
Gut Microbiome and Metabolites in Patients with NAFLD and after Bariatric Surgery: A Comprehensive Review

1
Department of Internal and Vascular Medicine, Amsterdam University Medical Centers, AMC, 1105 AZ Amsterdam, The Netherlands
2
Department of Bariatric and General Surgery, Spaarne Hospital, 2134 TM Hoofddorp, The Netherlands
3
Department of Internal Medicine, Spaarne Hospital, 2134 TM Hoofddorp, The Netherlands
*
Author to whom correspondence should be addressed.
Metabolites 2021, 11(6), 353; https://doi.org/10.3390/metabo11060353
Submission received: 12 April 2021
/
Revised: 26 May 2021
/
Accepted: 26 May 2021
/
Published: 31 May 2021
(This article belongs to the Special Issue Steroids in Non-alcoholic Fatty Liver Disease)
Abstract
:The prevalence of non-alcoholic fatty liver disease (NAFLD) is increasing, as are other manifestations of metabolic syndrome such as obesity and type 2 diabetes. NAFLD is currently the number one cause of chronic liver disease worldwide. The pathophysiology of NAFLD and disease progression is poorly understood. A potential contributing role for gut microbiome and metabolites in NAFLD is proposed. Currently, bariatric surgery is an effective therapy to prevent the progression of NAFLD and other manifestations of metabolic syndrome such as obesity and type 2 diabetes. This review provides an overview of gut microbiome composition and related metabolites in individuals with NAFLD and after bariatric surgery. Causality remains to be proven. Furthermore, the clinical effects of bariatric surgery on NAFLD are illustrated. Whether the gut microbiome and metabolites contribute to the metabolic improvement and improvement of NAFLD seen after bariatric surgery has not yet been proven. Future microbiome and metabolome research is necessary for elucidating the pathophysiology and underlying metabolic pathways and phenotypes and providing better methods for diagnostics, prognostics and surveillance to optimize clinical care.
1. Introduction
Non-alcoholic fatty liver disease (NAFLD) is considered the hepatic manifestation of metabolic syndrome, next to obesity, insulin resistance and hyperlipidemia. The prevalence of obesity is increasing worldwide, as is the prevalence of NAFLD [1]. The term NAFLD comprises a spectrum of liver diseases, ranging from hepatic steatosis to non-alcoholic steatohepatitis (NASH) with or without fibrosis [2]. Progression of NASH can ultimately lead to liver cirrhosis or hepatocellular carcinoma. Moreover, NAFLD is considered to become the number one cause of chronic liver disease worldwide, with end-stage fatty liver disease as the second indication for liver transplantation [3,4,5].
Consequently, health care costs of liver disease are increasing, including costs for regular follow-up necessary to detect disease progression. The early stages of fatty liver disease can be reversible with a healthy lifestyle and weight loss. Currently, bariatric surgery is a long-term effective therapy to prevent the progression of NAFLD and other manifestations of metabolic syndrome, such as obesity and type 2 diabetes [4,6].
However, the exact pathophysiology of fatty liver disease progression remains to be clarified. The gut microbiome has been proposed as one potential contributing factor in the pathogenesis of NAFLD. The gut microbiome consists of microorganisms present in the gastrointestinal tract. The bacteria produce metabolites which are transported to the liver, where they can act as signaling molecules and have systemic effects. In addition, several studies have shown an influence of the gut microbiome on metabolism and suggest a causal role in the pathogenesis of obesity, diabetes and atherosclerotic vascular disease, which are all associated with NAFLD [7,8,9]. However, causal relations were mostly demonstrated in animal models. Translating and reproducing insights to and establishing causality in humans remains challenging.
In this review, we summarize the current evidence in humans on the changes in the gut microbiome and microbial metabolites after bariatric surgery, with a focus on the impact of gut microbiome and metabolites on NAFLD (Figure 1).
2. Gut Microbiome and Metabolites
The microbiota consists of all host microorganisms, including bacteria, protozoa, viruses and bacteriophages. The genetic material of all microorganisms is called the microbiome. The microbiome can be differentiated by location, such as the gut microbiome, oral microbiome, dermal microbiome and vaginal microbiome [10].
The gut microbiome is shaped by environmental, dietary and host factors, such as gastro-intestinal anatomy and pH [11]. Mode of delivery influences the infant gut microbiome at birth [12]. Most gut microbiota are not (directly) pathogenic, and do not cause local or systemic infection [13].
The total bacteria count in the human body is approximately 4 × 10 12, equal to the number of human cells [10]. The most abundant in the gut microbiome of average lean adults are the phyla Bacteroidetes and Firmicutes, followed by Proteobacteria, Fusobacteria and Actinobacteria [14]. The composition of the microbiome can be affected by diet, antibiotics, and other medication such as metformin and proton pump inhibitors [15].
One of the functions of the gut microbiome is to contribute to host metabolism and homeostasis. Therefore, a distinct balance between bacterial species should be maintained. Disturbance of this balance is recognized as dysbiosis and has been associated with gastrointestinal complaints and metabolic alterations. For example, in a comparison of compositions of the gut microbiome between those with normal glucose, impaired glucose, and a diabetic glucose control, differences were observed [16,17]. Methods to determine microbiome composition include 16S ribosomal RNA (16S RNA) sequencing of ubiquitous genes and whole genome shotgun sequencing of the entire gene component. Additional functional analysis of the microbiome can provide insight into the metabolic microbial capacity.
The gut microbiota are metabolically active. Ingested food is metabolized by specific microorganisms and gut microbiota are also able to excrete metabolites. These gut microbiome-derived metabolites are absorbed by enterocytes and enter the circulation where they are able to exert systemic effects (Figure 1). Some of these metabolites are thought to be the key components in influencing host metabolism [18]. Dysbiosis, the disbalance of the gut microbiome, alters (diet-derived) metabolite production and thus crosstalk between the host and gut microbiome. Research on gut microbiome-derived metabolites helps to distinguish which ones are key players in regulating metabolism at cell level [18,19].
A major class of intestinal bacteria produced metabolites are short-chain fatty acids (SCFAs). They are produced during the fermentative activity of microbiota when digesting dietary fiber, and SCFA production is suggested to modulate intestinal pH and intestinal barrier integrity [20,21,22]. The main SCFAs are propionate, butyrate and acetate, which also function as signaling molecules in immune response regulation, intestinal homeostasis and energy metabolism [23]. Other classes of metabolites include amino acids such as glycine and its precursor serine, aromatase amino acids (AAAs) including phenylalanine and tyrosine, and branched chain amino acids (BCAAs) such as leucine or valine. BCAAs promote glucose uptake and skeletal protein synthesis [24,25]. However, increased serum levels of BCAAs are seen in individuals with insulin resistance, and two bacterial species were identified as the main species driving the association between the biosynthesis of BCAAs and insulin resistance [26]. The relationship between the gut microbiome and glucose metabolism has been reviewed elsewhere [27,28].
Furthermore, the gut microbiome metabolizes dietary choline to trimethylamine (TMA), which is metabolized by the liver to trimethylamine N oxide (TMAO) [29]. Elevated TMAO has been associated with atherosclerotic vascular disease [30].
Gut microbiota conjugate primary bile acids, produced by hepatocytes, into secondary bile acids [31,32]. Bile salt hydrolase (BSH) is one of the microbial enzymes involved in driving the deconjugation of bile acids, with metabolites such as glycine and taurine as end products [33]. Besides direct involvement in fat digestion, secondary bile acids also function as signaling molecules in lipid, glucose and energy metabolism [34,35]. Most bile acids are actively reabsorbed from the intestinal lumen to the liver. This gut-to-liver axis plays a role in the enterohepatic circulation of bile acids, influencing host metabolism. The Farnesoid X receptor (FXR) is involved in bile acid synthesis control and enterohepatic circulation [36,37]. Bile acid-induced TGR5 activation decreases inflammation, and alterations of bile acid metabolism are associated with gut microbiota dysbiosis and obesity [33,38,39]. Furthermore, imbalance in bile acids is associated with gut barrier dysfunction [40].
Lipopolysaccharides (LPS) are endotoxins, components of the Gram-negative bacterial cell membrane, and play a role in immune response [41]. They can also be transported beyond the liver (that filters most of these) into the systemic circulation, where they dysregulate the inflammatory tone and contribute to metabolic disease [42].
The development of advanced sequencing techniques and the use of machine learning has increased the possibilities to gain insight in the human microbiome and derived metabolites and differences between specific populations, forming the foundation of a whole new scale of possible therapies [43,44,45]. The influence of the gut microbiome in diseases such as diabetes mellitus or inflammatory bowel disease has been intensely investigated [46,47]. Intervention studies with prebiotics, probiotics, antibiotics and fecal microbiota transplantations (FMT) focus on elucidating a potential causal relationship between gut microbiome and development as therapy for various human diseases as well.
3. Gut Microbiome and Metabolites in NAFLD
3.1. Fatty Liver Disease
As briefly mentioned before, NAFLD includes various forms of fatty liver disease and can progress to severe and irreversible liver disease (Figure 1). Fatty liver disease encompasses hepatic steatosis and non-alcoholic steatohepatitis (NASH). NAFLD is defined as lipid droplet accumulation in more than 5% of hepatocytes, and is sometimes referred to “simple” steatosis [2]. NASH, characterized by inflammation, includes steatohepatitis with and without fibrosis and requires a liver biopsy for diagnosis. NASH is histologically characterized by inflammation and hepatocyte ballooning in combination with steatosis. Histological evaluation of liver biopsies is performed via scoring systems to classify liver disease, such as the NAFLD Activity Score (NAS) and the Steatosis Activity and Fibrosis Score (SAF). Another frequently used score is the NASH Clinical Research Network (NASH CRN) scoring system: F0 = no fibrosis; F1 = perisinusoidal or portal/periportal fibrosis; F2 = perisinusoidal and portal/periportal fibrosis; F3 = bridging fibrosis; and F4 = cirrhosis. European guidelines recommend the SAF score because of its higher accuracy in distinguishing the intermediate category from mild and severe fatty liver disease [48].
It is important to realize that although NAFLD is seen as the hepatic manifestation of metabolic syndrome, not all patients with NAFLD are obese, and several patients with obesity do not have NAFLD either [49].
3.2. Gut Microbiome in Non-Alcoholic Fatty Liver Diesease
Differences in composition of the gut microbiome of patients with fatty liver disease have been observed (Table 1) [50,51]. Compared to individuals without NAFLD, fatty liver disease is associated with an increased abundance of Gram-negative microbiota: the abundance of the phylum Proteobacteria is increased [52,53]. The genera Escherichia, Enterobacteriaceae, Dorea and Peptinophilus are enriched, and abundances of Anaerosporobacter, Coprococcus, Eubacterium, and Faecalibacterium and Prevotella, Rikenellaceae and Ruminococcaceae are decreased compared to healthy controls [52,54,55,56]. Functional analyses of these microbiome differences reported increased microbial capacity for the metabolism of BCAAs and AAAs [52,54,55,56]. Furthermore, increased intestinal permeability, which is associated with dysbiosis, is seen in NAFLD [57]. Increased intestinal permeability and inflammation was observed in NAFLD patients in combination with dysbiosis of the gut microbiota: higher abundance of Escherichia was observed in fatty liver disease [58]. Development of fatty liver disease to NASH is associated with increased innate immune activation and inflammation [59,60]. The specific inflammatory factors associated with NAFLD are reviewed elsewhere [61].
Furthermore, differences between the gut microbiome of the different manifestations of fatty liver disease are also reported. One study comparing the gut microbiome found higher abundance of the phylum Firmicutes in patients with mild NAFLD compared to advanced fibrosis. The species Eubacterium rectale and Bacteroides vulgatus were the most abundant species in the mild NAFLD group. The advanced fibrosis group had higher abundance of the phylum Proteobacteria and, the most abundant species were Bacteroides vulgatus and Escherichia coli. Ruminococcus obeum CAG: 39, R. obeum, and E. rectale were significantly lower in advanced fibrosis than in mild/moderate NAFLD. A study including 203 individuals compared patients with biopsy-proven NAFLD–cirrhosis to individuals with fatty liver without advanced fibrosis, and to controls without fatty liver disease, as determined via imaging. An enriched abundance of Streptococcus was observed in both NAFLD groups. Megasphaera was only enriched in NAFLD–cirrhosis, and the highest abundance in this group was family Enterobacteriaceae and genera Streptococci and Gallibacterium. Patients with fatty liver disease without advanced fibrosis and controls had similar abundances of Bacillus and Lactococcus, which was enriched compared to NAFLD-cirrhosis. Controls had a higher abundance of Faecalibacterium prausnitzii species compared to both NAFLD groups [62]. A recent prospective study, where fatty liver status was assessed via transient elastography, found decreased relative abundance of Clostridium (sensu stricto) in patients with liver fibrosis, and an enrichment of Enterobacteriaceae, Escherichia and Shigella compared to individuals with severe steatosis without fibrosis [63].
A study comparing liver biopsies of 57 patients with NAFLD reported that increased abundance of the family Bacteroidaceae and decreased abundance of the families Prevotellacea and Erysipelotrichaceae is associated with increased disease severity. Compared to patients without NASH, NASH was associated with decreased Prevotella and increased Bacteroides abundance. Fatty liver disease with significant fibrosis (F3/F4) is associated with the increased abundance of Ruminococcus and Bacteroides, and increased abundance of Prevotella compared to mild fibrosis [64]. In children, high abundance of Prevotella copri is associated with more severe fibrosis [65]. Another study found that compared to controls, the genus Collinsella was most strongly associated with patients with NASH, and Ruminococcoaceae, a SCFA-producing genus, was decreased [66].
Certain strains, such as the Proteobacteria species Klebsiella pneumonia, are associated with endogenous alcohol production and are found to be increased in fecal microbiome of individuals with NAFLD and NASH, and abundance decreased with clinical improvement of the fatty liver disease [9]. In children, NASH was associated with an increased abundance of Proteobacteria and increased peripheral blood ethanol levels. Between obese children and children with NASH, the microbiome abundance was only statistically different for the abundance of Proteobacteria, Enterobacteriaceae and Escherichia [52].
As already mentioned, NAFLD can also develop in non-obese patients. Additionally, it appears that microbiome and metabolome signatures of non-obese patients with NAFLD differ from those of obese patients [67]; however, this requires further study.
3.3. Metabolites in Fatty Liver Disease
The liver plays a central role in metabolism and homeostasis, and the majority of blood to it is supplied via the portal vein. The portal vein drains a large part of the gastrointestinal tract, and thus the liver is the first organ to process absorbed components, such as gut microbiome-derived metabolites.
