
Isolation, Characterization and Neuroprotective Activity of Folecitin: An In Vivo Study

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Bioactive Compound Isolation
2.2.1. Plant Materials
2.2.2. Folecitin Isolation and Characterization
2.3. Neuroprotective Efficacy of Folecitin
2.3.1. Animals Used in the Experiment
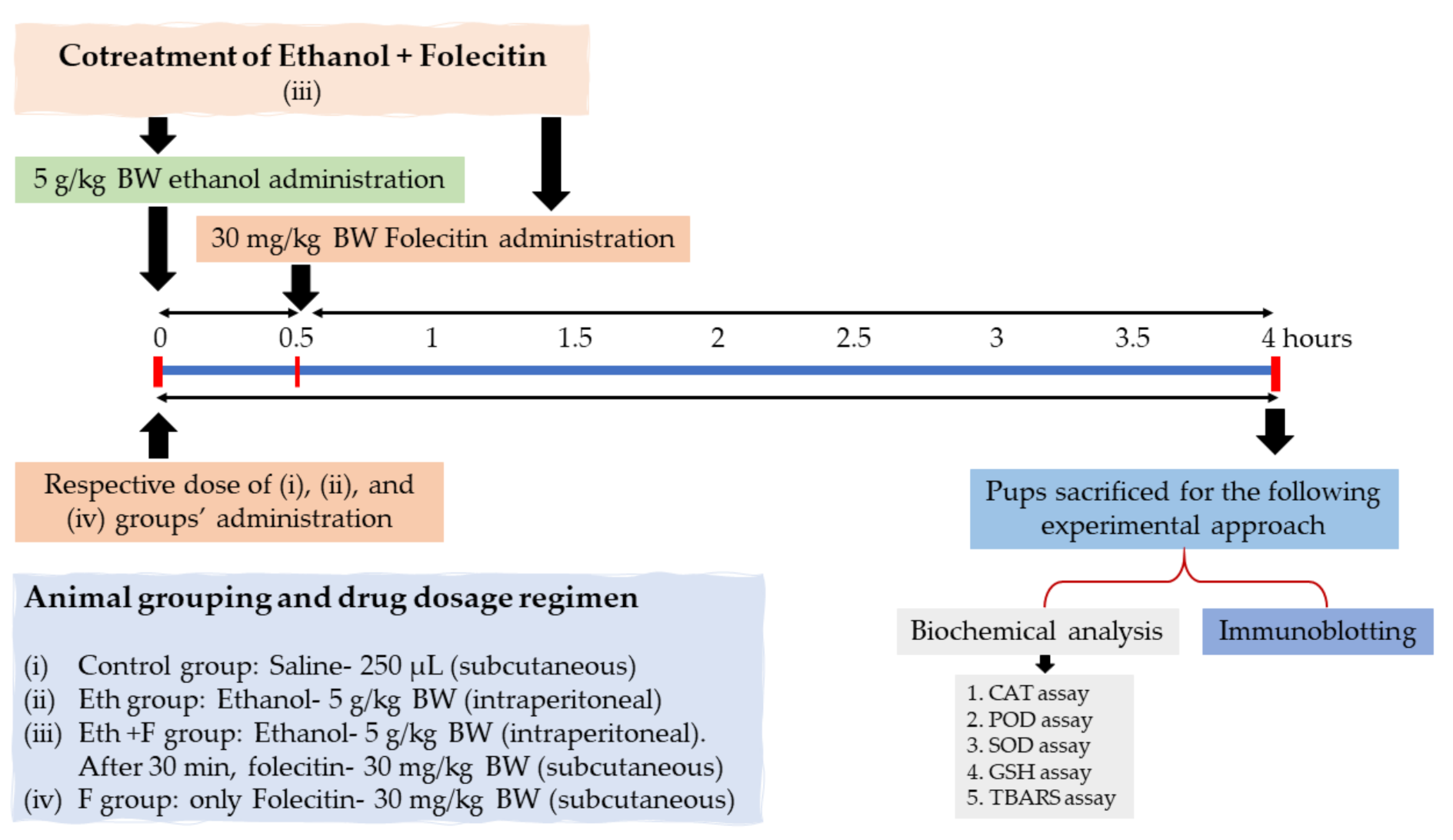
2.3.2. Experimental Design and Approach
2.3.3. Protein Extraction for Biochemical Analysis and Immunoblotting
2.3.4. Biochemical Analysis
Catalase Assay (CAT)
Peroxidase Assay (POD)
Superoxide Dismutase Assay (SOD)
Reduced Glutathione Assay (GSH)
Approximation of Lipid Peroxidation
2.4. Western Blotting
2.5. Statistical Analysis
3. Results
3.1. Folecitin’s Yield
3.2. Folecitin’s Characterization
3.2.1. Electrospray Ionization Mass Spectrometry (ESI–MS) Spectrum
3.2.2. Infrared Spectroscopy (IR) Spectrum
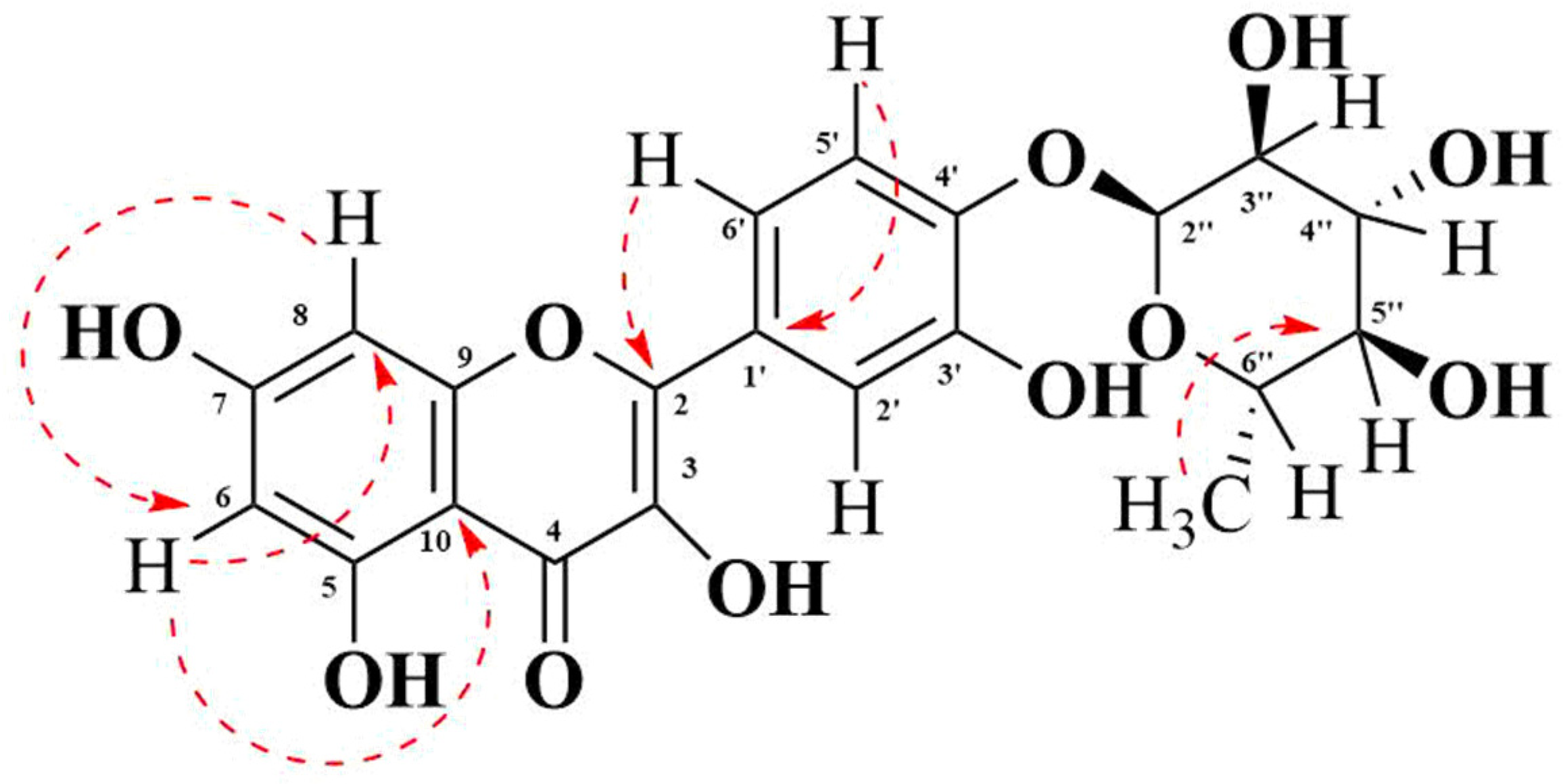
3.2.3. Nuclear Magnetic Resonance Spectroscopy (NRM) Spectrum
3.3. Neuroprotective Pharmacology
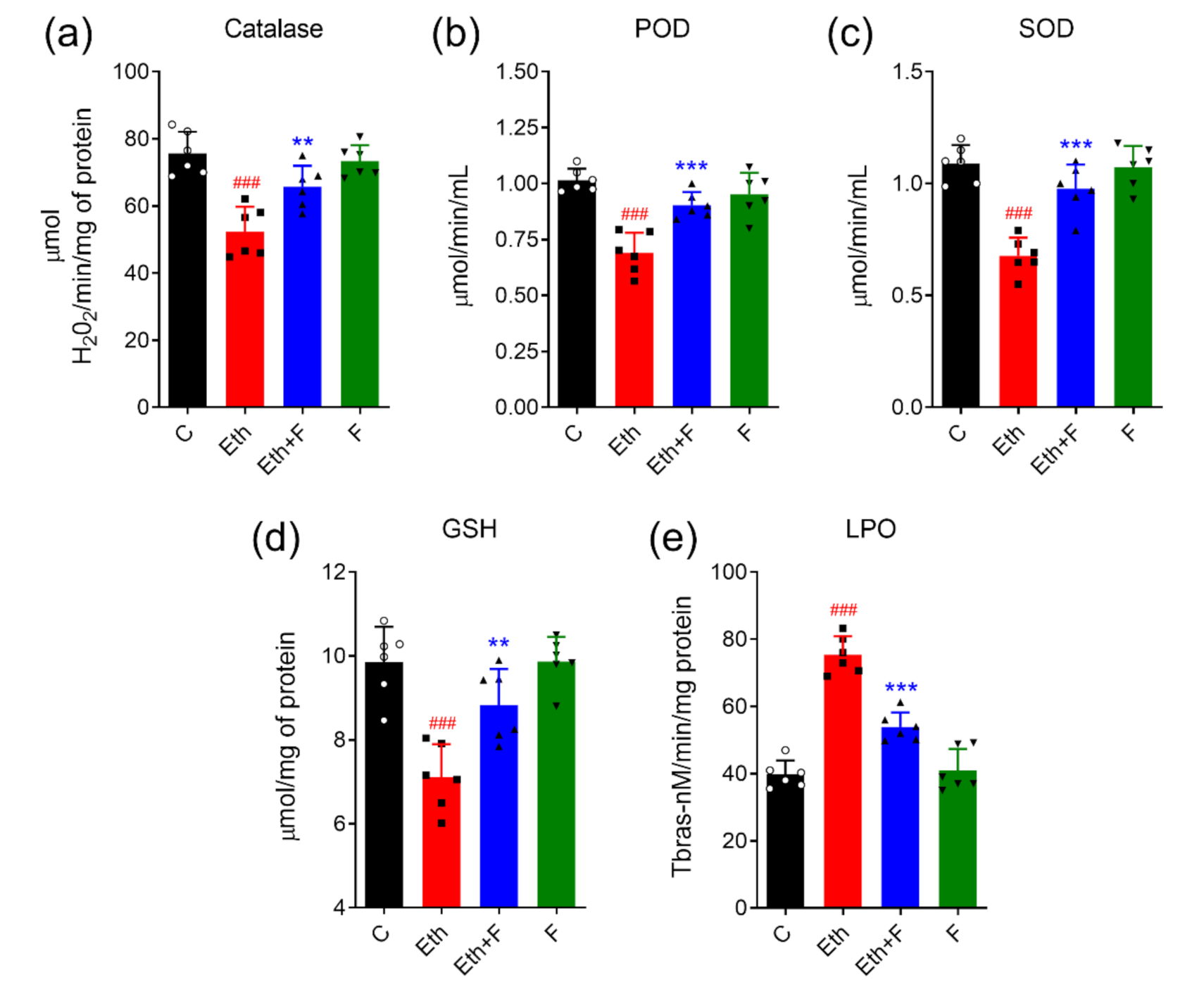
3.3.1. Folecitin Reduced Oxidative Stress Induced by Ethanol
3.3.2. Folecitin Inhibited Neuroinflammatory Markers
3.3.3. Folecitin Deactivated the NLRP-3 Inflammasome Complex
3.3.4. Folecitin Reversed the Expression of the Neuro-Apoptotic Proteins
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Erkkinen, M.G.; Kim, M.O.; Geschwind, M.D. Clinical Neurology and Epidemiology of the Major Neurodegenerative Diseases. Cold Spring Harb. Perspect. Biol. 2018, 10, 4. [Google Scholar] [CrossRef] [Green Version]
