Transferable Extended-Spectrum β-Lactamase (ESBL) Plasmids in Enterobacteriaceae from Irrigation Water

1
Microbiological Food Safety, Agroscope, 8820 Wädenswil, Switzerland
2
Microbiological Food Safety, Agroscope, 3003 Liebefeld, Switzerland
3
Institute for Epidemiology and Pathogen Diagnostics, Julius Kühn-Institut—Federal Research Centre for Cultivated Plants (JKI), 38104 Braunschweig, Germany
4
Department of Life Sciences, Albstadt-Sigmaringen University, 72488 Sigmaringen, Germany
*
Author to whom correspondence should be addressed.
Microorganisms 2020, 8(7), 978; https://doi.org/10.3390/microorganisms8070978
Submission received: 13 June 2020
/
Revised: 26 June 2020
/
Accepted: 28 June 2020
/
Published: 30 June 2020
(This article belongs to the Special Issue Antimicrobial Resistance: From the Environment to Human Health)
Abstract
:Extended-spectrum β-lactamase (ESBL)-producing Enterobacteriaceae are classified as serious threats to human health by the U.S. Centers for Disease Control and Prevention. Water used for irrigation of fresh produce can transmit such resistant bacteria directly to edible plant parts. We screened ESBL-producing Escherichia coli, Enterobacter cloacae, and Citrobacter freundii isolated from irrigation water for their potential to transmit resistance to antibiotic-susceptible E. coli. All strains were genome-sequenced and tested in vitro for transmission of resistance to third-generation cephalosporins on solid agar as well as in liquid culture. Of the 19 screened isolates, five ESBL-producing E. coli were able to transfer resistance with different efficiency to susceptible recipient E. coli. Transconjugant strains were sequenced for detection of transferred antibiotic resistance genes (ARGs) and compared to the known ARG pattern of their respective donors. Additionally, phenotypic resistance patterns were obtained for both transconjugant and corresponding donor strains, confirming ESBL-producing phenotypes of all obtained transconjugants.
1. Introduction
The discovery and subsequent broad use of antibiotics in the first half of the last century has revolutionized human and veterinarian medicine. However, a setback promptly followed, as upon discovery of each novel antibiotic, emergence of resistant bacteria soon followed [1]. Antibiotic resistance is conferred by three mechanisms: (1) bacterial target alteration, e.g., by mutation, (2) drug inactivation, e.g., by hydrolyzing enzymes, and (3) decreased intracellular drug concentration through low cell permeability or drug efflux pumps [2]. The genes conferring such antibiotic resistance (ARGs) are of special concern to human health when carried by mobile elements such as phages or plasmids, as these can spread rapidly within bacterial communities and, thus, from commensal or environmental bacteria to human pathogens [3]. The extent and speed of this so-called horizontal gene transfer (HGT) depends on various factors such as bacterial density (biofilms vs. planktonic cells), conditions supporting bacterial growth, or plasmid type (broad- or narrow-host-range plasmids). Barriers to HGT exist, such as recipient helicase compatibility or host restriction system susceptibility, which can reduce but not completely prevent the transfer and establishment of a transferable plasmid in the bacterial host cell [4]. Of note, antibiotic resistance (AR) plasmids usually carry several ARGs, often conferring resistance to multiple antibiotic classes. Antibiotic-resistant bacteria (ARB) with acquired resistance to at least one agent in three or more antibiotic classes are referred to as multidrug-resistant (MDR) [5].
ARB can be classified into different threat groups—concerning, serious, and urgent threats—based on the burden they pose for human health. Treatment options for these ARB become more and more limited, leaving only a few last-resort antibiotics at choice for serious and urgent threat ARB. Extended-spectrum β-lactamase (ESBL)-producing Enterobacteriaceae are listed by the U.S. Centers for Disease Control and Prevention (CDC) among the serious threats [6]. In its 2019 antibiotic resistance threat report, the CDC counted more than 2.8 million antibiotic-resistant infections causing 35,000 deaths in the United States, and they observed an increase by 50% in infections caused by ESBL-producing Enterobacteriaceae. These encompassed ESBL-producing Escherichia coli (E. coli), Enterobacter spp., Citrobacter spp., and Klebsiella pneumoniae.
The β-lactamase enzyme family can be subdivided into four groups based on protein homology, known as Ambler scheme classes A to D [7]. Extended-spectrum β-lactamases almost exclusively belong to Ambler class A and can be defined as enzymes conferring resistance to penicillins, first- to third-generation cephalosporins, and aztreonam [8]. ESBL-encoding genes are often located on transferable plasmids and can thus spread among bacteria. Such genes are far from confined to clinical settings, and they have also been detected in bacteria from companion and food-producing animals [9,10], in various foods including raw fresh produce [11,12,13,14], as well as different environmental sources such as surface waters [15,16] or agricultural as well as undisturbed soils [17,18]. One of the most important ESBL enzyme families is the CTX-M family [19]. They are assumed to originate from chromosomal blaCTX-M-related genes from different environmental Kluyvera spp. In this process, the involvement of mobile genetic elements (MGEs) such as insertion sequences, class 1 integrons, and transposons has been crucial [20].
Foods such as fresh produce that are usually consumed raw can easily transfer their bacterial communities to the consumer, and many studies have documented the presence of ARB in such products [21,22,23,24]. Fresh produce is often exposed to a broad variety of potential contamination sources, especially when cultured in open greenhouses or fields [25]. One contamination source is surface water such as rivers, channels, or lakes that are often used for irrigation, especially in areas with limited water resources [26]. Additionally, wastewater treatment plants often release treated wastewater into the nearby streams and have been identified as point sources of micropollutants such as antibiotics, ARB, and ARGs into the environment [27]. By irrigating fresh produce, especially overhead, edible plant parts can get contaminated with these micropollutants. Moreover, uptake of pharmaceutical pollutants such as antibiotics by different crops has been shown in real agricultural systems [28], and the introduction of ARB can affect the plant microbiome, which represents a major pathway of human exposure to microbes [29]. Nevertheless, irrigation water is not monitored routinely for pathogenic bacteria, let alone for ARB or ARG content. In Switzerland, a risk analysis for irrigation water has been proposed by SwissGAP since 2017, including reference values for maximal loads of the fecal indicators E. coli and Enterococcus spp. [30]. Only in case of operational changes on the farm, the risk analysis must be updated. This, however, is not comparable to systematic monitoring. In a previous study, we found that a large proportion (22%) of irrigation water from major Swiss vegetable growing areas contained ESBL-producing E. coli [15]. In the present study, we investigated the ability of these ESBL-producing E. coli as well as isolates belonging to ESBL-producing Enterobacter cloacae (E. cloacae) and Citrobacter freundii (C. freundii) complexes from the same irrigation water sources for their ability to transfer their ESBL-resistance phenotype to a β-lactam susceptible E. coli. We conclude that 5 of the investigated 11 ESBL-producing E. coli were able to transfer ESBL-encoding genes and ESBL-phenotype by conjugation in both solid and liquid medium experiments, while no conjugation was observed for the ESBL-producing E. cloacae or C. freundii complex isolates.
2. Materials and Methods
2.1. Bacterial Strains and Culture Conditions
ESBL-producing Enterobacteriaceae were isolated from irrigation water originating from important Swiss vegetable growing areas [15]. Irrigation water sources included groundwater as well as various surface waters such as rivers, creeks, or ponds. Briefly, ESBL-producing Enterobacteriaceae were isolated by bacterial enrichment in EE broth Mossel (Beckton Dickinson, Franklin Lakes, NJ, USA) and subsequent streaking onto commercial, ready-to-use Brilliance ESBL plates (Oxoid Ltd., Hampshire, UK). Single colonies with morphologies corresponding to the target bacteria ESBL-producing E. coli, Enterobacter spp., or Citrobacter spp. according to the manufacturer were identified at species level by MALDI biotyping (see Section 2.2), re-grown in lysogeny broth (LB; Sigma-Aldrich, St. Louis, MO, USA) containing ceftazidime (CAZ, 8 or 0.8 mg/L; Sigma-Aldrich), and stored with glycerol at −80 °C. Strains for conjugation experiments and antibiotic susceptibility testing were re-grown in LB with the appropriate CAZ concentration. Representative strains from each bacterial genus were selected for conjugation experiments: 11 ESBL-producing E. coli strains (H2, H6, H10, H17, H22, H25, H30, H38, H40, H44, and H45), 5 E. cloacae complex strains (H9, H16, H24, H34, and H36), and 3 C. freundii complex strains (H12, H41, and H42).
