Reorganized Genomic Taxonomy of Francisellaceae Enables Design of Robust Environmental PCR Assays for Detection of Francisella tularensis

 , , , , , ,
, , , , , ,  , , , and add
Show full author list
, , , and add
Show full author list
Abstract
:1. Introduction
2. Materials and Methods
2.1. Library Preparation
2.2. Genome Sequencing
2.3. Genome Assembly
2.4. Genome Sequences
2.5. Phylogenetic Trees
2.6. Average Nucleotide Identity
2.7. Genome Taxonomy of Francisellaceae
2.8. Identification of Clade-Specific Regions
2.9. Design of TaqMan qPCR Assays
2.10. In Silico Evaluation of Francisella Primers
2.11. Validation of Francisella tularensis Specific Assays
2.12. Rationale for Francisella Panel Selection
3. Results
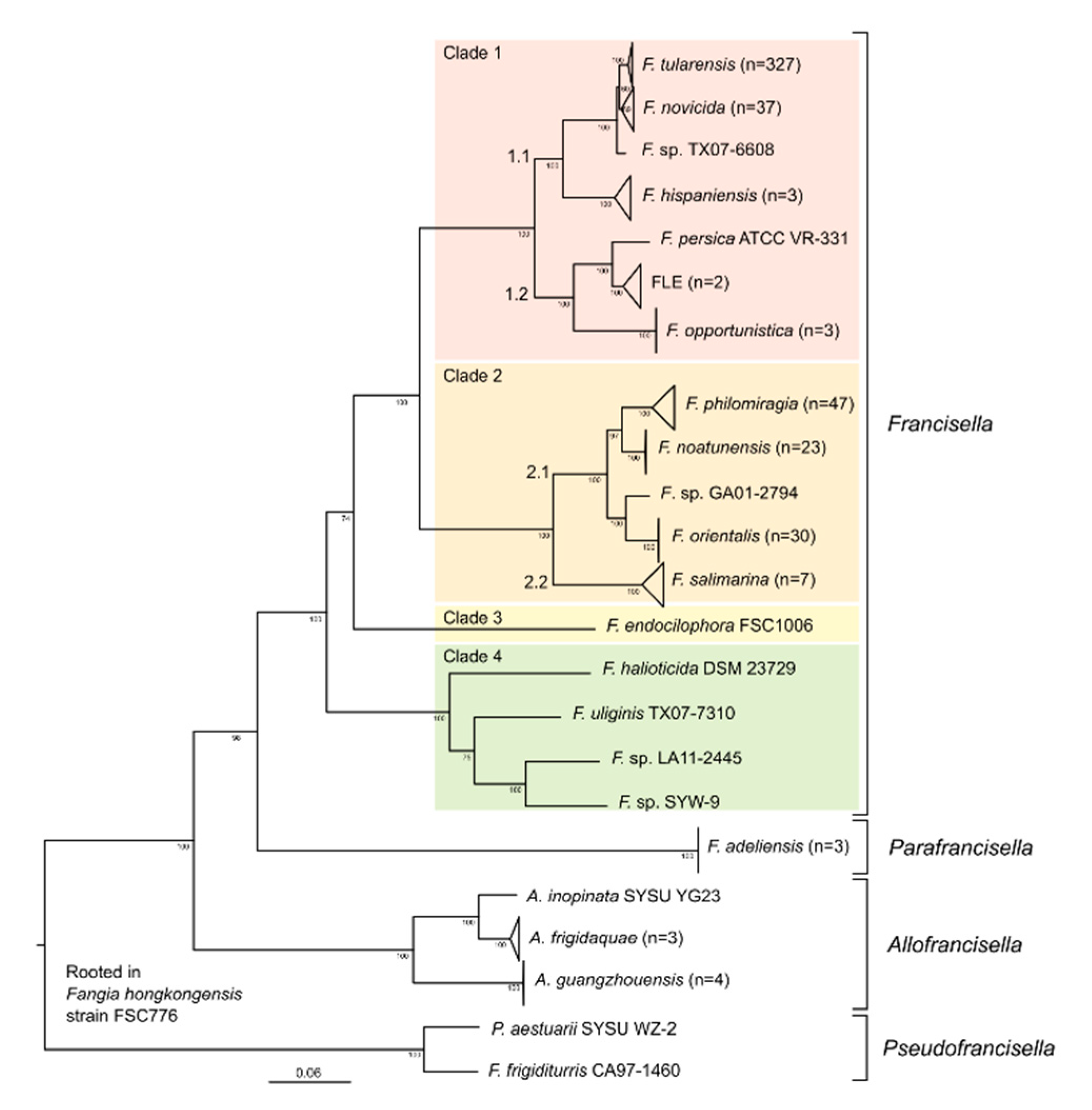
3.1. Organisation of Genera and Species Based on Whole-Genome Analysis
3.2. Phylogenetic Tree
3.3. Average Nucleotide Identity
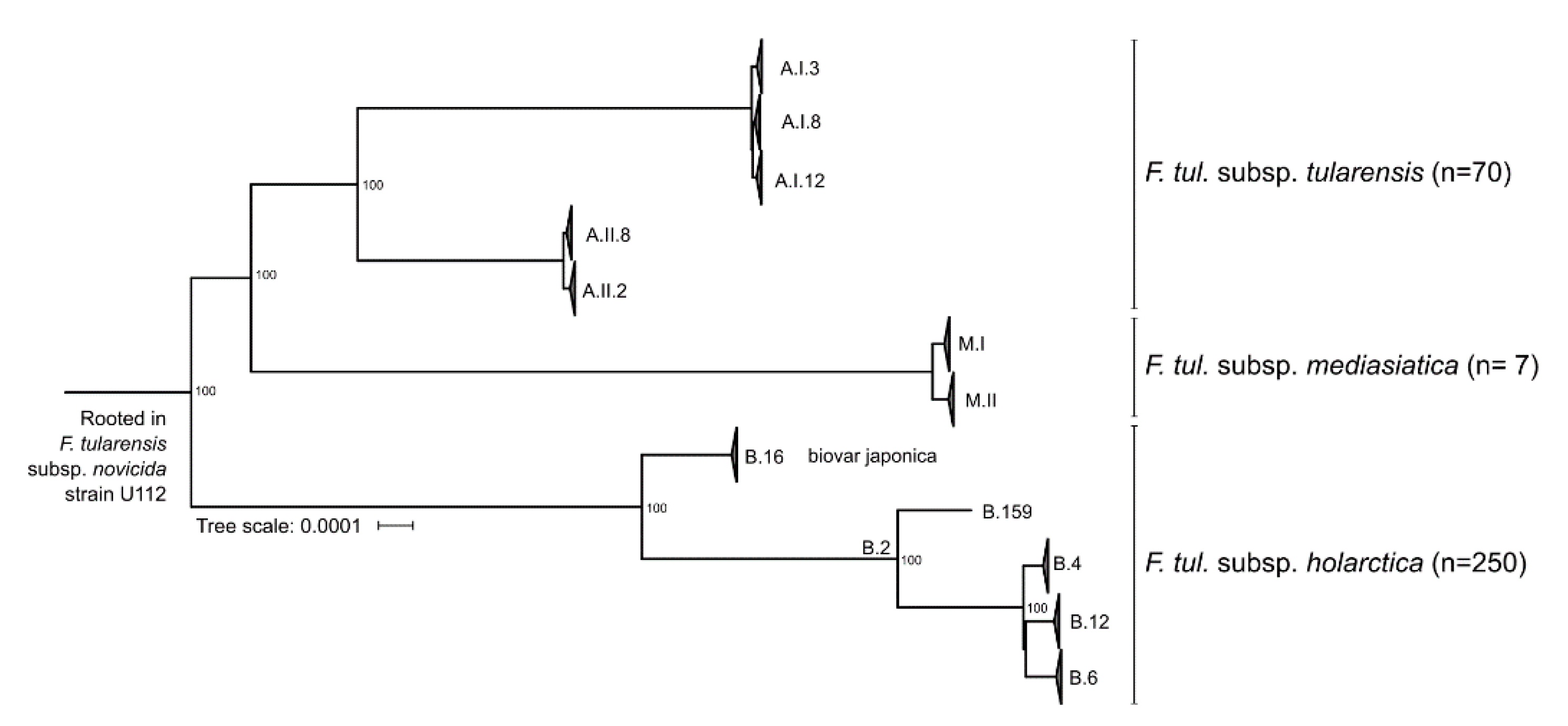
3.4. Subspecies and Major Genotypes within Francisella tularensis
3.5. Francisella Tularensis Specific Genomic Regions
3.6. Genomic Regions Specific to Individual Species
3.7. Genomic Regions Specific to Francisella Tularensis Subspecies and Major Phylogentic Groups
3.8. In Silico Evaluation of PCR Assay Specificity
3.9. Validation of Francisella tularensis Specific Assays
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Francisellaceae genus | Francisellaceae species | Heterotypic Synonyms | Type Strains | References |
|---|---|---|---|---|
| Francisella | Francisella tularensis | [93] | ||
| Francisella tularensis subspecies tularensis | SchuS4 | [17,42,94,95] | ||
| Francisella tularensis subspecies holarctica | Francisella tularensis var. palaearctica | FSC257; GIEM 503 | [17,95,96] | |
| FSC200 | ||||
| Francisella tularensis subspecies mediasiatica | FSC147 | [41,95,97,98] | ||
| Francisella tularensis subspecies novicida | F. novicida, Pasteurella novicida | ATCC 15482; CCUG 33449; CIP 56.12 | [15,17,19] | |
| U112 | ||||
| Francisella hispaniensis | CCUG 58020; DSM 22475; F62; FhSp1; FnSp1; FSC454 | [19] | ||
| Francisella persica (FLE) | Wolbachia persica | ATCC VR-331; DSM 101678; FSC845 | [22,99,100,101] | |
| Francisella opportunistica | ATCC BAA-2974; DSM 107100; PA05-1188 | [23,102,103] | ||
| Francisella philomiragia | F. philomiragia subspecies philomiragia, Yersinia philomiragia | ATCC 25015; CCUG 19700; CCUG 4992; CIP 82.98; DSM 7535 | [18,82,104] | |
| Francisella noatunensis | F. piscicida; F. philomiragia subspecies noatunensis Francisella noatunensis subspecies noatunensis, | 2005/50/F292-6; DSM 23596; LMG 23800; NCIMB 14265 | [11,12,13,105] | |
| Francisella orientalis | F. victoria, F. asiatica, Francisella noatunensis subspecies orientalis | DSM 21254; Ehime-1; LMG 24544 | [11,12,13] | |
| CCUG 56120T; DSM 21294; PQ 1104 | ||||
| Francisella salina | TX07-7308 | [23,66] | ||
| Francisella marina | E103-15, E95-16 | [9] | ||
| Francisella salimarina | CGMCC 1.17031; NBRC 113781; SYSU SYW-1 | [83] | ||
| Francisella endociliophora | F. noatunensis subspecies endociliophora | [20,86] | ||
| Francisella halioticida | DSM 23729; LMG 26062; Shimane-1 | [10,26] | ||