- Sun, A.Y.; Sun, G.Y. Ethanol and oxidative mechanisms in the brain. J. Biomed. Sci. 2001, 8, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Ramezani, A.; Goudarzi, I.; Lashkarboluki, T.; Ghorbanian, M.T.; Abrari, K.; Elahdadi Salmani, M. Role of Oxidative Stress in Ethanol-induced Neurotoxicity in the Developing Cerebellum. Iran J. Basic Med. Sci. 2012, 15, 965–974. [Google Scholar] [PubMed]
- Wilson, C.A.; Finch, E.; Kerr, C.; Shakespeare, J. Alcohol, smoking, and other substance use in the perinatal period. BMJ 2020, 369, m1627. [Google Scholar] [CrossRef]
- Cohen-Kerem, R.; Koren, G. Antioxidants and fetal protection against ethanol teratogenicity. I. Review of the experimental data and implications to humans. Neurotoxicol. Teratol. 2003, 25, 1–9. [Google Scholar] [CrossRef]
- Singh, A.; Kukreti, R.; Saso, L.; Kukreti, S. Oxidative Stress: A Key Modulator in Neurodegenerative Diseases. Molecules 2019, 24, 1583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, S.K.; Vasudevan, D.M. Alcohol-induced oxidative stress. Life Sci. 2007, 81, 177–187. [Google Scholar] [CrossRef]
- Ghosh, A.P.; Walls, K.C.; Klocke, B.J.; Toms, R.; Strasser, A.; Roth, K.A. The proapoptotic BH3-only, Bcl-2 family member, Puma is critical for acute ethanol-induced neuronal apoptosis. J. Neuropathol. Exp. Neurol. 2009, 68, 747–756. [Google Scholar] [CrossRef] [Green Version]
- Young, C.; Roth, K.A.; Klocke, B.J.; West, T.; Holtzman, D.M.; Labruyere, J.; Qin, Y.Q.; Dikranian, K.; Olney, J.W. Role of caspase-3 in ethanol-induced developmental neurodegeneration. Neurobiol. Dis. 2005, 20, 608–614. [Google Scholar] [CrossRef]
- Haorah, J.; Ramirez, S.H.; Floreani, N.; Gorantla, S.; Morsey, B.; Persidsky, Y. Mechanism of alcohol-induced oxidative stress and neuronal injury. Free Radic. Biol. Med. 2008, 45, 1542–1550. [Google Scholar] [CrossRef] [Green Version]
