Molecular Typing of Mycoplasma genitalium-Positive Specimens Discriminates between Persistent and Recurrent Infections in Cases of Treatment Failure and Supports Contact Tracing


Abstract
:1. Introduction
2. Materials and Methods
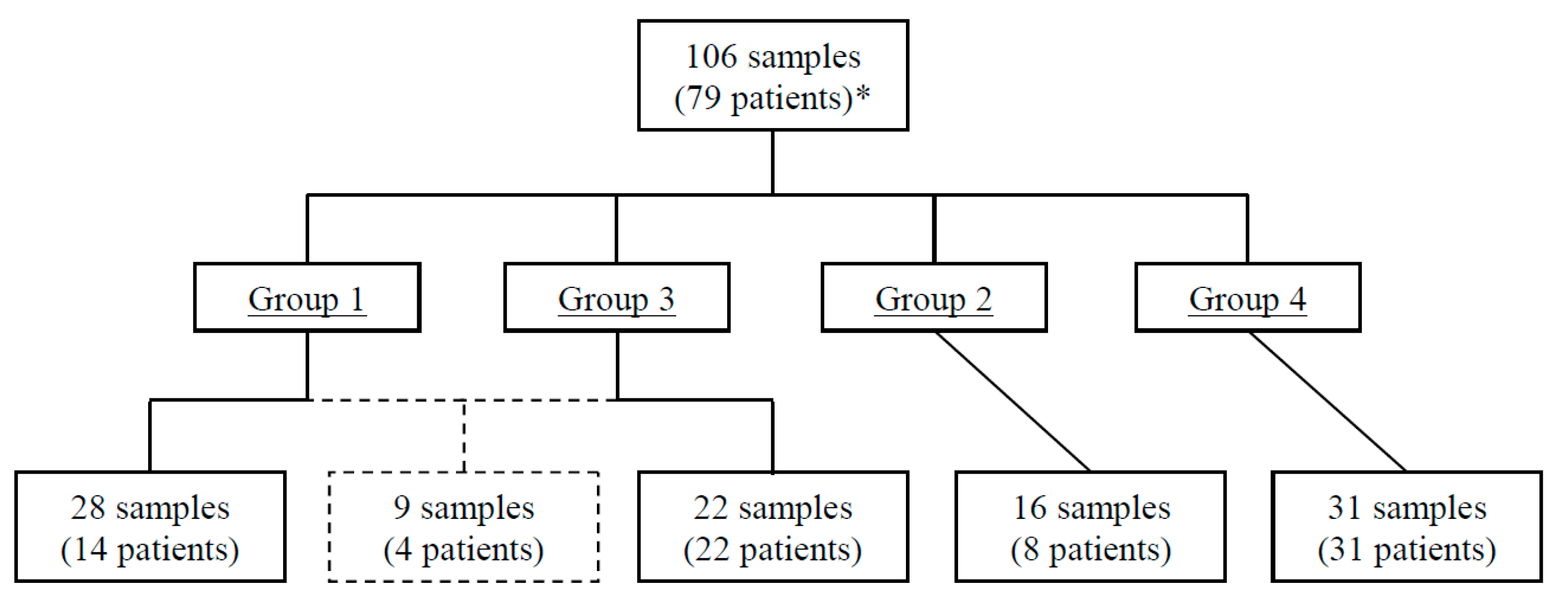
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Jensen, J.S.; Cusini, M.; Gomberg, M.; Moi, H. 2016 European guideline on Mycoplasma genitalium infections. J. Eur. Acad. Dermatol. Venereol. 2016, 30, 1650–1656. [Google Scholar] [CrossRef] [PubMed]
- Workowski, K.A.; Bolan, G.A. Centers for Disease Control and Prevention. Sexually transmitted diseases treatment guidelines, 2015. MMWR Recomm. Rep. 2015, 64, 1–137. [Google Scholar] [PubMed]
- Piñeiro, L.; Idigoras, P.; de la Caba, I.; López-Olaizola, M.; Cilla, G. Guided antibiotic therapy for Mycoplasma genitalium infections: Analysis of mutations associated with resistance to macrolides and fluoroquinolones. Enferm. Infecc. Microbiol. Clin. 2019, 37, 394–397. [Google Scholar] [CrossRef] [PubMed]
- Hjorth, S.V.; Björnelius, E.; Lidbrink, P.; Falk, L.; Dohn, B.; Berthelsen, L.; Ma, L.; Martin, D.H.; Jensen, J.S. Sequence-based typing of Mycoplasma genitalium reveals sexual transmission. J. Clin. Microbiol. 2006, 44, 2078–2083. [Google Scholar] [CrossRef]
- Sweeney, E.L.; Trembizki, E.; Bletchly, C.; Bradshaw, C.S.; Menon, A.; Francis, F.; Langton-Lockton, J.; Nimmo, G.R.; Whiley, D.M. Levels of Mycoplasma genitalium Antimicrobial Resistance Differ by Both Region and Gender in the State of Queensland, Australia: Implications for Treatment Guidelines. J. Clin. Microbiol. 2019, 57, e01555-18. [Google Scholar] [CrossRef]
- Touati, A.; Peuchant, O.; Jensen, J.S.; Bébéar, C.; Pereyre, S. Direct detection of macrolide resistance in Mycoplasma genitalium isolates from clinical specimens from France by use of real-time PCR and melting curve analysis. J. Clin. Microbiol. 2014, 52, 1549–1555. [Google Scholar] [CrossRef]
- Jensen, J.S.; Uldum, S.A.; Søndergård-Andersen, J.; Vuust, J.; Lind, K. Polymerase chain reaction for detection of Mycoplasma genitalium in clinical samples. J. Clin. Microbiol. 1991, 29, 46–50. [Google Scholar]
- Ma, L.; Martin, D.H. Single-nucleotide polymorphisms in the rRNA operon and variable numbers of tandem repeats in the lipoprotein gene among Mycoplasma genitalium strains from clinical specimens. J. Clin. Microbiol. 2004, 42, 4876–4878. [Google Scholar] [CrossRef]
- Ma, L.; Taylor, S.; Jensen, J.S.; Myers, L.; Lillis, R.; Martin, D.H. Short tandem repeat sequences in the Mycoplasma genitalium genome and their use in a multilocus genotyping system. BMC Microbiol. 2008, 8, 130. [Google Scholar] [CrossRef]
- Musatovova, O.; Baseman, J.B. Analysis identifying common and distinct sequences among Texas clinical strains of Mycoplasma genitalium. J. Clin. Microbiol. 2009, 47, 1469–1475. [Google Scholar] [CrossRef]