| Francisella sp. LA11-2445 | LA11-2445 | [25] | ||
| Francisella uliginis | TX07-7310 | [23,66] | ||
| Francisella adeliensis | FSC1327 | [21] | ||
| Francisella frigiditurris | CA97-1460 | [23] | ||
| Allofrancisella | Allofrancisella inopinata | DSM 101834; KCTC 42968; SYSU 23; SYSU YG23 | [1,27] | |
| Allofrancisella frigidaquae | DSM 101835; KCTC 42969; SYSU 10HL1970 | [1,24,27] | ||
| Allofrancisella guanzhousensis | F. guanzhousensis, F. cantonensis | 08HL01032; ATCC BAA-2361; CCUG 60119; NCTC 13503 | [1,31,106] | |
| Pseudofrancisella | Pseudofrancisella aestuarii | CGMCC 1.13718; KCTC 52557; SYSU WZ-2 | [2] |

References
- Qu, P.-H.; Li, Y.; Salam, N.; Chen, S.; Liu, L.; Gu, Q.; Fang, B.-Z.; Xiao, M.; Li, M.; Chen, C.; et al. Allofrancisella inopinata gen. nov., sp. nov. and Allofrancisella frigidaquae sp. nov., isolated from water-cooling systems, and transfer of Francisella guangzhouensis Qu et al. 2013 to the new genus as Allofrancisella guangzhouensis comb. nov. Int. J. Syst. Evol. Microbiol. 2016, 66, 4832–4838. [Google Scholar] [CrossRef] [PubMed]
- Zheng, M.-L.; Jiao, J.-Y.; Dong, L.; Han, M.-X.; Li, L.-H.; Xiao, M.; Chen, C.; Qu, P.-H.; Li, W.-J. Pseudofrancisella aestuarii gen. nov., sp. nov., a novel member of the family Francisellaceae isolated from estuarine seawater. Antonie Van Leeuwenhoek 2019, 112, 877–886. [Google Scholar] [CrossRef] [PubMed]
- Colquhoun, D.J.; Larsson, P.; Duodu, S.; Forsman, M. The Family Francisellaceae. Prokaryotes 2014, 287–314. Available online: https://link.springer.com/referenceworkentry/10.1007/978-3-642-38922-1_236 (accessed on 1 December 2020).
- Penn, R.L. Francisella tularensis (Tularemia). In Mandell, Douglas, and Bennett’s Principles and Practice of Infectious Diseases; Elsevier: Amsterdam, The Netherlands, 2010; pp. 2927–2937. [Google Scholar]
- Timofeev, V.; Bakhteeva, I.; Titareva, G.; Kopylov, P.; Christiany, D.; Mokrievich, A.; Dyatlov, I.; Vergnaud, G. Russian isolates enlarge the known geo-graphic diversity of Francisella tularensis subsp. mediasiatica. PLoS ONE 2017, 12, e0183714. Available online: http://www.ncbi.nlm.nih.gov/pubmed/28873421 (accessed on 1 November 2020). [CrossRef] [PubMed] [Green Version]
- Timofeev, V.; Titareva, G.; Bahtejeva, I.; Kombarova, T.; Kravchenko, T.; Mokrievich, A.; Dyatlov, I. The Comparative Virulence of Fran-cisella tularensis Subsp. mediasiatica for Vaccinated Laboratory Animals. Microorganisms 2020, 8, 1403. Available online: https://www.mdpi.com/2076-2607/8/9/1403 (accessed on 1 November 2020). [CrossRef] [PubMed]
- Dennis, D.T.; Inglesby, T.V.; Henderson, D.A.; Bartlett, J.G.; Ascher, M.S.; Eitzen, E.; Fine, A.D.; Friedlander, A.M.; Hauer, J.; Layton, M.; et al. Tularemia as a biological weapon: Medical and public health management. JAMA Am. Med Assoc. 2001, 285, 2763–2773. Available online: http://jama.ama-assn.org/content/285/21/2763.abstract (accessed on 1 November 2020). [CrossRef] [PubMed]
- Maurin, M. Francisella tularensis as a potential agent of bioterrorism? Expert Rev. Anti. Infect. Ther. 2015, 13, 141–144. Available online: https://pubmed.ncbi.nlm.nih.gov/25413334/ (accessed on 1 November 2020).