- Sokol, R.J.; Delaney-Black, V.; Nordstrom, B. Fetal alcohol spectrum disorder. JAMA 2003, 290, 2996–2999. [Google Scholar] [CrossRef] [PubMed]
- Hirsch, E.C. Animal models in neurodegenerative diseases. In Neuropsychiatric Disorders: An Integrative Approach; Springer Science + Business Media: New York, NY, USA, 2007; pp. 87–90. [Google Scholar]
- Gurney, M.E.; Pu, H.; Chiu, A.Y.; Dal Canto, M.C.; Polchow, C.Y.; Alexander, D.D.; Caliendo, J.; Hentati, A.; Kwon, Y.W.; Deng, H.X.; et al. Motor neuron degeneration in mice that express a human Cu,Zn superoxide dismutase mutation. Science 1994, 264, 1772–1775. [Google Scholar] [CrossRef]
- Carlsson, A.; Lindqvist, M.; Magnusson, T. 3,4-Dihydroxyphenylalanine and 5-hydroxytryptophan as reserpine antagonists. Nature 1957, 180, 1200. [Google Scholar] [CrossRef]
- Dawson, T.M.; Golde, T.E.; Lagier-Tourenne, C. Animal models of neurodegenerative diseases. Nat Neurosci. 2018, 21, 1370–1379. [Google Scholar] [CrossRef]
- Ikonomidou, C.; Bittigau, P.; Ishimaru, M.J.; Wozniak, D.F.; Koch, C.; Genz, K.; Price, M.T.; Stefovska, V.; Horster, F.; Tenkova, T.; et al. Ethanol-induced apoptotic neurodegeneration and fetal alcohol syndrome. Science 2000, 287, 1056–1060. [Google Scholar] [CrossRef] [PubMed]
- Jevtovic-Todorovic, V.; Hartman, R.E.; Izumi, Y.; Benshoff, N.D.; Dikranian, K.; Zorumski, C.F.; Olney, J.W.; Wozniak, D.F. Early exposure to common anesthetic agents causes widespread neurodegeneration in the developing rat brain and persistent learning deficits. J. Neurosci. 2003, 23, 876–882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anand, K.S.; Dhikav, V. Hippocampus in health and disease: An overview. Ann. Indian Acad. Neurol. 2012, 15, 239–246. [Google Scholar] [CrossRef]