A cohort study involving obese, non-diabetic women, associated hepatic steatosis with microbiome-derived metabolite phenylacetic acid [29]. Furthermore, low microbiome gene richness was correlated with plasma BCAAs leucine, valine and isoleucine and hepatic steatosis, and the microbial capacity for metabolism of BCAA and AAAs such as phenylalanine, tyrosine and tryptophan was increased in patients with steatosis [29]. Plasma choline and phosphocholine were not found to be negatively correlated with steatosis, but urinary choline excretion was increased [29]. However, TMA (the microbial processed form of dietary choline and carnitine) and hepatic (FMO3-processed TMA) TMAO were found to be inversely correlated with steatosis [29]. In a study with 86 patients with biopsy-proven NAFLD plasma metabolites, hypoxanthine and inosine were found to be enriched in individuals with mild NAFLD. In contrast, the plasma metabolites succinate, malate, alfa-ketoglutarate, glutamine, serine and fumarate (associated with carbon metabolism) were enriched in individuals with advanced NAFLD with fibrosis [53].
Analysis of the plasma metabolome from patients with NAFLD, NASH and cirrhosis identified several metabolites as biomarkers relevant for determining disease stage. The metabolites isocitric acid and isoleucine were decreased in controls and increased with disease progression. In contrast, xanthine, glutathione and glycolic acid were found to be higher in controls and decreased with disease progression. Valine (BCAA), asparagine, propanoic acid (SCFA), butanoic acid (SCFA), phenylalanine (AAA), palmitic acid (FA), stearic acid (FA) and taurocholic acid (BA) were also identified as relevant for the metabolic signature of NAFLD; these metabolites are linked to pathways involved in bile acid, lipid, and amino acid metabolism. With increases in disease severity, taurocholic acid, phenylalanine and BCAAs increase, while glycolic acid (SCFA) and glutathione decrease [68]. Interestingly, a deficiency of serine, a precursor of glycine which is a precursor of glutathione, is associated with fatty liver disease [69,70]. In rodents, treatment with glycine improves NAFLD via glutathione synthesis [71].
SCFAs are associated with anti-inflammatory properties, in accordance with the observed decrease in NAFLD disease severity [72]. In contrast, BCAAs are associated with inflammation and insulin resistance [25,73]. Increased serum levels of BCAAs are seen in individuals with insulin resistance, and Prevotella copri and Bacteroidetes vulgatus have been identified as the main species for the association between the biosynthesis of BCAAs and insulin resistance [26]. Insulin resistance is associated with increased hepatic de novo lipogenesis, contributing to fatty liver disease [74]. Although Masarone and colleagues did not describe gut microbiome data [68], the reported metabolites such as BCAAs and AAAs, SCFAs and bile acids are in line with other studies on the gut microbiome, metabolites and NAFLD, as described in this review (Table 1).
As mentioned before, bile acids are metabolized and reabsorbed in enterohepatic circulation. Only approximately 5% of bile acids are excreted in the feces, and thus can be measured as stool metabolites. Higher serum and fecal bile acids levels are associated with advanced NAFLD and fibrosis in obese patients [75,76]. Increased serum bile acids were primary conjugated bile acids such as glycocholic acid (GCA) and secondary conjugated bile acids [75]. Increased fecal bile acids were mainly secondary unconjugated bile acids such as deoxycholic acid (DCA). Serum CGA levels and fecal DCA levels correlated with the abundance of Bacteroidaceae and Lachnospiraceae [75]. In patients with NASH, total primary bile acids were elevated and secondary bile acids were decreased [77]. In patients with fibrosis, primary bile acids were also elevated, mainly, glycine-conjugated bile acids, but secondary bile acids were the same compared to patients without fibrosis [76]. Glycogenodeoxycholic acid (GCDCA) and glycocholic acid (CGA) had strongest association with advancing fibrosis grade, as well as secondary bile acids 7-keto-deoxycholic acid (7-Keto-DCA) and glycoursodeoxycholic acid (GUDCA) [76]. Patients with borderline NASH (NAS = 3–4) had higher levels of total bile acids, total primary bile acids, and primary glycine-conjugated bile acids compared to simple steatosis (NAS = 1–2); no differences were observed between simple steatosis and definite NASH (NAS 5–8) [76]. Secondary bile acids 7-Keto-DCA and 7-Keto-lithocholic acid (7-Keto-LCA), both formed by microbial transformation, were increased in patients with definite NASH [76]. Primary bile acids, including total glycine- and taurine-conjugated bile acids, were increased in lobular inflammation and were also more increased with higher inflammation grade [76]. In steatosis, serum total cholate/chenodeoxycholate ratio is increased. Compared to steatosis or controls, in patients with NASH, total conjugated primary bile acids, conjugated/unconjugated chenodeoxycholate, cholate, and total primary bile acids are increased [77].
Different patterns of bacterial taxa-metabolites networks are observed between non-obese NAFLD and obese NAFLD [67]. In non-obese patients with worsening fibrosis severity, stool metabolites cholic acid (CA), chenodeoxycholic acid (CDCA), ursodeoxycholic acid (UDCA), glycogenodeoxycholic acid (GCDCA) and glycoursodeoxycholic acid (GUDCA) are increased. In obese individuals with significant fibrosis, LCA was significantly elevated. Of the three SCFAs, stool propionate levels gradually increased as fibrosis became more severe in non-obese patients [67].
3.4. Causality
Causality can be established with intervention studies. One of the first indicators for causality regarding gut microbiome and metabolic syndrome was when the transfer of intestinal microbiota of lean donors to patients with metabolic syndrome increased the insulin sensitivity [8,78,79]. Interestingly, metformin alters the gut microbiome of individuals with treatment-naive type 2 diabetes, probably contributing to the therapeutic effects of the drug [80]. However, most studies are cross-sectional and therefore cannot imply causality. A causal role for the gut microbiome and metabolites in the pathogenesis of NAFLD was indicated when donor feces of patients with hepatic steatosis induced steatosis after transplantation into germ-free mice. In addition, chronic treatment with phenylacetic acid also triggered steatosis [29]. FMT of human NAFLD gut microbiota into mice promoted the progression of NASH, by increasing the accumulation of intrahepatic B-cells, thus suggesting gut microbial-driven factors contributing to hepatic inflammation and fibrosis [81].
A recent publication studied the effect of FMT from vegan donors on histologic improvements of liver biopsies from patients with NAFLD. Although no significant histological changes were observed, positive changes in liver gene expression level were detected, as well as changes in metabolites and gut microbiome [82]. Another recently published study showed that donor FMT can improve liver stiffness in obese subjects [83]. Although promising, larger studies are needed to support these data.
4. Gut Microbiome and Metabolites after Bariatric Surgery and Other Weight Loss Interventions
4.1. Bariatric Surgery and Other Weight Loss Interventions
Bariatric surgery is a successful therapy for obesity and related comorbidities, resulting in permanent weight loss and improvements of metabolic and inflammatory status, such as insulin resistance and fatty liver disease [86,87,88,89]. The most performed types of surgery are laparoscopic Roux-en Y gastric bypass (RYGB) and laparoscopic sleeve gastrectomy (SG). Other surgical interventions include omega-loop gastric bypass (OAGB), biliopancreatic diversion, adjustable gastric banding (AdGB) and vertical banded gastroplasty (VGB).
4.2. Gut Microbiome in Obese Population and after Bariatric Surgery
The gut microbiome of individuals with obesity differs from that of lean individuals [90]. The gut microbiome of obese patients has increased the capacity for energy harvest [91]. Bacterial diversity and gene richness are usually decreased in patients with obesity, and microbial gene richness is inversely correlated with the severity of obesity [7]. Moreover, a small but significant negative association between microbial species alpha diversity (the species variation at individual sample level) and BMI has been reported [90].
When comparing the microbiome of lean individuals with obese individuals, the gut microbiome of obese patients was characterized by a change in relative abundance of the phyla Firmicutes compared to Bacteroidetes: Firmicutes were increased and Bacteroidetes were decreased [91]. Of the Firmicutes, abundance of the genera Rikenellaceae was increased [91]. The species Ruminococcus gnavus, Ruminococcus torques, Ruminococcus obeum, Dorea longicatena, Dorea formicigenterans, Coprococcus comes, Lachnospiraceae bacterium, Fusobacterium ulcerans and Fusobacterium varium were increased [90,92]. From the phylum Bacteroidetes, decreased species were Alistipes shahii and Alistipes senegalensis, as well as Akkermansia muciniphila, Fecalibacterium prausnitzii, and multiple species from the genus Bacteroides: Bacteroides thetaiotaomicron Bacteroides uniformis, Bacterioides xylanisolvens, Bacteroides ovatus, Bacteroides intestinalis [90,92]. A recent study found Actinomyces odontolyticus, Streptococcus australis, Streptococcus thermomphilus, Collinsella aerofaciens and Ruminococcus torques to be the most predictive bacterial species for obesity status [90]. The computational model found that 50% variance in body fat composition and BMI can be explained by the fecal microbiome.
After bariatric surgery, both anatomy and physiology rapidly adapt to the new physiological state [93,94]. The specific mechanisms of how bariatric surgery affects the composition of the gut microbiome, however, remains to be clarified. However, changes in diet habits, gastrointestinal anatomy, nutrients and gastrointestinal transit time, bile acid metabolism and gastrointestinal pH are probably all contributing factors in altering the gut microbiome [94]. Studies show that bariatric surgery increases microbial gene richness, although it still remains low and does not increase to the average richness seen in lean controls (Table 2) [7,95].
Several studies investigated the influence of RYGB on gut microbiome. In a study of 16 patients who underwent RYGB, the gut microbiome before and 3 months after surgery was analyzed. Before surgery, the phyla Firmicutes and Actinobacteria were more abundant and Verrucomicrobia was less abundant compared to lean subjects. After RYGB, the abundance of these phyla was similar to the healthy controls. Only Proteobacteria abundance was enriched after RYGB and lower in controls [96]. Thus, compared to before surgery, the phyla Firmicutes, Verrucomicrobia and Proteobacteria were increased, and the phylum Actinobacteria was decreased [96], as was Bacteroidetes [95]. At the genus level, Blautia, Roseburia, Faecalibacterium (Firmicutes) and Bifidobacterium (Actinobacteria) were decreased; however, these genera were still more abundant when compared to lean controls [96]. Furthermore, at the species level, Streptococcus spp., Akkermansia muciniphila (Verrucomicrobia), Roseburia feces, Roseburia hominis and Enterococcus faecalis were increased [96,97], and Faecalibacterium prausnitzii was decreased after RYGB [97,98]. F. prausnitzii, a butyrate producer, was associated with beneficial effects on host metabolism and negatively correlated with inflammation markers [99,100,101,102].
Studies specifically analyzing gut microbiome composition after SG found enriched abundance of the species C. comes, D. longicatena, Clostridiales bacterium, Anaerotruncus colihominis, Akkermansia muciniphila and B. thetaiotaomicron [92,103]. Relative abundance of Firmicutes, Prevotella and Bacteroides fragilis was decreased, and the abundance of species Akkermansia muciniphila, Roseburia spp., Bacteroidetes, and Bifidobacterium was increased. One year after SG, the abundance of phylum Actinobacteria was increased compared to baseline and three months post-operation [103].
After both RYGB and SG, the species Escherichia coli (Proteobacteria), Klebsiella pneumoniae (Proteobacteria) and Haemophilus parainfluenzae (Proteobacteria) were increased [97]. Abundance of aero-tolerant bacteria, such as Escherichia coli and buccal species, such as Streptococcus and Veillonella spp., was relatively higher after RYGB. In contrast, anaerobes such as Clostridium were more abundant after SG. Akkermansia muciniphila was enriched in both surgeries [97]. Functional analysis found that pathways involved in cholesterol transporters, nitrate respiration and propionate production via kinase increased in RYGB, and glutamate degradation module was more abundant in SG [97].
Long-term effects of RYGB on the gut microbial composition in seven women nine years after surgery showed enriched Gammaproteobacteria and Proteobacteria, and at the genus level, increased relative abundance of Escherichia, Klebsiella and Pseudomonas. Three species of Firmicutes, Clostridium difficile, Clostridium hiranonis and Gemella sanguinis, were lower after RYGB [104]. Functional analysis found enriched microbial genes associated with TMAO in both RYGB and ABG compared to controls, but only after RYGB increased was plasma TMAO observed.
Some patients have considerable weight regain after RYGB, and the gut microbiome may be involved. A study comparing patients at least 5 years after RYGB with and without weight regain found differences in the composition of the gut microbiome [105] In the non-regain group, higher Akkermansia genus abundance was found, compared both to the control group and weight-regain group. Compared to the control group, the non-regain group also had a higher abundance of Phascolarctobacterium genus and lower abundance of the SMB53 genus. Bacteroidetes still had a lower abundance after RYGB, both with and without weight regain compared to the control group, and in the control group genera Bacteroides and SMB53 were increased [105].
Multiple studies report changes in microbial composition after bariatric surgery. Different interventions showed different microbial profiles and only partial restoration towards lean microbiome composition. However, variations in results were observed, possibly due to different methods, small sample sizes, and/or comorbidities such as type 2 diabetes [106,107].
4.3. Plasma Metabolites after Bariatric Surgery
Plasma metabolites such as tyrosine, phenylalanine, leucine, isoleucine, valine and glutamate are all increased in patients with obesity [92]. Plasma glycine and glutamine levels are lower in obese patients [73]. Lower branched chain fatty acids (BCFAs) and higher BCAA levels are found in obese patients compared to controls [109]. Alterations of SCFA production are thought to play a contributing role in the development of obesity [20,110]. Colonic infusion with SCFA mixture increased fat oxidation and energy expenditure and decreased lipolysis in obese men [111]. Studies correlate metabolites with findings in the gut microbiome. For example, Liu et al. reported a decreased abundance of Bacteroides thetaiotaomicron, a glutamate-fermenting commensal, in Chinese patients with obesity, and this decreased abundance was inversely correlated with serum glutamate concentration [92].
As described previously, the obesity-associated gut microbiome after bariatric surgery is partially restored towards that of lean patients [92]. This phenomenon can also be observed in metabolite levels (Table 2). After both SG and RYGB, the production of AAAs and BCAAs is reduced and normalizes towards lean control levels [92]. Glycine levels relatively increase and normalize towards lean control levels [92]. In addition to the BCAAs leucine, isoleucine, and valine, the aromatic AAs phenylalanine and tyrosine, as well as ornithine, citrulline, and histidine, all decreased after RYGB. In a study comparing diet intervention with RYGB, this effect was not seen after diet intervention [108]. Furthermore, during 50 weeks of dietary weight loss, plasma SCFA levels did not change; only acetate concentration decreased with overall weight loss [112].