- Soto, E.; Griffin, M.J.; Morales, J.A.; Calvo, E.B.; Sebastião, F.D.A.; Porras, A.L.; Víquez-Rodríguez, X.; Reichley, S.R.; Rosser, T.G.; Ware, C.; et al. Francisella marinasp. nov., Etiologic Agent of Systemic Disease in Cultured Spotted Rose Snapper (Lutjanus guttatus) in Central America. Appl. Environ. Microbiol. 2018, 84, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Brevik, Ø.J.; Ottem, K.; Kamaishi, T.; Watanabe, K.; Nylund, A. Francisella halioticida sp. nov., a pathogen of farmed giant abalone (Haliotis gigantea) in Japan. J. Appl. Microbiol. 2011, 111, 1044–1056. [Google Scholar] [CrossRef]
- Ottem, K.; Nylund, A.; Karlsbakk, E.; Friis-Møller, A.; Kamaishi, T. Elevation of Francisella philomiragia subsp. noatunensis Mikalsen et al. (2007) to Francisella noatunensis comb. nov.[syn. Francisella piscicida Ottem et al.(2008) syn. nov.] and characterization of Francisella noatunensis subsp. orientalis subsp. nov., two important fish pathogens. J. Appl. Microbiol. 2009, 106, 1231–1243. Available online: http://www3.interscience.wiley.com/cgi-bin/abstract/121660927/ABSTRACT (accessed on 1 November 2020).
- Mikalsen, J.; Colquhoun, D.J. Francisella asiatica sp. nov. isolated from farmed tilapia (Oreochromis sp.) and elevation of Francisella philomiragia subsp. noatunensis to species rank as Francisella noatunensis comb. nov., sp. nov. Int. J. Syst. Evol. Microbiol. 2009. Available online: http://ijs.sgmjournals.org/content/ijs.0.002139-0v1/.abstract (accessed on 1 November 2020). [CrossRef] [PubMed]
- Ramirez-Paredes, J.G.; Larsson, P.; Thompson, K.D.; Penman, D.J.; Busse, H.-J.; Öhrman, C.; Sjödin, A.; Soto, E.; Richards, R.H.; Adams, A.; et al. Reclassification of Francisella noatunensis subsp. orientalis Ottem et al. 2009 as Francisella orientalis sp. nov., Francisella noatunensis subsp. chilensis subsp. nov. and emended description of Francisella noatunensis. Int. J. Syst. Evol. Microbiol. 2020, 70, 2034–2048. [Google Scholar] [CrossRef] [PubMed]
- Colquhoun, D.J.; Duodu, S. Francisella infections in farmed and wild aquatic organisms. Veter-Res. 2011, 42, 47. [Google Scholar] [CrossRef] [Green Version]
- Larson, C.L.; Wicht, W.; Jellison, W.L. A new organism resembling P. tularensis isolated from water. Public Health Rep. 1955, 70, 253–258. Available online: http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2024510/ (accessed on 1 November 2020). [CrossRef]
- Aravena-Román, M.; Merritt, A.J.; Inglis, T.J. First case of Francisella bacteraemia in Western Australia. New Microbes New Infect. 2015, 8, 75–77. [Google Scholar] [CrossRef] [Green Version]
- Olsufiev, N.G.; Emelyanova, O.S.; Dunayeva, T.N. Comparative study of strains of B. tularense in the old and new world and their taxonomy. J. Hyg. Epidemiol. Microbiol. Immunol. 1959, 3, 138–149. [Google Scholar]
- Hollis, D.G.; Weaver, R.; Steigerwalt, A.G.; Wenger, J.D.; Moss, C.W.; Brenner, D.J. Francisella philomiragia comb. nov. (formerly Yersinia philomiragia) and Francisella tularensis biogroup novicida (formerly Francisella novicida) associated with human disease. J. Clin. Microbiol. 1989, 27, 1601–1608. [Google Scholar] [CrossRef] [Green Version]
- Huber, B.; Escudero, R.; Busse, H.-J.; Seibold, E.; Scholz, H.C.; Anda, P.; Kämpfer, P.; Splettstoesser, W.D. Description of Francisella hispaniensis sp. nov., isolated from human blood, reclassification of Francisella novicida (Larson et al. 1955) Olsufiev et al. 1959 as Francisella tularensis subsp. novicida comb. nov. and emended description of the genus Francisella. Int. J. Syst. Evol. Microbiol. 2010, 60, 1887–1896. [Google Scholar] [CrossRef]
- Sjödin, A.; Öhrman, C.; Bäckman, S.; Lärkeryd, A.; Granberg, M.; Lundmark, E.; Karlsson, E.; Nilsson, E.; Vallesi, A.; Tellgren-Roth, C.; et al. Complete Genome Sequence of Francisella endociliophora Strain FSC1006, Isolated from a Laboratory Culture of the Marine Ciliate Euplotes raikovi. Genome Announc. 2014, 2, e01227-14. [Google Scholar] [CrossRef] [Green Version]
- Vallesi, A.; Sjödin, A.; Petrelli, D.; Luporini, P.; Taddei, A.R.; Thelaus, J.; Öhrman, C.; Nilsson, E.; Di Giuseppe, G.; Gutiérrez, G.; et al. A New Species of the γ-Proteobacterium Francisella, F. adeliensis sp. Nov., Endocytobiont in an Antarctic Marine Ciliate and Potential Evolutionary Forerunner of Pathogenic Species. Microb. Ecol. 2019, 77, 587–596. [Google Scholar] [CrossRef]
- Larson, M.A.; Nalbantoglu, U.; Sayood, K.; Zentz, E.B.; Cer, R.Z.; Iwen, P.C.; Francesconi, S.C.; Bishop-Lilly, K.A.; Mokashi, V.P.; Sjöstedt, A.; et al. Reclassification of Wolbachia persica as Francisella persica comb. nov. and emended description of the family Francisellaceae. Int. J. Syst. Evol. Microbiol. 2016, 66, 1200–1205. [Google Scholar] [CrossRef] [PubMed]
- Challacombe, J.F.; Petersen, J.M.; Gallegos-Graves, L.V.; Hodge, D.; Pillai, S.; Kuske, C.R. Whole-Genome Relationships among Francisella Bacteria of Diverse Origins Define New Species and Provide Specific Regions for Detection. Appl. Environ. Microbiol. 2016, 83, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rydzewski, K.; Schulz, T.; Brzuszkiewicz, E.; Holland, G.; Lück, C.; Fleischer, J.; Grunow, R.; Heuner, K. Genome sequence and phenotypic analysis of a first German Francisella sp. isolate (W12-1067) not belonging to the species Francisella tularensis. BMC Microbiol. 2014, 14, 169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Respicio-Kingry, L.; Byrd, L.; Allison, A.; Brett, M.; Scott-Waldron, C.; Galliher, K.; Hannah, P.; Mead, P.; Petersen, J.M. Cutaneous Infection Caused by a Novel Francisella sp. J. Clin. Microbiol. 2013, 51, 3456–3460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uneklint, I.; Öhrman, C.; Karlsson, L.; Byström, M.; Hägglund, E.; Forsman, M.; Sjödin, A. Complete Genome Sequence of Francisella hali-oticida Type Strain DSM 23729 (FSC1005). Microbiol. Resour. Announc. 2020, 9, e01418-17. Available online: https://mra.asm.org/content/9/37/e00541-20 (accessed on 1 November 2020). [CrossRef] [PubMed]
- Öhrman, C.; Uneklint, I.; Karlsson, L.; Svensson, D.; Forsman, M.; Sjödin, A. Complete Genome Sequences of Allofrancisella inopinata SYSU YG23 and Allofrancisella frigidaquae SYSU 10HL1970, Isolated from Water from Cooling Systems in China. Microbiol. Resour. Announc. 2020, 9. Available online: https://mra.asm.org/content/9/48/e00554-20 (accessed on 1 November 2020).