- Abrahao, K.P.; Salinas, A.G.; Lovinger, D.M. Alcohol and the Brain: Neuronal Molecular Targets, Synapses, and Circuits. Neuron 2017, 96, 1223–1238. [Google Scholar] [CrossRef] [Green Version]
- Nagy, J. Alcohol related changes in regulation of NMDA receptor functions. Curr. Neuropharmacol. 2008, 6, 39–54. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Zhao, J.; Liu, Z.; Guo, F.; Wang, Y.; Wang, X.; Zhang, R.; Vreugdenhil, M.; Lu, C. Acute Ethanol Inhibition of gamma Oscillations Is Mediated by Akt and GSK3beta. Front. Cell. Neurosci. 2016, 10, 189. [Google Scholar] [CrossRef] [Green Version]
- Shah, S.A.; Khan, M.; Jo, M.H.; Jo, M.G.; Amin, F.U.; Kim, M.O. Melatonin stimulates the SIRT 1/Nrf2 signaling pathway counteracting lipopolysaccharide (LPS)-induced oxidative stress to rescue postnatal rat brain. CNS Neurosci. Ther. 2017, 23, 33–44. [Google Scholar] [CrossRef]
- Shah, S.A.; Yoon, G.H.; Kim, M.O. Protection of the developing brain with anthocyanins against ethanol-induced oxidative stress and neurodegeneration. Mol. Neurobiol. 2015, 51, 1278–1291. [Google Scholar] [CrossRef] [PubMed]
- Shahid, M.; Subhan, F.; Islam, N.U.; Ahmad, N.; Farooq, U.; Abbas, S.; Akbar, S.; Ullah, I.; Raziq, N.; Din, Z.U. The antioxidant N-(2-mercaptopropionyl)-glycine (tiopronin) attenuates expression of neuropathic allodynia and hyperalgesia. Naunyn-Schmiedebergs Arch. Pharmacol. 2021, 394, 603–617. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.S.; Urbi, Z. Effect of Naphthalene Acetic Acid on the Adventitious Rooting in Shoot Cuttings ofAndrographis paniculata (Burm.f.) Wall. ex Nees: An Important Therapeutical Herb. Int. J. Agron. 2016, 2016, 1617543. [Google Scholar] [CrossRef] [Green Version]
