
Bacillus Cyclic Lipopeptides Iturin and Fengycin Control Rice Blast Caused by Pyricularia oryzae in Potting and Acid Sulfate Soils by Direct Antagonism and Induced Systemic Resistance

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material, Strains, Media and Growth Conditions
2.2. Construction of Bacillus velezensis GA1 Mutants
2.3. Preparation of Cell-Free Supernatants and UPLC–MS for Detection of CLiPs
2.4. Potential of Indigenous Bacillus Strains to Trigger ISR against Rice Blast Caused by P. oryzae in Potting and Acid Sulfate Soil
2.5. Role of Cyclic Lipopeptides in ISR Triggered by B. velezensis against P. oryzae in Potting and Acid Sulfate Soil Conditions
2.6. Root Colonization Assay
2.7. In Vitro Antagonism against P. oryzae Using Cell-Free Supernatants of B. velezensis Strains and GA1 CLiP Mutants
2.8. Influence of Cell-Free Culture Filtrates on P. oryzae Spore Germination and Appressoria Formation
2.9. Direct Effect of Cell-Free Supernatants to Reduce Rice Blast Symptoms Caused by P. oryzae Spores
2.10. Statistical Data Analysis
3. Results
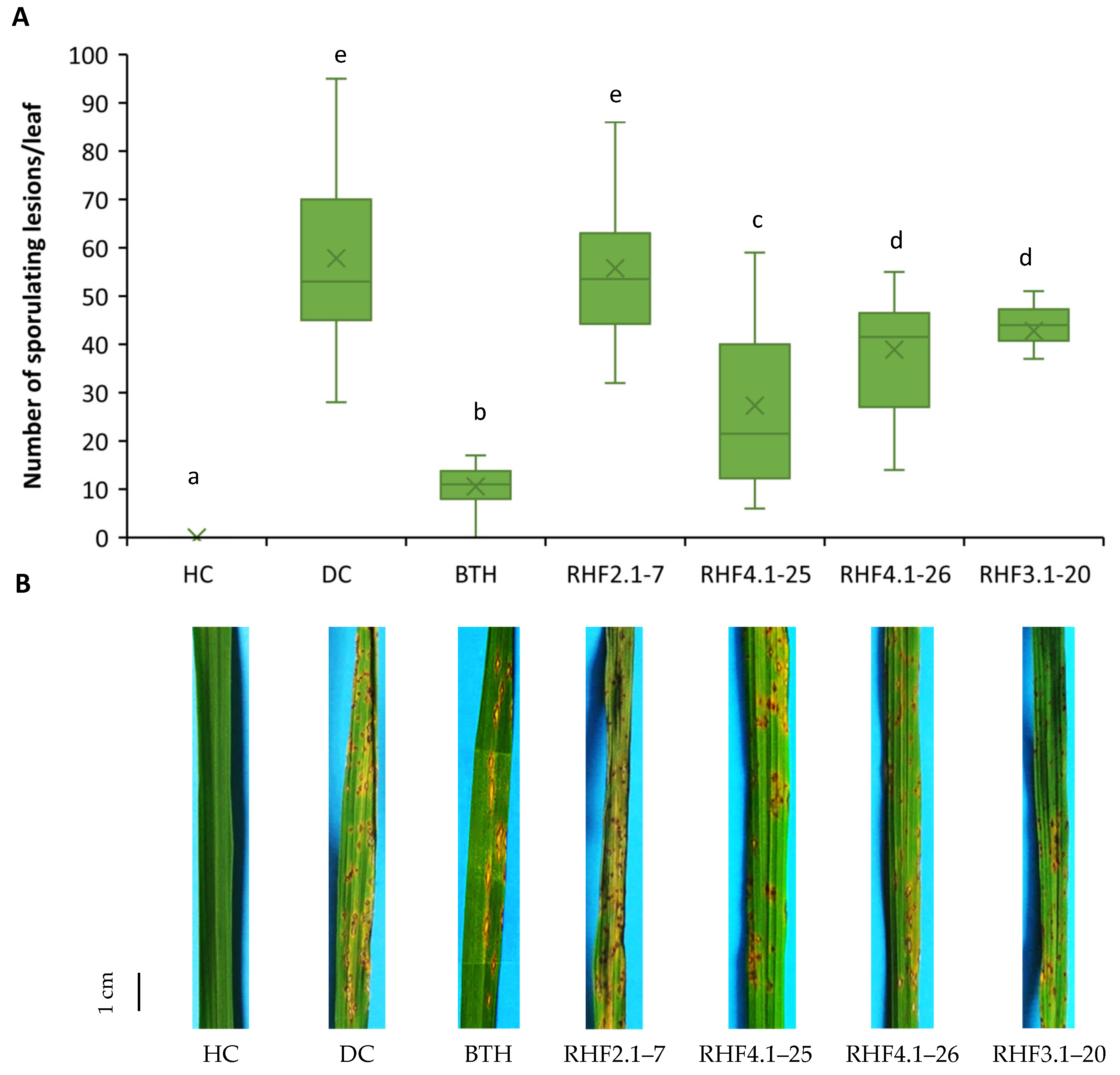
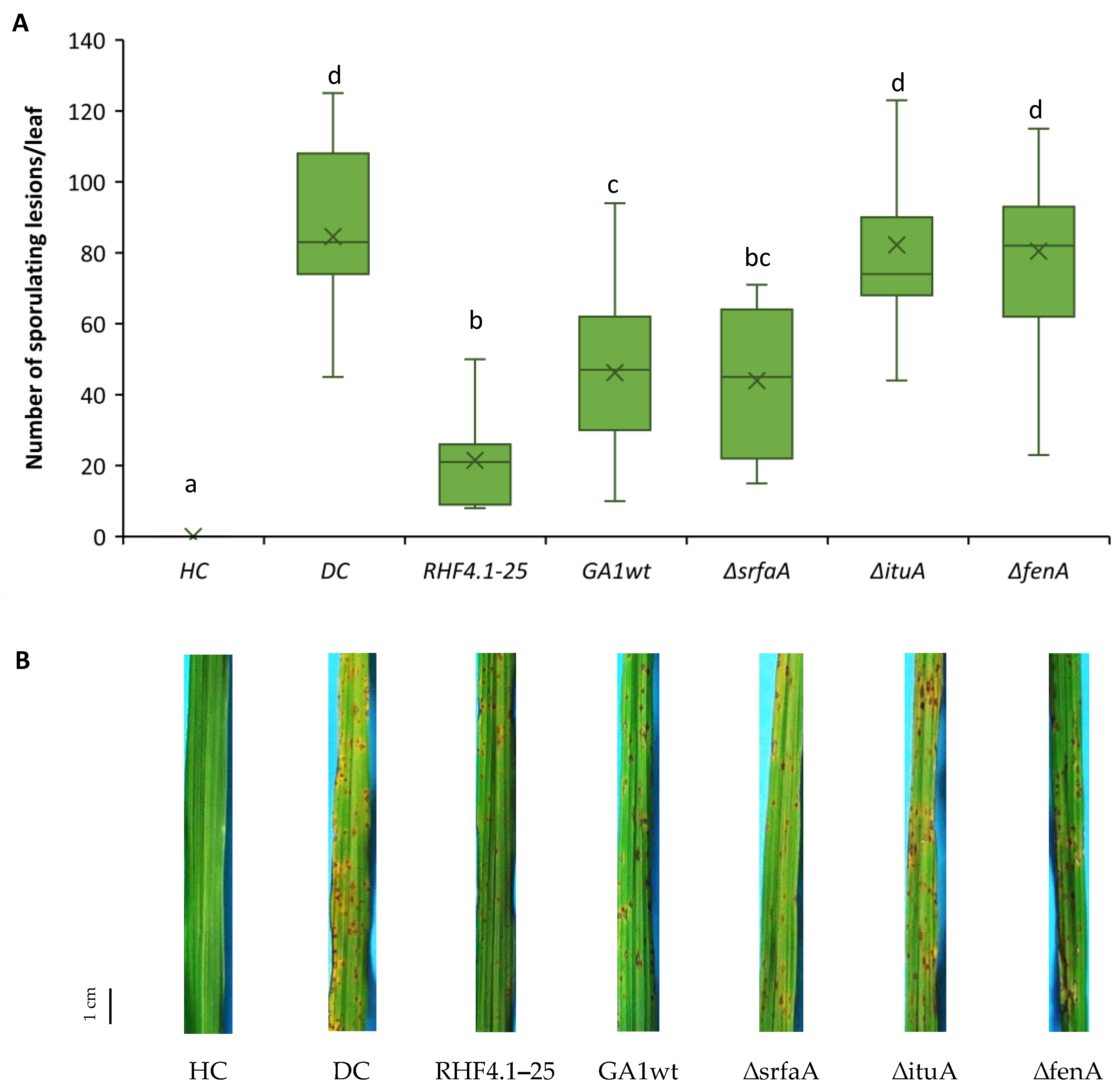
3.1. Potential of Indigenous Bacillus Strains to Control Rice Blast by ISR in Potting Soil and ASS Conditions
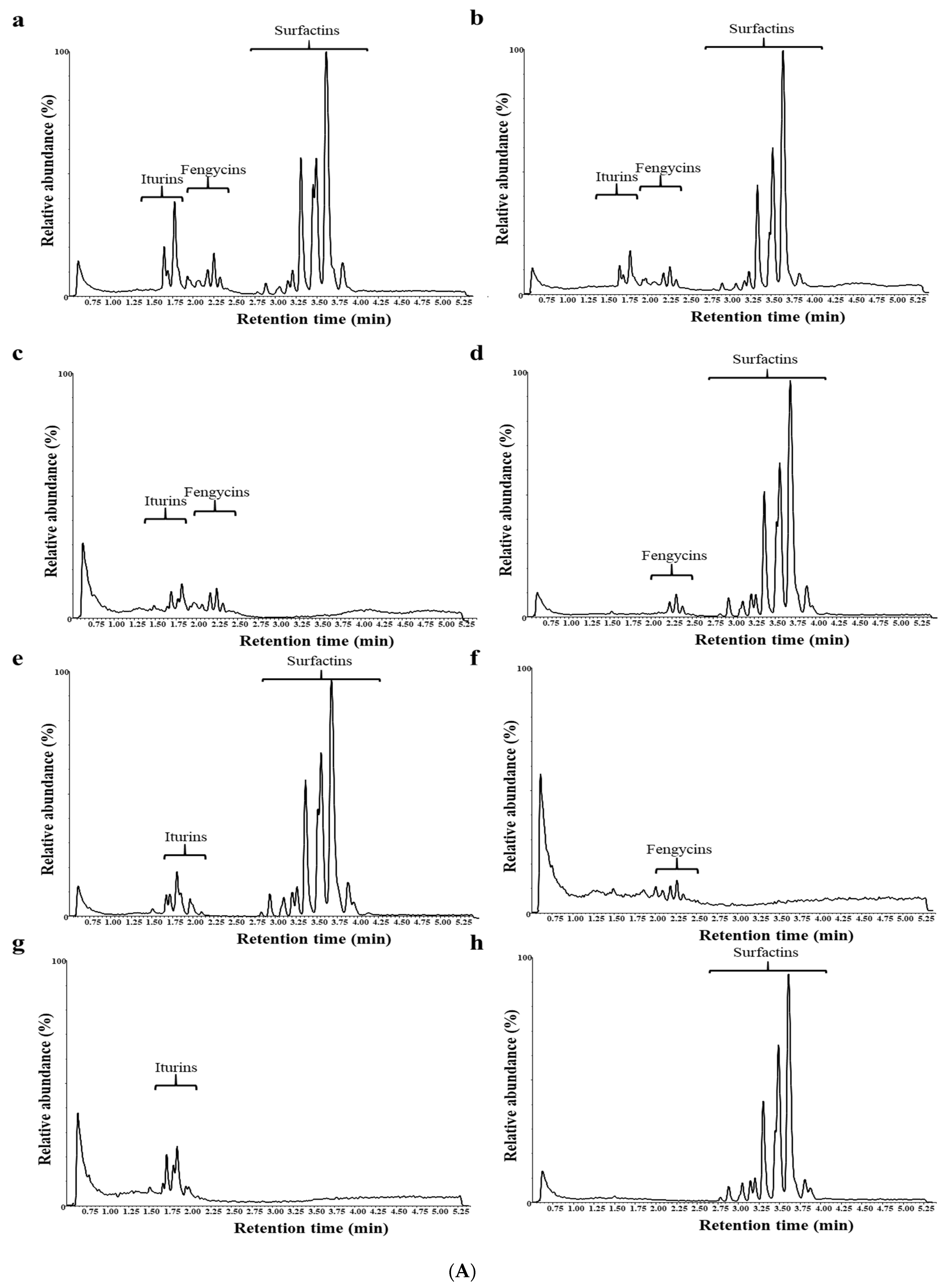
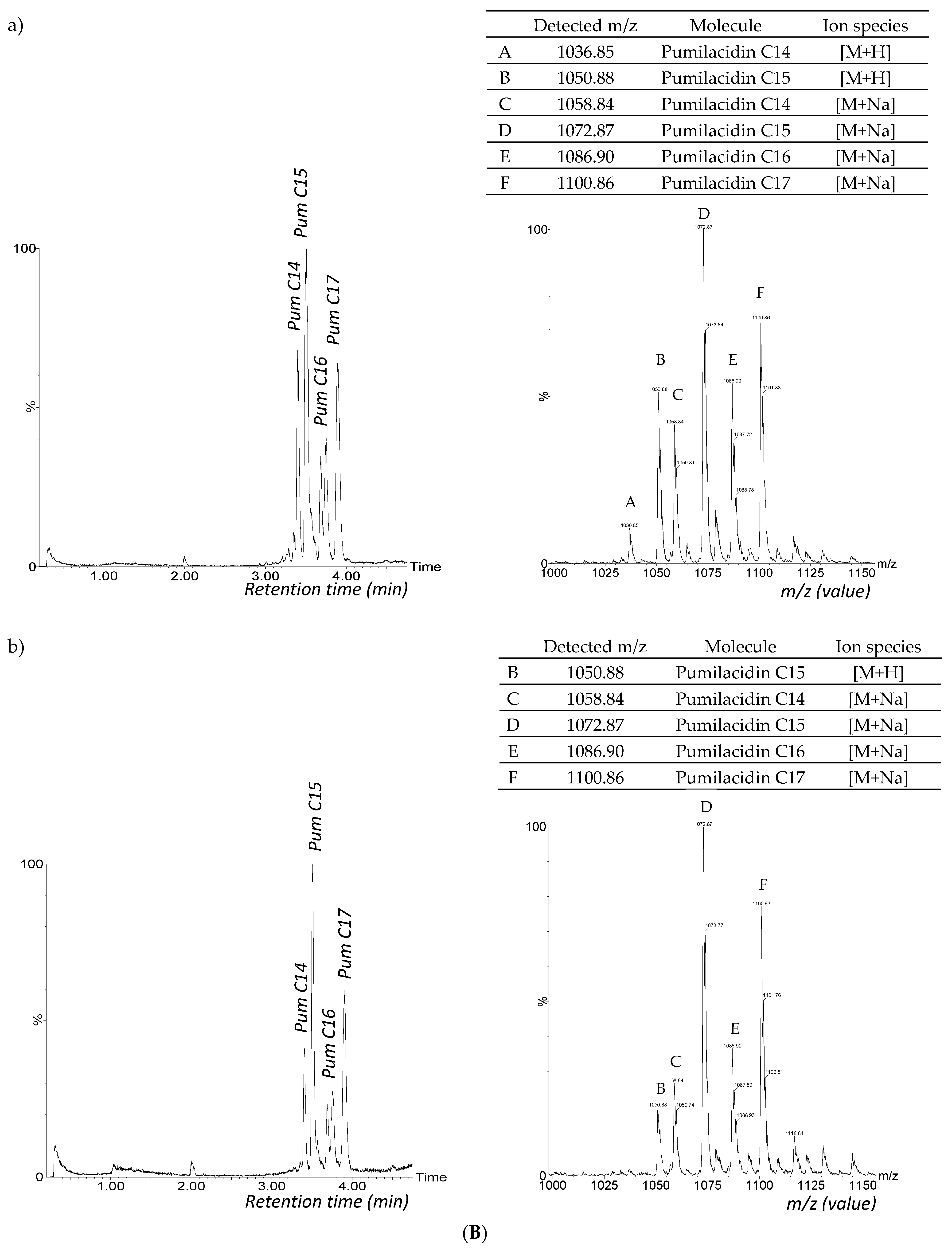
3.2. CLiP Production in B. altitudinis, B. velezensis and Mutants
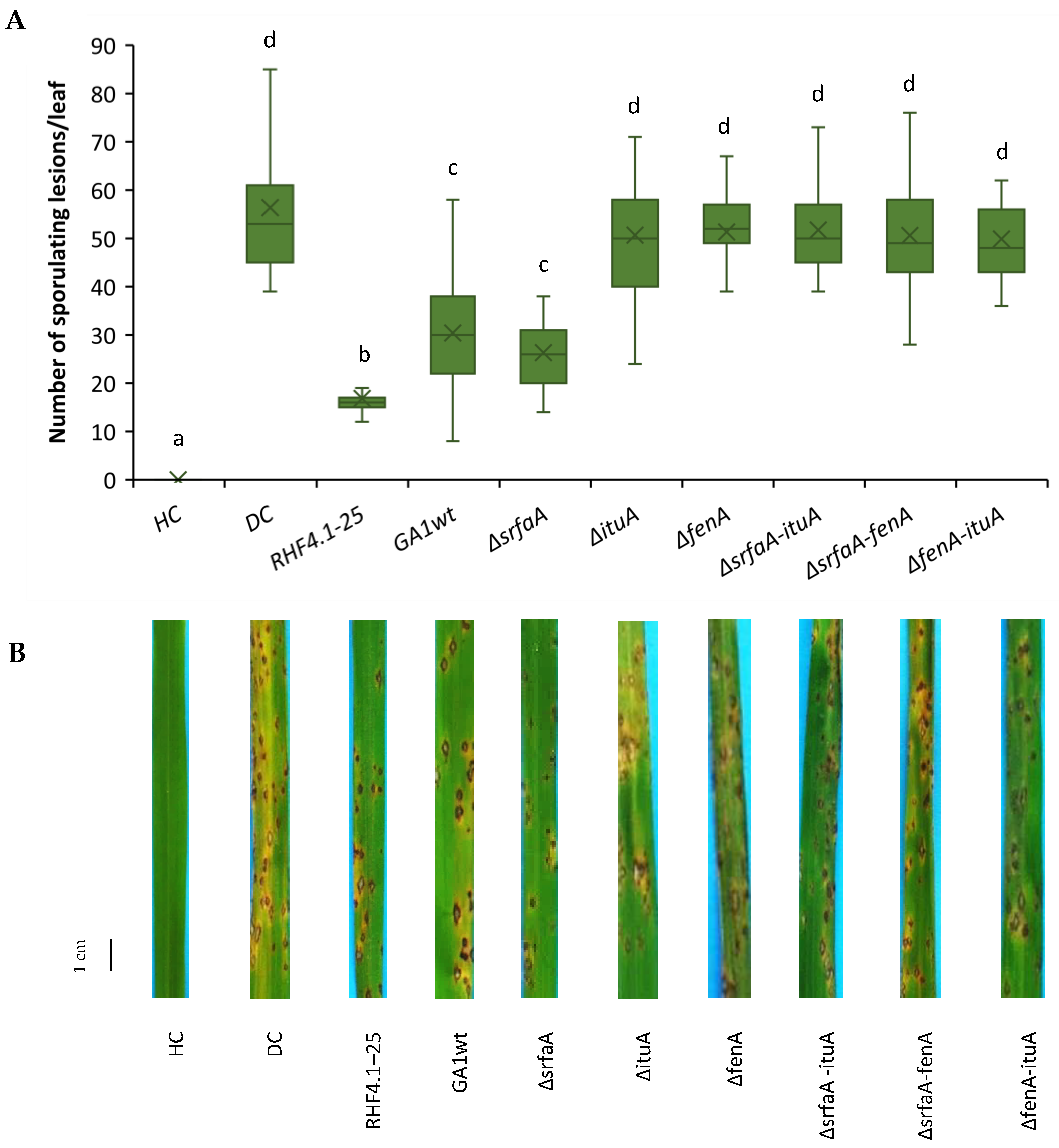
3.3. Role of Cyclic Lipopeptides Produced by B. velezensis in ISR against P. oryzae VT5M1 in Potting Soil
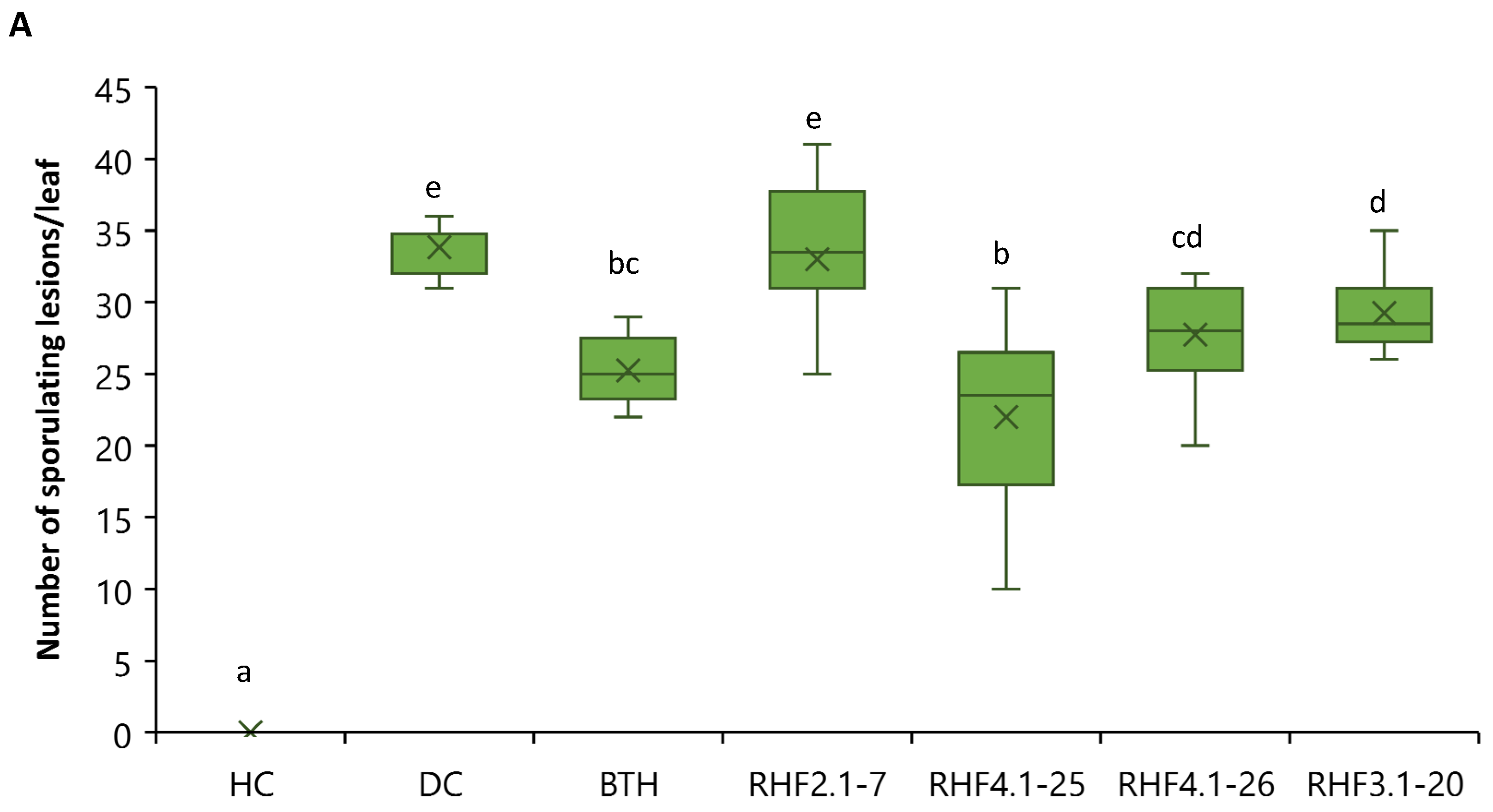
3.4. Role of Cyclic Lipopeptides in ISR of B. velezensis against P. oryzae VT5M1 in Acid Sulfate Soil
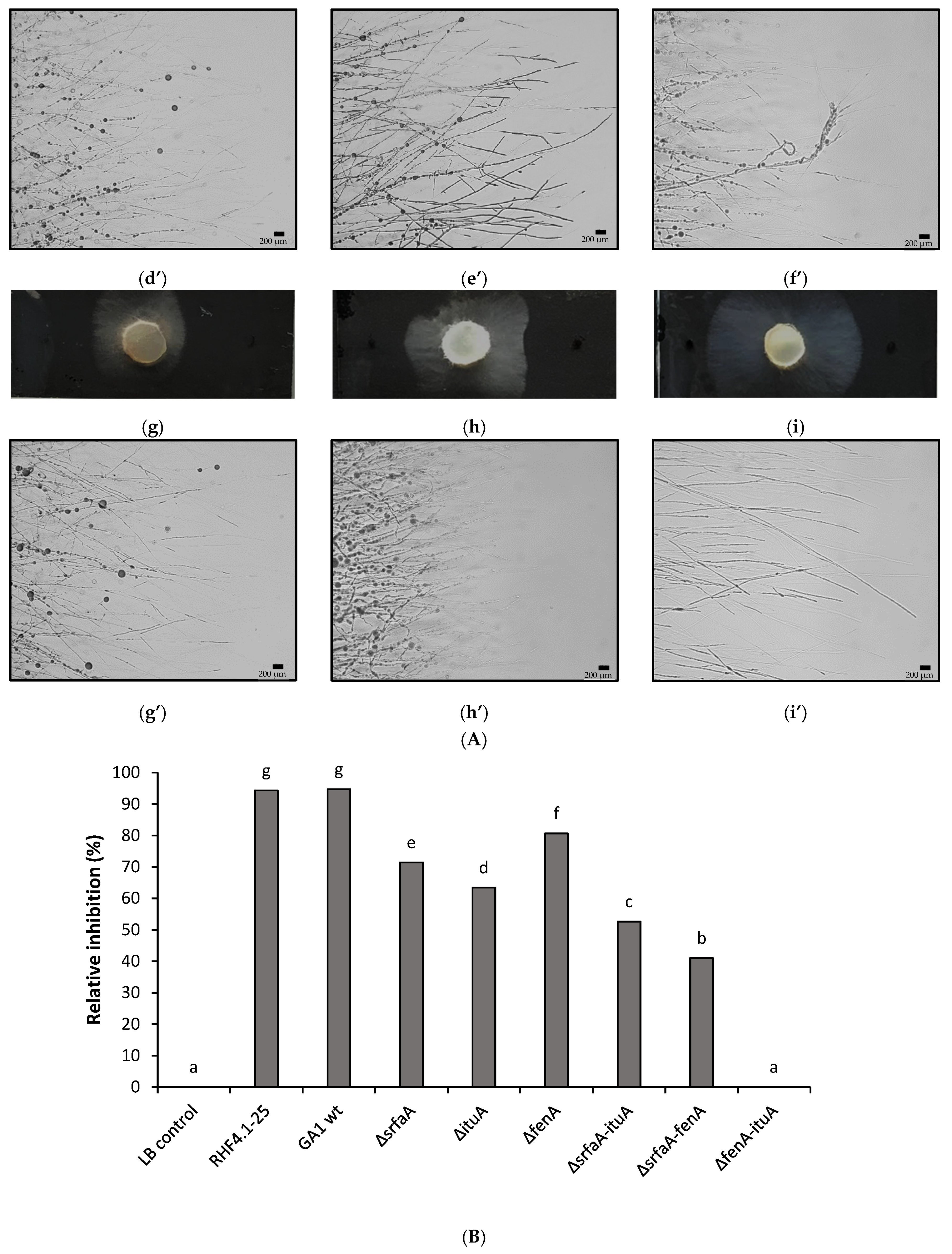
3.5. In Vitro Antagonism of P. oryzae Using Cell-Free Supernatants of B. velezensis Wild Type Strains and CLiP Mutants
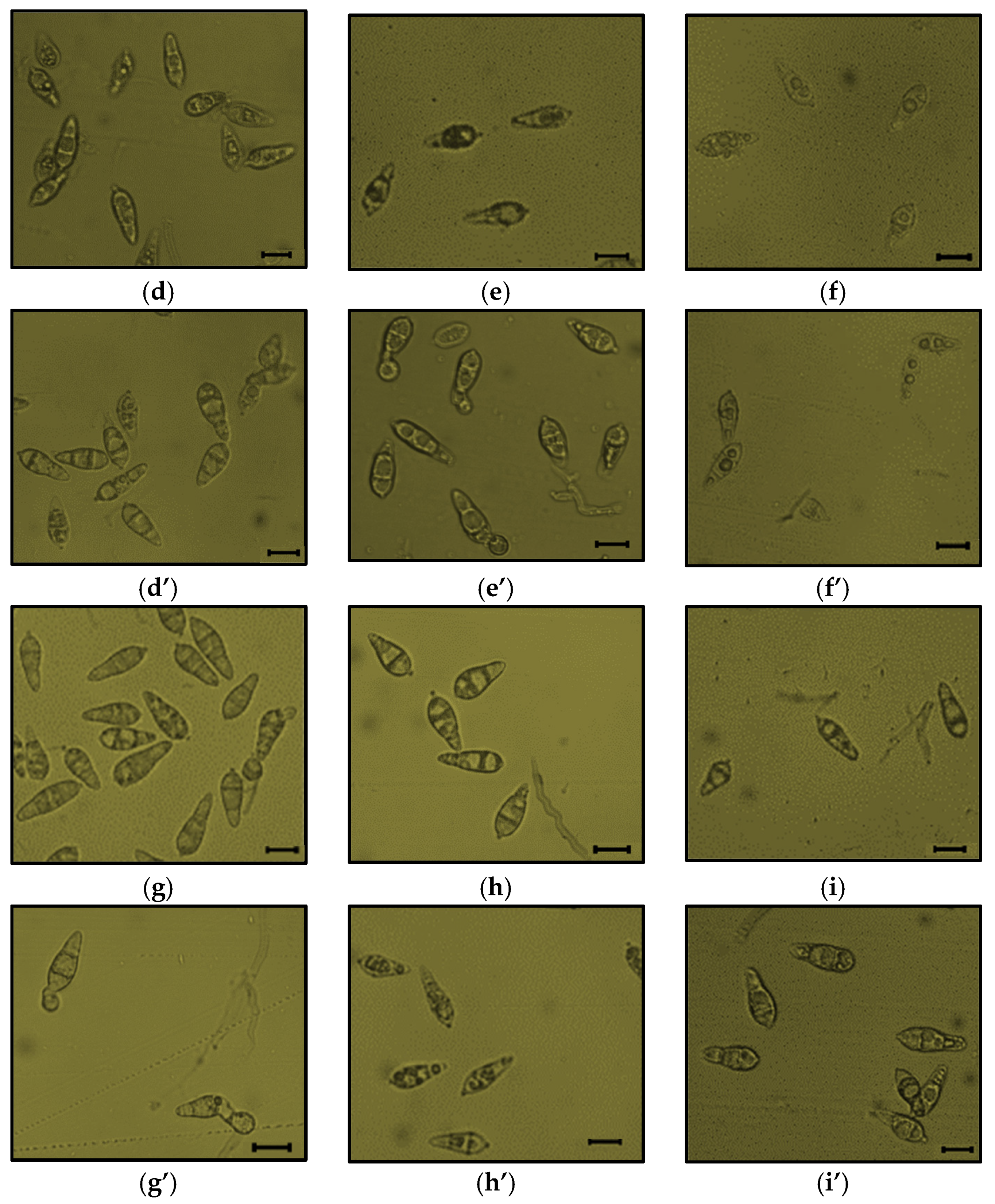
3.6. Influence of Cell-Free Culture Filtrates on P. oryzae Spore Germination and Appressoria Formation
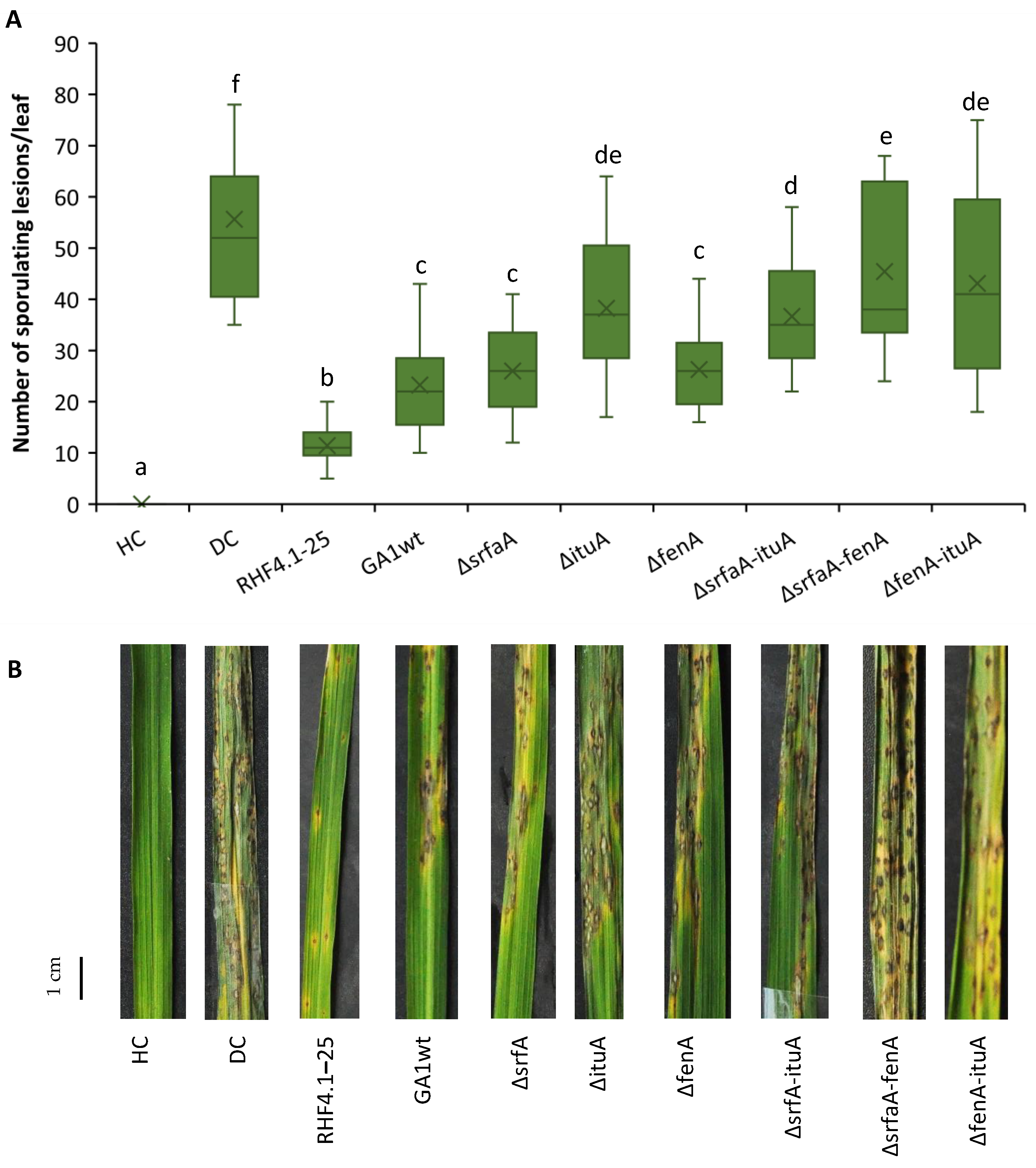
3.7. Direct Effect of Cell-Free Supernatants to Reduce Rice Blast Symptoms Caused by P. oryzae Spores
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Primer Sequence (5′->3′) | Targeted Genes | Reference | |
|---|---|---|---|---|
| Deletion Mutant | ||||
| B. velezensisGA1 | ||||
| UpsrfaAF | TCAGCAAAACTGCGTGGTAG | srfaA | [37] | |
| UpsrfaAR | CCAATTTTCGAATTCTTTTACCGCGATAAAAAGTTATTTCCATATGTGTGC | |||
| DwsrfAF | CAGCTCCAGATCCTCTACGCCGGACACGCTTTATATCGTGCCGAA | |||
| DwsrfAR | AAGAAATGATCATAAATACC | |||
| UpFenAF | AGCAAAAACCGGGTCACTAA | fenA | [37] | |