- Zhou, H.; Yang, Q.; Shen, L.; Yao, Y.; Xu, J.; Ye, J.; Wu, X.; Yu, Y.; Li, Z.; Zhou, J.; et al. Seawater-Associated Highly Pathogenic Francisella hispaniensis Infections Causing Multiple Organ Failure. Emerg. Infect. Dis. 2020, 26, 2424–2428. [Google Scholar] [CrossRef]
- Whitehouse, C.; Kesterson, K.; Duncan, D.; Eshoo, M.; Wolcott, M. Identification and characterization of Francisella species from natural warm springs in Utah, USA. Lett. Appl. Microbiol. 2012, 54, 313–324. [Google Scholar] [CrossRef]
- Brett, M.; Respicio-Kingry, L.B.; Yendell, S.; Ratard, R.; Hand, J.; Balsamo, G.; Scott-Waldron, C.; O’Neal, C.; Kidwell, D.; Yockey, B.; et al. Outbreak of Francisella novicida Bacteremia Among Inmates at a Louisiana Correctional Facility. Clin. Infect. Dis. 2014, 59, 826–833. [Google Scholar] [CrossRef] [Green Version]
- Qu, P.-H.; Chen, S.; Scholz, H.C.; Busse, H.-J.; Gu, Q.; Kämpfer, P.; Foster, J.T.; Glaeser, S.P.; Chen, C.; Yang, Z.-C. Francisella guangzhouensis sp. nov., isolated from air-conditioning systems. Int. J. Syst. Evol. Microbiol. 2013, 63, 3628–3635. [Google Scholar] [CrossRef]
- Person, T.; Currie, C. Biosurveillance: DHS Should Not Pursue BioWatch Upgrades or Enhancements Until System Capabilities Are Established; United States Government Accountability Office: Washington, DC, USA, 2015. Available online: https://www.gao.gov/assets/680/673309.pdf (accessed on 1 November 2020).
- Friend, M. Tularemia. Acta Vet. Scand. 2016. Available online: http://wildpro.twycrosszoo.org/000ADOBES/Lagomorphs/B527Circ1297Tularemia.pdf (accessed on 1 November 2020).
- Parker, R.R.; Steinhaus, E.; Kohls, G.M.; Jellison, W.L. Contamination of natural waters and mud with Pasteurella tularensis and tularemia in beavers and muskrats in the northwestern United States. Bull. Natl. Institutes Heal. (U.S.) 1951, 193, 1–161. [Google Scholar]
- Karadenizli, A.; Forsman, M.; Şimşek, H.; Taner, M.; Öhrman, C.; Myrtennäs, K.; Lärkeryd, A.; Johansson, A.; Özdemir, L.; Sjödin, A. Genomic analyses of Francisella tularensis strains confirm disease transmission from drinking water sources, Turkey, 2008, 2009 and 2012. Eurosurveillance 2015, 20. Available online: http://www.ncbi.nlm.nih.gov/pubmed/26062561 (accessed on 1 November 2020). [CrossRef] [PubMed]
- Kuske, C.R.; Barns, S.M.; Grow, C.C.; Merrill, L.; Dunbar, J. Environmental Survey for Four Pathogenic Bacteria and Closely Related Species Using Phylogenetic and Functional Genes*. J. Forensic. Sci. 2006, 51, 548–558. [Google Scholar] [CrossRef] [PubMed]
- Barns, S.M.; Grow, C.C.; Okinaka, R.T.; Keim, P.; Kuske, C.R. Detection of diverse new Francisella-like bacteria in environmental sam-ples. Appl. Environ. Microbiol. 2005, 71, 5494–5500. Available online: http://aem.asm.org/content/71/9/5494.abstract (accessed on 1 November 2020). [CrossRef] [PubMed] [Green Version]
- Gerhart, J.G.; Auguste Dutcher, H.; Brenner, A.E.; Moses, A.S.; Grubhoffer, L.; Raghavan, R. Multiple Acquisitions of Patho-gen-Derived Francisella Endosymbionts in Soft Ticks. Genome Biol. Evol. 2018, 10, 607–615. [Google Scholar] [CrossRef] [Green Version]
- Sjödin, A.; Svensson, K.; Öhrman, C.; Ahlinder, J.; Lindgren, P.; Duodu, S.; Johansson, A.; Colquhoun, D.J.; Larsson, P.; Forsman, M. Genome characterisation of the genus Francisella reveals insight into similar evolutionary paths in pathogens of mammals and fish. BMC Genom. 2012, 13, 268. [Google Scholar] [CrossRef] [Green Version]
- Kumar, R.; Bröms, J.E.; Sjöstedt, A. Exploring the Diversity within the Genus Francisella—An Integrated Pan-Genome and Ge-nome-Mining Approach. Front. Microbiol. 2020, 11. Available online: https://www.frontiersin.org/article/10.3389/fmicb.2020.01928/full (accessed on 1 November 2020). [CrossRef]
- Larsson, P.; Elfsmark, D.; Svensson, K.; Wikström, P.; Forsman, M.; Brettin, T.; Keim, P.; Johansson, A. Molecular Evolutionary Consequences of Niche Restriction in Francisella tularensis, a Facultative Intracellular Pathogen. PLoS Pathog. 2009, 5, e1000472. [Google Scholar] [CrossRef] [Green Version]
- Larsson, P.; Oyston, P.C.F.; Chain, P.; Chu, M.C.; Duffield, M.; Fuxelius, H.-H.; Garcia, E.; Hälltorp, G.; Johansson, D.; Isherwood, K.E.; et al. The complete genome sequence of Francisella tu-larensis, the causative agent of tularemia. Nat. Genet. 2005, 37, 153–159. Available online: http://www.ncbi.nlm.nih.gov/pubmed/15640799 (accessed on 1 November 2020). [CrossRef] [Green Version]
- Sampson, T.R.; Weiss, D.S. Degeneration of a CRISPR/Cas system and its regulatory target during the evolution of a pathogen. RNA Biol. 2013, 10, 1618–1622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schunder, E.; Rydzewski, K.; Grunow, R.; Heuner, K. First indication for a functional CRISPR/Cas system in Francisella tularensis. Int. J. Med. Microbiol. 2013, 303, 51–60. [Google Scholar] [CrossRef] [Green Version]
- Kozarewa, I.; Turner, D.J. Amplification-Free Library Preparation for Paired-End Illumina Sequencing. Comput. Biol. 2011, 733, 257–266. [Google Scholar]
- Jackman, S.D.; Vandervalk, B.P.; Mohamadi, H.; Chu, J.; Yeo, S.; Hammond, S.A.; Jahesh, G.; Khan, H.; Coombe, L.; Warren, R.L.; et al. ABySS 2.0: Resource-efficient assembly of large genomes using a Bloom filter. Genome Res. 2017, 27, 768–777. [Google Scholar] [CrossRef] [Green Version]
- Walker, B.J.; Abeel, T.; Shea, T.; Priest, M.; Abouelliel, A.; Sakthikumar, S.; Cuomo, C.A.; Zeng, Q.; Wortman, J.; Young, S.K.; et al. Pilon: An Integrated Tool for Comprehensive Microbial Variant Detection and Genome Assembly Improvement. PLoS ONE 2014, 9, e112963. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. Available online: http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3342519/ (accessed on 1 November 2020). [CrossRef] [Green Version]
- Parks, D.H.; Chuvochina, M.; Chaumeil, P.-A.; Rinke, C.; Mussig, A.J.; Hugenholtz, P. A complete domain-to-species taxonomy for Bacteria and Archaea. Nat. Biotechnol. 2020, 38, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Darling, A.E.; Mau, B.; Perna, N.T. progressiveMauve: Multiple Genome Alignment with Gene Gain, Loss and Rearrangement. PLoS ONE 2010, 5, e11147. [Google Scholar] [CrossRef] [Green Version]
