 20 | 10 | |||||||||
| Salix alba | 10 | |||||||||||||
| Ulmus minor | 10 | 30 | 50 | 40 | 50 | 50 | 50 | 50 |
Table 4.
Shrub species composition and cover values (%) in the examined shelterbelts.
| Layer/Species | MS1 | MS2 | MS3 | MS4 | SA1 | SA2 | SA3 | SA4 | SH1 | SH2 | SH3 | VS1 | VS2 | VS3 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Shrub layer cover | 30 | 40 | 20 | 30 | 20 | 30 | 40 | 30 | 50 | 15 | 30 | 20 | ||
| Cornus mas | 20 | |||||||||||||
| Cornus sanguinea | 10 | |||||||||||||
| Crataegus monogyna | 10 | |||||||||||||
| Elaeagnus angustifolia | 10 | |||||||||||||
| Euonymus verrucosus | 20 | |||||||||||||
| Ligustrum vulgare | 20 | 20 | 20 | 20 | 20 | |||||||||
| Maclura pomifera | 5 | 20 | ||||||||||||
| Prunus spinosa | 10 | 10 | 15 | 10 | 20 | 20 | 5 | 20 | 10 | |||||
| Sambucus nigra | 10 | 10 | 5 | 10 | 10 | 5 | 5 | |||||||
| Ulmus minor | 10 | 20 | ||||||||||||
| Viburnum lantana | 20 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Szigeti, N.; Berki, I.; Vityi, A.; Rasran, L. Shelterbelts Planted on Cultivated Fields Are Not Solutions for the Recovery of Former Forest-Related Herbaceous Vegetation. Land 2021, 10, 930. https://doi.org/10.3390/land10090930
AMA Style
Szigeti N, Berki I, Vityi A, Rasran L. Shelterbelts Planted on Cultivated Fields Are Not Solutions for the Recovery of Former Forest-Related Herbaceous Vegetation. Land. 2021; 10(9):930. https://doi.org/10.3390/land10090930
Chicago/Turabian StyleSzigeti, Nóra, Imre Berki, Andrea Vityi, and Leonid Rasran. 2021. "Shelterbelts Planted on Cultivated Fields Are Not Solutions for the Recovery of Former Forest-Related Herbaceous Vegetation" Land 10, no. 9: 930. https://doi.org/10.3390/land10090930
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.