| UpFenAR | CCAATTTTCGAATTCTTTTACCGCGTTCGTCTGACATGACAAGCA | |||
| DwFenAF | CAGCTCCAGATCCTCTACGCCGGACAAAGGACTTTAATTTCATAAAAAGGTG | |||
| DwFenAR | CCTTTTTGAGAAGAGAAGAAAAAG | |||
| UpItuAF | ATGCAGGAAATAGGGGTGAA | ituA | [37] | |
| UpItuAR | CCAATTTTCGAATTCTTTTACCGCGGGTATACATAGGTCCCCTCCTG | |||
| DwItuAF | CAGCTCCAGATCCTCTACGCCGGACCAATTGAACTTTTAGGGAAAAGCA | |||
| DwItuAR | GCGACTAACGTATCGGGTTG | |||
| Antibiotic marker | ||||
| PhleoF | GTCATAGCTGTTTCCTGCCAAAAGGGGGTTTCATTTT | Phleomycin marker | [37] | |
| PhleoR | ACTGGCCGTCGTTTTACTCCAATAAATGCGACACCAA | |||
| Strain a | CLPs Produced (%) b | ||
|---|---|---|---|
| Surfactins | Fengycins | Iturins | |
| B. velezensis GA1wt | 100 ± 4.2 | 100 ± 2.2 | 100 ± 4.8 |
| B. velezensis GA1∆srfaA | 0 ± 0 | 32 ± 2.8 | 37 ± 4.3 |
| B. velezensis GA1∆ituA | 129 ± 1.3 | 82 ± 0.6 | 0 ± 0 |
| B. velezensis GA1∆fenA | 139 ± 1.3 | 0 ± 0 | 121 ± 0.9 |
| B. velezensis GA1∆srfaA-ituA | 0 ± 0 | 18 ± 9.7 | 0 ± 0 |
| B. velezensis GA1∆srfaA-fenA | 0 ± 0 | 0 ± 0 | 69 ± 2.1 |
| B. velezensis GA1∆fenA-ituA | 137 ± 2.9 | 0 ± 0 | 0 ± 0 |
| B. velezensis RHF4.1–25 | 81 ± 15 | 104 ± 12.9 | 96 ± 18 |
References
- Dent, D. A world perspective on acid sulphate soils. Geoderma 1995, 67, 263–276. [Google Scholar] [CrossRef]
- Fältmarsch, R.M.; Åström, M.E.; Vuori, K.M. Environmental risks of metals mobilised from acid sulphate soils in Finland: A literature review. Boreal Environ. Res. 2008, 13, 444–456. [Google Scholar]
- Tanaka, A.; Navasero, S.A. Growth of the rice plant on acid sulfate soils. Soil Sci. Plant Nutr. 1966, 12, 23–30. [Google Scholar] [CrossRef]
- Olivier, H.; Hanhart, K.; Phung, M.; BoumaWater, J. Management for rice cultivation on acid sulpahte soils in the plain of reeds, Vietnam. Agric. Water Manag. 2000, 46, 91–109. [Google Scholar]
- Ren, D.T.T.; Tinh, T.K.; Minh, N.T.N.; Linh, T.B. Applying mixed manure and inorganic phosphorus fertiliser to improve rice yield on acid sulfate soil (Hydraquentic Sulfaquept). Soil Res. 2004, 42, 693. [Google Scholar] [CrossRef]
- Aktar, W.; Sengupta, D.; Chowdhury, A. Impact of pesticides use in agriculture: Their benefits and hazards. Interdiscip. Toxicol. 2009, 2, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bourguet, D.; Guillemaud, T. Sustainable Agriculture Reviews; Lichtfouse, E., Ed.; Springer International Publishing: Cham, Switzerland, 2016; Volume 19, ISBN 978-3-319-26776-0. [Google Scholar]
- Dinh, T.X. An Overview of Agricultural Pollution in Vietnam. Agric. Pollut. Vietnam 2017. [Google Scholar] [CrossRef]
- Wilson, R.A.; Talbot, N.J. Under pressure: Investigating the biology of plant infection by Magnaporthe oryzae. Nat. Rev. Microbiol. 2009, 7, 185–195. [Google Scholar] [CrossRef] [PubMed]