- Page, A.J.; Taylor, B.; Delaney, A.J.; Soares, J.; Seemann, T.; Keane, J.A.; Harris, S.R. SNP-sites: Rapid efficient extraction of SNPs from multi-FASTA alignments. Microb. Genom. 2016, 2, e000056. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Pritchard, L.; Glover, R.H.; Humphris, S.; Elphinstone, J.G.; Toth, I.K. Genomics and taxonomy in diagnostics for food security: Soft-rotting enterobacterial plant pathogens. Anal. Methods R. Soc. Chem. 2016, 8, 12–24. Available online: www.rsc.org/methods (accessed on 1 November 2020). [CrossRef]
- Hyatt, D.; Chen, G.-L.; Locascio, P.F.; Land, M.L.; Larimer, F.W.; Hauser, L.J. Prodigal: Prokaryotic gene recognition and translation initiation site identification. BMC Bioinform. 2010, 11, 119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, L.; Niu, B.; Zhu, Z.; Wu, S.; Li, W. CD-HIT: Accelerated for clustering the next-generation sequencing data. Bioinformatics 2012, 28, 3150–3152. [Google Scholar] [CrossRef] [PubMed]
- Camacho, C.E.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.S.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasko, D.; Myers, G.S.A.; Ravel, J. Visualization of comparative genomic analyses by BLAST score ratio. BMC Bioinform. 2005, 6, 2. [Google Scholar] [CrossRef] [Green Version]
- Letunic, I.; Bork, P. Interactive tree of life (iTOL) v3: An online tool for the display and annotation of phylogenetic and other trees. Nucleic Acids Res. 2016, 44, W242–W245. [Google Scholar] [CrossRef]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S. Primer3—New capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, J.L.; Chatwell, N.; Christensen, D.; Diaper, H.; Minogue, T.D.; Parsons, T.M.; Walker, B.; Weller, S.A. Development of real-time PCR assays for the specific detection of Francisella tularensis ssp. tularensis, holarctica and mediaasiatica. Mol. Cell. Probes 2010, 24, 72–76. [Google Scholar] [CrossRef]
- Larson, M.A.; Sayood, K.; Bartling, A.M.; Meyer, J.R.; Starr, C.; Baldwin, J.; Dempsey, M.P. Differentiation of Francisella tularensis Subspecies and Subtypes. J. Clin. Microbiol 2020, 58, 1–16. Available online: https://jcm.asm.org/content/58/4/e01495-19 (accessed on 1 November 2020). [CrossRef] [Green Version]
- Birdsell, D.N.; Vogler, A.J.; Buchhagen, J.; Clare, A.; Kaufman, E.; Naumann, A.; Driebe, E.; Wagner, D.M.; Keim, P.S. TaqMan Real-Time PCR Assays for Sin-gle-Nucleotide Polymorphisms Which Identify Francisella tularensis and Its Subspecies and Subpopulations. PLoS ONE 2014, 9, e107964. Available online: http://www.ncbi.nlm.nih.gov/pubmed/25238067 (accessed on 24 September 2014). [CrossRef]
- Liu, C.M.; Aziz, M.; Kachur, S.; Hsueh, P.-R.; Huang, Y.-T.; Keim, P.; Price, L.B. BactQuant: An enhanced broad-coverage bacterial quantitative real-time PCR assay. BMC Microbiol. 2012, 12, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johansson, A.; Celli, J.; Conlan, W.; Elkins, K.L.; Forsman, M.; Keim, P.S.; Larsson, P.; Manoil, C.; Nano, F.E.; Petersen, J.M.; et al. Objections to the transfer of Francisella novicida to the subspecies rank of Francisella tularensis. Int. J. Syst. Evol. Microbiol. 2010, 60, 1717–1718. [Google Scholar] [CrossRef] [PubMed]
- Busse, H.-J.; Scholz, H.C.; Escudero, R.; Anda, P.; Kämpfer, P.; Splettstoesser, W.D.; Huber, B.; Seibold, E. Objections to the transfer of Francisella novicida to the subspecies rank of Francisella tularensis—Response to Johansson et al. Int. J. Syst. Evol. Microbiol. 2010, 60, 1718–1720. [Google Scholar] [CrossRef]
- Petersen, J.M.M.; Carlson, J.; Yockey, B.; Pillai, S.; Kuske, C.; Garbalena, G.; Pottumarthy, S.; Chalcraft, L. Direct isolation of Francisella spp. from environmental samples. Lett. Appl. Microbiol. 2009, 48, 663–667. Available online: http://www.ingentaconnect.com/content/bsc/lappm/2009/00000048/00000006/art00003 (accessed on 1 November 2020). [PubMed]
- Birdsell, D.N.; Johansson, A.; Öhrman, C.; Kaufman, E.; Molins, C.; Pearson, T. Francisella tularensis subsp. tularensis Group A.I, United States. Emerg. Infect. Dis. 2014, 20, 861–865. Available online: http://wwwnc.cdc.gov/eid/article/20/5/13-1559_article.htm (accessed on 1 November 2020). [CrossRef] [Green Version]
- Svensson, K.; Granberg, M.; Karlsson, L.; Neubauerova, V.; Forsman, M.; Johansson, A. A real-time PCR array for hierarchical identification of Francisella isolates. PLoS ONE 2009, 4, e8360. Available online: http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=2793073&tool=pmcentrez&rendertype=abstract (accessed on 1 November 2020). [CrossRef]
- Vogler, A.J.; Birdsell, D.; Price, L.B.; Bowers, J.R.; Beckstrom-Sternberg, S.M.; Auerbach, R.K.; Beckstrom-Sternberg, J.S.; Johansson, A.; Clare, A.; Buchhagen, J.L.; et al. Phylogeography of Francisella tu-larensis: Global expansion of a highly fit clone. J. Bacteriol. 2009, 191, 2474–2484. Available online: http://jb.asm.org/content/191/8/2474.abstract (accessed on 1 November 2020). [CrossRef] [Green Version]
- Birdsell, D.N.; Yaglom, H.; Rodriguez, E.; Engelthaler, D.M.; Maurer, M.; Gaither, M.; Vinocur, J.; Weiss, J.; Terriquez, J.; Komatsu, K.; et al. Phylogenetic Analysis of Francisella tularensis Group A.II Isolates from 5 Patients with Tularemia, Arizona, USA, 2015–2017. Emerg. Infect. Dis. 2019, 25, 944–946. [Google Scholar] [CrossRef]
- Mokrievich, A.N.; Kislichkina, A.A.; Kudryavtseva, T.Y.; Mironova, R.I.; Vakhrameeva, G.M.; Shishkova, N.A.; Timofeev, V.S.; Bogun, A.G.; Pavlov, V.M.; Dyatlov, I.A.; et al. Draft Genome Sequences of Three Francisella tularensis subsp. mediasiatica Strains Isolated in the Altai Territory, Russian Federation. Microbiol Resour Announc [Internet]. American Society for Microbiology; 2020 [cited 2020 Dec 2];9. Available online: https://doi.org/10.1371/journal.pone.0183714 (accessed on 1 November 2020).