Correlation analysis of metabolites and clinical parameters after RYGB indicated that a cluster of metabolites, including glycine, acetylglycine, and methylmalonate, were increased and were negatively correlated with decreases in body corpulence and adipocyte diameter [7]. After RYGB and AGB, acetylglycine and glycine were negatively associated with improved weight and body composition [7]. Corresponding pathways involved in carbohydrate fermentation, the citrate cycle, glycosaminoglycan degradation, and LPS synthesis were normalized towards levels of lean controls. In patients with type 2 diabetes who underwent RYGB, plasma LPS, the endotoxin associated with insulin resistance and increased inflammatory tone, was reduced by 20% during follow-up [113]. Additionally, inflammatory stress markers decreased, and gut permeability was reduced after RYGB. Although an increase in Gram-negative, LPS-producing Proteobacteria was seen after RYGB, plasma LPS decreased, possibly due to the decreased gut permeability in combination with the increase in SCFAs, which also decreased gut permeability.
When correlated with gut microbiome composition, plasma metabolites positively associated with microbial gene richness are 3-methoxyphenylacetic acid, phloretate, hippurate, 3-hydroxphenylacetate, L-histidin, and three unidentified metabolites. Specifically, plasma glutamate is negatively correlated with microbial gene richness [7]. Changes in methylmalonate and glycine are significantly correlated with the change of Bacteroides finegoldii and Coprobacillus spp., which were also associated with improvements in body composition. Moreover, stool metabolite SCFAs acetate, propionate and butyrate were decreased after RYGB and VBG. After RYGB, the SCFA/BCFA ratio was decreased without changes in total fiber intake and with decreased protein intake, indicating that the findings were not due to dietary consumption [104].
Finally, plasma bile acids are also altered after SG and RYGB [84,114]. Especially, after RYGB, the bilio-enteric flow is altered with changes in plasma bile acid concentrations and decreases in FGF-19 [114]. Changes in bile acid profile after laparoscopic sleeve gastrectomy are associated with improvements in metabolic profile and fatty liver disease (as measured via serum cytokeratin 18) [84]. A study analyzing the gut microbiome and serum bile acids of seven patients after RYGB with controls provided evidence for an increased abundance of microbial genes involved in converting primary to secondary bile acids [104].
5. NAFLD after Bariatric Surgery
NAFLD improves following bariatric surgery, both histologically and as measured with non-invasive methods such as laboratory results and transient elastography (Table 3). In a prospective study of patients after SG and RYGB, changes in non-invasive measures of NAFLD before surgery and 1 year after surgery were measured. Improvements in the laboratory-based fibrosis score ASAT/ALAT ratio (0.8 ± 0.3 vs. 1.1 ± 0.4), NAFLD fibrosis score (− 1.0 ± 1.8 vs. − 1.7 ± 1.3), APRI score (0.3 ± 0.2 vs. 0.3 ± 0.1), and BARD score (2.3 ± 1.2 vs. 2.8 ± 1.1) were seen [115]. Liver function tests in biopsy-proven NAFLD improved in 84% of patients after RYGB and sleeve gastrectomy [116]. Liver stiffness measured by transient elastography improved from before surgery to post-operation (12.9 ± 10.4 vs. 7.1 ± 3.7 kPa), and RYGB showed more improvements to liver stiffness than SG [115].
Both SG and RYGB lead to histological improvements of NAFLD, as scored by NAS [117]. A prospective analysis on the effect of bariatric surgery on hepatic inflammation and fibrosis, as assessed in liver biopsies from 32 patients, found significant improvements in steatosis, lobular inflammation, ballooning, and fibrosis, and NASH was resolved in three out of four patients [118]. A recent large, retrospective, cohort analysis reported decreased progression of NAFLD to cirrhosis after bariatric surgery: 1.7% of the nonsurgical population and 0.5% of the bariatric surgery cohort had progression to cirrhosis [119]. In a retrospective analysis comparing RYGB with diet intervention on the potential effects on the severity and course of NASH, a 93.3% regression of NASH was shown after RYGB, as well as a 27.3% regression after diet [120]. In addition, bariatric surgery can be performed in selected patients with liver cirrhosis [121,122]. However, with more profound fibrosis, the long-term net clinical benefit compared to conservative weight loss therapy needs to be determined.
Bariatric surgery has positive effects on the clinical parameters of NAFLD; therefore, the pathophysiological mechanism has been studied intensively. However, studies focusing on evaluating the effect of microbiota of individuals before and after bariatric surgery on NAFLD are scarce. Whether gut microbiomes and metabolites contribute to the metabolic improvement and improvements of NAFLD seen after bariatric surgery has not been proven. For example, the plasma metabolite glycine is relatively decreased in individuals with obesity or NAFLD, and after bariatric surgery, it increases. Glycine levels are inversely correlated with BCAA levels. In rodent studies, glycine suppletion improved fatty liver disease [71], but suppletion did not lower elevated BCAA levels [123,124]. The glycine increase after bariatric surgery possibly contributes to the beneficial effect on NAFLD. Furthermore, a low-isoleucine diet in rodents reprogrammed liver and adipose metabolism, increasing hepatic insulin sensitivity and increasing energy expenditure [125]. However, further research in humans is necessary to validate hypotheses regarding the underlying metabolic processes of how bariatric surgery affects the pathophysiology of fatty liver disease.
As mentioned before, overlap of differences in microbial composition exists between different metabolic diseases, including NAFLD and diabetes. To determine the exact isolated microbial effect of bariatric surgery on NAFLD, including its bacterial metabolites, more research is necessary in human studies with large sample sizes to find the subtle differences between the metabolic diseases. Additionally, it is expected that adding machine learning bioinformatics will further increase insights into genes and pathways involved in the improvement of NASH after RYGB [126].
6. Discussion and Future Perspectives
Gut microbiome and plasma metabolite research is rapidly evolving, and heterogeneous new methods have been introduced. This certainly sometimes contributes to contradictory findings. Study population, chosen time points after intervention, differences in the handling of samples and storage also have an impact on study results. Validation and reproduction of results is necessary before findings can be declared as facts. Correct sample sizes with sufficient power can contribute to that. Furthermore, conflicting results and overlap of altered microbiota between metabolic diseases such as obesity, NAFLD, and type 2 diabetes are complicating factors in interpretation and in allocating findings to a distinct signature per disease: a recent review focused on microbial signatures of patients with NAFLD and the overlap with other metabolic disorders [54,67]. Thus, further research is necessary for validation and the continued comparing and interpreting of results.
Gut microbiome and metabolome research continues to evolve in two directions. First, it is evolving towards better understanding of the pathophysiology and underlying metabolic pathways of various diseases and phenotypes, using an integration of metabolome, microbiome, and other omics data [127]. For example, besides the gut microbiome, the intrahepatic microbiome and its relationship with fatty liver disease has been investigated to better understand the gut–liver axis [128,129].
Secondly, the research is evolving towards improving clinical care via determining biomarkers (serum and/or stool) for providing better methods for diagnostics, prognostics and surveillance. Furthermore, the possibilities of clinical therapies with next-generation (bacterial strain based) probiotics, albeit via fecal microbiota transplantations or oral probiotics, are explored [130]. One study showed that administration of the probiotic Akkermansia muciniphila reduces insulin resistance in overweight individuals [131]. A randomized controlled trial of patients with NAFLD showed that 1 year of administration of a synbiotic combination (probiotic and prebiotic) with Bifidobacterium animalis subspecies lactis BB-12 as a component, altered the fecal microbiome but did not reduce liver fat content or markers of liver fibrosis [132]. An RCT including patients with NAFLD scheduled for SG did not find improvements of hepatic, inflammatory or clinical outcomes when comparing the effect of generic probiotics and placebo [133]. Limitations of intervention therapies with microorganisms include the inability to control the development of the particular gut microbiome, specific abundances, or optimal ratios. Furthermore, antibiotic therapies have taught us that influencing the gut microbiome comes with adverse effects. Nowadays, the only accepted microbiome intervention to restore dysbiosis is the treatment of Clostridium difficile [134,135]. Developing therapies focused on specific receptors identified in pathophysiological studies could enable more specific interventions, with fewer uncontrolled influencing variables and thus less variance in preferred outcomes.
Although outside the scope of this review, animal studies can be used to further develop and evaluate hypotheses [136,137,138,139]. For example, a recent study in rats concluded that RYGB led to greater liver fat loss compared to a low-calorie diet, possibly due to increased fasting bile acid levels and the increased expression of modulators of liver fat oxidation, FXR and PPARα (peroxisome proliferator activator receptor alpha) [140]. Intervention with resveratrol altered the gut microbiota and improved hepatic steatosis and insulin resistance in mice [141]. Whether microbiota and their metabolites are a causal factor interacting with pathophysiological processes remains to be demonstrated. Intervention studies including FMT studies continue to explore the causality of the gut microbiome and derived metabolites in fatty liver disease [82,142].
Thus, the positive effects of bariatric surgery on NAFLD and its underlying mechanisms become more elucidated every year. Bioinformatics are used to exert insights into genes and pathways involved in the improvement of NASH after RYGB [46]. However, most patients with NAFLD will not undergo bariatric surgery. Therefore, expanding knowledge and the translation of gained insights is necessary to optimize general care. Although research can focus and reveal processes that cannot be seen by the human eye, we cannot escape the bigger picture of the impact of metabolic syndrome and NAFLD: there are limits to what health care can comprehend and support. The boundaries of resilience have become painfully clear during the ongoing COVID-19 pandemic: simple steatosis is not as benign as assumed [143].
Author Contributions
Conceptualization, J.H. and V.G; writing—original draft preparation, J.H.; writing—review and editing, V.G., M.N. and M.d.B. All authors have read and agreed to the published version of the manuscript.
Funding
J.H. is supported by an NNF GUTMMM grant (NNF15OC0016798) M.N. is supported by a ZONMW VICI grant 2020 [09150182010020].
Acknowledgments
Figure 1 was created with BioRender.com.
Conflicts of Interest
M.N. is on the Scientific Advisory Board of Caelus Health, the Netherlands, and Kaleido Biosciences, USA. None of these possible conflicts of interest bear direct relation to the content of this review. All other authors declare that they have no competing interests.
Abbreviations
FA: fatty acid; SCFA, short-chain fatty acid; BCFA, branched-chain fatty acid; AA, amino acid; BCAA, branched-chain amino acid; AAA, aromatase amino acid; TMA, trimethylamine; TMAO, trimethylamine N-oxide; BA, bile acid; LPS, lipopolysaccharides; FMT, fecal microbiome transplantation; BS, bariatric surgery; SG, sleeve gastrectomy; RYGB, Roux-en Y gastric bypass; AGB/VGB, adjustable/ventral gastric banding; FU follow-up; ASAT, aspartate aminotransferase; ALAT, alanine aminotransferase.