- Martin-Urdiroz, M.; Oses-Ruiz, M.; Ryder, L.S.; Talbot, N.J. Investigating the biology of plant infection by the rice blast fungus Magnaporthe oryzae. Fungal Genet. Biol. 2016, 90, 61–68. [Google Scholar] [CrossRef]
- Hamer, J.E.; Howard, R.J.; Chumley, F.G.; Valent, B. A mechanism for surface attachment in spores of a plant pathogenic fungus. Science 1988, 239, 288–290. [Google Scholar] [CrossRef] [PubMed]
- Thuan, N.T.N.; Bigirimana, J.; Roumen, E.; Van Der Straeten, D.; Höfte, M. Molecular and pathotype analysis of the rice blast fungus in North Vietnam. Eur. J. Plant. Pathol. 2006, 114, 381–396. [Google Scholar] [CrossRef]
- Chen, W.C.; Chiou, T.Y.; Delgado, A.L.; Liao, C.S. The control of rice blast disease by the novel biofungicide formulations. Sustainability 2019, 11, 3449. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Zhao, L.; Chen, W.; Dong, Y.; Yang, C.; Li, C.; Xu, H.; Gao, X.; Chen, R.; Li, L.; et al. Isolation and evaluation of Bacillus velezensis ZW-10 as a potential biological control agent against Magnaporthe oryzae. Biotechnol. Biotechnol. Equip. 2020, 34, 714–724. [Google Scholar] [CrossRef]
- He, Y.; Zhu, M.; Huang, J.; Hsiang, T.; Zheng, L. Biocontrol potential of a Bacillus subtilis strain BJ-1 against the rice blast fungus Magnaporthe oryzae. Can. J. Plant. Pathol. 2019, 41, 47–59. [Google Scholar] [CrossRef]
- Ma, Z.; Zhang, S.; Sun, K.; Hu, J. Identification and characterization of a cyclic lipopeptide iturin A from a marine-derived Bacillus velezensis 11-5 as a fungicidal agent to Magnaporthe oryzae in rice. J. Plant. Dis. Prot. 2020, 127, 15–24. [Google Scholar] [CrossRef]
- Omoboye, O.O.; Geudens, N.; Duban, M.; Chevalier, M.; Flahaut, C.; Martins, J.C.; Leclère, V.; Oni, F.E.; Höfte, M. Pseudomonas sp. COW3 produces new bananamide-type cyclic lipopeptides with antimicrobial activity against Pythium myriotylum and Pyricularia oryzae. Molecules 2019, 24, 4170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rong, S.; Xu, H.; Li, L.; Chen, R.; Gao, X.; Xu, Z. Antifungal activity of endophytic Bacillus safensis B21 and its potential application as a biopesticide to control rice blast. Pestic. Biochem. Physiol. 2020, 162, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Sahu, K.P.; Kumar, A.; Patel, A.; Kumar, M.; Gopalakrishnan, S.; Prakash, G.; Rathour, R.; Gogoi, R. Rice Blast Lesions: An Unexplored Phyllosphere Microhabitat for Novel Antagonistic Bacterial Species Against Magnaporthe oryzae. Microb. Ecol. 2020, 731–745. [Google Scholar] [CrossRef] [PubMed]