- Ismail, N.A.; Hossain, M.S.; Mohd Mustafa, N.H.; Chia Phang, I. Morpho-physiological characteristics, selected macronutrient uptake, and oxidative stress level of Andrographis paniculata under saline condition. J. Teknol. 2015, 77, 135–140. [Google Scholar] [CrossRef] [Green Version]
- Hossain, M.S.; Urbi, Z.; Sule, A.; Hafizur Rahman, K.M. Andrographis paniculata (Burm. f.) Wall. ex Nees: A review of ethnobotany, phytochemistry, and pharmacology. Sci. World J. 2014, 2014, 274905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hossain, M.S.; Urbi, Z.; Phang, I.C. Auxin increased adventitious root development in the medicinal plant Andrographis paniculata (Burm. f.) Wall. ex Nees. Agron. J. 2021. [Google Scholar] [CrossRef]
- Hossain, M.S. The Effect of Salinity Stress on the Morpho-physiology and Protein Profile of Andrographis Paniculata. Master’s Thesis, International Islamic University Malaysia, Kuantan, Pahang, Malaysia, 2016. [Google Scholar]
- Hossain, S.; Urbi, Z.; Karuniawati, H.; Mohiuddin, R.B.; Moh Qrimida, A.; Allzrag, A.M.M.; Ming, L.C.; Pagano, E.; Capasso, R. Andrographis paniculata (Burm. f.) Wall. ex Nees: An Updated Review of Phytochemistry, Antimicrobial Pharmacology, and Clinical Safety and Efficacy. Life 2021, 11, 348. [Google Scholar] [CrossRef] [PubMed]
- Urbi, Z.; Hossain, M.S.; Rahman, K.M.H.; Zayed, T.M. Grape: A medicinal fruit species in the holy Qur’an and its ethnomedicinal importance. World Appl. Sci. J. 2014, 30, 253–265. [Google Scholar]
- Hossain, M.S.; Urbi, Z.; Evamoni, F.Z.; Zohora, F.T.; Rahman, K.M.H. A secondary research on medicinal plants mentioned in the Holy Qur’an. J. Med. Plants 2016, 3, 81–97. [Google Scholar]