Kanamycin (KAN)- and rifampin (RIF)-resistant, green fluorescent protein (gfp)-tagged E. coli CV601 [31], serving as a recipient strain in conjugation experiments, was grown in LB containing 50 mg/L of each KAN and RIF.
2.2. MALDI Biotyping
MALDI biotyping was performed by direct smearing as described previously [32] using a microflex LT MALDI-TOF mass spectrometer (Bruker Daltonics, Bremen, Germany) and the associated MALDI biotyper RTC Software (Version 3.1; Bruker Daltonics). Notably, species belonging to either E. cloacae complex (such as E. ludwigii or E. asburiae) or C. freundii complex (such as C. braakii or C. gillenii) could not be distinguished according to the manufacturer and were grouped therefore into the corresponding complex.
2.3. Antibiotic Susceptibility Testing
For all 19 ESBL-producing Enterobacteriaceae from irrigation water, resistance to 32 clinically relevant antibiotics was determined in disk diffusion assays as described previously [15]. Additionally, all ESBL-producing donor and transconjugant strains obtained from filter conjugation experiments were screened for resistance against 31 microbial agents by broth dilution using the MicroScan autoSCAN-4 System (Beckman Coulter Life Sciences, Indianapolis, IN, USA) with the Neg MIC 44 panel. Susceptibility testing was performed according to the manufacturer’s guidelines.
2.4. DNA Extractions, Sequencing, and Bioinformatics
Genomic DNA was extracted from all selected ESBL-producing Enterobacteriaceae and transconjugants using the commercial GenEluteTM Bacterial Genomic DNA Kit (Sigma-Aldrich) according to the manufacturer’s instructions. Genomic DNA was sequenced at Eurofins Genomics (Konstanz, Germany) on an Illumina HiSeq4000 instrument (paired-end, 2 × 150 bp). Raw data were assembled to contigs using CLC Genomics Workbench 10.0 (Qiagen, Venlo, Netherlands) for further analysis. The Illumina raw reads (ERR4065331–ERR4065341; ERR4065362–ERR4065369; and ERR4065372–ERR4065376) were submitted to the European Nucleotide Archive (ENA) under BioProject PRJEB37967. Additionally, plasmid DNA from ESBL-producing E. coli and their transconjugants was enriched using a commercial PureYield Plasmid Maxiprep System (Promega, Fitchburg, WI, USA). Enriched plasmid DNA was sequenced on a Pacific Biosciences RSII instrument and used for hybrid assemblies as described previously [15]. The PacBio raw reads (ERR4235917–ERR4235921; ERR4234695; ERR4235916) were also submitted to ENA under PRJEB37967.
In order to estimate the diversity of the tested strains, multi-locus sequence typing (MLST) was performed and a phylogenetic tree constructed from the assembled contigs, using the Center for Genomic Epidemiology (CGE) online tools MLST 2.0 and CSI Phylogeny 1.4 [33,34]. Phylogenetic trees were illustrated using FigTree v1.4.4 [35]. Antibiotic resistance genes and plasmid replicons were identified using ResFinder 3.2 and PlasmidFinder 2.1 [36,37]. All CGE tools were run with default settings for the respective tool version. One type strain per ESBL-producing species was included in the analysis (C. freundii ATCC8090, E. cloacae ATCC13047, and E. coli ATCC11775).
2.5. Plasmid Transfer Assays
To examine the frequency of ESBL resistance genes and phenotype transfer to non-ESBL-producers, ESBL-producing irrigation water isolates were used as donors in filter and broth conjugation assays. The donor strain collection comprised 11 ESBL E. coli, 5 ESBL E. cloacae complex, and 3 ESBL C. freundii complex strains. Escherichia coli CV601 served as a recipient strain in all experiments.
The filter conjugation assay was adapted from a previously described method [38]. Donor and recipient strains were grown overnight in Tryptic Soy Broth (TSB, Merck, Darmstadt, Germany) supplemented with CAZ (8 or 0.8 mg/L), RIF (50 mg/L), and KAN (50 mg/L), respectively. Absorbance was measured and adjusted to OD600 = 4.0 ± 0.5 for all cultures. Recipient and donor cultures were centrifuged (3100 × g, 5 min), the supernatants discarded, the pellets washed twice with 1 mL of 0.1 × TSB, and finally re-suspended in 1 mL of 0.1 × TSB. Then, 500 µL of each donor culture was added to 500 µL of recipient culture and mixed by pipetting. The resulting conjugation mixture was centrifuged (3100 × g, 5 min), the supernatant discarded, and the pellet re-suspended in 200 µL of 0.1 × TSB. For conjugation, Durapore® membrane filters (Merck Millipore, Burlington, MA, USA) were placed on Plate Count Agar (PCA; Thermo Fisher Scientific, Waltham, MA, USA), and the entire conjugation mixture was transferred to the center of the filter. As controls, after washing, 1 mL of each donor culture only or recipient culture only was centrifuged, re-suspended in 200 µL of 0.1 × TSB, and transferred to a filter. After overnight incubation at 28 °C, filters were placed in 2 mL of sterile 0.9% NaCl solution and vortexed for 1 min. Conjugation suspensions and controls were serially diluted and appropriate dilutions plated in duplicates on PCA plates containing RIF (50 mg/L), KAN (50 mg/L), and CAZ (8 or 0.8 mg/L). To determine the number of recipient cells, spot plating was performed for each conjugation mixture by spotting three 20 µL drops per dilution (10−5 to 10−8) on PCA supplemented with RIF (50 mg/L) and KAN (50 mg/L). To determine transfer frequencies, the total number of transconjugants was divided by the number of recipients as determined by spot plating. All plates were incubated at 28 °C for up to 72 h.
The broth conjugation assay was performed as described previously [38]. In short, donor and recipient strains were grown in LB overnight (37 °C, 180 rpm). In 2 mL Eppendorf tubes containing 1 mL of LB, 500 µL of each donor strain were mixed with 500 µL of recipient strain and incubated for 24 h at 37 °C. Conjugation mixtures were serially diluted, and 100 µL of appropriate dilutions were plated in duplicate onto LB agar plates containing KAN (50 mg/L), RIF (50 mg/L), and CAZ (8 or 0.8 mg/L) followed by incubation at 37 °C for up to 48 h.
For both, filter and broth conjugation assays, green fluorescence was confirmed under UV light for putative transconjugants. Per replicate, three transconjugants were picked and re-streaked. For confirmation of E. coli CV601 background in transconjugants, repetitive sequence-based PCR fingerprinting (REP-PCR) was performed using the (GTG)5 primer (5′-GTGGTGGTGGTGGTG-3′) [39]. Template DNA of putative transconjugants and recipient was obtained by suspending a single colony in 50 µL of sterile water followed by boiling for 10 min at 99 °C. For PCR conditions see Table A1.