References
- Sayiner, M.; Koenig, A.; Henry, L.; Younossi, Z.M. Epidemiology of Nonalcoholic Fatty Liver Disease and Nonalcoholic Steatohepatitis in the United States and the Rest of the World. Clin. Liver Dis. 2016, 20, 205–214. [Google Scholar] [CrossRef]
- Sberna, A.L.; Bouillet, B.; Rouland, A.; Brindisi, M.C.; Nguyen, A.; Mouillot, T.; Duvillard, L.; Denimal, D.; Loffroy, R.; Vergès, B.; et al. European Association for the Study of the Liver (EASL); European Association for the Study of Diabetes (EASD); European Association for the Study of Obesity (EASO). EASL-EASD-EASO Clinical Practice Guidelines for the management of non-alcoholic fatty liver disease. J. Hepatol. 2016, 64, 1388–1402. [Google Scholar]
- Setiawan, V.W.; Stram, D.O.; Porcel, J.; Lu, S.C.; Le Marchand, L.; Noureddin, M. Prevalence of chronic liver disease and cirrhosis by underlying cause in understudied ethnic groups: The multiethnic cohort. Hepatology 2016, 64, 1969–1977. [Google Scholar] [CrossRef] [Green Version]
- Chalasani, N.; Younossi, Z.; LaVine, J.E.; Charlton, M.; Cusi, K.; Rinella, M.; Harrison, S.A.; Brunt, E.M.; Sanyal, A.J. The diagnosis and management of nonalcoholic fatty liver disease: Practice guidance from the American Association for the Study of Liver Diseases. Hepatology 2018, 67, 328–357. [Google Scholar] [CrossRef]
- Sayiner, M.; Younossi, Z.M. Nonalcoholic Steatohepatitis Is Becoming a Top Indication for Liver Transplantation Worldwide. Liver Transplant. 2019, 25, 10–11. [Google Scholar] [CrossRef] [Green Version]
- Syn, N.L.; Cummings, D.E.; Wang, L.Z.; Lin, D.J.; Zhao, J.J.; Loh, M.; Koh, Z.J.; Chew, C.A.; Loo, Y.E.; Tai, B.C.; et al. Association of metabolic–bariatric surgery with long-term survival in adults with and without diabetes: A one-stage meta-analysis of matched cohort and prospective controlled studies with 174 772 participants. Lancet 2021, 397, 1830–1841. [Google Scholar] [CrossRef]
- Aron-Wisnewsky, J.; Prifti, E.; Belda, E.; Ichou, F.; Kayser, B.D.; Dao, M.C.; O Verger, E.; Hedjazil, L.; Bouillot, J.-L.; Chevallier, J.-M.; et al. Major microbiota dysbiosis in severe obesity: Fate after bariatric surgery. Gut 2019, 68, 70–82. [Google Scholar] [CrossRef]
- De Groot, P.; Scheithauer, T.; Bakker, G.J.; Prodan, A.; Levin, E.; Khan, M.T.; Herrema, H.; Ackermans, M.; Serlie, M.J.M.; de Brauw’, M.; et al. Gut microbiota Donor metabolic characteristics drive effects of faecal microbiota transplantation on recipient insulin sensitivity, energy expenditure and intestinal transit time. Gut 2019, 69, 502–512. [Google Scholar] [CrossRef] [Green Version]
- Yuan, J.; Chen, C.; Cui, J.; Lu, J.; Yan, C.; Wei, X.; Zhao, X.; Li, N.; Li, S.; Xue, G.; et al. Fatty Liver Disease Caused by High-Alcohol-Producing Klebsiella pneumoniae. Cell Metab. 2019, 30, 675–688. [Google Scholar] [CrossRef]
- Sender, R.; Fuchs, S.; Milo, R. Are We Really Vastly Outnumbered? Revisiting the Ratio of Bacterial to Host Cells in Humans. Cell 2016, 164, 337–340. [Google Scholar] [CrossRef] [Green Version]
- Rothschild, D.; Weissbrod, O.; Barkan, E.; Kurilshikov, A.; Korem, T.; Zeevi, D.; Costea, P.I.; Godneva, A.; Kalka, I.N.; Bar, N.; et al. Environment dominates over host genetics in shaping human gut microbiota. Nature 2018, 555, 210–215. [Google Scholar] [CrossRef]
- Jakobsson, H.E.; Abrahamsson, T.R.; Jenmalm, M.C.; Harris, K.; Quince, C.; Jernberg, C.; Björkstén, B.; Engstrand, L.; Andersson, A.F. Decreased gut microbiota diversity, delayed Bacteroidetes colonisation and reduced Th1 responses in infants delivered by Caesarean section. Gut 2014, 63, 559–566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, Y.; Pedersen, O. Gut microbiota in human metabolic health and disease. Nat. Rev. Microbiol. 2021, 19, 55–71. [Google Scholar] [CrossRef]
- Stearns, J.C.; Lynch, M.D.J.; Senadheera, D.B.; Tenenbaum, H.C.; Goldberg, M.B.; Cvitkovitch, D.G.; Croitoru, K.; Moretno-Hagelsieb, G.; Neufeld, J.D. Bacterial biogeography of the human digestive tract. Sci. Rep. 2011, 1, 170. [Google Scholar] [CrossRef] [Green Version]
- Asnicar, F.; Berry, S.E.; Valdes, A.M.; Nguyen, L.H.; Piccinno, G.; Drew, D.A.; Leeming, E.; Gibson, R.; Le Roy, C.; Al Khatib, H.; et al. Microbiome connections with host metabolism and habitual diet from 1,098 deeply phenotyped individuals. Nat. Med. 2021, 27, 321–332. [Google Scholar] [CrossRef]
- Karlsson, F.H.; Tremaroli, V.; Nookaew, I.; Bergström, G.; Behre, C.J.; Fagerberg, B.; Nielsen, J.; Bäckhed, F.; Bäckhed, F. Gut metagenome in European women with normal, impaired and diabetic glucose control. Nature 2013, 498, 99–103. [Google Scholar] [CrossRef]
- Wang, J.; Qin, J.; Li, Y.; Cai, Z.; Li, S.; Zhu, J.; Zhang, F.; Liang, S.; Zhang, W.; Guan, Y.; et al. A metagenome-wide association study of gut microbiota in type 2 diabetes. Nature 2012, 490, 55–60. [Google Scholar]
- Krautkramer, K.A.; Fan, J.; Bäckhed, F. Gut microbial metabolites as multi-kingdom intermediates. Nat. Rev. Microbiol. 2020, 19, 77–94. [Google Scholar] [CrossRef] [PubMed]
- Tabone, M.; Bressa, C.; García-Merino, J.A.; Moreno-Pérez, D.; Van, E.C.; Castelli, F.A.; Fetnaille, F.; Lalrrosa, M. The effect of acute moderate-intensity exercise on the serum and fecal metabolomes and the gut microbiota of cross-country endurance athletes. Sci. Rep. 2021, 11, 3558. [Google Scholar] [CrossRef]
- Overby, H.B.; Ferguson, J.F. Gut Microbiota-Derived Short-Chain Fatty Acids Facilitate Microbiota:Host Cross talk and Modulate Obesity and Hypertension. Curr. Hypertens. Rep. 2021, 23, 8. [Google Scholar] [CrossRef]
- Canfora, E.E.; Jocken, J.W.; Blaak, E.E. Short-chain fatty acids in control of body weight and insulin sensitivity. Nat. Rev. Endocrinol. 2015, 11, 577–591. [Google Scholar] [CrossRef] [PubMed]
- Kelly, C.J.; Zheng, L.; Campbell, E.L.; Saeedi, B.; Scholz, C.C.; Bayless, A.J.; Wilson, K.E.; Glover, L.E.; Kominsky, D.J.; Magnuson, A.; et al. Crosstalk between microbiota-derived short-chain fatty acids and intestinal epithelial HIF augments tissue barrier function. Cell Host Microbe. 2015, 17, 662–671. [Google Scholar] [CrossRef] [Green Version]
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; Bäckhed, F. From dietary fiber to host physiology: Short-chain fatty acids as key bacterial metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef] [Green Version]
- Gibala, M.J. Protein metabolism and endurance exercise. Sports Med. 2007, 34, 337–340. [Google Scholar] [CrossRef]
- Zhou, M.; Shao, J.; Wu, C.-Y.; Shu, L.; Dong, W.; Liu, Y.; Chen, M.; Wynn, R.M.; Wang, J.; Wang, J.; et al. Targeting BCAA catabolism to treat obesity-associated insulin resistance. Diabetes 2019, 68, 1730–1746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedersen, H.K.; Gudmundsdottir, V.; Nielsen, H.B.; Hyoötylaäinen, T.; Nielsen, T.; Jensen, B.A.H.; Forslund, K.; Hildebrand, F.; Prifti, E.; Falony, G.; et al. Human gut microbes impact host serum metabolome and insulin sensitivity. Nature 2016, 535, 376–381. [Google Scholar] [CrossRef]
- Meijnikman, A.S.; Gerdes, V.E.; Nieuwdorp, M.; Herrema, H. Evaluating causality of gut microbiota in obesity and diabetes in humans. Endocr. Rev. 2018, 39, 133–153. [Google Scholar] [CrossRef] [PubMed]
- Aydin, Ö.; Nieuwdorp, M.; Gerdes, V. The Gut Microbiome as a Target for the Treatment of Type 2 Diabetes. Curr. Diabetes Rep. 2018, 18, 55. [Google Scholar] [CrossRef] [Green Version]
- Hoyles, L.; Fernández-Real, J.M.; Federici, M.; Serino, M.; Abbott, J.; Charpentier, J.; Heymes, C.; Luque, J.L.; Anthony, E.; Barton, R.H.; et al. Molecular phenomics and metagenomics of hepatic steatosis in non-diabetic obese women. Nat. Med. 2018, 24, 1070–1080. [Google Scholar] [CrossRef] [PubMed]
- Sherriff, J.L.; O’Sullivan, T.A.; Properzi, C.; Oddo, J.L.; Adams, L.A. Choline, its potential role in nonalcoholic fatty liver disease, and the case for human and bacterial genes. Adv. Nutr. 2016, 7, 5–13. [Google Scholar] [CrossRef] [Green Version]
- Wahlström, A.; Kovatcheva-Datchary, P.; Ståhlman, M.; Bäckhed, F.; Marschall, H.U. Crosstalk between Bile Acids and Gut Microbiota and Its Impact on Farnesoid X Receptor Signalling. Dig. Dis. 2017, 35, 246–250. [Google Scholar] [CrossRef] [PubMed]
- Molinero, N.; Ruiz, L.; Sánchez, B.; Margolles, A.; Delgado, S. Intestinal bacteria interplay with bile and cholesterol metabolism: Implications on host physiology. Front. Physiol. 2019, 10, 185. [Google Scholar] [CrossRef] [Green Version]
- Chiang, J.Y.L.; Ferrell, J.M. Bile acid metabolism in liver pathobiology. Gene Expr. 2018, 18, 71–87. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, T.R.; Haeusler, R.A. Bile acids in glucose metabolism and insulin signalling—mechanisms and research needs. Nat. Rev. Endocrinol. 2019, 15, 701–712. [Google Scholar] [CrossRef]
- Thomas, C.; Gioiello, A.; Noriega, L.; Strehle, A.; Oury, J.; Rizzo, G.; Macchiarulo, A.; Yamamoto, H.; Mataki, C.; Pruzanski, M.; et al. TGR5-Mediated Bile Acid Sensing Controls Glucose Homeostasis. Cell Metab. 2009, 10, 167–177. [Google Scholar] [CrossRef] [Green Version]
- Sayin, S.I.; Wahlström, A.; Felin, J.; Jäntti, S.; Marschall, H.U.; Bamberg, K.; Angelin, B.; Hyötyläinen, T.; Orešič, M.; Bäckhed, M. Gut microbiota regulates bile acid metabolism by reducing the levels of tauro-beta-muricholic acid, a naturally occurring FXR antagonist. Cell Metab. 2013, 17, 225–235. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Cai, J.; Gonzalez, F.J. The role of farnesoid X receptor in metabolic diseases, and gastrointestinal and liver cancer. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 1–13. [Google Scholar] [CrossRef]
- Kuipers, F.; Bloks, V.W.; Groen, A.K. Beyond intestinal soap—Bile acids in metabolic control. Nat. Rev. Endocrinol. 2014, 10, 488–498. [Google Scholar] [CrossRef] [PubMed]
- van Nierop, F.S.; Scheltema, M.J.; Eggink, H.M.; Pols, T.W.; Sonne, D.P.; Knop, F.K.; Soeters, M.R. Clinical relevance of the bile acid receptor TGR5 in metabolism. Lancet Diabetes Endocrinol. 2017, 5, 224–233. [Google Scholar] [CrossRef]
- Stenman, L.K.; Holma, R.; Eggert, A.; Korpela, R. A novel mechanism for gut barrier dysfunction by dietary fat: Epithelial disruption by hydrophobic bile acids. Am. J. Physiol. Gastrointest. Liver Physiol. 2013, 304, G227–G234. [Google Scholar] [CrossRef] [Green Version]
- Heianza, Y.; Ma, W.; Manson, J.A.E.; Rexrode, K.M.; Qi, L. Gut microbiota metabolites and risk of major adverse cardiovascular disease events and death: A systematic review and meta-analysis of prospective studies. J. Am. Heart Assoc. 2017, 6, e004947. [Google Scholar] [CrossRef] [PubMed]