- Raziq, N.; Saeed, M.; Ali, M.S.; Zafar, S.; Ali, M.I. In vitro anti-oxidant potential of new metabolites from Hypericum oblongifolium (Guttiferae). Nat. Prod. Res. 2015, 29, 2265–2270. [Google Scholar] [CrossRef]
- Raziq, N.; Saeed, M.; Shahid, M.; Muhammad, N.; Khan, H.; Gul, F. Pharmacological basis for the use of Hypericum oblongifolium as a medicinal plant in the management of pain, inflammation and pyrexia. BMC Complement. Altern. Med. 2016, 16, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butola, J.S.; Pant, S.; Samant, S.S. Diversity, distribution and indigenous uses of the Hypericum Species in Indian Himalayan Region. Ethnobot. Leafl. 2007, 2007, 4. [Google Scholar]
- Hassan, M.J.; Saddiqe, Z.; Shahzad, M.; Hussain, J.; Parveen, S.; Maimoona, A. Non-Toxic fractions of Hypericum perforatum and Hypericum oblongifolium inhibit protein glycation, free radicals production and lipid peroxidation in vitro. International J. Phytomedicine 2013, 5, 191–196. [Google Scholar]
- Sajid, A.; Ahmed, E.; Sharif, A.; Arshed, F.; Arshad, M.; Sher, M.; Sajid, A.; Amanat, S. Bioassay Directed Isolation Studies on Hypericum oblongifolium. J. Chem. Soc. Pak. 2018, 40, 249–254. [Google Scholar]
- Ayaz, M.; Sadiq, A.; Junaid, M.; Ullah, F.; Ovais, M.; Ullah, I.; Ahmed, J.; Shahid, M. Flavonoids as prospective neuroprotectants and their therapeutic propensity in aging associated neurological disorders. Front. Aging Neurosci. 2019, 11, 155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Animal Testing and Research: Guidance for the Regulated Community. Available online: https://www.gov.uk/guidance/research-and-testing-using-animals (accessed on 7 July 2019).