3. Results
3.1. Phylogenetic Diversity of Donor Strains
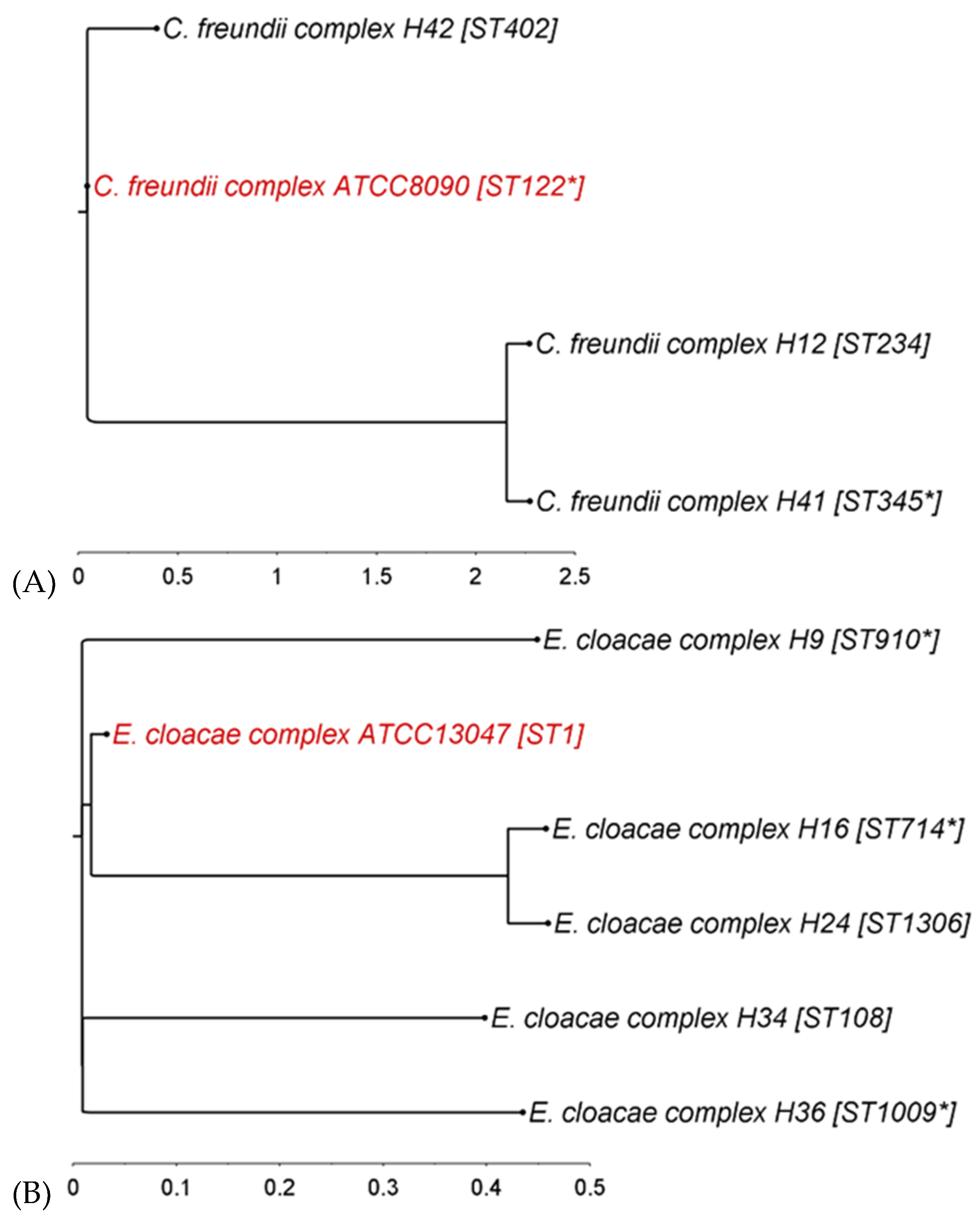
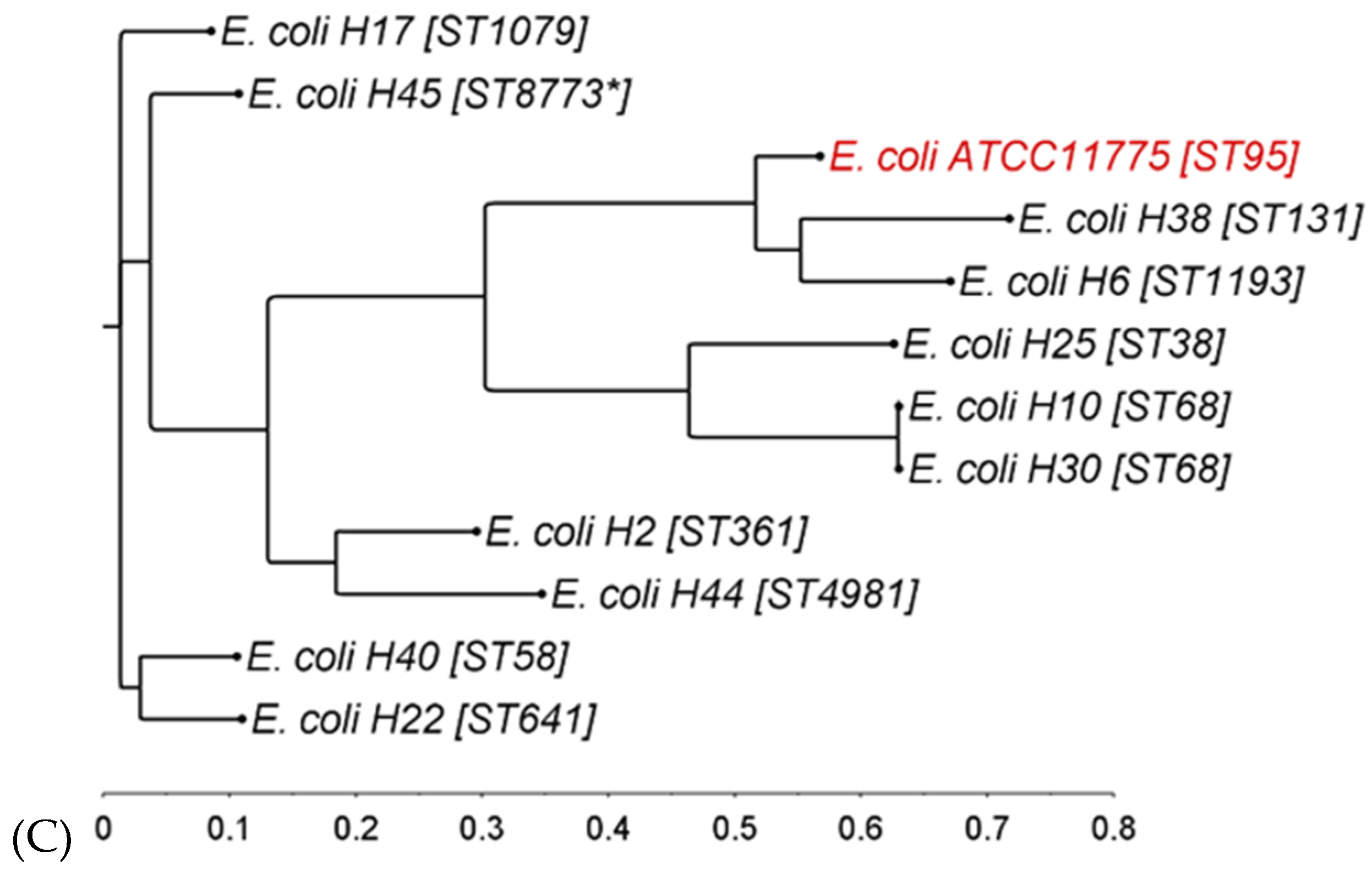
The phylogenetic diversity of the ESBL-producing strains was analyzed by MLST typing and by calculating single-nucleotide polymorphism (SNP)-based phylogenetic trees (Figure 1).
For C. freundii complex strains, 78.5% of the reference type strain genome was covered by all three isolates, with the individual strains sharing at least 80.6% with the reference. Two strains (H12 and H41) clustered together in one clade, while the third strain (H42) was separated from them by the reference strain (Figure 1A). All four strains belonged to different sequence types (STs).
In the E. cloacae complex, 64.7% of the reference strain genome was covered by all five irrigation water E. cloacae strains and at least 71.1% by the single strains. Strains H16 and H24 clustered closely in one clade, while the others were separated clearly, with the reference strain placed in between them. They all belonged to different STs, while three belonged to unknown sequence types for which the nearest known ST is indicated (Figure 1B).
All together, the analyzed ESBL-producing E. coli covered 72.6% of the reference strain genome with a minimum of 80.2% shared genome when comparing individual strains to the reference. Two strains (H10 and H30) belonged to ST68 and consequently clustered together in the tree (Figure 1C). Strain H38, belonging to the global pandemic clone ST131, closely clustered with strain H6 (ST1193) and the reference strain. Taken together, the ESBL-producing E. coli used in this study represented a broad diversity of STs, with the reference type strain lying in between them (Figure 1C). Interestingly, three strains originating from the same water source (H2, H6, and H17) did not cluster together in the tree, demonstrating that irrigation water can contain multiple and distantly related ESBL-producing E. coli. On the other hand, H10 and H25, also originating from one irrigation water sample, clustered closely together, although they belonged to different STs.
Finally, SNP-distance matrices were calculated for each species group, featuring the number of SNPs detected by pairwise comparison. Based on the minimum and maximum SNP differences detected, E. coli showed the largest range from nearest to farthest strain (min. 25; max. 46,239), followed by E. cloacae (min. 6913; max. 52,453) and C. freundii (min. 13,962; max. 66,505). In terms of maximum SNP difference, the C. freundii strains were the most dispersed.