- Lärkeryd, A.; Myrtennäs, K.; Karlsson, E.; Dwibedi, C.K.; Forsman, M.; Larsson, P.; Johansson, A.; Sjödin, A. CanSNPer: A hierarchical genotype classifier of clonal pathogens. Bioinformatics. 2014, 30, 1762–1764. Available online: http://cancerres.aacrjournals.org/cgi/doi/10.1158/0008-5472.CAN-15-3332 (accessed on 1 November 2020).
- Chanturia, G.; Birdsell, D.N.; Kekelidze, M.; Zhgenti, E.; Babuadze, G.; Tsertsvadze, N.; Tsanava, S.; Imnadze, P.; Beckstrom-Sternberg, S.M.; Beckstrom-Sternberg, J.; et al. Phylogeography of Francisella tularensis subspecies holarctica from the country of Georgia. BMC Microbiol. 2011, 11, 139. [Google Scholar] [CrossRef] [Green Version]
- Karlsson, E.; Svensson, K.; Lindgren, P.; Byström, M.; Sjödin, A.; Forsman, M.; Johansson, A. The phylogeographic pattern ofFrancisella tularensisin Sweden indicates a Scandinavian origin of Eurosiberian tularaemia. Environ. Microbiol. 2012, 15, 634–645. [Google Scholar] [CrossRef] [PubMed]
- Johansson, A.; Lärkeryd, A.; Widerström, M.; Mörtberg, S.; Myrtännäs, K.; Ohrman, C.; Birdsell, D.; Keim, P.; Wagner, D.M.; Forsman, M.; et al. A respiratory tularemia outbreak caused by diverse clones of Francisella tularensis. Clin. Infect. Dis. 2014, 59, 1546–1553. Available online: http://cid.oxfordjournals.org/content/early/2014/08/05/cid.ciu621.abstract (accessed on 1 November 2020). [CrossRef] [PubMed] [Green Version]
- Gyuranecz, M.; Szeredi, L.; Makrai, L.; Fodor, L.; Meszaros, A.R.; Szépe, B.; Füleki, M.; Erdélyi, K. Tularemia of European Brown Hare (Lepus euro-paeus). Vet. Pathol. Online 2010, 47, 958–963. Available online: http://vet.sagepub.com/content/47/5/958.abstract (accessed on 1 November 2020). [CrossRef] [PubMed]
- Svensson, K.; Larsson, P.; Johansson, D.; Byström, M.; Forsman, M.; Johansson, A. Evolution of Subspecies of Francisella tularensis. J. Bacteriol. 2005, 187, 3903–3908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerhart, J.G.; Moses, A.S.; Raghavan, R. A Francisella-like endosymbiont in the Gulf Coast tick evolved from a mammalian patho-gen. Sci. Rep. 2016, 6, 33670. Available online: http://www.ncbi.nlm.nih.gov/pubmed/27645766 (accessed on 1 November 2020). [CrossRef] [PubMed]
- Liu, J.N.; Yu, Z.J.; Liu, L.M.; Li, N.X.; Wang, R.R.; Zhang, C.M.; Liu, J.Z. Identification, Distribution and Population Dynamics of Francisella-like Endosymbiont in Haemaphysalis doenitzi (Acari: Ixodidae). Sci. Rep. 2016, 6, 35178. Available online: http://www.ncbi.nlm.nih.gov/pubmed/27731377%5Cnhttp://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=PMC5059625 (accessed on 1 November 2020). [CrossRef] [Green Version]
- Angelina, W.-F.; Zając, V.; Sawczyn, A.; Cisak, E.; Sroka, J.; Dutkiewicz, J. Occurrence of Francisella spp. in Dermacentor reticulatus and Ixodes ricinus ticks collected in eastern Poland. Ticks Tick-Borne Dis. 2015, 6, 253–257. [Google Scholar] [CrossRef]
- Ivanov, I.N.; Mitkova, N.; Reye, A.L.; Hübschen, J.M.; Vatcheva-Dobrevska, R.S.; Dobreva, E.G.; Kantardjiev, T.V.; Muller, C.P. Detection of New Francisella-Like Tick Endosymbionts in Hyalomma spp. and Rhipicephalus spp. (Acari: Ixodidae) from Bulgaria. Appl. Environ. Microbiol. 2011, 77, 5562–5565. [Google Scholar] [CrossRef] [Green Version]
- Kingry, L.; Petersen, J.M. Comparative review of Francisella tularensis and Francisella novicida. Front. Cell. Infect. Microbiol. 2014, 4, 35. [Google Scholar] [CrossRef]
- Whipp, M.J.; Davis, J.M.; Lum, G.; De Boer, J.; Zhou, Y.; Bearden, S.W.; Petersen, J.M.; Chu, M.C.; Hogg, G. Characterization of a novicida-like subspecies of Francisella tularensis isolated in Australia. J. Med. Microbiol. 2003, 52, 839–842. [Google Scholar] [CrossRef]
- Berrada, Z.L.; Telford, S.R., III. Survival of Francisella tularensis Type A in brackish-water. Arch. Microbiol. 2011, 193, 223–226. Available online: http://www.springerlink.com/content/f277542lg8860482 (accessed on 1 November 2020). [CrossRef] [PubMed] [Green Version]
- Wenger, J.D.; Hollis, D.G.; Weaver, R.E.; Baker, C.N.; Brown, G.R.; Brenner, D.J.; Broome, C.V. Infection Caused byFrancisella philomiragia(FormerlyYersinia philomiragia). Ann. Intern. Med. 1989, 110, 888–892. [Google Scholar] [CrossRef] [PubMed]
- Li, L.-H.; Luo, H.-M.; Feng, J.-H.; Ming, Y.-Z.; Zheng, M.-L.; Deng, G.-Y.; Chen, C.; Li, W.; Xiao, M.; Qu, P. Francisella salimarina sp. nov., isolated from coastal seawater. Int. J. Syst. Evol. Microbiol. 2020, 70, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Hennebique, A.; Boisset, S.; Maurin, M. Tularemia as a waterborne disease: A review. Emerg. Microbes Infect. 2019, 8, 1027–1042. [Google Scholar] [CrossRef] [PubMed]
- Schrallhammer, M.; Schweikert, M.; Vallesi, A.; Verni, F.; Petroni, G. Detection of a Novel Subspecies of Francisella noatunensis as Endosymbiont of the Ciliate Euplotes raikovi. Microb. Ecol. 2010, 61, 455–464. [Google Scholar] [CrossRef]
- Rohmer, L.; Fong, C.; Abmayr, S.; Wasnick, M.; Freeman, T.J.L.; Radey, M.; Guina, T.; Svensson, K.; Hayden, H.S.; Jacobs, M.A.; et al. Comparison of Francisella tularensis genomes reveals evolutionary events associated with the emergence of human pathogenic strains. Genome Biol. 2007, 8, R102–R116. [Google Scholar] [CrossRef] [Green Version]