- Sha, Y.; Zeng, Q.; Sui, S. Screening and application of Bacillus strains isolated from nonrhizospheric rice soil for the biocontrol of rice blast. Plant. Pathol. J. 2020, 36, 231–243. [Google Scholar] [CrossRef]
- Wei, Y.; Li, L.; Hu, W.; Ju, H.; Zhang, M.; Qin, Q.; Zhang, S.; Li, G. Suppression of rice blast by bacterial strains isolated from cultivated soda saline-sodic soils. Int. J. Environ. Res. Public Health 2020, 17, 5248. [Google Scholar] [CrossRef]
- Rais, A.; Shakeel, M.; Hafeez, F.Y.; Hassan, M.N. Plant growth promoting rhizobacteria suppress blast disease caused by Pyricularia oryzae and increase grain yield of rice. BioControl 2016, 61, 769–780. [Google Scholar] [CrossRef]
- Rais, A.; Jabeen, Z.; Shair, F.; Hafeez, F.Y.; Hassan, M.N. Bacillus spp., a bio-control agent enhances the activity of antioxidant defense enzymes in rice against Pyricularia oryzae. PLoS ONE 2017, 12, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Ma, Z.; Ongena, M.; Höfte, M. The cyclic lipopeptide orfamide induces systemic resistance in rice to Cochliobolus miyabeanus but not to Magnaporthe oryzae. Plant. Cell Rep. 2017, 36, 1731–1746. [Google Scholar] [CrossRef] [PubMed]
- Omoboye, O.O.; Oni, F.E.; Batool, H.; Yimer, H.Z.; De Mot, R.; Höfte, M. Pseudomonas cyclic lipopeptides suppress the rice blast fungus Magnaporthe oryzae by induced resistance and direct antagonism. Front. Plant. Sci. 2019, 10, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wulff, E.G.; Mguni, C.M.; Mansfeld-Giese, K.; Fels, J.; Lübeck, M.; Hockenhull, J. Biochemical and molecular characterization of Bacillus amyloliquefaciens, B. subtilis and B. pumilus isolates with distinct antagonistic potential against Xanthomonas campestris pv. campestris. Plant Pathol. 2002, 51, 574–584. [Google Scholar] [CrossRef]
- Gopal, N.; Hill, C.; Ross, P.R.; Beresford, T.P.; Fenelon, M.A.; Cotter, P.D. The prevalence and control of Bacillus and related spore-forming bacteria in the dairy industry. Front. Microbiol. 2015, 6, 1–38. [Google Scholar] [CrossRef]
- Cochrane, S.A.; Vederas, J.C. Lipopeptides from Bacillus and Paenibacillus spp.: A Gold Mine of Antibiotic Candidates. Med. Res. Rev. 2016, 36, 4–31. [Google Scholar] [CrossRef] [PubMed]
- Luo, C.; Liu, X.; Zhou, H.; Wang, X.; Chen, Z. Nonribosomal peptide synthase gene clusters for lipopeptide biosynthesis in Bacillus subtilis 916 and their phenotypic functions. Appl. Environ. Microbiol. 2015, 81, 422–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ongena, M.; Jourdan, E.; Adam, A.; Paquot, M.; Brans, A.; Joris, B.; Arpigny, J.L.; Thonart, P. Surfactin and fengycin lipopeptides of Bacillus subtilis as elicitors of induced systemic resistance in plants. Environ. Microbiol. 2007, 9, 1084–1090. [Google Scholar] [CrossRef]