3.2. Antibiotic Resistance and Plasmid Profiles of Donor Strains
For the ESBL-producing E. coli from different irrigation waters, phenotypic resistance determined by disk diffusion as well as ARGs and plasmid replicons detected by sequencing have been published previously [15]. Briefly, the ARGs responsible for the observed ESBL phenotype all belonged to the blaCTX-M gene family. The most frequently detected were blaCTX-M-1 (4 of 11) and blaCTX-M-15 (3 of 11). Five of the 11 detected blaCTX-M genes were linked to a known plasmid replicon, 3 of which were identified as IncI1.
Disk diffusion profiles as well as ARGs and plasmid replicons detected in ESBL-producing E. cloacae and C. freundii are shown in Table 1. In four of five E. cloacae strains, β-lactam resistance genes of the AmpC-type were detected (blaACT), while no β-lactamase gene was detected in one E. cloacae strain. In C. freundii strains as well, β-lactamases of Ambler class C were detected (blaCMY genes). Many of these blaCMY-2-like genes occur naturally in C. freundii, among others the detected blaCMY-48 and blaCMY-83 [40]. Additionally, in C. freundii H42 a blaTEM-1B gene was detected. Apart from ARGs encoding β-lactamases, ARGs associated with resistance to fosfomycin (fosA genes), colistin (mcr-9), and the quinolone ciprofloxacin (qnrE1 and qnrB) were detected. Again, C. freundii H42 harbored additional ARGs in good correspondence to its observed richer phenotypic resistance profile (Table 1).
3.3. Filter and Broth Conjugation
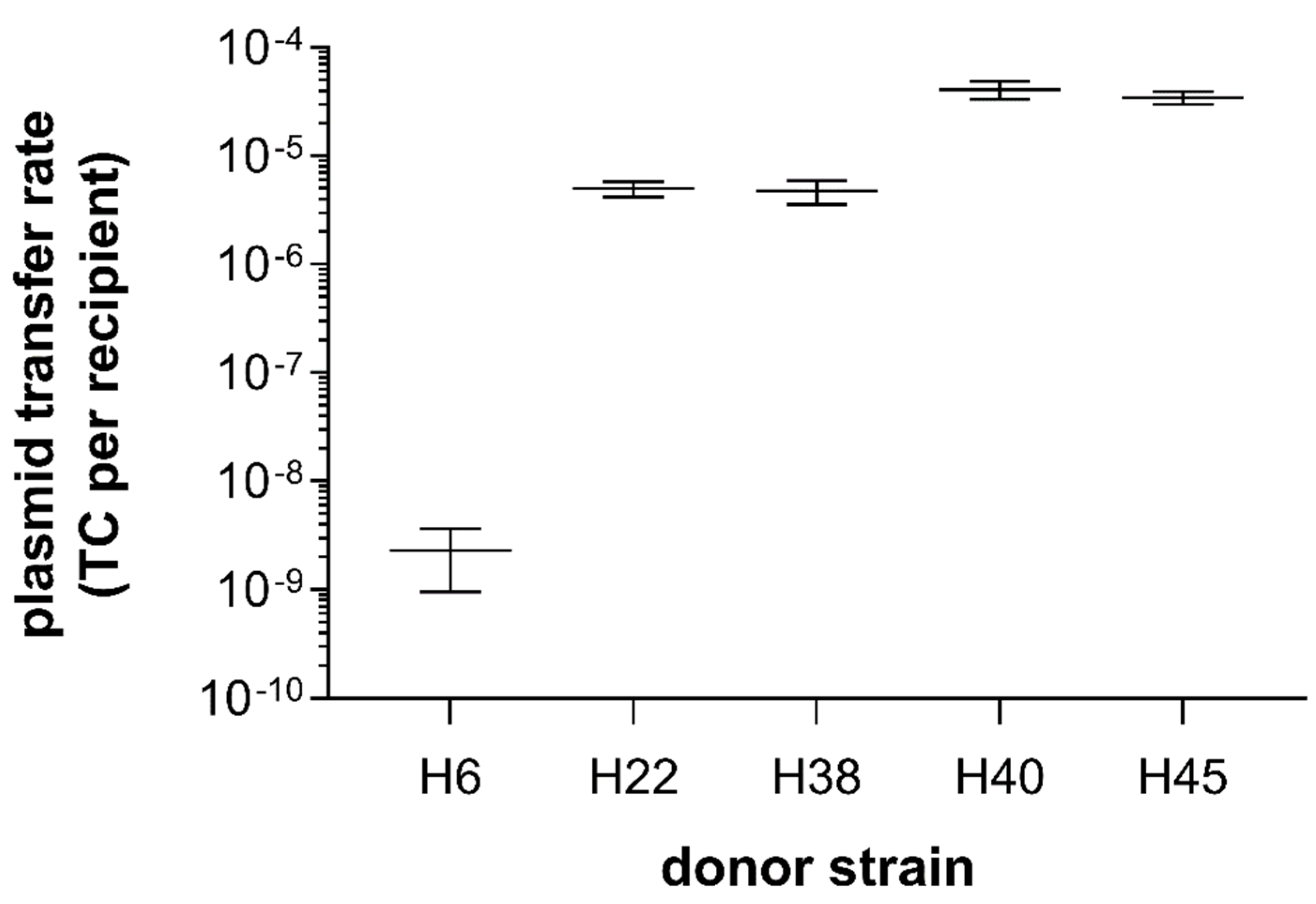
From the 19 ESBL-producing Enterobacteriaceae strains used in the filter conjugation assay, only 5 E. coli strains (for H6, H22, H38, H40, and H45) could transfer their resistance to a non-ESBL-producing recipient strain, resulting in ESBL-producing transconjugants (Figure 2). In the broth conjugation assay, these same five strains were the only ones to transfer successfully the ESBL-phenotype to the recipient strain. The successful transfer was confirmed by the matching REP-PCR fingerprint profiles of the transconjugants and the recipient. Plasmid transfer frequencies in filter conjugation assays were 2.3 × 10−9 (± 1.9 × 10−9) for H6, 5 × 10−6 (± 1.1 × 10−6) for H22, 4.7 × 10−6 (± 1.7 × 10−6) for H38, 4.1 × 10−5 (± 1.1 × 10−5) for H40, and 3.5 × 10−5 (± 6.8 × 10−6) for H45, respectively (Figure 2). Overall, conjugation rates were very similar between biological replications, with the exception of H6, whose conjugation rates were close to the detection limit. Of note, strain H6 harbors the blaCTX-M gene with the most associated ARGs, summing up to a total of nine ARGs on one contig (see below).
3.4. Donor and Transconjugant Strain Comparison
3.4.1. Phenotypic Antibiotic Resistance
Broth microdilution results from ESBL-producing E. coli and their respective transconjugants are shown in Table 2. No resistance was observed for any strain towards cefoxitin, chloramphenicol, colistin, doripenem, ertapenem, fosfomycin, gentamycin, imipenem, meropenem, piperacillin–tazobactam, and tigecycline.
The recipient strain E. coli CV601 showed intermediate resistance towards amikacin (AMK) and tobramycin (TOB), as expected due to presence of a gfp-aph(3′)-III cassette. These two resistances were therefore detected in most transconjugants as well (Table 2). Most importantly, the ESBL phenotype was detected phenotypically in all transconjugants. Resistance phenotypes of H6 and H6-TC were identical with the exception of fluoroquinolone resistance (ciprofloxacin (CIP) and levofloxacin (LVX)) observed in the donor strain only. Strains H22 and H22-TC shared all phenotypic resistances, with the exception of recipient-associated aminoglycoside resistance (Table 2). Strain H38 and the respective transconjugant strain were identical in their resistances towards β-lactams. They differed in H38 having fluoroquinolone resistances (CIP and LVX; Table 2). Comparing H40 to its transconjugant, we again observed a common β-lactam resistance profile. In terms of differences, H40 showed resistance to trimethoprim–sulfamethoxazole (SXT), minocycline (MI), and tetracycline (TE), while H40-TC to TE only (and one in three replicates resistant towards MI). Finally, H45 and H45-TC shared all β-lactam resistances.