- Champion, M.D.; Zeng, Q.; Nix, E.B.; Nano, F.E.; Keim, P.; Kodira, C.D.; Borowsky, M.; Young, S.; Koehrsen, M.; Engels, R.; et al. Comparative Genomic Characterization of Francisella tularensis Strains Belonging to Low and High Virulence Subspecies. PLoS Pathog. 2009, 5, e1000459. [Google Scholar] [CrossRef]
- Skaldin, M.; Tuittila, M.; Zavialov, A.V.; Zavialov, A.V. Secreted Bacterial Adenosine Deaminase Is an Evolutionary Precursor of Adenosine Deaminase Growth Factor. Mol. Biol 2018, 35, 2851–2861. Available online: https://academic.oup.com/mbe/article/35/12/2851/5133304 (accessed on 1 November 2020). [CrossRef]
- Gallagher, L.A.; McKevitt, M.; Ramage, E.R.; Manoil, C. Genetic Dissection of the Francisella novicida Restriction Barrier. J. Bacteriol. 2008, 190, 7830–7837. [Google Scholar] [CrossRef] [Green Version]
- Dorofe’ev, K.A. Classification of the causative agent of tularemia. Symp. Res. Work Inst. Epidemiol. Microbiol. Chita 1947, 170–180. [Google Scholar]
- McCoy, G.G.W.; Chapin, C.W.C. Further observations on a plague-like disease of rodents with a preliminary note on the causative agent, Bacterium tularense. J Infect Dis. 1912, 10, 61–72. Available online: http://www.jstor.org/stable/10.2307/30071893 (accessed on 1 November 2020). [CrossRef]
- Olsufiev, N.G.; Meshcheryakova, I.S. Subspecific Taxonomy of Francisella tularensis McCoy and Chapin 1912. Int. J. Syst. Bacteriol. 1983, 33, 872–874. Available online: http://ijs.sgmjournals.org/content/33/4/872.abstract (accessed on 1 November 2020). [CrossRef] [Green Version]
- Svensson, K.; Sjödin, A.; Byström, M.; Granberg, M.; Brittnacher, M.J.; Rohmer, L.; Jacobs, M.A.; Sims-Day, E.H.; Levy, R.; Zhou, Y.; et al. Genome sequence of Francisella tularensis subspecies holarctica strain FSC200, isolated from a child with tularemia. J. Bacteriol. 2012, 194, 6965–6966. Available online: http://jb.asm.org/content/194/24/6965.abstract (accessed on 1 November 2020). [CrossRef] [PubMed] [Green Version]
- Aikimbaev, M. Taxonomy of Genus Francisella. Rep. Acad. Sci. Kaz. SSR Ser. Biol. 1966, 5, 42–44. [Google Scholar]
- Olsufjev, N.G.; Meshcheryakova, I.S. Infraspecific taxonomy of tularemia agent Francisella tularensis McCoy et Chapin. J. Hyg. Epidemiol. Microbiol. Immunol. 1982, 26, 291–299. Available online: http://www.ncbi.nlm.nih.gov/pubmed/7142690 (accessed on 1 November 2020).
- Jr, E.C.S.; Weiss, E.; Suitor, E. Isolation of a Rickettsialike Microorganism (Wolbachia persica, n. sp.) from Argas persicus (Oken). J. Infect. Dis. 1961, 108, 95–106. Available online: http://jid.oxfordjournals.org/content/108/1/95.full.pdf (accessed on 1 November 2020).
- Forsman, M.; Sandström, G.; Sjöstedt, A. Analysis of 16S ribosomal DNA sequences of Francisella strains and utilization for determination of the phylogeny of the genus and for identification of strains by PCR. Int. J. Syst. Bacteriol. 1994, 44, 38–46. Available online: http://ijs.sgmjournals.org/content/44/1/38.abstract (accessed on 1 November 2020). [CrossRef]
- Noda, H.; Munderloh, U.G.; Kurtti, T.J. Endosymbionts of ticks and their relationship to Wolbachia spp. and tick-borne pathogens of humans and animals. Appl. Environ. Microbiol. 1997, 63, 3926–3932. Available online: http://aem.asm.org/cgi/content/abstract/63/10/3926 (accessed on 1 November 2020). [CrossRef] [Green Version]
- Kugeler, K.J.; Mead, P.S.; McGowan, K.L.; Burnham, J.M.; Hogarty, M.D.; Ruchelli, E.; Pollard, K.; Husband, B.; Conley, C.; Rivera, T.; et al. Isolation and characterization of a novel Francisella sp. from human cerebrospinal fluid and blood. J. Clin. Microbiol. 2008, 46, 2428–2431. [Google Scholar] [CrossRef] [Green Version]
- Dietrich, E.A.; Kingry, L.C.; Kugeler, K.J.; Levy, C.; Yaglom, H.; Young, J.W.; Mead, P.S.; Petersen, J.M. Francisella opportunistica sp. Nov., isolated from human blood and cerebrospinal fluid. Int. J. Syst. Evol. Microbiol. 2020, 70, 1145–1151. [Google Scholar] [CrossRef]
- Jensen, W.I.; Owen, C.R.; Jellison, W.L. Yersinia philomiragia sp. n., a new member of the Pasteurella group of bacteria, naturally pathogenic for the muskrat (Ondatra zibethica). J. Bacteriol. 1969, 100, 1237–1241. Available online: http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=250302&tool=pmcentrez&rendertype=abstract (accessed on 1 November 2020). [CrossRef] [PubMed] [Green Version]
- Ottem, K.F.; Nylund, A.; Karlsbakk, E.; Friis-Møller, A.; Krossøy, B.; Knappskog, D. New species in the genus Francisella (Gammaproteobacteria; Francisellaceae); Francisella piscicida sp. nov. isolated from cod (Gadus morhua). Arch Microbiol. 2007, 188, 547–550. Available online: http://www.springerlink.com/content/q464n38w86k248l4 (accessed on 1 November 2020). [CrossRef] [PubMed]
- Svensson, D.; Öhrman, C.; Bäckman, S.; Karlsson, E.; Nilsson, E.; Byström, M.; Lärkeryd, A.; Myrtennäs, K.; Stenberg, P.; Qu, P.H.; et al. Complete Genome Sequence of Francisella guangzhouensis Strain 08HL01032T, Isolated from Air-Conditioning Systems in China. Genome Announc. 2015, 3, e00024-15. Available online: http://genomea.asm.org/lookup/doi/10.1128/genomeA.00024-15 (accessed on 1 November 2020). [CrossRef] [PubMed] [Green Version]