- Badshah, H.; Ali, T.; Kim, M.O. Osmotin attenuates LPS-induced neuroinflammation and memory impairments via the TLR4/NFκB signaling pathway. Sci. Rep. 2016, 6, 24493. [Google Scholar] [CrossRef]
- Olney, J.W.; Wozniak, D.F.; Jevtovic-Todorovic, V.; Farber, N.B.; Bittigau, P.; Ikonomidou, C. Drug-induced apoptotic neurodegeneration in the developing brain. Brain Pathol. 2002, 12, 488–498. [Google Scholar] [CrossRef] [PubMed]
- Vauzour, D.; Vafeiadou, K.; Rodriguez-Mateos, A.; Rendeiro, C.; Spencer, J.P. The neuroprotective potential of flavonoids: A multiplicity of effects. Genes Nutr. 2008, 3, 115–126. [Google Scholar] [CrossRef] [Green Version]
- Samarghandian, S.; Farkhondeh, T.; Azimi-Nezhad, M. Protective Effects of Chrysin Against Drugs and Toxic Agents. Dose Response 2017, 15, 1559325817711782. [Google Scholar] [CrossRef] [PubMed]
- Salehi, B.; Machin, L.; Monzote, L.; Sharifi-Rad, J.; Ezzat, S.M.; Salem, M.A.; Merghany, R.M.; El Mahdy, N.M.; Kilic, C.S.; Sytar, O.; et al. Therapeutic Potential of Quercetin: New Insights and Perspectives for Human Health. ACS Omega 2020, 5, 11849–11872. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Zhang, N.; Yang, D.; Yang, M.; Guo, X.; He, J.; Wu, W.; Ji, B.; Cheng, Q.; Zhou, F. Protective Effects of Five Structurally Diverse Flavonoid Subgroups against Chronic Alcohol-Induced Hepatic Damage in a Mouse Model. Nutrients 2018, 10, 1754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anand David, A.V.; Arulmoli, R.; Parasuraman, S. Overviews of Biological Importance of Quercetin: A Bioactive Flavonoid. Pharm. Rev 2016, 10, 84–89. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.Y.; Li, Q.; Bi, K.S. Bioactive flavonoids in medicinal plants: Structure, activity and biological fate. Asian J. Pharm. Sci. 2018, 13, 12–23. [Google Scholar] [CrossRef]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An overview. J. Nutr. Sci. 2016, 5, e47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abo-Ouf, H.; Hooper, A.W.; White, E.J.; Janse van Rensburg, H.J.; Trigatti, B.L.; Igdoura, S.A. Deletion of tumor necrosis factor-alpha ameliorates neurodegeneration in Sandhoff disease mice. Hum. Mol. Genet. 2013, 22, 3960–3975. [Google Scholar] [CrossRef] [Green Version]
- Rosenzweig, H.L.; Planck, S.R.; Rosenbaum, J.T. NLRs in immune privileged sites. Curr. Opin. Pharmacol. 2011, 11, 423–428. [Google Scholar] [CrossRef] [Green Version]
- Abais, J.M.; Xia, M.; Zhang, Y.; Boini, K.M.; Li, P.-L. Redox regulation of NLRP3 inflammasomes: ROS as trigger or effector? Antioxid. Redox Signal. 2015, 22, 1111–1129. [Google Scholar] [CrossRef] [Green Version]
- Mao, L.; Kitani, A.; Strober, W.; Fuss, I.J. The role of NLRP3 and IL-1β in the pathogenesis of inflammatory bowel disease. Front. Immunol. 2018, 9, 2566. [Google Scholar] [CrossRef] [Green Version]
- Kalejaiye, O.; Getachew, B.; Ferguson, C.L.; Taylor, R.E.; Tizabi, Y. Alcohol-Induced Increases in Inflammatory Cytokines Are Attenuated by Nicotine in Region-Selective Manner in Male Rats. J. Drug Alcohol Res. 2017, 6, 236036. [Google Scholar] [CrossRef] [Green Version]
- Kane, C.J.; Drew, P.D. Inflammatory responses to alcohol in the CNS: Nuclear receptors as potential therapeutics for alcohol-induced neuropathologies. J. Leukoc Biol. 2016, 100, 951–959. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Yu, H.; Wang, C.; Liu, Y.; You, J.; Wang, P.; Xu, G.; Shen, H.; Yao, H.; Lan, X.; et al. Chronic ethanol exposure induces neuroinflammation in H4 cells through TLR3/NF-kappaB pathway and anxiety-like behavior in male C57BL/6 mice. Toxicology 2020, 446, 152625. [Google Scholar] [CrossRef] [PubMed]