- Chandler, S.; Van Hese, N.; Coutte, F.; Jacques, P.; Höfte, M.; De Vleesschauwer, D. Role of cyclic lipopeptides produced by Bacillus subtilis in mounting induced immunity in rice (Oryza sativa L.). Physiol. Mol. Plant. Pathol. 2015, 91, 20–30. [Google Scholar] [CrossRef]
- Abdellaziz, L.; Chollet, M.; Abderrahmani, A.; Béchet, M.; Yaici, L.; Chataigné, G.; Arias, A.A.; Leclère, V.; Jacques, P. Lipopeptide biodiversity in antifungal Bacillus strains isolated from Algeria. Arch. Microbiol. 2018, 200, 1205–1216. [Google Scholar] [CrossRef]
- Ongena, M.; Jacques, P. Bacillus lipopeptides: Versatile weapons for plant disease biocontrol. Trends Microbiol. 2008, 16, 115–125. [Google Scholar] [CrossRef]
- Arguelles-Arias, A.; Ongena, M.; Halimi, B.; Lara, Y.; Brans, A.; Joris, B.; Fickers, P. Bacillus amyloliquefaciens GA1 as a source of potent antibiotics and other secondary metabolites for biocontrol of plant pathogens. Microb. Cell Fact. 2009, 8, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Touré, Y.; Ongena, M.; Jacques, P.; Guiro, A.; Thonart, P. Role of lipopeptides produced by Bacillus subtilis GA1 in the reduction of grey mould disease caused by Botrytis cinerea on apple. J. Appl. Microbiol. 2004, 96, 1151–1160. [Google Scholar] [CrossRef] [PubMed]
- Talbot, N.J.; Ebbole, D.J.; Hamer, J.E. Identification and characterization of MPG1, a gene involved in pathogenicity from the rice blast fungus Magnaporthe grisea. Plant. Cell 1993, 5, 1575–1590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andric, S.; Meyer, T.; Rigolet, A.; Mccann, A.; Pauw, E. De Chelator sensing and lipopeptide interplay mediates molecular interspecies interactions between soil bacilli and pseudomonads. bioRxiv 2021. [Google Scholar] [CrossRef]
- Oni, F.E.; Geudens, N.; Omoboye, O.O.; Bertier, L.; Hua, H.G.K.; Adiobo, A.; Sinnaeve, D.; Martins, J.C.; Höfte, M. Fluorescent Pseudomonas and cyclic lipopeptide diversity in the rhizosphere of cocoyam (Xanthosoma sagittifolium). Environ. Microbiol. 2019, 21, 1019–1034. [Google Scholar] [CrossRef] [Green Version]
- Olorunleke, F.E.; Hua, G.K.H.; Kieu, N.P.; Ma, Z.; Höfte, M. Interplay between orfamides, sessilins and phenazines in the control of Rhizoctonia diseases by Pseudomonas sp. CMR12a. Environ. Microbiol. Rep. 2015, 7, 774–781. [Google Scholar] [CrossRef] [PubMed]
- Sahu, P.K.; Singh, S.; Gupta, A.R.; Gupta, A.; Singh, U.B.; Manzar, N.; Bhowmik, A.; Singh, H.V.; Saxena, A.K. Endophytic bacilli from medicinal-aromatic perennial Holy basil (Ocimum tenuiflorum L.) modulate plant growth promotion and induced systemic resistance against Rhizoctonia solani in rice (Oryza sativa L.). Biol. Control. 2020, 150, 104353. [Google Scholar] [CrossRef]