| Assay | Region in Genome of SCHU S4 a | Sequences of Forward and Reverse Primers b and Probes c | Primer (µM) d | MGB e | Probe (µM) f | Tm °C g |
|---|---|---|---|---|---|---|
| Ft-sp. FTT0376 | 377718–377824 | F CCATATCACTGGCTTTGCTAGACTAGT R TGTTGGCAAAAGCTAAAGAGTCTAAA FAM-AAATTATAAAACCAAACCCAGACCTTCAAACCACA | 0.9 | no | 0.2 | 60 |
| Ft-sp.3Pan | 8967–9049 | F TTTACACCCGTCTCCGTTAGT R CTCTTAAGGATGCAATTTGGGATT FAM-AAGAGGCAAAGCTGGAATTACACTCTCTC | 0.9 | no | 0.2 | 60 |
| Ft-sp. FTS_0772 | 1187646–1187807 | F CAAGGTAAAGAAATTAAGAGTAGTAAAGTTGAATTC R ATTTAATCTAGTATTATCAATTGGGTAAAAAGGTA FAM-GATGTGGCAACAACTGAAAT | 0.9 | yes | 0.2 | 63 |
| Genus | Clade | Species | #Genomes | #genomes | #Unique Regions | Pan-Genome Size (#Genes) | Core-Genome Size (#Genes) |
|---|---|---|---|---|---|---|---|
| Francisella | clade 1 | F. tularensis | 374 | 327 | 6 | 2401 | 1541 |
| F. novicida | 37 | 2 | 2949 | 1499 | |||
| F. sp. TX07-6608 | 1 | 250 | N/A | N/A | |||
| F. hispaniensis | 3 | 121 | 2015 | 1599 | |||
| FLE and F. persica | 3 | 170 | 2293 | 1006 | |||
| F. opportunistica | 3 | 464 | 1751 | 1713 | |||
| clade 2 | F. philomiragia | 113 | 47 | 10 | 2992 | 1611 | |
| F. noatunensis | 23 | 74 | 1944 | 1766 | |||
| F. sp. GA01-2794 | 1 | 180 | N/A | N/A | |||
| F. orientalis | 30 | 36 | 2287 | 2025 | |||
| F. salimarina | 7 | 191 | 2371 | 1786 | |||
| clade 3 | F. endociliophora | 1 | 1 | 1340 | N/A | N/A | |
| clade 4 | F. halioticida | 4 | 1 | 1190 | N/A | N/A | |
| F. uliginis | 1 | 736 | N/A | N/A | |||
| F. sp. LA11-2445 | 1 | 587 | N/A | N/A | |||
| F. sp. SYW 9 | 1 | 1179 | N/A | N/A | |||
| Parafrancisella | F. adeliensis | 3 | 3 | 1874 | 1936 | 1925 | |
| Allofrancisella | A. inopinata | 7 | 1 | 377 | N/A | N/A | |
| A. frigidaquae | 2 | 211 | 1479 | 1356 | |||
| A. guangzhouensis | 4 | 715 | 1570 | 1455 | |||
| Pseudofrancisella | P. aestuarii | 2 | 1 | 375 | N/A | N/A | |
| F. frigiditurris | 1 | 406 | N/A | N/A |
| Region | LVS * Coordinates | SCHU S4 ** Coordinates | Marker Length | Annotation | Notes |
|---|---|---|---|---|---|
| 1 | 1490829–1491161 | 1187496–1187828 | 333 | hsdS, type I restriction modification domain protein | partial locus |
| 2 | 84841–85406 | 378437–377872 | 567 | FTT_0376c, hypothetical membrane protein | partial locus |
| 3 | 1673808–1673858 | 547774–547824 | 51 | FTT_0525, conserved hypothetical protein | |
| 4 | 43283–43397 | 951193–951307 | 114 | add1, adenosine deaminase | partial locus |
| 5 | 43766–43874 | 950716–950824 | 109 | intergenic, region between bioA and add1 | |
| 6 | 135869–136156 | 765604–765891 | 288 | FTT_0742, hypothetical protein | partial locus |
| Species | Subspecies | Genetic Group | #Unique Regions | #Regions Lost | Number of F. tularensis Subspecies Region is Present in |
|---|---|---|---|---|---|
| F. tularensis | tularensis | A | 70 | 1 | 3 |
| A1 | 39 | 0 | 2 | ||
| A2 | 31 | 0 | 2 | ||
| holarctica | B (excluding japonica) | 250 | 0 | 0 | |
| japonica | 5 | 1 | 2 | ||
| B including japonica | 255 | 7 | 2 | ||
| mediasiatica | M | 7 | 3 | 2 |
| #Isolates | Ft-sp.FTT0376 | Ft-sp.3Pan | Ft-sp.FTS_0772 | |
|---|---|---|---|---|
| Francisella tularensis subspecies tularensis | 25 | 25 | 25 | 25 |
| Francisella tularensis subspecies holarctica | 13 | 13 | 13 | 13 |
| Francisella tularensis subspecies mediasiatica | 2 | 2 | 2 | 2 |
| Francisella novicida | 7 | |||
| Francisella hispaniensis | 2 | |||
| Francisella persica | 1 | |||
| Francisella FLE | 1 | |||
| Francisella opportunistica | 1 | |||
| Francisella philomiragia | 23 | 2 | ||
| Francisella halioticida | 1 | |||
| Francisella noatunensis | 1 | |||
| Francisella orientalis | 1 | |||
| Francisella salimarina | 1 | |||
| Francisella endociliophora | 1 | |||
| Parafrancisella adeliensis | 1 | |||
| Allofrancisella inopinata | 1 | |||
| Allofrancisella frigidaquae | 1 | |||
| Allofrancisella guangzhouensis | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Öhrman, C.; Sahl, J.W.; Sjödin, A.; Uneklint, I.; Ballard, R.; Karlsson, L.; McDonough, R.F.; Sundell, D.; Soria, K.; Bäckman, S.; et al. Reorganized Genomic Taxonomy of Francisellaceae Enables Design of Robust Environmental PCR Assays for Detection of Francisella tularensis. Microorganisms 2021, 9, 146. https://doi.org/10.3390/microorganisms9010146
Öhrman C, Sahl JW, Sjödin A, Uneklint I, Ballard R, Karlsson L, McDonough RF, Sundell D, Soria K, Bäckman S, et al. Reorganized Genomic Taxonomy of Francisellaceae Enables Design of Robust Environmental PCR Assays for Detection of Francisella tularensis. Microorganisms. 2021; 9(1):146. https://doi.org/10.3390/microorganisms9010146
Chicago/Turabian StyleÖhrman, Caroline, Jason W. Sahl, Andreas Sjödin, Ingrid Uneklint, Rebecca Ballard, Linda Karlsson, Ryelan F. McDonough, David Sundell, Kathleen Soria, Stina Bäckman, and et al. 2021. "Reorganized Genomic Taxonomy of Francisellaceae Enables Design of Robust Environmental PCR Assays for Detection of Francisella tularensis" Microorganisms 9, no. 1: 146. https://doi.org/10.3390/microorganisms9010146