- Cani, P.D.; Amar, J.; Iglesias, M.A.; Poggi, M.; Knauf, C.; Bastelica, D.; Neyrinck, A.M.; Fava, F.; Tuohy, K.; Chabo, C.; et al. Metabolic endotoxemia initiates obesity and insulin resistance. Diabetes 2007, 56, 1761–1772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.C.Y.; Tyler, A.D. Systematic evaluation of supervised machine learning for sample origin prediction using metagenomic sequencing data. Biol. Direct. 2020, 15, 1–12. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Ziemski, M.; Robeson, M.S.; Kaehler, B.D. Measuring the microbiome: Best practices for developing and benchmarking microbiomics methods. Comput. Struct. Biotechnol. J. 2020, 18, 4048–4062. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.S.; Spakowicz, D.J.; Hong, B.Y.; Petersen, L.M.; Demkowicz, P.; Chen, L.; Leopold, S.M.; Hanson, B.M.; Agresta, O.M.; Gerstein, M.; et al. Evaluation of 16S rRNA gene sequencing for species and strain-level microbiome analysis. Nat. Commun. 2019, 10, 5029. [Google Scholar] [CrossRef] [Green Version]
- Herrema, H.; Niess, J.H. Intestinal microbial metabolites in human metabolism and type 2 diabetes. Diabetologia 2020, 63, 2533–2547. [Google Scholar] [CrossRef]
- Pittayanon, R.; Lau, J.T.; Leontiadis, G.I.; Tse, F.; Yuan, Y.; Surette, M.; Moayyetdi, P. Differences in Gut Microbiota in Patients With vs. Without Inflammatory Bowel Diseases: A Systematic Review. Gastroenterology 2020, 158, 930–946.e1. [Google Scholar] [CrossRef]
- European Association for the Study of the Liver (EASL). EASL–EASD–EASO Clinical Practice Guidelines for the management of non-alcoholic fatty liver disease. Diabetologia 2016, 59, 1121–1140. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Jiang, X.; Cao, M.; Ge, J.; Bao, Q.; Tang, L.; Chen, Y.; Li, L. Altered fecal microbiota correlates with liver biochemistry in nonobese patients with non-alcoholic fatty liver disease. Sci. Rep. 2016, 6, 32002. [Google Scholar] [CrossRef] [PubMed]
- Shen, F.; Zheng, R.D.; Sun, X.Q.; Ding, W.J.; Wang, X.Y.; Fan, J.G. Gut microbiota dysbiosis in patients with non-alcoholic fatty liver disease. Hepatobiliary Pancreat Dis. Int. 2017, 16, 375–381. [Google Scholar] [CrossRef]
- Augustyn, M.; Grys, I.; Kukla, M. Small intestinal bacterial overgrowth and nonalcoholic fatty liver disease. Clin. Exp. Hepatol. 2019, 5, 1–10. [Google Scholar] [CrossRef]
- Zhu, L.; Baker, S.S.; Gill, C.; Liu, W.; Alkhouri, R.; Baker, R.D.; Gill, S.R. Characterization of gut microbiomes in nonalcoholic steatohepatitis (NASH) patients: A connection between endogenous alcohol and NAS.H. Hepatology 2013, 57, 601–609. [Google Scholar] [CrossRef] [PubMed]
- Loomba, R.; Seguritan, V.; Li, W.; Long, T.; Klitgord, N.; Bhatt, A.; Dulai, P.S.; Caussy, C.; Bettencourt, R.; Highlander, S.K.; et al. Gut Microbiome-Based Metagenomic Signature for Non-invasive Detection of Advanced Fibrosis in Human Nonalcoholic Fatty Liver Disease. Cell Metab. 2017, 25, 1054–1062.e5. [Google Scholar] [CrossRef] [PubMed]
- Aron-Wisnewsky, J.; Vigliotti, C.; Witjes, J.; Le, P.; Holleboom, A.G.; Verheij, J.; Nieuwdorp, M.; Clément, K. Gut microbiota and human NAFLD: Disentangling microbial signatures from metabolic disorders. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 279–297. [Google Scholar] [CrossRef] [PubMed]
- Raman, M.; Ahmed, I.; Gillevet, P.M.; Probert, C.S.; Ratcliffe, N.M.; Smith, S.; Greenwood, R.; Sikaroodi, M.; Lam, V.; Crotty, P.; et al. Fecal microbiome and volatile organic compound metabolome in obese humans with nonalcoholic fatty liver disease. Clin. Gastroenterol. Hepatol. 2013, 11, 868–875.e3. [Google Scholar] [CrossRef]
- Ruuskanen, M.O.; Åberg, F.; Männistö, V.; Havulinna, A.S.; Méric, G.; Liu, Y.; Loomba, R.; Vázquez-Baeza, Y.; Tripathi, A.; Valsta, L.M.; et al. Links between gut microbiome composition and fatty liver disease in a large population sample. Gut Microbes 2021, 13, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Miele, L.; Valenza, V.; La Torre, G.; Montalto, M.; Cammarota, G.; Ricci, R.; Mascianà, R.; Forgione, A.; Gabrieli, M.L.; Perotti, G.; et al. Increased intestinal permeability and tight junction alterations in nonalcoholic fatty liver disease. Hepatology 2009, 49, 1877–1887. [Google Scholar] [CrossRef]
- Jiang, W.; Wu, N.; Wang, X.; Chi, Y.; Zhang, Y.; Qiu, X.; Hu, a.; Li, J.; Liu, Y. Dysbiosis gut microbiota associated with inflammation and impaired mucosal immune function in intestine of humans with non-alcoholic fatty liver disease. Sci. Rep. 2015, 5, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Arrese, M.; Cabrera, D.; Kalergis, A.M.; Feldstein, A.E. Innate Immunity and Inflammation in NAFLD/NAS.H. Dig. Dis. Sci. 2016, 61, 1294–1303. [Google Scholar] [CrossRef] [Green Version]
- Fei, N.; Bruneau, A.; Zhang, X.; Wang, R.; Wang, J.; Rabot, S.; Gerard, P.; Zhao, L.P. Endotoxin producers overgrowing in human gut microbiota as the causative agents for nonalcoholic fatty liver disease. MBio 2020, 11. [Google Scholar] [CrossRef] [Green Version]
- Pan, X.; Wen, S.W.; Kaminga, A.C.; Liu, A. Gut metabolites and inflammation factors in non-alcoholic fatty liver disease: A systematic review and meta-analysis. Sci. Rep. 2020, 10, 8848. [Google Scholar] [CrossRef]
- Caussy, C.; Tripathi, A.; Humphrey, G.; Bassirian, S.; Singh, S.; Faulkner, C.; Bettencourt, R.; Rizo, E.; Richards, L.; Xu, Z.Z.; et al. A gut microbiome signature for cirrhosis due to nonalcoholic fatty liver disease. Nat. Commun. 2019, 10, 1406. [Google Scholar] [CrossRef] [PubMed]
- Lanthier, N.; Rodriguez, J.; Nachit, M.; Hiel, S.; Trefois, P.; Neyrinck, A.M.; Cani, P.D.; Bindels, L.B.; Thissen, J.-P.; Delzenne, N.M. Microbiota analysis and transient elastography reveal new extra-hepatic components of liver steatosis and fibrosis in obese patients. Sci. Rep. 2021, 11, 659. [Google Scholar] [CrossRef] [PubMed]
- Boursier, J.; Mueller, O.; Barret, M.; Machado, M.V.; Fizanne, L.; Araujo-Perez, F.; Guy, C.D.; Seed, P.C.; Rawls, J.F.; David, L.A.; et al. The severity of nonalcoholic fatty liver disease is associated with gut dysbiosis and shift in the metabolic function of the gut microbiota. Hepatology 2016, 63, 764–775. [Google Scholar] [CrossRef] [Green Version]
- Schwimmer, J.B.; Johnson, J.S.; Angeles, J.E.; Behling, C.; Belt, P.H.; Borecki, I.; Bross, C.; Durelle, J.; Goyal, N.P.; Hamilton, G.; et al. Microbiome Signatures Associated With Steatohepatitis and Moderate to Severe Fibrosis in Children With Nonalcoholic Fatty Liver Disease. Gastroenterology 2019, 157, 1109–1122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Astbury, S.; Atallah, E.; Vijay, A.; Aithal, G.P.; Grove, J.I.; Valdes, A.M. Lower gut microbiome diversity and higher abundance of proinflammatory genus Collinsella are associated with biopsy-proven nonalcoholic steatohepatitis. Gut Microbes 2020, 11, 569–580. [Google Scholar] [CrossRef]
- Lee, G.; You, H.J.; Bajaj, J.S.; Joo, S.K.; Yu, J.; Park, S.; Kang, H.; Park, J.H.; Kim, J.H.; Lee, D.H.; et al. Distinct signatures of gut microbiome and metabolites associated with significant fibrosis in non-obese NAFLD. Nat. Commun. 2020, 11, 4982. [Google Scholar] [CrossRef] [PubMed]
- Masarone, M.; Troisi, J.; Aglitti, A.; Torre, P.; Colucci, A.; Dallio, M.; Fetderico, A.; Balsano, C.; Persico, M. Untargeted metabolomics as a diagnostic tool in NAFLD: Discrimination of steatosis, steatohepatitis and cirrhosis. Metabolomics 2021, 17, 12. [Google Scholar] [CrossRef]
- Mardinoglu, A.; Agren, R.; Kampf, C.; Asplund, A.; Uhlen, M.; Nielsen, J. Genome-scale metabolic modelling of hepatocytes reveals serine deficiency in patients with non-alcoholic fatty liver disease. Nat. Commun. 2014, 5, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Mardinoglu, A.; Shoaie, S.; Bergentall, M.; Ghaffari, P.; Zhang, C.; Larsson, E.; Bäckhetd, F.; Nielsen, J. The gut microbiota modulates host amino acid and glutathione metabolism in mice. Mol. Syst. Biol. 2015, 11, 834. [Google Scholar] [CrossRef]
- Rom, O.; Liu, Y.; Liu, Z.; Zhao, Y.; Wu, J.; Ghrayeb, A.; Villacorta, L.; Fan, Y.; Chang, L.; Wang, L.; et al. Glycine-based treatment ameliorates NAFLD by modulating fatty acid oxidation, glutathione synthesis, and the gut microbiome. Sci. Transl. Med. 2020, 12, eaaz2841. [Google Scholar] [CrossRef] [PubMed]
- Coppola, S.; Avagliano, C.; Calignano, A.; Canani, R.B. The Protective Role of Butyrate against Obesity and Obesity-Related Diseases. Molecules 2021, 26, 682. [Google Scholar] [CrossRef] [PubMed]
- Newgard, C.B.; An, J.; Bain, J.R.; Muehlbauer, M.J.; Stevens, R.D.; Lien, L.F.; Haqq, A.M.; Shah, S.H.; Arlotto, M.; Slentz, C.A.; et al. A Branched-Chain Amino Acid-Related Metabolic Signature that Differentiates Obese and Lean Humans and Contributes to Insulin Resistance. Cell Metab. 2009, 9, 311–326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, G.I.; Shankaran, M.; Yoshino, M.; Schweitzer, G.G.; Chondronikola, M.; Beals, J.W.; Okunade, A.L.; Patterson, B.W.; Nyangau, E.; Field, T.; et al. Insulin resistance drives hepatic de novo lipogenesis in nonalcoholic fatty liver disease. J. Clin. Invest. 2020, 130, 1453–1460. [Google Scholar] [CrossRef]
- Adams, L.A.; Wang, Z.; Liddle, C.; Melton, P.E.; Ariff, A.; Chandraratna, H.; Tan, J.; Ching, H.; Coulter, S.; de Boer, B.; et al. Bile acids associate with specific gut microbiota, low-level alcohol consumption and liver fibrosis in patients with non-alcoholic fatty liver disease. Liver Int. 2020, 40, 1356–1365. [Google Scholar] [CrossRef] [PubMed]
- Nimer, N.; Choucair, I.; Wang, Z.; Nemet, I.; Li, L.; Gukasyan, J.; Weeks, T.L.; Alkhouri, N.; Nizar Zein, W.H.; Tang, W.; et al. Bile acids profile, histopathological indices and genetic variants for non-alcoholic fatty liver disease progression. Metabolism 2021, 116, 154457. [Google Scholar] [CrossRef]
- Puri, P.; Daita, K.; Joyce, A.; Mirshahi, F.; Santhekadur, P.K.; Cazanave, S.; A Luketic, V.; Siddiqui, M.S.; Boyett, S.; Min, H.; et al. The presence and severity of nonalcoholic steatohepatitis is associated with specific changes in circulating bile acids. Hepatology 2018, 67, 534–548. [Google Scholar] [CrossRef]
- Vrieze, A.; Van Nood, E.; Holleman, F.; Salojärvi, J.; Kootte, R.S.; Bartelsman, J.F.W.M.; Dallinga-Thie, G.M.; Ackermans, M.T.; Serlie, M.J.; Oozee, R.; et al. Transfer of intestinal microbiota from lean donors increases insulin sensitivity in individuals with metabolic syndrome. Gastroenterology 2012, 143, 913–916.e7. [Google Scholar] [CrossRef]
- Kootte, R.S.; Levin, E.; Salojärvi, J.; Smits, L.P.; Hartstra, A.V.; Udayappan, S.D.; Hermes, G.; Bouter, K.E.; Koopen, A.M.; Holst, J.J.; et al. Improvement of Insulin Sensitivity after Lean Donor Feces in Metabolic Syndrome Is Driven by Baseline Intestinal Microbiota Composition. Cell Metab. 2017, 26, 611–619.e6. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Esteve, E.; Tremaroli, V.; Khan, M.T.; Caesar, R.; Mannerås-Holm, L.; Ståhlman, M.; Olsson, L.M.; Serino, M.; Planas-Fèlix, M.; et al. Metformin alters the gut microbiome of individuals with treatment-naive type 2 diabetes, contributing to the therapeutic effects of the drug. Nat. Med. 2017, 23, 850–858. [Google Scholar] [CrossRef]
- B Barrow, F.; Khan, S.; Fredrickson, G.; Wang, H.; Dietsche, K.; Parthiban, P.; Robert, S.; Kaiser, T.; Winer, S.; Herman, A.; et al. Microbiota-Driven Activation of Intrahepatic B Cells Aggravates Nonalcoholic Steatohepatitis through Innate and Adaptive Signaling. Hepatology 2021. [Google Scholar] [CrossRef]