- Budiharjo, A.; Jeong, H.; Wulandari, D.; Lee, S.; Ryu, C.-M. Complete Genome Sequence of Bacillus altitudinis P-10, a potential bioprotectant against Xanthomonas oryzae pv. oryzae, isolated from rice rhizosphere in Java, Indonesia. Genome Announc. 2017, 5, e01388-17. [Google Scholar]
- Théatre, A.; Cano-Prieto, C.; Bartolini, M.; Laurin, Y.; Deleu, M.; Niehren, J.; Fida, T.; Gerbinet, S.; Alanjary, M.; Medema, M.H.; et al. The Surfactin-Like Lipopeptides From Bacillus spp.: Natural Biodiversity and Synthetic Biology for a Broader Application Range. Front. Bioeng. Biotechnol. 2021, 9. [Google Scholar] [CrossRef] [PubMed]
- Andrić, S.; Meyer, T.; Ongena, M. Bacillus Responses to Plant-Associated Fungal and Bacterial Communities. Front. Microbiol. 2020, 11, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Naruse, N.; Tenmyo, O.; Kobaru, S.; Kamei, H.; Miyaki, T.; Konishi, M.; Oki, T. Pumilacidin, a complex of new antiviral antibiotics. Production, isolation, chemical properties, structure and biological activity. J. Antibiot. 1990, 43, 267–280. [Google Scholar] [CrossRef] [Green Version]
- De Araujo, L.L.G.C.; Sodré, L.G.P.; Brasil, L.R.; Domingos, D.F.; de Oliveira, V.M.; da Cruz, G.F. Microbial enhanced oil recovery using a biosurfactant produced by Bacillus safensis isolated from mangrove microbiota—Part I biosurfactant characterization and oil displacement test. J. Pet. Sci. Eng. 2019, 180, 950–957. [Google Scholar] [CrossRef]
- Rahman, A.; Uddin, W.; Wenner, N.G. Induced systemic resistance responses in perennial ryegrass against Magnaporthe oryzae elicited by semi-purified surfactin lipopeptides and live cells of Bacillus amyloliquefaciens. Mol. Plant. Pathol. 2015, 16, 546–558. [Google Scholar] [CrossRef]
- Crouzet, J.; Arguelles-Arias, A.; Dhondt-Cordelier, S.; Cordelier, S.; Pršić, J.; Hoff, G.; Mazeyrat-Gourbeyre, F.; Baillieul, F.; Clément, C.; Ongena, M.; et al. Biosurfactants in Plant Protection Against Diseases: Rhamnolipids and Lipopeptides Case Study. Front. Bioeng. Biotechnol. 2020, 8, 1–11. [Google Scholar] [CrossRef]
- Tendulkar, S.R.; Saikumari, Y.K.; Patel, V.; Raghotama, S.; Munshi, T.K.; Balaram, P.; Chattoo, B.B. Isolation, purification and characterization of an antifungal molecule produced by Bacillus licheniformis BC98, and its effect on phytopathogen Magnaporthe grisea. J. Appl. Microbiol. 2007, 103, 2331–2339. [Google Scholar] [CrossRef] [PubMed]