- Iverson-Cabral, S.L.; Astete, S.G.; Cohen, C.R.; Rocha, E.P.; Totten, P.A. Intrastrain heterogeneity of the mgpB gene in Mycoplasma genitalium is extensive in vitro and in vivo and suggests that variation is generated via recombination with repetitive chromosomal sequences. Infect. Immun. 2006, 74, 3715–3726. [Google Scholar] [CrossRef] [PubMed]
- Cazanave, C.; Charron, A.; Renaudin, H.; Bébéar, C. Method comparison for molecular typing of French and Tunisian Mycoplasma genitalium-positive specimens. J. Med. Microbiol. 2012, 61 Pt 4, 500–506. [Google Scholar] [CrossRef] [PubMed]
- Chrisment, D.; Charron, A.; Cazanave, C.; Pereyre, S.; Bébéar, C. Detection of macrolide resistance in Mycoplasma genitalium in France. J. Antimicrob. Chemother. 2012, 67, 2598–2601. [Google Scholar] [CrossRef] [PubMed]
- Pond, M.J.; Nori, A.V.; Witney, A.A.; Lopeman, R.C.; Butcher, P.D.; Sadiq, S.T. High prevalence of antibiotic-resistant Mycoplasma genitalium in nongonococcal urethritis: The need for routine testing and the inadequacy of current treatment options. Clin. Infect. Dis. 2014, 58, 631–637. [Google Scholar] [CrossRef] [PubMed]
- Hunter, P.R.; Gaston, M.A. Numerical index of the discriminatory ability of typing systems: An application of Simpson’s index of diversity. J. Clin. Microbiol. 1988, 26, 2465–2466. [Google Scholar]
- Jensen, J.S.; Bradshaw, C.S.; Tabrizi, S.N.; Fairley, C.K.; Hamasuna, R. Azithromycin treatment failure in Mycoplasma genitalium-positive patients with nongonococcal urethritis is associated with induced macrolide resistance. Clin. Infect. Dis. 2008, 47, 1546–1553. [Google Scholar] [CrossRef]
- Falk, L.; Enger, M.; Jensen, J.S. Time to eradication of Mycoplasma genitalium after antibiotic treatment in men and women. J. Antimicrob. Chemother. 2015, 70, 3134–3140. [Google Scholar] [CrossRef]
- Vester, B.; Douthwaite, S. Macrolide resistance conferred by base substitutions in 23S rRNA. Antimicrob. Agents Chemother. 2001, 45, 1–12. [Google Scholar] [CrossRef]
- Bradshaw, C.S.; Jensen, J.S.; Waites, K.B. New Horizons in Mycoplasma genitalium Treatment. J. Infect. Dis. 2017, 216 (Suppl. 2), S412–S419. [Google Scholar] [CrossRef]
- Mena, L.A.; Mroczkowski, T.F.; Nsuami, M.; Martin, D.H. A randomized comparison of azithromycin and doxycycline for the treatment of Mycoplasma genitalium-positive urethritis in men. Clin. Infect. Dis. 2009, 48, 1649–1654. [Google Scholar] [CrossRef]
- Schwebke, J.R.; Rompalo, A.; Taylor, S.; Seña, A.C.; Martin, D.H.; Lopez, L.M.; Lensing, S.; Lee, J.Y. Re-evaluating the treatment of nongonococcal urethritis: Emphasizing emerging pathogens—A randomized clinical trial. Clin. Infect. Dis. 2011, 52, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Manhart, L.E.; Gillespie, C.W.; Lowens, M.S.; Khosropour, C.M.; Colombara, D.V.; Golden, M.R.; Hakhu, N.R.; Thomas, K.K.; Hughes, J.P.; Jensen, N.L.; et al. Standard treatment regimens for nongonococcal urethritis have similar but declining cure rates: A randomized controlled trial. Clin. Infect. Dis. 2013, 56, 934–942. [Google Scholar] [CrossRef] [PubMed]