3.4.2. Molecular Detection of ARGs and Resistance Point Mutations
On the molecular side, we identified ARGs, plasmid replicons, and point mutations known to confer antibiotic resistance in transconjugant strains and their respective ESBL-producing donors. The detected ARGs grouped by contig are in complete accordance between donor and respective transconjugant strain for all five donor-transconjugant pairs (Table 3). Each of the contigs carrying the blaCTX-M gene was transferred as an entity to the recipient strain, including the long ARG contig of strain H6 encompassing nine ARGs in total. Moreover, the plasmid replicons associated with the blaCTX-M contigs in donor strains were detected in the transconjugant strains as well, with H6-TC forming the only exception. Finally, all transconjugants contained aph(3′)-III originating from the recipient strain.
Strain H40 was the only donor to harbor a second, non-ESBL plasmid of the IncFIB/IncFIC replicon family with seven ARGs (Table 3). However, transfer to the recipient strain was not detected—as opposed to the IncI1-associated blaCTX-M-1, sul2, and tet(A) genes—since the plasmid did not carry ESBL genes; thus, transconjugants were not selected.
Several point mutations conferring resistance to fluoroquinolones were identified in the donors H6 and H38. These included mutations in the A-subunit of the DNA gyrase as well as in the genes parC and parE, encoding subunits of the DNA topoisomerase IV (Table 3). No point mutations were detected in any of the transconjugant strains.
4. Discussion
Bacterial MLST can be used in source tracking. As opposed to extensive E. coli isolate databases revealing associations between known STs and strain origin, information about E. cloacae and C. freundii isolates is scarcer. As described in our previous work [15], the ESBL-producing E. coli STs detected in Swiss irrigation water have been detected previously in water and sewage, but also in humans, domesticated and wild animals, and different food items. As for C. freundii, the corresponding public MLST database [42] contains one entry for a ST234 strain from a diarrheal patient in China and one record for ST345 isolated from food in China. No records exist to date for C. freundii ST402. Enterobacter cloacae ST108 seems to be among the globally most widespread STs [43]. Additionally, three major lineages have been identified among MDR E. cloacae, one of which was defined as ST108-like [44]. Finally, E. cloacae ST910 has been associated with carbapenemase-carrying E. cloacae from sewage water of different Chinese hospitals [45] as well as German surface water [46]. Overall, the diversity and geographic distance of the different STs that were isolated from these sources emphasize the global distribution of these strains.
No transfer of ESBL phenotype or genes could be observed for any of the investigated E. cloacae or C. freundii strains, although known ESBL genes were detected in all but one isolate. However, no plasmid replicons could be identified, suggesting that these genes were not plasmid-borne. The blaCMY-2-like genes detected in all three C. freundii strains have been identified on plasmids previously [47]. However, they are also known to occur naturally in this species (that is, to confer intrinsic resistance), and could have been mobilized from the C. freundii chromosome onto plasmids by mediation of the insertion sequence ISEcp1 [47]. In E. cloacae strains blaACT genes were detected. Transmission of such genes between bacteria has also been described [48]. However, in Enterobacter spp. they are known to be intrinsic [49], making them unable of transferring to other bacteria unless gene mobilization takes place first.
Resistance phenotype and genotype of C. freundii and E. cloacae were in good accordance (Table 1). The ESBL phenotype relied on the presence of the above-mentioned AmpC-type ARGs. The presence of qnr genes was accompanied by phenotypic quinolone resistance. The colistin ARG mcr-9 has been documented to not always confer phenotypic resistance [40], and accordingly we did not observe any colistin resistance phenotypically. Finally, the resistance phenotype of C. freundii H42 nicely reflected all ARGs detected, including tetracycline, sulfonamide, and trimethoprim resistance. Additionally, a plasmid replicon was detected in this strain that was not, however, associated with the blaCMY gene after assembly, but with sul2 only.
Conjugation experiments were conducted on filter/solid medium as well as in liquid broth cultures, as conjugation success can vary between the two systems. We did not observe, however, a difference as the same strains conjugated under both conditions. The five E. coli strains able to transfer their ESBL phenotype by conjugation were the same five in which a known plasmid replicon was associated with the blaCTX-M gene. Plasmid-mediated transmission was further confirmed by the detection of all donor plasmid replicons associated with the blaCTX-M genes in the transconjugants, except for H6-TC. This latter displayed the lowest conjugation rates while harboring the most ARG on the blaCTX-M contig. This heavy ARG burden possibly associated with a large plasmid might be one reason for the low conjugation rates.
The phenotypic resistance profiles of donor and respective TC strains observed in broth microdilution are in good accordance with the molecular resistance profiles. Their different genetic backgrounds can explain most differences between donor and respective transconjugant strains. First, we observed intermediate to full resistance towards the tested aminoglycosides amikacin and tobramycin in most transconjugants. This resistance originates from the recipient strain E. coli CV601, which carries aph(3′)-III encoded by a gfp-cassette and conferring resistance to kanamycin and other aminoglycosides. On the other hand, the donors H6 and H38 were resistant to fluoroquinolones (CIP and LVX), which rely on point mutations in the respective strains and cannot transfer to the recipient. Finally, the H40-TC lost SXT resistance as compared to H40, since dfrA1 for trimethoprim resistance was located on the second, non-ESBL plasmid of this donor strain. Tetracycline resistance, however, transferred along with blaCTX-M-1 and was observed phenotypically in both H40 and H40-TC.
In conclusion, none of the investigated ESBL-producing E. cloacae and C. freundii strains transferred ESBL-encoding genes by conjugation, due to the intrinsic nature of these ARGs. On the other hand, we could show significant conjugative transfer of ESBL-encoding MDR plasmids from irrigation water-borne ESBL-producing E. coli to ESBL-susceptible E. coli, with conjugation in 5 out of 11 strains (up to 4.1 × 10−5 TC per recipient bacterium). These results highlight the importance of monitoring irrigation water quality for selected ARB in order to mitigate contamination of the irrigated produce with clinically significant bacteria, whose MDR plasmids can moreover spread in bacterial communities.
Author Contributions
Conceptualization, M.-T.G., A.K., K.S. and D.D.; methodology, M.-T.G., A.K. and K.S.; validation, M.-T.G. and A.K.; formal analysis, M.-T.G. and A.K.; investigation, M.-T.G. and A.K.; resources, K.S. and D.D.; data curation, M.-T.G. and A.K.; writing—original draft preparation, M.-T.G., A.K.; writing—review and editing, M.-T.G., A.K., K.S. and D.D.; visualization, M.-T.G., A.K.; supervision, M.-T.G. and D.D.; project administration, D.D.; funding acquisition, D.D. All authors have read and agreed to the published version of the manuscript.
Funding
Part of this research was funded by the National Research Program “Antimicrobial Resistance” (NRP 72, grant number 407240_167068) of the Swiss National Science Foundation.
Acknowledgments
We are grateful to Khald Blau (Julius Kühn-Institut, Braunschweig) for sharing his valuable technical skills and scientific discussions. We thank Weihong Qi (Functional Genomics Center Zurich) for bioinformatics assistance.
Conflicts of Interest
The authors declare no conflicts of interest.
Appendix A

{kind=link}
{kind=link}
{kind=link}
Table A1.
Repetitive sequence-based PCR fingerprinting (REP-PCR) cycling conditions. Used for confirming E. coli CV601 background in putative transconjugants using the (GTG)5 primer (5′-GTGGTGGTGGTGGTG-3). PCRs were performed using the HotStarTaq® Master Mix Kit (Qiagen, Hilden, Germany) in 25 µL reaction volumes.
Table A1.
Repetitive sequence-based PCR fingerprinting (REP-PCR) cycling conditions. Used for confirming E. coli CV601 background in putative transconjugants using the (GTG)5 primer (5′-GTGGTGGTGGTGGTG-3). PCRs were performed using the HotStarTaq® Master Mix Kit (Qiagen, Hilden, Germany) in 25 µL reaction volumes.
| Cycling Step | PCR Conditions | ||
|---|---|---|---|
| Enzyme activation | 95 °C, 5 min |  | |
| Denaturation | 94 °C, 3 s | ||
| 92 °C, 30 s | 30 cycles | ||
| Annealing | 40 °C, 1 min | ||
| Extension | 65 °C, 8 min | ||
| Final extension | 65 °C, 16 min | ||
References
- Kupferschmidt, K. Resistance fighters. Science 2016, 352, 758–761. [Google Scholar] [CrossRef] [Green Version]
- Morar, M.; Wright, G.D. The genomic enzymology of antibiotic resistance. Annu. Rev. Genet. 2010, 44, 25–51. [Google Scholar] [CrossRef] [PubMed]
- Svara, F.; Rankin, D.J. The evolution of plasmid-carried antibiotic resistance. BMC Evol. Biol. 2011, 11, 130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, C.M.; Nielsen, K.M. Mechanisms of, and barriers to, horizontal gene transfer between bacteria. Nat. Rev. Microbiol. 2005, 3, 711–721. [Google Scholar] [CrossRef] [PubMed]
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [Green Version]
- Centers for Disease Control and Prevention (CDC): Antibiotic/Antimicrobial Resistance—Biggest Threats; c2019. Available online: https://www.cdc.gov/drugresistance/biggest_threats.html (accessed on 29 June 2020).