- Pérez-García, A.; Romero, D.; de Vicente, A. Plant protection and growth stimulation by microorganisms: Biotechnological applications of Bacilli in agriculture. Curr. Opin. Biotechnol. 2011, 22, 187–193. [Google Scholar] [CrossRef]
- Mihalache, G.; Balaes, T.; Gostin, I.; Stefan, M.; Coutte, F.; Krier, F. Lipopeptides produced by Bacillus subtilis as new biocontrol products against fusariosis in ornamental plants. Environ. Sci. Pollut. Res. 2018, 25, 29784–29793. [Google Scholar] [CrossRef]













| Strain | Relevant Characteristics | References |
|---|---|---|
| Bacillus altitudinis group | ||
| RHF4.1–26 | Pumilacidin-producer from rice rhizosphere grown in ASS, Vietnam | This study |
| RHF3.1–20 | Pumilacidin-producer from rice rhizosphere grown in ASS, Vietnam | This study |
| Bacillus marisflavi group | ||
| RHF2.1–7 | Non-CLiP-producer from rice rhizosphere grown in ASS, Vietnam | This study |
| Bacillus velezensis group | ||
| RHF4.1–25 | Surfactin, iturin and fengycin-producer from rice rhizosphere grown in ASS, Vietnam | This study |
| GA1wt | Wild type: srf+, itu+, fen+ | [35] |
| GA1∆srfaA | Surfactin mutant: srf−, itu+, fen+ | [37] |
| GA1∆ituA | Iturin mutant: itu−, srf+, fen+ | [37] |
| GA1∆fenA | Fengycin mutant: fen−, itu+, srf+, | [37] |
| GA1∆srfaA-ituA | Surfactin and iturin mutant: srf−, itu−, fen+ | This study |
| GA1∆srfaA-fenA | Surfactin and fengycin mutant: srf−, fen−, itu+ | This study |
| GA1∆fenA-ituA | Iturin and fengycin mutant: itu−, fen−, srf+ | This study |
| Pyricularia oryzae VT5M1 | Rice blast pathogen from Vietnam | [12] |
| Treatment | Population Density (log CFU g−1 of Fresh Root) |
|---|---|
| B. marisflavi RHF2.1–7 | 8.41 ± 0.06 c |
| B. velezensis RHF4.1–25 | 7.97 ± 0.11 b |
| B. altitudinis RHF4.1–26 | 7.70 ± 0.08 a |
| B. altitudinis RHF3.1–20 | 8.15 ± 0.32 b |
| Treatment | Population Density (log CFU g−1 of Fresh Root) |
|---|---|
| B. marisflavi RHF2.1–7 | 7.31 ± 0.59 b |
| B. velezensis RHF4.1–25 | 6.56 ± 0.59 a |
| B. altitudinis RHF4.1–26 | 7.41 ± 0.65 b |
| B. altitudinis RHF3.1–20 | 7.54 ± 0.37 b |
| Treatment | CLPs Produced | Population Density (in log CFU g−1 of Fresh Root) |
|---|---|---|
| B. velezensis RHF4.1–25 | Surfactin, iturin and fengycin | 6.16 ± 0.63 a |
| B. velezensis GA1wt | Surfactin, iturin and fengycin | 6.12 ± 0.57 a |
| B. velezensis GA1∆srfaA | Iturin and fengycin | 6.60 ± 0.31 a |
| B. velezensis GA1∆ituA | Surfactin and fengycin | 6.41 ± 0.52 a |
| B. velezensis GA1∆fenA | Surfactin and iturin | 6.38 ± 0.50 a |
| B. velezensis GA1∆fenA-ituA | Surfactin, iturin and fengycin | 6.26 ± 0.40 a |
| Treatment | CLPs Produced | Population Density (in log CFU g−1 of Fresh Root) |