- Witjes, J.J.; Smits, L.P.; Pekmez, C.T.; Prodan, A.; Meijnikman, A.S.; Troelstra, M.A.; Bouter, K.E.C.; Herrema, H.; Levin, E.; Holleboom, A.G.; et al. Donor Fecal Microbiota Transplantation Alters Gut Microbiota and Metabolites in Obese Individuals with Steatohepatitis. Hepatol. Commun. 2020, 4, 1578–1590. [Google Scholar] [CrossRef] [PubMed]
- Ng, S.C.; Xu, Z.; Mak, J.W.Y.; Yang, K.; Liu, Q.; Zuo, T.; Tang, W.; Lau, L.; Lui, R.N.; Wong, S.H.; et al. Microbiota engraftment after faecal microbiota transplantation in obese subjects with type 2 diabetes: A 24-week, double-blind, randomised controlled trial. Gut 2021. [Google Scholar] [CrossRef] [PubMed]
- Belgaumkar, A.P.; Vincent, R.P.; Carswell, K.A.; Hughes, R.D.; Alaghband-Zadeh, J.; Mitry, R.R.; Le Roux, C.W.; Pael, A.G. Changes in Bile Acid Profile After Laparoscopic Sleeve Gastrectomy are Associated with Improvements in Metabolic Profile and Fatty Liver Disease. Obes. Surg. 2016, 26, 1195–1202. [Google Scholar] [CrossRef]
- Caussy, C.; Hsu, C.; Lo, M.T.; Liu, A.; Bettencourt, R.; Ajmera, V.H.; Bassirian, S.; Hooker, J.; Sy, E.; Richards, L.; et al. Link between gut-microbiome derived metabolite and shared gene-effects with hepatic steatosis and fibrosis in NAFL.D. Hepatology 2018, 68, 918–932. [Google Scholar] [CrossRef] [Green Version]
- Sjöström, L.; Narbro, K.; Sjöström, C.D.; Karason, K.; Larsson, B.; Wedel, H.; Lystig, T.; Sullivan, M.; Bouchard, C.; Carlsson, B.; et al. Effects of bariatric surgery on mortality in Swedish obese subjects. N. Engl. J. Med. 2007, 357, 741–752. [Google Scholar] [CrossRef] [Green Version]
- Adams, T.D.; Davidson, L.E.; Litwin, S.E.; Kim, J.; Kolotkin, R.L.; Nanjee, M.N.; Gutierrez, J.M.; Frogley, S.J.; Ibele, A.R.; Brinton, E.A.; et al. Weight and metabolic outcomes 12 years after gastric bypass. N. Engl. J. Med. 2017, 377, 1143–1155. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Li, L.; Chen, Y.; Lian, G.; Wang, J.; Zhang, J. Role of Gut Microbiome and Microbial Metabolites in Alleviating Insulin Resistance After Bariatric Surgery. Obes. Surg. 2021, 31, 327–336. [Google Scholar] [CrossRef]
- Yoshino, M.; Kayser, B.D.; Yoshino, J.; Stein, R.I.; Reeds, D.; Eagon, J.C.; Eckhouse, S.R.; Watrous, J.D.; Jain, M.; Knight, R.; et al. Effects of Diet versus Gastric Bypass on Metabolic Function in Diabetes. N. Engl. J. Med. 2020, 383, 721–732. [Google Scholar] [CrossRef]
- Meijnikman, A.S.; Aydin, O.; Prodan, A.; Tremaroli, V.; Herrema, H.; Levin, E.; Acherman, Y.; Bruin, S.; Gerdes, V.; Backhed, F.; et al. Distinct differences in gut microbial composition and functional potential from lean to morbidly obese subjects. J. Intern. Med. 2020, 288, 699–710. [Google Scholar] [CrossRef] [PubMed]
- Ley, R.E.; Turnbaugh, P.J.; Klein, S.; Gordon, J.I. Microbial ecology: Human gut microbes associated with obesity. Nature 2006, 444, 1022–1023. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Hong, J.; Xu, X.; Feng, Q.; Zhang, D.; Gu, Y.; Shi, J.; Zhao, S.; Liu, W.; Wang, X.; et al. Gut microbiome and serum metabolome alterations in obesity and after weight-loss intervention. Nat. Med. 2017, 23, 859–868. [Google Scholar] [CrossRef]
- Koffert, J.; Lahti, L.; Nylund, L.; Salminen, S.; Hannukainen, J.C.; Salminen, P.; De Vos, W.M.; Nuutila, P. Partial restoration of normal intestinal microbiota in morbidly obese women six months after bariatric surgery. PeerJ. 2020, 8, e10442. [Google Scholar] [CrossRef]
- Steenackers, N.; Vanuytsel, T.; Augustijns, P.; Tack, J.; Mertens, A.; Lannoo, M.; Van der Schueren, B.; Matthys, C. Adaptations in gastrointestinal physiology after sleeve gastrectomy and Roux-en-Y gastric bypass. Lancet Gastroenterol. Hepatol. 2021, 6, 225–237. [Google Scholar] [CrossRef]
- Karami, R.; Kermansaravi, M.; Pishgahroudsari, M.; Talebi, M.; Mohammadzadeh, N.; Pazouki, A. Changes in gut microbial flora after Roux-en-Y gastric bypass and sleeve gastrectomy and their effects on post-operative weight loss. Updates Surg. 2020. [Google Scholar] [CrossRef] [PubMed]
- Steinert, R.E.; Rehman, A.; Souto Lima, E.J.; Agamennone, V.; Schuren, F.H.J.; Gero, D.; Schirnier, P.; Vonlanthen, R.; Ismaeil, A.; Tzafos, S.; et al. Roux-en-Y gastric bypass surgery changes fungal and bacterial microbiota in morbidly obese patients-A pilot study. PLoS ONE 2020, 15, e0236936. [Google Scholar] [CrossRef] [PubMed]
- Farin, W.; Oñate, F.P.; Plassais, J.; Bonny, C.; Beglinger, C.; Woelnerhanssen, B.; Nocca, D.; Magloues, F.; Le Chatelier, E.; Pons, N.; et al. Impact of laparoscopic Roux-en-Y gastric bypass and sleeve gastrectomy on gut microbiota: A metagenomic comparative analysis. Surg. Obes. Relat. Dis. 2020, 16, 852–862. [Google Scholar] [CrossRef] [PubMed]
- Palleja, A.; Kashani, A.; Allin, K.H.; Nielsen, T.; Zhang, C.; Li, Y.; Brach, T.; Liang, S.; Feng, Q.; Jørgensen, N.B.; et al. Roux-en-Y gastric bypass surgery of morbidly obese patients induces swift and persistent changes of the individual gut microbiota. Genome. Med. 2016, 8, 67. [Google Scholar] [CrossRef] [Green Version]
- Barcenilla, A.; Pryde, S.E.; Martin, J.C.; Duncan, S.H.; Stewart, C.S.; Henderson, C.; Flint, H.J. Phylogenetic relationships of butyrate-producing bacteria from the human gut. Appl. Environ. Microbiol. 2000, 66, 1654–1661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez-Siles, M.; Duncan, S.H.; Garcia-Gil, L.J.; Martinez-Medina, M. Faecalibacterium prausnitzii: From microbiology to diagnostics and prognostics. ISME J. 2017, 11, 841–852. [Google Scholar] [CrossRef]
- Lenoir, M.; Martín, R.; Torres-Maravilla, E.; Chadi, S.; González-Dávila, P.; Sokol, H.; Langella, P.; Chain, F.; Bermúdez-Humarán, L.G. Butyrate mediates anti-inflammatory effects of Faecalibacterium prausnitzii in intestinal epithelial cells through Dact3. Gut Microbes. 2020, 12, 1826748. [Google Scholar] [CrossRef]
- Zhang, M.; Zhou, L.; Wang, Y.; Dorfman, R.G.; Tang, D.; Xu, L.; Pan, Y.D.; Zhou, Q.; Li, Y.; Yin, Y.Y.; et al. Faecalibacterium prausnitzii produces butyrate to decrease c-Myc-related metabolism and Th17 differentiation by inhibiting histone deacetylase 3. Int. Immunol. 2019, 31, 499–514. [Google Scholar] [CrossRef]
- Tabasi, M.; Eybpoosh, S.; Siadat, S.D.; Elyasinia, F.; Soroush, A.; Bouzari, S. Modulation of the Gut Microbiota and Serum Biomarkers After Laparoscopic Sleeve Gastrectomy: A 1-Year Follow-Up Study. Obes. Surg. 2021, 31, 1949–1956. [Google Scholar] [CrossRef] [PubMed]
- Tremaroli, V.; Karlsson, F.; Werling, M.; Ståhlman, M.; Kovatcheva-Datchary, P.; Olbers, T.; Fändriks, L.; Roux, C.W.; Nielsen, J.; Bäckhed, F. Roux-en-Y Gastric Bypass and Vertical Banded Gastroplasty Induce Long-Term Changes on the Human Gut Microbiome Contributing to Fat Mass Regulation. Cell Metab. 2015, 22, 228–238. [Google Scholar] [CrossRef] [Green Version]
- Faria, S.L.; Santos, A.; Magro, D.O.; Cazzo, E.; Assalin, H.B.; Guadagnini, D.; Vieira, F.T.; Dutra, E.S.; Saad, M.J.A.; Ito, M.K. Gut Microbiota Modifications and Weight Regain in Morbidly Obese Women After Roux-en-Y Gastric Bypass. Obes. Surg. 2020, 30, 4958–4966. [Google Scholar] [CrossRef] [PubMed]
- Murphy, R.; Tsai, P.; Jüllig, M.; Liu, A.; Plank, L.; Booth, M. Differential Changes in Gut Microbiota After Gastric Bypass and Sleeve Gastrectomy Bariatric Surgery Vary According to Diabetes Remission. Obes. Surg. 2017, 27, 917–925. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.; Shu, X.O.; Howard, E.F.; Long, J.; English, W.J.; Flynn, C.R. Fecal metagenomics and metabolomics reveal gut microbial changes after bariatric surgery. Surg. Obes. Relat. Dis. 2020, 16, 1772–1782. [Google Scholar] [CrossRef] [PubMed]
- Laferrère, B.; Reilly, D.; Arias, S.; Swerdlow, N.; Gorroochurn, P.; Bawa, B.; Bose, M.; Teixeira, J.; Stevens, R.D.; Wenner, B.R.; et al. Differential metabolic impact of gastric bypass surgery versus dietary intervention in obese diabetic subjects despite identical weight loss. Sci. Transl. Med. 2011, 3, 80re2. [Google Scholar] [CrossRef] [Green Version]
- Pakiet, A.; Wilczynski, M.; Rostkowska, O.; Korczynska, J.; Jabłonska, P.; Kaska, L.; Proczko-Stepaniak, M.; Sobczak, E.; Stepnowski, P.; Magkos, P.; et al. The Effect of One Anastomosis Gastric Bypass on Branched-Chain Fatty Acid and Branched-Chain Amino Acid Metabolism in Subjects with Morbid Obesity. Obes. Surg. 2020, 30, 304–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alhabeeb, H.; AlFaiz, A.; Kutbi, E.; AlShahrani, D.; Alsuhail, A.; AlRajhi, S.; Alotaibi, N.; Alotaibi, K.; AlAmri, S.; Alghamdi, S.; et al. Gut hormones in health and obesity: The upcoming role of short chain fatty acids. Nutrients 2021, 13, 481. [Google Scholar] [CrossRef] [PubMed]
- Canfora, E.E.; Van Der Beek, C.M.; Jocken, J.W.E.; Goossens, G.H.; Holst, J.J.; Olde Damink, S.W.M.O.; Lenaerts, K.; DeJong, C.H.C.; Blalak, E.E. Colonic infusions of short-chain fatty acid mixtures promote energy metabolism in overweight/obese men: A randomized crossover trial. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef]
- Sowah, S.A.; Hirche, F.; Milanese, A.; Johnson, T.S.; Grafetstätter, M.; Schübel, R.; Kirsten, R.; Ulrich, C.M.; Kaaks, R.; Zeller, G.; et al. Changes in plasma short-chain fatty acid levels after dietary weight loss among overweight and obese adults over 50 weeks. Nutrients 2020, 12, 452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monte, S.V.; Caruana, J.A.; Ghanim, H.; Sia, C.L.; Korzeniewski, K.; Schentag, J.J.; Dandona, P. Reduction in endotoxemia, oxidative and inflammatory stress, and insulin resistance after Roux-en-Y gastric bypass surgery in patients with morbid obesity and type 2 diabetes mellitus. Surgery 2012, 151, 587–593. [Google Scholar] [CrossRef] [PubMed]
- Eiken, A.; Fuglsang, S.; Eiken, M.; Svane, M.S.; Kuhre, R.E.; Wewer Albrechtsen, N.J.W.; Hansen, S.H.; Trammell, S.A.J.; Svenningsen, J.S.; Rehfeld, J.F.; et al. Bilio-enteric flow and plasma concentrations of bile acids after gastric bypass and sleeve gastrectomy. Int. J. Obes. 2020, 44, 1872–1883. [Google Scholar] [CrossRef] [PubMed]
- Nickel, F.; Tapking, C.; Benner, L.; Sollors, J.; Billeter, A.T.; Kenngott, H.G.; Bokhary, L.; Schmid, M.; von Frankenberg, M.; Fischer, L.; et al. Bariatric Surgery as an Efficient Treatment for Non-Alcoholic Fatty Liver Disease in a Prospective Study with 1-Year Follow-up: BariScan Study. Obes. Surg. 2018, 28, 1342–1350. [Google Scholar] [CrossRef]
- Cherla, D.V.; Rodriguez, N.A.; Vangoitsenhoven, R.; Singh, T.; Mehta, N.; McCullough, A.J.; Brethauer, S.A.; Schauer, P.R.; Aminian, A. Impact of sleeve gastrectomy and Roux-en-Y gastric bypass on biopsy-proven non-alcoholic fatty liver disease. Surg. Endosc. 2020, 34, 2266–2272. [Google Scholar] [CrossRef]
- von Schönfels, W.; Beckmann, J.H.; Ahrens, M.; Hendricks, A.; Röcken, C.; Szymczak, S.; Hampe, J.; Schalfmayer, C. Histologic improvement of NAFLD in patients with obesity after bariatric surgery based on standardized NAS (NAFLD activity score). Surg. Obes. Relat. Dis. 2018, 14, 1607–1616. [Google Scholar] [CrossRef]
- Garg, H.; Aggarwal, S.; Shalimar; Yadav, R.; Gupta, S.D.; Agarwal, L.; Agarwal, S. Utility of transient elastography (fibroscan) and impact of bariatric surgery on nonalcoholic fatty liver disease (NAFLD) in morbidly obese patients. Surg. Obes. Relat. Dis. 2018, 14, 81–91. [Google Scholar] [CrossRef]