- Schaffner, S.L.; Lussier, A.A.; Baker, J.A.; Goldowitz, D.; Hamre, K.M.; Kobor, M.S. Neonatal Alcohol Exposure in Mice Induces Select Differentiation- and Apoptosis-Related Chromatin Changes Both Independent of and Dependent on Sex. Front. Genet. 2020, 11, 35. [Google Scholar] [CrossRef]
- Topper, L.A.; Baculis, B.C.; Valenzuela, C.F. Exposure of neonatal rats to alcohol has differential effects on neuroinflammation and neuronal survival in the cerebellum and hippocampus. J. Neuroinflammation 2015, 12, 160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cobley, J.N.; Fiorello, M.L.; Bailey, D.M. 13 reasons why the brain is susceptible to oxidative stress. Redox Biol. 2018, 15, 490–503. [Google Scholar] [CrossRef]
- Massaad, C.A.; Klann, E. Reactive oxygen species in the regulation of synaptic plasticity and memory. Antioxid. Redox Signal. 2011, 14, 2013–2054. [Google Scholar] [CrossRef] [Green Version]
- Behl, C. Vitamin E protects neurons against oxidative cell death in vitro more effectively than 17-beta estradiol and induces the activity of the transcription factor NF-kappaB. J. Neural Transm. 2000, 107, 393–407. [Google Scholar] [CrossRef] [PubMed]
- Marino, M.D.; Aksenov, M.Y.; Kelly, S.J. Vitamin E protects against alcohol-induced cell loss and oxidative stress in the neonatal rat hippocampus. Int. J. Dev. Neurosci. 2004, 22, 363–377. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S.# | Antibodies Name | Catalogue # |
|---|---|---|
| 1 | Anti-PARP-1 | sc-8007 |
| 2 | Anti-NLRP3 | ab270449 |
| 3 | Anti-TNFα | sc-52746 |
| 4 | Anti-NF-kB | sc-8008 |
| 5 | Anti-Caspase-1 | sc-56036 |
| 6 | Anti-Caspase-3 | sc-7272 |
| 7 | Anti-IL-1β | sc-32294 |
| 8 | Anti-BAX | sc-7480 |
| 9 | Anti- BCL-2 | sc-7382 |
| 10 | Anti-ASC | sc-514414 |
| 11 | Anti-p-JNK | sc-6254 |
| 12 | Anti-beta actin | sc-47778 |
| 13 | Goat anti-mouse (IgG-HRPs) secondary antibodies | sc-2031 |
| Traits | Results/Description |
|---|---|
| Physical state | Yellowish-brown crystalline solid powders |
| UV activity | UV active over TLC |
| Rf value | 0.4 [methanol (1): ethyl acetate (9)] |
| Molecular formula | C21H20O11 |
| Crude extract | 400 g (ethyl acetate fraction) |
| Isolated quantity | 75 mg |
| Yield (kg/crude extract) | 175 |
| Melting point (°C) | 187–189 |
| Solubility | Sparingly soluble in methanol at room temperature |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Farooq, U.; Khan, T.; Shah, S.A.; Hossain, M.S.; Ali, Y.; Ullah, R.; Raziq, N.; Shahid, M.; Capasso, R. Isolation, Characterization and Neuroprotective Activity of Folecitin: An In Vivo Study. Life 2021, 11, 825. https://doi.org/10.3390/life11080825
Farooq U, Khan T, Shah SA, Hossain MS, Ali Y, Ullah R, Raziq N, Shahid M, Capasso R. Isolation, Characterization and Neuroprotective Activity of Folecitin: An In Vivo Study. Life. 2021; 11(8):825. https://doi.org/10.3390/life11080825
Chicago/Turabian StyleFarooq, Umar, Taous Khan, Shahid Ali Shah, Md. Sanower Hossain, Yousaf Ali, Rahim Ullah, Naila Raziq, Muhammad Shahid, and Raffaele Capasso. 2021. "Isolation, Characterization and Neuroprotective Activity of Folecitin: An In Vivo Study" Life 11, no. 8: 825. https://doi.org/10.3390/life11080825