- Ambler, R.P. The structure of beta-lactamases. Philos. Trans. R. Soc. Lond. 1980, 289, 321–331. [Google Scholar]
- Rawat, D.; Nair, D. Extended-spectrum β-lactamases in Gram Negative Bacteria. J. Glob. Infect. Dis. 2010, 2, 263–274. [Google Scholar] [CrossRef] [PubMed]
- Dahmen, S.; Métayer, V.; Gay, E.; Madec, J.-Y.; Haenni, M. Characterization of extended-spectrum beta-lactamase (ESBL)-carrying plasmids and clones of Enterobacteriaceae causing cattle mastitis in France. Vet. Microbiol. 2013, 162, 793–799. [Google Scholar] [CrossRef]
- Ewers, C.; Grobbel, M.; Bethe, A.; Wieler, L.H.; Guenther, S. Extended-spectrum beta-lactamases-producing Gram-negative bacteria in companion animals: Action is clearly warranted. Berl. Munch. Tierarztl. Wochenschr. 2011, 124, 4–101. [Google Scholar]
- Nüesch-Inderbinen, M.; Zurfluh, K.; Peterhans, S.; Hächler, H.; Stephan, R. Assessment of the prevalence of extended-spectrum beta-lactamase-producing Enterobacteriaceae in ready-to-eat salads, fresh-cut fruit, and sprouts from the Swiss market. J. Food Prot. 2015, 78, 1178–1181. [Google Scholar] [CrossRef]
- Szmolka, A.; Nagy, B. Multidrug resistant commensal Escherichia coli in animals and its impact for public health. Front. Microbiol. 2013, 4, 258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geser, N.; Stephan, R.; Hächler, H. Occurrence and characteristics of extended-spectrum β-lactamase (ESBL) producing Enterobacteriaceae in food producing animals, minced meat and raw milk. BMC Vet. Res. 2012, 8, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reuland, E.A.; al Naiemi, N.; Raadsen, S.A.; Savelkoul, P.H.M.; Kluytmans, J.A.J.W.; Vandenbroucke-Grauls, C.M.J.E. Prevalence of ESBL-producing Enterobacteriaceae in raw vegetables. Eur. J. Clin. Microbiol. Infect. Dis. 2014, 33, 1843–1846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gekenidis, M.T.; Qi, W.; Hummerjohann, J.; Zbinden, R.; Walsh, F.; Drissner, D. Antibiotic-resistant indicator bacteria in irrigation water: High prevalence of extended-spectrum beta-lactamase (ESBL)-producing Escherichia coli. PLoS ONE 2018, 13, e0207857. [Google Scholar] [CrossRef]
- Zurfluh, K.; Hächler, H.; Nüesch-Inderbinen, M.; Stephan, R. Characteristics of extended-spectrum beta-lactamase- and carbapenemase-producing Enterobacteriaceae isolates from rivers and lakes in Switzerland. Appl. Environ. Microbiol. 2013, 79, 3021–3026. [Google Scholar] [CrossRef] [Green Version]
- Graham, D.W.; Knapp, C.W.; Christensen, B.T.; McCluskey, S.; Dolfing, J. Appearance of β-lactam Resistance Genes in Agricultural Soils and Clinical Isolates over the 20th Century. Sci. Rep. 2016, 6, 21550. [Google Scholar] [CrossRef] [Green Version]
- Gatica, J.; Yang, K.; Pagaling, E.; Jurkevitch, E.; Yan, T.; Cytryn, E. Resistance of Undisturbed Soil Microbiomes to Ceftriaxone Indicates Extended Spectrum β-Lactamase Activity. Front. Microbiol. 2015, 6, 1233. [Google Scholar] [CrossRef] [Green Version]
- Guenther, S.; Ewers, C.; Wieler, L.H. Extended-spectrum beta-lactamases producing E. coli in wildlife, yet another form of environmental pollution? Front. Microbiol. 2011, 2, 246. [Google Scholar] [CrossRef] [Green Version]
- Cantón, R.; González-Alba, J.M.; Galán, J.C. CTX-M Enzymes: Origin and Diffusion. Front. Microbiol. 2012, 3, 110. [Google Scholar] [CrossRef] [Green Version]
- Reid, C.J.; Blau, K.; Jechalke, S.; Smalla, K.; Djordjevic, S.P. Whole Genome Sequencing of Escherichia coli From Store-Bought Produce. Front. Microbiol. 2020, 10, 3050. [Google Scholar] [CrossRef]
- Pesavento, G.; Calonico, C.; Ducci, B.; Magnanini, A.; Lo Nostro, A. Prevalence and antibiotic resistance of Enterococcus spp. isolated from retail cheese, ready-to-eat salads, ham, and raw meat. Food Microbiol. 2014, 41, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Gekenidis, M.T.; Schöner, U.; von Ah, U.; Schmelcher, M.; Walsh, F.; Drissner, D. Tracing back multidrug-resistant bacteria in fresh herb production: From chive to source through the irrigation water chain. FEMS Microbiol. Ecol. 2018, 94, fiy149. [Google Scholar] [CrossRef] [PubMed]
- Vital, P.G.; Caballes, M.B.D.; Rivera, W.L. Antimicrobial resistance in Escherichia coli and Salmonella spp. isolates from fresh produce and the impact to food safety. J. Environ. Sci. Health B 2017, 52, 683–689. [Google Scholar] [CrossRef] [PubMed]
- Gekenidis, M.T.; Gossin, D.; Schmelcher, M.; Schöner, U.; Remus-Emsermann, M.N.P.; Drissner, D. Dynamics of culturable mesophilic bacterial communities of three fresh herbs and their production environment. J. Appl. Microbiol. 2017, 123, 916–932. [Google Scholar] [CrossRef] [Green Version]
- Steele, M.; Odumeru, J. Irrigation water as source of foodborne pathogens on fruit and vegetables. J. Food Prot. 2004, 67, 2839–2849. [Google Scholar] [CrossRef]
- Czekalski, N.; Díez, E.G.; Bürgmann, H. Wastewater as a point source of antibiotic-resistance genes in the sediment of a freshwater lake. ISME J. 2014, 8, 1381–1390. [Google Scholar] [CrossRef]
- Liu, X.; Liang, C.; Liu, X.; Zhao, F.; Han, C. Occurrence and human health risk assessment of pharmaceuticals and personal care products in real agricultural systems with long-term reclaimed wastewater irrigation in Beijing, China. Ecotoxicol. Environ. Saf. 2020, 190, 110022. [Google Scholar] [CrossRef]
- Chen, Q.-L.; Cui, H.-L.; Su, J.-Q.; Penuelas, J.; Zhu, Y.-G. Antibiotic resistomes in plant microbiomes. Trends Plant. Sci. 2019, 24, 530–541. [Google Scholar] [CrossRef]
- SwissGAP: Risikoanalyse Bewässerungswasser (2017); c2018. Available online: https://agrosolution.ch/swissgap/ (accessed on 29 June 2020).