- Wirth, K.M.; Sheka, A.C.; Kizy, S.; Irey, R.; Benner, A.; Sieger, G. Bariatric Surgery is Associated With Decreased Progression of Nonalcoholic Fatty Liver Disease to Cirrhosis. Ann. Surg. 2020, 272, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Mazzini, G.S.; Khoraki, J.; Dozmorov, M.; Browning, M.G.; Wijesinghe, D.; Wolfe, L.; Gurski, R.R.; Campos, G.M. Concomitant PPARα and FXR Activation as a Putative Mechanism of NASH Improvement after Gastric Bypass Surgery: A GEO Datasets Analysis. J. Gastrointest. Surg. 2019, 23, 51–57. [Google Scholar] [CrossRef]
- Rebibo, L.; Gerin, O.; Verhaeghe, P.; Dhahri, A.; Cosse, C.; Regimbeau, J.M. Laparoscopic sleeve gastrectomy in patients with NASH-related cirrhosis: A case-matched study. Surg. Obes. Relat. Dis. 2014, 10, 405–410. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.; Pouwels, S.; Parmar, C.; Kassir, R.; de Luca, M.; Graham, Y.; Mahawar, K.; for the Global Bariatric Research Collaborative. Outcomes of Bariatric Surgery in Patients with Liver Cirrhosis: A Systematic Review. Obes. Surg. 2021, 31, 2255–2267. [Google Scholar] [CrossRef] [PubMed]
- Glynn, E.L.; Piner, L.W.; Huffman, K.M.; Slentz, C.A.; Elliot-Penry, L.; Abouassi, H.; White, P.; Bain, J.R.; Muehlbauer, M.J.; Ilkayeva, O.R.; et al. Impact of combined resistance and aerobic exercise training on branched-chain amino acid turnover, glycine metabolism and insulin sensitivity in overweight humans. Diabetologia 2015, 58, 2324–2335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, P.J.; Lapworth, A.L.; McGarrah, R.W.; Kwee, L.C.; Crown, S.B.; Ilkayeva, O.; An, J.; Carson, M.W.; Christopher, B.A.; Ball, J.R.; et al. Muscle-Liver Trafficking of BCAA-Derived Nitrogen Underlies Obesity-Related Glycine Depletion. Cell Rep. 2020, 33, 108375. [Google Scholar] [CrossRef]
- Yu, D.; Richardson, N.E.; Green, C.L.; Spicer, A.B.; Murphy, M.E.; Flores, V.; Jang, C.; Kasza, I.; Nikodemova, M.; Wakai, M.H.; et al. The adverse metabolic effects of branched-chain amino acids are mediated by isoleucine and valine. Cell Metab. 2021, 33, 905–922.e6. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Zhou, Y.; Wu, Z.; Li, Y.; Zhou, W.; Wang, Y. Integrated Analysis of Key Genes and Pathways Involved in Nonalcoholic Steatohepatitis Improvement After Roux-en-Y Gastric Bypass Surgery. Front. Endocrinol. 2021, 11, 611213. [Google Scholar] [CrossRef] [PubMed]
- Van Olden, C.C.; Van de Laar, A.W.; Meijnikman, A.S.; Aydin, O.; Van Olst, N.; Hoozemans, J.B.; De Brauw, L.M.; Bruin, S.C.; Acherman, Y.I.Z.; Verheij, J.; et al. A Systems Biology approach to understand gut microbiota and host metabolism in morbid obesity: Design of the BARIA Longitudinal Cohort Study. J. Intern. Med. 2020. [Google Scholar] [CrossRef]
- Sookoian, S.; Salatino, A.; Castaño, G.O.; Landa, M.S.; Fijalkowky, C.; Garaycoechea, M.; Pirola, G.J. Intrahepatic bacterial metataxonomic signature in non-alcoholic fatty liver disease. Gut 2020, 69, 1483–1491. [Google Scholar] [CrossRef] [PubMed]
- Tilg, H.; Burcelin, R.; Tremaroli, V. Liver tissue microbiome in NAFLD: Next step in understanding the gut-–liver axis? . Gut 2020, 69, 1373–1374. [Google Scholar] [CrossRef]
- Verdier, C.; Denis, S.; Gasc, C.; Boucinha, L.; Uriot, O.; Delmas, D.; Dore, J.; Le Camus, C.; Schwintner, C.; Blanquet-Diot, S. An Oral FMT Capsule as Efficient as an Enema for Microbiota Reconstruction Following Disruption by Antibiotics, as Assessed in an In Vitro Human Gut Model. Microorganisms 2021, 9, 358. [Google Scholar] [CrossRef]
- Depommier, C.; Everard, A.; Druart, C.; Plovier, H.; Van Hul, M.; Vieira-Silva, S.; Falony, G.; Raes, J.; Maiter, D.; Delzenne, N.; et al. Supplementation with Akkermansia muciniphila in overweight and obese human volunteers: A proof-of-concept exploratory study. Nat. Med. 2019, 25, 1096–1103. [Google Scholar] [CrossRef]
- Scorletti, E.; Afolabi, P.R.; Miles, E.A.; Smith, D.E.; Almehmadi, A.; Alshathry, A.; Childs, C.E.; Del Fabbro, S.; Bilson, J.; Moyses, H.E.; et al. Synbiotics Alter Fecal Microbiomes, But Not Liver Fat or Fibrosis, in a Randomized Trial of Patients with Nonalcoholic Fatty Liver Disease. Gastroenterology 2020, 158, 1597–1610.e7. [Google Scholar] [CrossRef] [PubMed]
- Sherf-Dagan, S.; Zelber-Sagi, S.; Zilberman-Schapira, G.; Webb, M.; Buch, A.; Keidar, A.; Sakran, N.; Goitein, D.; Goldenberg, N.; Mahdi, J.A.; et al. Probiotics administration following sleeve gastrectomy surgery: A randomized double-blind trial. Int. J. Obes. 2018, 42, 147–155. [Google Scholar] [CrossRef]
- Taur, Y.; Pamer, E.G. Harnessing microbiota to kill a pathogen: Fixing the microbiota to treat Clostridium difficile infections. Nat. Med. 2014, 20, 246–247. [Google Scholar] [CrossRef]
- Tixier, E.N.; Verheyen, E.; Luo, Y.; Grinspan, L.T.; Du, C.H.; Ungaro, R.C.; Walsh, S.; Grinspan, A.M. Systematic Review with Meta-Analysis: Fecal Microbiota Transplantation for Severe or Fulminant Clostridioides difficile. Dig. Dis. Sci. 2021. [Google Scholar] [CrossRef] [PubMed]
- Bozadjieva Kramer, N.; Evers, S.S.; Shin, J.H.; Silverwood, S.; Wang, Y.; Burant, C.F.; Sandoval, D.A.; Seeley, R.J. The Role of Elevated Branched-Chain Amino Acids in the Effects of Vertical Sleeve Gastrectomy to Reduce Weight and Improve Glucose Regulation. Cell Rep. 2020, 33, 108239. [Google Scholar] [CrossRef] [PubMed]
- Chaudhari, S.N.; Luo, J.N.; Harris, D.A.; Aliakbarian, H.; Yao, L.; Paik, D.; Subramaniam, R.; Adhikari, A.A.; Vernon, A.H.; Kiliç, A.; et al. A microbial metabolite remodels the gut-liver axis following bariatric surgery. Cell Host Microbe. 2021, 29, 408–424.e7. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.; Li, Y.; Wang, Y.; Jin, L.; Yang, L.; Wang, L.; Liao, J.; Wang, H.; Peng, Y.; Zhang, Z.; et al. Da-Chai-Hu Decoction Ameliorates High Fat Diet-Induced Nonalcoholic Fatty Liver Disease Through Remodeling the Gut Microbiota and Modulating the Serum Metabolism. Front. Pharmacol. 2020, 11, 584090. [Google Scholar] [CrossRef] [PubMed]
- Littmann, E.R.; Lee, J.J.; Denny, J.E.; Alam, Z.; Maslanka, J.R.; Zarin, I.; Matsuda, R.; Carter, R.A.; Susac, B.; Saffern, M.S.; et al. Host immunity modulates the efficacy of microbiota transplantation for treatment of Clostridioides difficile infection. Nat. Commun. 2021, 12, 755. [Google Scholar] [CrossRef]
- Mazzini, G.S.; Khoraki, J.; Browning, M.G.; Wu, J.; Zhou, H.; Price, E.T.; Wolfe, L.G.; Mangino, M.J.; Campos, G.M. Gastric Bypass Increases Circulating Bile Acids and Activates Hepatic Farnesoid X Receptor (FXR) but Requires Intact Peroxisome Proliferator Activator Receptor Alpha (PPARα) Signaling to Significantly Reduce Liver Fat Content. J. Gastrointest. Surg. 2021, 25, 871–879. [Google Scholar] [CrossRef]
- Du, F.; Huang, R.; Lin, D.; Wang, Y.; Yang, X.; Huang, X.; Zheng, B.; Chen, Z.; Huang, Y.; Wang, X.; et al. Resveratrol Improves Liver Steatosis and Insulin Resistance in Non-alcoholic Fatty Liver Disease in Association With the Gut Microbiota. Front. Microbiol. 2021, 12, 611323. [Google Scholar] [CrossRef] [PubMed]
- Burz, S.; Monnoye, M.; Philippe, C.; Farin, W.; Ratziu, V.; Strozzi, F.; Paillarse, J.-M.; Chêne, L.; Blottière, H.; Gérard, P. Fecal Microbiota Transplant from Human to Mice Gives Insights into the Role of the Gut Microbiota in Non-Alcoholic Fatty Liver Disease (NAFLD). Microorg. 2021, 9, 199. [Google Scholar] [CrossRef]
- Meijnikman, A.S.; Bruin, S.; Groen, A.K.; Nieuwdorp, M.; Herrema, H. Increased expression of key SARS-CoV-2 entry points in multiple tissues in individuals with NAFLD. J. Hepatol. 2020, 74, 748. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Overview of the gut microbiome and metabolites, and relationship with NAFLD and bariatric surgery. (A) A systematic overview of how gut microbiota contribute to producing metabolites such as LPS, bile acids, SCFAs and trimethylamine. The metabolites enter the portal circulation, where they are further metabolized and enter the systemic circulation. (B) The spectrum of non-alcoholic fatty liver disease. The black arrow indicates that the different stages of NAFLD are associated with a different composition of gut microbiome and plasma metabolites. The smaller grey arrow indicates the possible relationship of gut microbiome composition and plasma metabolites on non-alcoholic fatty liver disease; however, this causal role remains to be verified. (C) Three types of bariatric interventions are displayed: sleeve gastrectomy, Roux-en-Y gastric bypass, and vertical gastric banding. Bariatric surgery is associated with changes in gut microbiome composition and altered metabolite levels. Bariatric surgery is also associated with improvements of fatty liver disease. (D) Different gut microbiota compositions and relative changes in metabolites are observed in individuals with NAFLD and after bariatric surgery compared to (lean and/or pre-operative) controls. Evidence points toward a causal role for microbiome and metabolites in the pathophysiology of fatty liver disease and improvement after bariatric surgery; however, this remains to be verified. LPS, lipopolysaccharides; SCFAs, short-chain fatty acids; ATP, adenosine triphosphate; NAFLD, non-alcoholic fatty liver disease; NASH, non-alcoholic steatohepatitis.
Figure 1.
Overview of the gut microbiome and metabolites, and relationship with NAFLD and bariatric surgery. (A) A systematic overview of how gut microbiota contribute to producing metabolites such as LPS, bile acids, SCFAs and trimethylamine. The metabolites enter the portal circulation, where they are further metabolized and enter the systemic circulation. (B) The spectrum of non-alcoholic fatty liver disease. The black arrow indicates that the different stages of NAFLD are associated with a different composition of gut microbiome and plasma metabolites. The smaller grey arrow indicates the possible relationship of gut microbiome composition and plasma metabolites on non-alcoholic fatty liver disease; however, this causal role remains to be verified. (C) Three types of bariatric interventions are displayed: sleeve gastrectomy, Roux-en-Y gastric bypass, and vertical gastric banding. Bariatric surgery is associated with changes in gut microbiome composition and altered metabolite levels. Bariatric surgery is also associated with improvements of fatty liver disease. (D) Different gut microbiota compositions and relative changes in metabolites are observed in individuals with NAFLD and after bariatric surgery compared to (lean and/or pre-operative) controls. Evidence points toward a causal role for microbiome and metabolites in the pathophysiology of fatty liver disease and improvement after bariatric surgery; however, this remains to be verified. LPS, lipopolysaccharides; SCFAs, short-chain fatty acids; ATP, adenosine triphosphate; NAFLD, non-alcoholic fatty liver disease; NASH, non-alcoholic steatohepatitis.


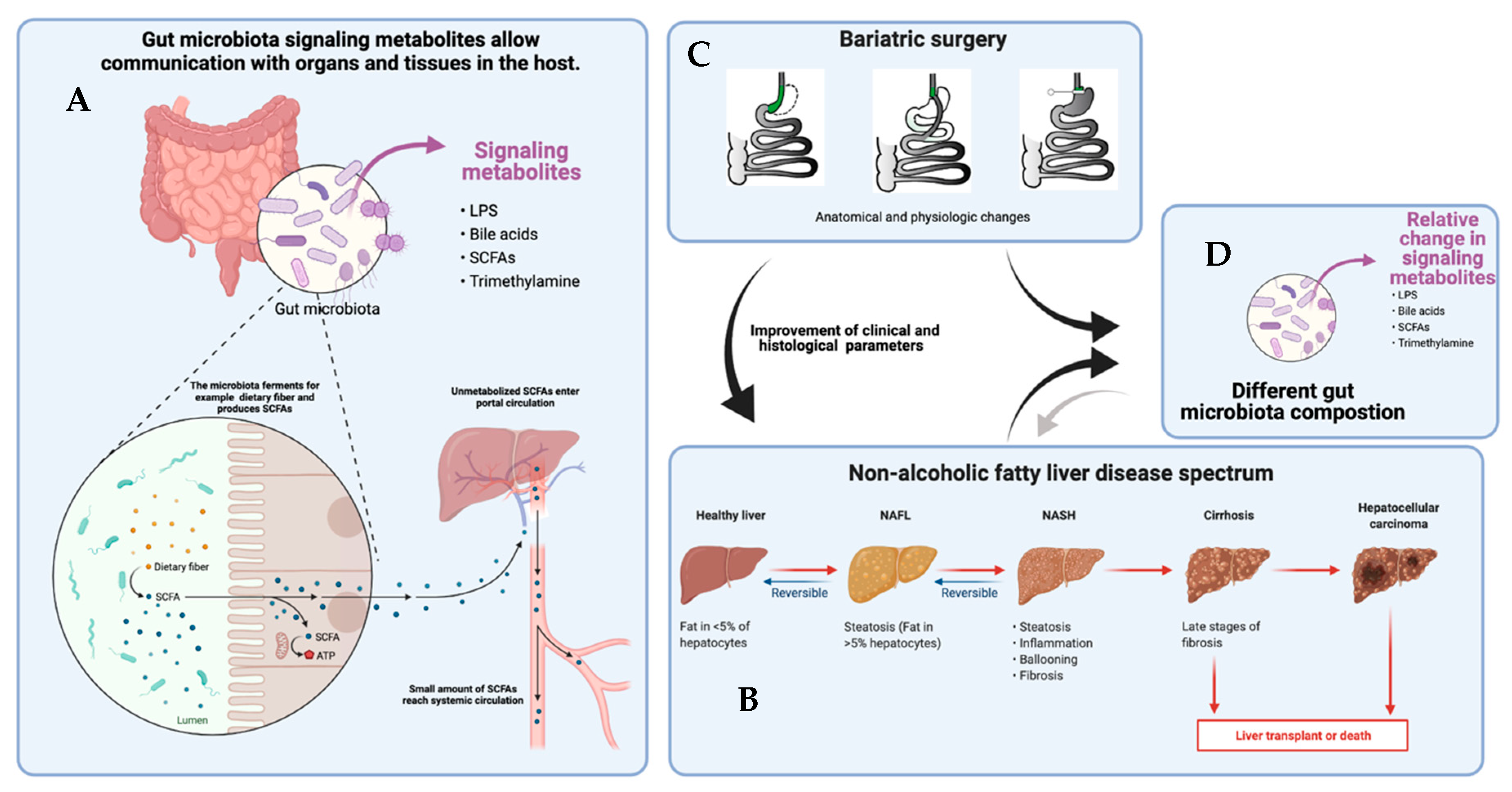{kind=link}
Table 1.
NAFLD, gut microbiome and metabolites.