- Horner, P.J.; Blee, K.; Falk, L.; van der Meijden, W.; Moi, H. 2016 European guideline on the management of non-gonococcal urethritis. Int. J. STD AIDS 2016, 27, 928–937. [Google Scholar] [CrossRef] [PubMed]
- Read, T.R.H.; Fairley, C.K.; Murray, G.L.; Jensen, J.S.; Danielewski, J.; Worthington, K.; Doyle, M.; Mokany, E.; Tan, L.; Chow, E.P.F.; et al. Outcomes of Resistance-guided Sequential Treatment of Mycoplasma genitalium Infections: A Prospective Evaluation. Clin. Infect. Dis. 2019, 68, 554–560. [Google Scholar] [CrossRef] [PubMed]
- Quinn, T.C.; Gaydos, C.A. Treatment for Chlamydia Infection—Doxycycline versus Azithromycin. N. Engl. J. Med. 2015, 373, 2573–2575. [Google Scholar] [CrossRef] [PubMed]
- Khosropour, C.M.; Manhart, L.E.; Colombara, D.V.; Gillespie, C.W.; Lowens, M.S.; Totten, P.A.; Golden, M.R.; Simoni, J. Suboptimal adherence to doxycycline and treatment outcomes among men with non-gonococcal urethritis: A prospective cohort study. Sex Transm. Infect. 2014, 90, 3–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levinson, G.; Gutman, G.A. Slipped-strand mispairing: A major mechanism for DNA sequence evolution. Mol. Biol. Evol. 1987, 4, 203–221. [Google Scholar]


{kind=link}
| Study Group(s) (Method) | No. of Types | Size (%) of Largest Type | Discriminatory Index |
|---|---|---|---|
| G1–3 (Adh+Lip+ITS1+ITS2) | 44 | 4 | 0.9871 |
| G1–3 (Adh+Lip) | 43 | 4 | 0.9863 |
| G1–3 (ITS1+ITS2) | 4 | 64 | 0.5056 |
| G1–3 (Adh) | 21 | 14 | 0.9334 |
| G1–3 (Lip) | 21 | 9 | 0.9496 |
| G4 (Adh) | 23 | 16 | 0.9634 |
| G1–4 (Adh) * | 33 | 14 | 0.9524 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piñeiro, L.; Idigoras, P.; Cilla, G. Molecular Typing of Mycoplasma genitalium-Positive Specimens Discriminates between Persistent and Recurrent Infections in Cases of Treatment Failure and Supports Contact Tracing. Microorganisms 2019, 7, 609. https://doi.org/10.3390/microorganisms7120609
Piñeiro L, Idigoras P, Cilla G. Molecular Typing of Mycoplasma genitalium-Positive Specimens Discriminates between Persistent and Recurrent Infections in Cases of Treatment Failure and Supports Contact Tracing. Microorganisms. 2019; 7(12):609. https://doi.org/10.3390/microorganisms7120609
Chicago/Turabian StylePiñeiro, Luis, Pedro Idigoras, and Gustavo Cilla. 2019. "Molecular Typing of Mycoplasma genitalium-Positive Specimens Discriminates between Persistent and Recurrent Infections in Cases of Treatment Failure and Supports Contact Tracing" Microorganisms 7, no. 12: 609. https://doi.org/10.3390/microorganisms7120609