- Heuer, H.; Krögerrecklenfort, E.; Wellington, E.M.; Egan, S.; van Elsas, J.D.; van Overbeek, L.; Collard, J.M.; Guillaume, G.; Karagouni, A.D.; Nikolakopoulou, T.L.; et al. Gentamicin resistance genes in environmental bacteria: Prevalence and transfer. FEMS Microbiol. Ecol. 2002, 42, 289–302. [Google Scholar] [CrossRef]
- Gekenidis, M.-T.; Studer, P.; Wüthrich, S.; Brunisholz, R.; Drissner, D. Beyond the matrix-assisted laser desorption ionization (MALDI) biotyping workflow: In search of microorganism-specific tryptic peptides enabling discrimination of subspecies. Appl. Environ. Microbiol. 2014, 80, 4234–4241. [Google Scholar] [CrossRef] [Green Version]
- Larsen, M.V.; Cosentino, S.; Rasmussen, S.; Friis, C.; Hasman, H.; Marvig, R.L.; Jelsbak, L.; Pontén, T.S.; Ussery, D.W.; Aarestrup, F.M. Multilocus sequence typing of total genome sequenced bacteria. J. Clin. Microbiol. 2012, 50, 1355–1361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaas, R.S.; Leekitcharoenphon, P.; Aarestrup, F.M.; Lund, O. Solving the problem of comparing whole bacterial genomes across different sequencing platforms. PLoS ONE 2014, 9, e104984. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A. FigTree v1.4.4, a Graphical Viewer of Phylogenetic Trees, 2018. 2014 Java. Available online: https://github.com/rambaut/figtree (accessed on 29 June 2020).
- Zankari, E.; Hasman, H.; Cosentino, S.; Vestergaard, M.; Rasmussen, S.; Lund, O.; Aarestrup, F.M.; Larsen, M.V. Identification of acquired antimicrobial resistance genes. J. Antimicrob. Chemother. 2012, 67, 2640–2644. [Google Scholar] [CrossRef]
- Carattoli, A.; Zankari, E.; Garcìa-Fernandez, A.; Larsen, M.V.; Lund, O.; Villa, L.; Aarestrup, F.M.; Hasman, H. PlasmidFinder and pMLST: In silico detection and typing of plasmids. Antimicrob. Agents Chemother. 2014, 58, 3895–3903. [Google Scholar] [CrossRef] [Green Version]
- Blau, K.; Bettermann, A.; Jechalke, S.; Fornefeld, E.; Vanrobaeys, Y.; Stalder, T.; Top, E.M.; Smalla, K. The Transferable Resistome of Produce. mBio 2018, 9, e01300–e01318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohapatra, B.R.; Mazumder, A. Comparative efficacy of five different rep-PCR methods to discriminate Escherichia coli populations in aquatic environments. Water Sci. Technol. 2008, 58, 537–547. [Google Scholar] [CrossRef] [PubMed]
- Bortolaia, V. Genomic Epidemiology—ResFinder 4 database 2020. Available online: https://bitbucket.org/genomicepidemiology/resfinder_db/src/master/phenotypes.txt (accessed on 29 June 2020).
- Kaase, M.; Szabados, F.; Anders, A.; Gatermann, S.G. Fosfomycin susceptibility in carbapenem-resistant Enterobacteriaceae from Germany. J. Clin. Microbiol. 2014, 52, 1893–1897. [Google Scholar] [CrossRef] [Green Version]
- Jolley, K.A.; Bray, J.E.; Maiden, M.C.J. Open-access bacterial population genomics: BIGSdb software, the PubMLST.org website and their applications. Wellcome Open Res. 2018, 3, 124. [Google Scholar] [CrossRef]
- Izdebski, R.; Baraniak, A.; Herda, M.; Fiett, J.; Bonten, M.J.M.; Carmeli, Y.; Goossens, H.; Hryniewicz, W.; Brun-Buisson, C.; Gniadkowski, M. MLST reveals potentially high-risk international clones of Enterobacter cloacae. J. Antimicrob. Chemoth. 2014, 70, 48–56. [Google Scholar] [CrossRef] [Green Version]
- Girlich, D.; Poirel, L.; Nordmann, P. Clonal distribution of multidrug-resistant Enterobacter cloacae. Diagnost. Microbiol. Infect. Dis. 2015, 81, 264–268. [Google Scholar] [CrossRef] [Green Version]
- Jin, L.; Wang, R.; Wang, X.; Wang, Q.; Zhang, Y.; Yin, Y.; Wang, H. Emergence of mcr-1 and carbapenemase genes in hospital sewage water in Beijing, China. J. Antimicrob. Chemother. 2017, 73, 84–87. [Google Scholar] [CrossRef] [PubMed]
- Falgenhauer, L.; Schwengers, O.; Schmiedel, J.; Baars, C.; Lambrecht, O.; Heß, S.; Berendonk, T.U.; Falgenhauer, J.; Chakraborty, T.; Imirzalioglu, C. Multidrug-Resistant and Clinically Relevant Gram-Negative Bacteria Are Present in German Surface Waters. Front. Microbiol 2019, 10, 2779. [Google Scholar] [CrossRef] [PubMed]
- Verdet, C.; Gautier, V.; Chachaty, E.; Ronco, E.; Hidri, N.; Decré, D.; Arlet, G. Genetic Context of Plasmid-Carried blaCMY-2-Like Genes in Enterobacteriaceae. Antimicrob. Agents Chemother. 2009, 53, 4002–4006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ku, Y.-H.; Lee, M.-F.; Chuang, Y.-C.; Yu, W.-L. Detection of Plasmid-Mediated β-Lactamase Genes and Emergence of a Novel AmpC (CMH-1) in Enterobacter cloacae at a Medical Center in Southern Taiwan. J. Clin. Med. 2018, 8, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piotrowska, M.; Kowalska, S.; Popowska, M. Diversity of β-lactam resistance genes in Gram-negative rods isolated from a municipal wastewater treatment plant. Ann. Microbiol. 2019, 69, 591–601. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Phylogenetic trees of ESBL-producing (A) C. freundii complex, (B) E. cloacae complex, and (C) E. coli strains based on single-nucleotide polymorphisms. Reference strains in red. ST, sequence type; for strains with unknown ST, the nearest known ST is marked by an asterisk (*) (illustration: FigTree v1.4.4).
Figure 1.
Phylogenetic trees of ESBL-producing (A) C. freundii complex, (B) E. cloacae complex, and (C) E. coli strains based on single-nucleotide polymorphisms. Reference strains in red. ST, sequence type; for strains with unknown ST, the nearest known ST is marked by an asterisk (*) (illustration: FigTree v1.4.4).


Figure 2.
Plasmid transfer rates from ESBL-producing E. coli donor strains to a susceptible recipient strain (E. coli CV601) in filter conjugation assays (n = 3). Transconjugant (TC) numbers are normalized by the number of recipient bacteria per assay. Mean and standard error (SEM) are shown.
Figure 2.
Plasmid transfer rates from ESBL-producing E. coli donor strains to a susceptible recipient strain (E. coli CV601) in filter conjugation assays (n = 3). Transconjugant (TC) numbers are normalized by the number of recipient bacteria per assay. Mean and standard error (SEM) are shown.

Table 1.
Antibiotic resistance phenotype, resistance genes, and plasmid replicons in ESBL-producing E. cloacae and C. freundii complex strains from irrigation water. Full phenotypic resistance (R) shown in bold, intermediate phenotypic resistance (I) in plain font; % ID, percent identical bases between query and sample sequence; HSP/query, alignment length of sample to query sequence. AM: ampicillin; TEMO: temocillin; AMC: amoxicillin–clavulanic acid; TPZ: piperacillin–tazobactam; KF: cefalotin; FOX: cefoxitin; CXM: cefuroxime; CPD: cefpodoxime; CRO: ceftriaxone; CTX: cefotaxime; CAZ: ceftazidime; FEP: cefepime; ETP: ertapenem; TE: tetracycline; MI: minocycline; NA: nalidixic acid; TMP: trimethoprim; SMZ: sulfonamide; SXT: trimethoprim–sulfamethoxazole; F100: nitrofurantoin.
Table 1.