|---|---|---|
| B. velezensis RHF4.1–25 | Surfactin, iturin and fengycin | 6.32 ± 0.42 a |
| B. velezensis GA1wt | Surfactin, iturin and fengycin | 6.49 ± 0.23 a |
| B. velezensis GA1∆srfaA | Iturin and fengycin | 6.46 ± 0.45 a |
| B. velezensis GA1∆ituA | Surfactin and fengycin | 6.52 ± 0.30 a |
| B. velezensis GA1∆fenA | Surfactin and iturin | 6.38 ± 0.29 a |
| B. velezensis GA1∆srfaA-ituA | Fengycin | 6.54 ± 0.28 a |
| B. velezensis GA1∆srfaA-fenA | Iturin | 6.45 ± 0.30 a |
| B. velezensis GA1∆fenA-ituA | Surfactin | 6.70 ± 0.64 a |
| Treatment | CLPs Produced | Population Density (in log CFU g−1 of Fresh Root) |
|---|---|---|
| B. velezensis RHF4.1–25 | Surfactin, iturin and fengycin | 6.77 ± 0.31 a |
| B. velezensis GA1wt | Surfactin, iturin and fengycin | 6.66 ± 0.40 a |
| B. velezensis GA1∆srfaA | Iturin and fengycin | 6.21 ± 0.13 a |
| B. velezensis GA1∆ItuA | Surfactin and fengycin | 6.33 ± 0.16 a |
| B. velezensis GA1∆fenA | Surfactin and iturin | 6.50 ± 0.17 a |
| Treatment | CLPs Produced | Spore Germination (%) |
|---|---|---|
| Control (LB broth) | - | 97 ± 1 |
| B. velezensis RHF4.1–25 | Surfactin, iturin and fengycin | 0 ± 0 |
| B. velezensis GA1wt | Surfactin, iturin and fengycin | 0 ± 0 |
| B. velezensis GA1∆srfaA | Iturin and fengycin | 0 ± 0 |
| B. velezensis GA1∆ituA | Surfactin and fengycin | 0 ± 0 |
| B. velezensis GA1∆fenA | Surfactin and iturin | 0 ± 0 |
| B. velezensis GA1∆srfaA-ituA | Fengycin | 6 ± 3 |
| B. velezensis GA1∆srfaA-fenA | Iturin | 0 ± 0 |
| B. velezensis GA1∆fenA-ituA | Surfactin | 0 ± 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lam, V.B.; Meyer, T.; Arias, A.A.; Ongena, M.; Oni, F.E.; Höfte, M. Bacillus Cyclic Lipopeptides Iturin and Fengycin Control Rice Blast Caused by Pyricularia oryzae in Potting and Acid Sulfate Soils by Direct Antagonism and Induced Systemic Resistance. Microorganisms 2021, 9, 1441. https://doi.org/10.3390/microorganisms9071441
Lam VB, Meyer T, Arias AA, Ongena M, Oni FE, Höfte M. Bacillus Cyclic Lipopeptides Iturin and Fengycin Control Rice Blast Caused by Pyricularia oryzae in Potting and Acid Sulfate Soils by Direct Antagonism and Induced Systemic Resistance. Microorganisms. 2021; 9(7):1441. https://doi.org/10.3390/microorganisms9071441
Chicago/Turabian StyleLam, Van Bach, Thibault Meyer, Anthony Arguelles Arias, Marc Ongena, Feyisara Eyiwumi Oni, and Monica Höfte. 2021. "Bacillus Cyclic Lipopeptides Iturin and Fengycin Control Rice Blast Caused by Pyricularia oryzae in Potting and Acid Sulfate Soils by Direct Antagonism and Induced Systemic Resistance" Microorganisms 9, no. 7: 1441. https://doi.org/10.3390/microorganisms9071441