| Author, Year | Study Descriptive (Population; Follow-up; Weight Loss) | Method | Microbiome | Metabolites |
|---|---|---|---|---|
| Belgaumkar, 2016 [84] | Prospective analysis; SG (n = 18) NAFLD defined by serum cytokeratin 18 (n = 14, 78% NAFL) FU 6 months; TWL-39.9 kg | Bile acids: LC/MS | Not described. | ↑ primary glycine- and ↑ taurine-conjugated BA, ↓ cholic acid decreased, and ↑ secondary BA, ↑ glycine-conjugated urodeoxycholic acid No change in total BA. |
| Boursier, 2016 [64] | Biopsy-proven NAFLD (n = 57) F0/FI n = 30 vs. F3/F4 n = 27 | Fecal microbiome: 16 S RNA sequencing analysis | Increased NAFLD severity: ↑ Bacteroidaceae, ↓ Prevotellacea; ↓ Erysipelotrichaceae. NASH (compared to no NASH): ↑ Bacteroides; ↓ Prevotella Significant fibrosis (F3/4) compared to F0/F1: ↑ Bacteroides; ↑ Ruminococcus; ↓ Prevotella. | Not described. |
| Loomba, 2017 [53] | Prospective analysis biopsy-proven NAFLD (n = 86): comparison mild/moderate (n = 72;) vs. advanced fibrosis (n = 14); | Fecal microbiome: whole-genome shotgun sequencing | NAFLD—mild/moderate: ↑ abundance Firmicutes; most abundant Eubacterium rectale, Bacteroides vulgates NAFLD-AF: ↑ abundance Proteobacteria; most abundant B. vulgates, Escherichia coli. ↓ Ruminococcus obeum CAG:39; R. obeum; E. rectale. | NAFLD—mild/moderate: serum: ↑ Hypoxanthine, ↑ Inosine; Stool: ↑ L-lactate; ↑ Acetate ↑ formate; NAFLD-AF: serum: ↑ Succinate; ↑ Malatae; ↑ alfa-ketoglutarate; ↑ Serine; ↑ Glutamine; ↑ Fumarate; ↑ Glutamate; ↑ Lactate; stool: ↑ butyrate, D-lactate, propionate, succinate |
| Caussy, 2018 [85] | Cross-sectional analysis twin family cohort, n = 156 validation cohort, n = 156 hepatic steatosis, n = 57 | Fecal microbiome: whole-genome shotgun metagenomic sequencing; Liver: MRI/MRE; Metabolites CG/MS and LC/MS/MS | Proteobacteria, Firmicutes, Bacteroidetes correlated with 3-(4-hydroxyphenyl)lactate and phenyllactate. | 6 microbial origins: 3-(4-hydroxyphenyl)lactate, N-formylmethionine, phenyllactate, mannitol, allantoine, N-(2-furoyl)glycine. 3-(4-hydroxyphenyl)lactate gut microbiome-linked metabolite assosated with liver fibrosis. |
| Caussy, 2019 [62] | Cross-sectional; n = 203 NAFLD-cirrhosis, NAFLD, without advanced fibrosis non-NAFLD controls | Fecal microbiome: 16S RNA sequencing analysis Liver: MRI/MRE. | NAFLD–cirrhosis: ↑ Streptococcus; ↑ Megashaera; most enriched abundance of family Enterobacteriaceae, genera Streptococci and Gallibacterium. NAFLD-AF: ↑ Streptococcus; ↑ Bacillus; ↑ Lactococcus Non-NAFLD: ↑ Bacillus; ↑ Lactococcus; ↑ Pseudomonas; ↑ Faecalibacterium prausnitzii, ↑ genus Catenibacterium; families ↑ Rikenellaceae, ↑ Mogibacterium, ↑ Peptostreptococcaceae | Not described. |
| Puri, 2018 [77] | Cross-sectional analysis biopsy-proven NAFLD and bile acids; n = 86 (controls n = 24. NAFL n = 25; NASH n = 37; BMI 31.9) | LC/MS | Not described. | NASH: ↑ total primary BAs; ↓ secondary BAs. NASH vs. NAFL, vs. controls: ↑ Total conjugated primary BAs ↑ conjugated/unconjugated chenodeoxycholate; ↑ cholate; ↑ total primary BAs. NAFL: ↑ Total cholate/chenodeoxycholate ratio ↑ total secondary/primary BA ratio -> ↓ likelihood of significant fibrosis (F ≥ 2) ↑ conjugated cholate -> ↑ likelihood of significant fibrosis (F ≥ 2). |
| Hoyles, 2018 [29] | Prospective analysis; obese women n= 105; liver biopsy (histology), NAFLD (n = 56); fecal microbiome (n =56, | Fecal microbiome: shotgun metagenomic sequencing; serum and urine Metabolites: LC/MS | Steatosis: ↑ Proteobacteria, ↑ Actinobacteria, Verrucomicrobia ↑ correlated Firmicutes and Euryarchaeta ↓ correlated. Species: ↑ Acidaminococcus, ↑ Escherichia; ↓ Lachnospiraceae, ↓ Ruminococcaceae Functional analysis: ↑ LPS and peptidoglycan biosynthesis. | Steatosis: Serum BCAAs: ↑ leucine, ↑ valine, ↑ isoleucine. ↑ phenylacetic acid (PAA) Urine: ↑ choline No-NAFLD: ↑ acetate; ↑ TMAO |
| Lee, 2020 [67] | Prospective analysis Non-obese NAFLD | Fecal metabolites: 16S RNA sequencing analysis | Elevated Ruminococcaceae and Veillonellaceae associated with fibrosis severity. | Fecal metabolites: bile acids and propionate elevated (especially with significant fibrosis). |
| Adams, 2020 [75] | Prospective analysis liver biopsy n = 122 (as part of clinical care or during bariatric surgery) | Fecal microbiome: 16S RNA sequencing analysis Metabolies: serum + fecal BA analysis: LCMS. | NAFLD-AF (F3/4): ↑ Firmicutes, ↑ Proteobacteria; ↑ Actinobacteria; ↓ Bacteriodetes. Family: ↑ Actinomycetaceae; ↑ Lachnospiraceae; ↓ Bacteroidaceae; ↓ unclassifiable of order Bacteroidales. | Progressive ↑ total serum BAs from controls, F0–2 NAFLD to F3/4 NAFLD. ↑ GCA (glycocholic acid); ↑ GDCA (glycodeoxycholic acid) Fecal BA: ↑ DCA (deoxycholic acid); ↑ LC (lithocholic acid). |
| Masarone, 2021 [68] | Cross-sectional analysis cohort biopsy-proven NAFLD n = 144 steatosis, n = 76, NASH n = 23, cirrhosis, n = 43 (NASH–cirrhosis n = 15, HCV n = 8, cryptogenic n = 20) | Serum metabolites GC/MS; machine learning model. | Not described. | Lower in controls and increase with disease progression: isocitric acid, isoleucine, not identified metabolite Higher in controls and decrease with disease progression: xanthine, glutathione, glycolic acid Valine, asparagine, 4-deoxy erythronic acid, propanoic acid, palmitic acid, butanoic acid, stearic acid, phenylalanine, taurocholic acid NASH-related cirrhosis, increased concentration of galactose, uric acid, glyceric acid, butanoic acid, histidine, phenylalanine, stearic acid, threonine and palmitic acid |
| Nimer, 2021 [76] | Prospective analysis; NAFLD n = 102 (30% simple steatosis, 43% borderline NASH, 27% NASH); controls n = 50 Liver biopsy; BMI 32.8 kg/m2 | Plasma bile acid profile: quantitative stable isotope dilution LC/MS/MS | Not described. | NAFLD vs. controls: ↑ almost all circulating BAs Fibrosis vs. NAFLD: ↑ glycine-conjugated primary BAs (↑ GCDCA, ↑ GCA), secondary BAs ↑ 7-keto-DCA, ↑ GUDCA NASH vs. simple steatosis: ↑ 7-keto-DCA, ↑ 7-keto, LCA |
Table 2.
Bariatric surgery, gut microbiome and metabolites.
| First author, year [Ref.] | Study Descriptive (Population; Follow-up; Weight Loss) | Method | Results Microbiome | Results Metabolites |
|---|---|---|---|---|
| Laferrere, 2011 [108] | Prospective analysis; diabetic patients; RYGB, n = 10 Diet intervention, n = 11 FU 1 month | Plasma metabolites: MS | Not described. | RYGB: ↓ BCAAs: ↓ leucine, ↓ isoleucine, ↓ valine; ↓ aromatic AAs: ↓ phenylalanine ↓ tyrosine; ↓ ornithine, ↓ citrulline, ↓ histidine. Diet: no differences |
| Tremaroli, 2015 [104] | Post-bariatric surgery, long-term effect: RYGB (n = 7) vs. VGB (n = 7); matched controls (severe obesity n = 7); FU 9.4 years | Fecal microbiome: shotgun metagenomic sequencing Metabolites: GC/MS, UPLC-MS-MS | RYGB vs. obese controls: ↑ Gammaproteobacteria; ↓ Firmicutes (↓ Clostridium difficile, ↓ Clostridium hiranonis, ↓ Gemella sanguinis); ↑ Proteobacteria (↑ Escherichia, ↑ Klebsiella, ↑ Pseudomonas) No significant differences VBG and controls. No significant differences VBG and RYGB. | RYGB, VBG: ↓SCFAs (↓ acetate, ↓ propionate, ↓ butyrate) BCFA (isobutryrate, isovaleratie): no change. RYGB: SCFA/BCFA ratio decreased |
| Palleja, 2016 [98] | Prospective analysis; RYGB, n = 13 FU 3 months, n = 12 12 months, n = 8 | Fecal microbiome: Shotgun metagenomic sequencing | Microbial diversity: ↑ 3 months and ↑ 1 year. ↑ Proteobacteria, ↑ Fusobacteria ↑ Escherichia coli, ↑ Klebsiella pneumoniae, ↑ Veillonella spp., ↑ Streptococcus spp., ↑ Alistipes spp., and ↑ Enterococcus faecalis; ↑ Bifidobacterium dentium; ↑ Fusobacterium nucleatum; ↑ Akkermansia muciniphila; ↓ Faecalibacterium prausnitzii | Not described. |
| Liu, 2017 [92] | Prospective analysisSG, n = 23 obese individuals, n = 72 lean controls, n = 79 FU 1 and 3 months. | Shotgun metagenomic sequencing; serum plasma metabolites: LC/MS | ↑ : C. comes; D. longicatena; Clostridiales bacterium; Anaerotruncus colihominis; Akkermansia muciniphila; B. thetaiotaomicron. | ↓ : Aromatic amino acids ↓; methionine ↓; alanine ↓; lysine ↓; serine ↓; glutamate ↓ decreased ↑ : Acetylglycine ↑; glycine ↑increased. |
| Aron, 2019 [7)] | Prospective analysisRYGB, n = 41 gastric banding/AGB, n = 20) n = 24 had post-operative follow-up. | Fecal microbiome: shotgun metagenomic sequencing Serum metabolites: LC/MS | RYGB: ↑ Oscillibacter; ↑ Clostridium sp; ↑ Alistipes shahii; ↑ Butyricimonas; ↑ Fusobacterium nucleatum; ↑ Roseburia; ↑ Dialister sp; ↑ Butyricimonas virosa; ↑ Hungatella hatewayi; ↓ Coprobacillus sp.; ↓ Anaerostipes hadrus (butyrate producer)AGB: ↑ Oscillibacter; ↑ Butyricimonas virosa; ↑ Bacteroides finegoldii | Metabolites associated with microbial gene richness (MGR): glutamate, negatively correlated; 3-methoxyphenylacetic acid, phloretate, hippurate, 3-hydroxphenylacetate, L-histidin and three unidentified positively correlated.. RYGB: ↑ glycine, ↑ acetylglycine, ↑ methylmalonate After BS: - ↑ Acetlyglycine and ↑ glycine: - ↓ BCAA when insulin resistance ↑ |
| Steinert, 2020 [96] | Prospective n= 25, RYGB, n = 16 (RYGB), Controls, n= 9 FU 3 months, not described | Fecal microbiome: 16S RNA sequencing analysis | RYGB vs. pre-operation: ↓ Blautia, ↓ Roseburia and ↓ Faecalibacterium (Firmicutes); compared to controls more abundant RYGB: ↓ Bifidobacterium, which was increased compared to lean controls | Not described. |
| Farin, 2020 [97] | RYGB (n = 89) SG (n= 108) FU 6 months; not described | Fecal microbiome: shotgun metagenomic sequencing | Phylum: 15% gene enrichment: ↑ Bacteroidetes, ↑ Proteobacteria, ↑ Actinobacteria, ↑ Verrucomicrobia; ↓ Firmicutes (Firmicutes slight decrease after surgeries).Both: ↑ A. muciniphila; ↑ E. coli, ↑ H. parainfluenzae, ↑ Klebsiella pneuoniae. RYGB > SG: ↑ E. coli and ↑ K. pneumoniae, ↑ R. faecis and ↑ R. hominis, ↑ E. faecalis more enriched by RYGB than SG. ↓ Faecalibacterium preusnitzii less abundant after RYGB. SG > RYGB: ↑ A. hadrus, C. sp KLE, F. plautii, O. sp. KLE, R. gnavus (Firmicutes order Clostridiales) | Not described. |
| Karami, 2020 [95] | Prospective analysis n = 30 RYGB, n = 12 SG, n = 18 FU 6 months; EWL 57.72 ± 15.08% | Fecal microbiome: 16S RNA sequencing analysis | RYGB: Bacteroidetes ↓ compared to pre-surgery. RYGB or SG: no changes in Firmicutes count. Firmicutes to Bacteroidetes ratio not different from baseline; B. fragilis count not different to baseline or between groups. BS (RYGB + SG): ↓ Bacteroidetes; ↑ Firmicutes/Bacteroidetes ratio. | Not described. |
| Faria, 2020 [105] | Retrospective analysis; RYGB: with vs. without weight regain, vs. control; FU > 5 year post-operative; TWL% 25.8 vs. 33.1. | Fecal microbiome: 16S RNA sequencing analysis. | RYGB, non-regain vs. control and weight regain: ↑ Akkermansia genus. RYGB, non-regain vs. control: ↑ Phascolarctobacterium genus and ↓ SMB53 genus. RYGB vs. control: ↓ Bacteroidetes Control vs. RYGB: ↑ Bacteroides, ↑ SMB53 | Not described. |
| Pakiet, 2020 [109] | Prospective analysis OAGB, n = 50 lean controls, n = 32 | Serum metabolites GM/LM/MS | Not described. | Baseline vs. controls: ↓BCFAs; ↑ BCAAs OAGB: ↑ BCFA; ↓BCAA. IR correlated inversely with BCFAs and positively with BCAAs |
| Tabasi, 2021 [103] | Prospective analysis SG, n = 126 FU 3 and 12 months | Fecal microbiome: qPCR | M3: ↓ Prevotella; ↓ Bacteroides fragilis group; ↓ Firmicutes spp; ↑ Akkermansia muciniphila; ↑ Roseburia spp.; ↑ Bacteroidetes; ↑ Bifidobacterium M12: ↑ Actinobacteria (compared to M3 and baseline) | Not described. |
Table 3.
Bariatric surgery and NAFLD.
| Author, Year [Ref.] | Study Design | Parameters to Assess Liver Disease | Changes/Outcomes |
|---|---|---|---|
| Nickel, 2018 [115] | Prospective analysis, n = 100 SG vs. RYGB FU 1 year. | Transient elastography: liver stiffness Laboratory-based fibrosis score: AST/ALT ratio; NAFLD fibrosis score; APRI score; BARD score | 12.9 ± 10.4 vs. 7.1 ± 3.7 kPa; (RYGB > SG) 0.8 ± 0.3 vs. 1.1 ± 0.4 −1.0 ± 1.8 vs. −1.7 ± 1.3; 0.3 ± 0.2 vs. 0.3 ± 0.1; 2.3 ± 1.2 vs. 2.8 ± 1.1. |
| Garg, 2018 [118] | Prospective analysis Bariatric surgery population n = 76 FU 1 year (32 biopsies) | Controlled attenuation parameter (CAP) Transient elastography: liver stiffness Liver biopsy | Pre-operation: 326.5 (301–360.5) dB/m; Success rate pre-operation 87.9% Pre-operation 7.0 (5.0–9.5) kPa Improvement in hepatic steatosis, lobular inflammation, ballooning and fibrosis; NASH resolved in 3 out of 4 patients. |
| Cherla, 2020 [116] | Prospective analysis of biopsy-proven NAFLD SG, n = 62 RYGB, n = 248 FU 4 years (median) | Liver function tests AST, SG AST, RYGB ALT, SG ALT, RYGB | 84% normalized after bariatric surgery 49.1 ± 21.5 to 28.0 ± 16.5 49.3 ± 22.0 to 26.5 ± 15.5 61.7 ± 30.0 to 27.2 ± 21.5 59.4 ± 24.9 to 26.1 ± 19.2 |
| Wirth, 2020 [119] | Retrospective analysis; Bariatric surgery, n = 2942 Control, n = 5884 FU 31 months | Risk of NAFLD progression to cirrhosis after bariatric surgery | Reduced risk: HR 0.31 (95% CI 0.19–0.52) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Hoozemans, J.; de Brauw, M.; Nieuwdorp, M.; Gerdes, V. Gut Microbiome and Metabolites in Patients with NAFLD and after Bariatric Surgery: A Comprehensive Review. Metabolites 2021, 11, 353. https://doi.org/10.3390/metabo11060353
AMA Style
Hoozemans J, de Brauw M, Nieuwdorp M, Gerdes V. Gut Microbiome and Metabolites in Patients with NAFLD and after Bariatric Surgery: A Comprehensive Review. Metabolites. 2021; 11(6):353. https://doi.org/10.3390/metabo11060353
Chicago/Turabian StyleHoozemans, Jacqueline, Maurits de Brauw, Max Nieuwdorp, and Victor Gerdes. 2021. "Gut Microbiome and Metabolites in Patients with NAFLD and after Bariatric Surgery: A Comprehensive Review" Metabolites 11, no. 6: 353. https://doi.org/10.3390/metabo11060353
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.