Antibiotic resistance phenotype, resistance genes, and plasmid replicons in ESBL-producing E. cloacae and C. freundii complex strains from irrigation water. Full phenotypic resistance (R) shown in bold, intermediate phenotypic resistance (I) in plain font; % ID, percent identical bases between query and sample sequence; HSP/query, alignment length of sample to query sequence. AM: ampicillin; TEMO: temocillin; AMC: amoxicillin–clavulanic acid; TPZ: piperacillin–tazobactam; KF: cefalotin; FOX: cefoxitin; CXM: cefuroxime; CPD: cefpodoxime; CRO: ceftriaxone; CTX: cefotaxime; CAZ: ceftazidime; FEP: cefepime; ETP: ertapenem; TE: tetracycline; MI: minocycline; NA: nalidixic acid; TMP: trimethoprim; SMZ: sulfonamide; SXT: trimethoprim–sulfamethoxazole; F100: nitrofurantoin.
| Strain ID | Antibiotic Resistance Phenotypes (R and I) | Acquired Antibiotic Resistance Genes | Plasmid Replicons (% ID; HSP/Query) |
|---|---|---|---|
| Ent.H9 | AM, TEMO, AMC, TPZ, KF, FOX, CXM, CPD, CRO, CTX, CAZ, ETP, SMZ, SXT | blaACT-9, fosA†, mcr-9 | none detected |
| Ent.H16 | AM, TEMO, AMC, TPZ, KF, FOX, CXM, CPD, CRO, CTX, CAZ, ETP, F100 | blaACT-12, fosA2† | none detected |
| Ent.H24 | AM, TEMO, AMC, TPZ, KF, FOX, CXM, CPD, CRO, CTX, CAZ, ETP, F100 | blaACT-12, fosA2† | none detected |
| Ent.H34 | AM, TEMO, AMC, TPZ, KF, FOX, CXM, CPD, CRO, CTX, CAZ, ETP, F100 | blaACT-7 | none detected |
| Ent.H36 | AM, TEMO, AMC, TPZ, KF, FOX, CXM, CPD, CRO, CTX, CAZ, ETP, MI, NA, F100 | fosA†, qnrE1 | none detected |
| Citr.H12 | AM, TEMO, AMC, TPZ, KF, FOX, CXM, CPD, CRO, CTX, CAZ, FEP, NA, F100 | blaCMY-83 | none detected |
| Citr.H41 | AM, TEMO, AMC, TPZ, KF, FOX, CXM, CPD, CRO, CTX, CAZ, FEP, ETP | blaCMY-101 | none detected |
| Citr.H42 | AM, AMC, TPZ, KF, FOX, CXM, CPD, CRO, CTX, CAZ, TE, MI, NA, TMP, SMZ, SXT | blaTEM-1B, blaCMY-48, tet(B), qnrB, dfrA17, sul1, sul2, aadA5 ˟, catA1 ˟ | IncQ1 (100; 529/796) * |
* truncated form of IncQ1, associated with sul2; ˟ resistance towards streptomycin and chloramphenicol was not tested in disk diffusion assays; † resistance towards fosfomycin cannot be determined reliably in disk diffusion assays [41].
Table 2.
Antibiotic resistance phenotypes of recipient strains E. coli CV601, ESBL-producing E. coli, and their respective transconjugants (TCs) obtained from filter conjugation, determined using broth microdilution. S (sensitive), I (intermediate), and R (resistant) phenotypes are based on EUCAST cut-off values. +, ESBL confirmation; AMK, amikacin; AMC, amoxicillin–clavulanic acid; SAM, ampicillin–sulbactam; AM, ampicillin; ATM, aztreonam; FEP, cefepime; CTX, cefotaxime; CAZ, ceftazidime; CXM, cefuroxime; CIP, ciprofloxacin; LVX, levofloxacin; MI, minocycline; PIP, piperacillin; TE, tetracycline; TOB, tobramycin; SXT, trimethoprim–sulfamethoxazole.
Table 2.
Antibiotic resistance phenotypes of recipient strains E. coli CV601, ESBL-producing E. coli, and their respective transconjugants (TCs) obtained from filter conjugation, determined using broth microdilution. S (sensitive), I (intermediate), and R (resistant) phenotypes are based on EUCAST cut-off values. +, ESBL confirmation; AMK, amikacin; AMC, amoxicillin–clavulanic acid; SAM, ampicillin–sulbactam; AM, ampicillin; ATM, aztreonam; FEP, cefepime; CTX, cefotaxime; CAZ, ceftazidime; CXM, cefuroxime; CIP, ciprofloxacin; LVX, levofloxacin; MI, minocycline; PIP, piperacillin; TE, tetracycline; TOB, tobramycin; SXT, trimethoprim–sulfamethoxazole.
| Antibiotic Resistant Phenotypes | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Strain ID | AMK | AMC | SAM | AM | ATM | FEP | CTX | CAZ | CXM | CIP | LVX | MI | PIP | TE | TOB | SXT |
| E. coli CV601 | I | S | S | S | S | S | S | S | S | S | S | S | S | S | I | S |
| H6 | S | S | R | R | + | R | + | + | R | R | R | S | R | R | S | R |
| H6-TC | S | S | R | R | + | R | + | + | R | S | S | S | R | R | S | R |
| H22 | S | R | R | R | + | R | + | + | R | S | S | I | R | R | S | S |
| H22-TC | I | R | R | R | + | R | + | + | R | S | S | I | R | R | I | S |
| H38 | S | S | R | R | + | R | + | + | R | R | R | S | R | S | S | S |
| H38-TC | I | S | R | R | + | R | + | + | R | S | S | S | R | S | S | S |
| H40 | S | R | R | R | + | R | + | + | R | S | S | I | R | R | S | R |
| H40-TC | I | R | R | R | + | R | + | + | R | S | S | S | R | R | I | S |
| H45 | S | R | R | R | + | R | + | + | R | S | S | S | R | S | S | S |
| H45-TC | R | R | R | R | + | R | + | + | R | S | S | S | R | S | R | S |
Table 3.
Antibiotic resistance genotype including plasmid replicons in ESBL-producing E. coli and their respective transconjugants obtained from filter conjugation. All detected plasmid replicons were full length.
Table 3.
Antibiotic resistance genotype including plasmid replicons in ESBL-producing E. coli and their respective transconjugants obtained from filter conjugation. All detected plasmid replicons were full length.
| ESBL-Producing Donors | Transconjugants | ||||||
|---|---|---|---|---|---|---|---|
| ID | Acquired ARGs (Grouped by Contig) | Plasmid Replicons (% ID) | Point Mutations | ID | Acquired ARGs (Grouped by Contig) | Plasmid Replicons (% ID) | Point Mutations |
| H6 | I. strA, strB, aadA5, blaCTX-M-27, mph(A), sul1, sul2, tet(A), dfrA17 | I. IncFIA (99.74), IncFIB (96.63) | parE p.L416F parC p.S80I gyrA p.S83L gyrA p.D87N | H6-TC | I. strA, strB, aadA5, blaCTX-M-27, mph(A), sul1, sul2, tet(A), dfrA17 II. aph(3′)-III | none detected | none detected |
| H22 | I. blaCTX-M-1, sul2, tet(A) | I. IncI1 (98.59) | none detected | H22-TC | I. blaCTX-M-1, sul2, tet(A) II. aph(3′)-III | I. IncI1 (99.3) | none detected |
| H38 | I. blaCTX-M-15, mph(A) | I. IncFIB (98.39) | parE p.I529L parC p.S80I parC p.E84V gyrA p.S83L gyrA p.D87N | H38-TC | I. blaCTX-M-15, mph(A) II. aph(3′)-III | I. IncFIB (98.39) | none detected |
| H40 | I. blaCTX-M-1, sul2, tet(A) II. strA, strB, aadA1, sul1, sul2, tet(A), dfrA1 | I. IncI1 (98.59) II. IncFIB (97.07), IncFIC (95.59) | none detected | H40-TC | I. blaCTX-M-1, sul2, tet(A) II. aph(3′)-III | I. IncI1 (98.59) | none detected |
| H45 | I. blaCTX-M-1 | I. IncI1 (98.59) | none detected | H45-TC | I. blaCTX-M-1 II. aph(3′)-III | I. IncI1 (99.3) | none detected |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Gekenidis, M.-T.; Kläui, A.; Smalla, K.; Drissner, D. Transferable Extended-Spectrum β-Lactamase (ESBL) Plasmids in Enterobacteriaceae from Irrigation Water. Microorganisms 2020, 8, 978. https://doi.org/10.3390/microorganisms8070978
AMA Style
Gekenidis M-T, Kläui A, Smalla K, Drissner D. Transferable Extended-Spectrum β-Lactamase (ESBL) Plasmids in Enterobacteriaceae from Irrigation Water. Microorganisms. 2020; 8(7):978. https://doi.org/10.3390/microorganisms8070978
Chicago/Turabian StyleGekenidis, Maria-Theresia, Anita Kläui, Kornelia Smalla, and David Drissner. 2020. "Transferable Extended-Spectrum β-Lactamase (ESBL) Plasmids in Enterobacteriaceae from Irrigation Water" Microorganisms 8, no. 7: 978. https://doi.org/10.3390/microorganisms8070978
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.