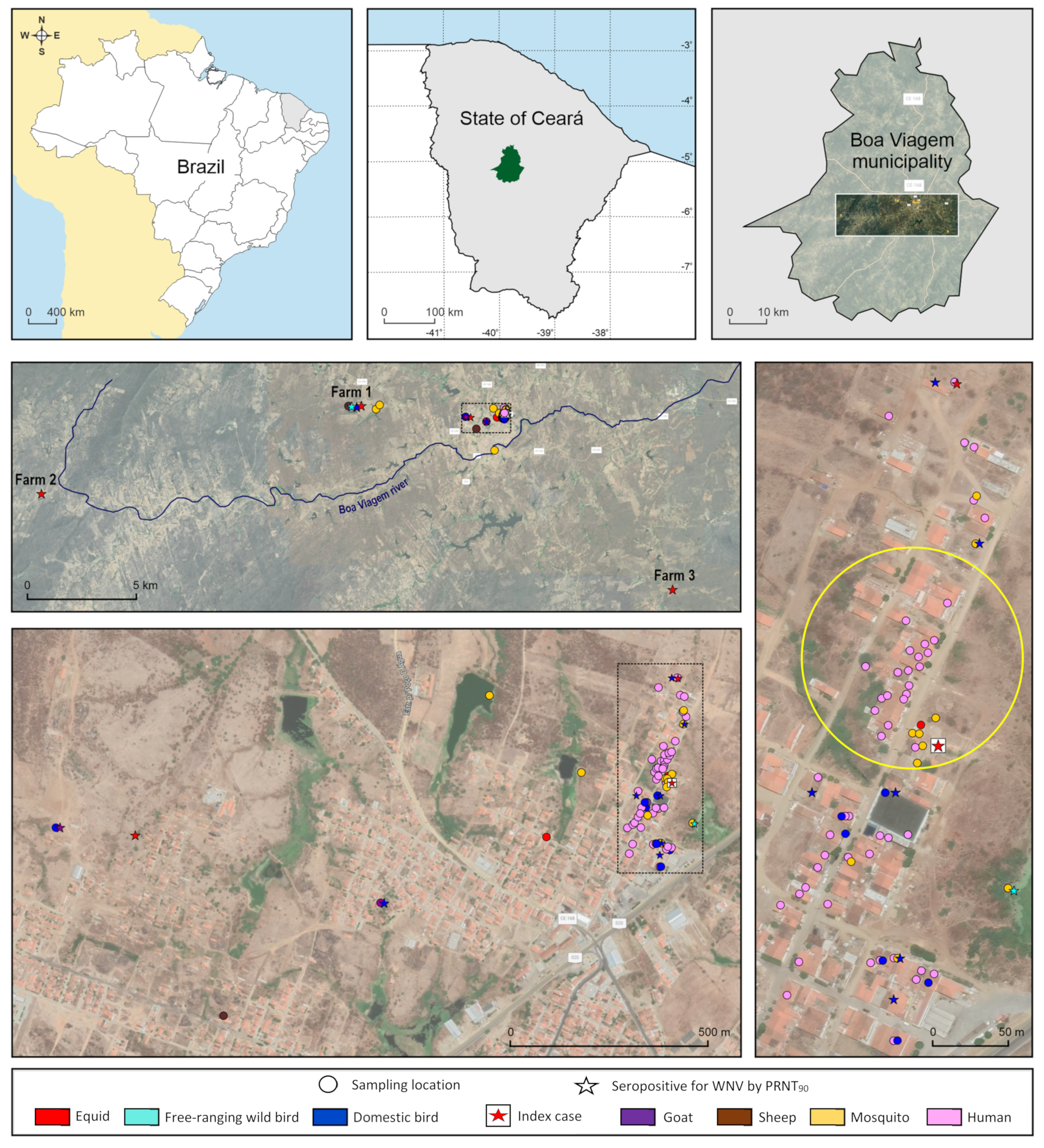
4.1. Circulation of WNV in Boa Viagem
Epidemiological data associated with clinical, molecular, and serological diagnosis are the most reliable approach to confirm arbovirus infection. Findings presented here, including the molecular confirmation of the clinical case and the detection of highly specific neutralizing antibodies in various local vertebrate hosts, including potential amplifying bird species, confirm the past circulation of WNV in the municipality of Boa Viagem, CE.
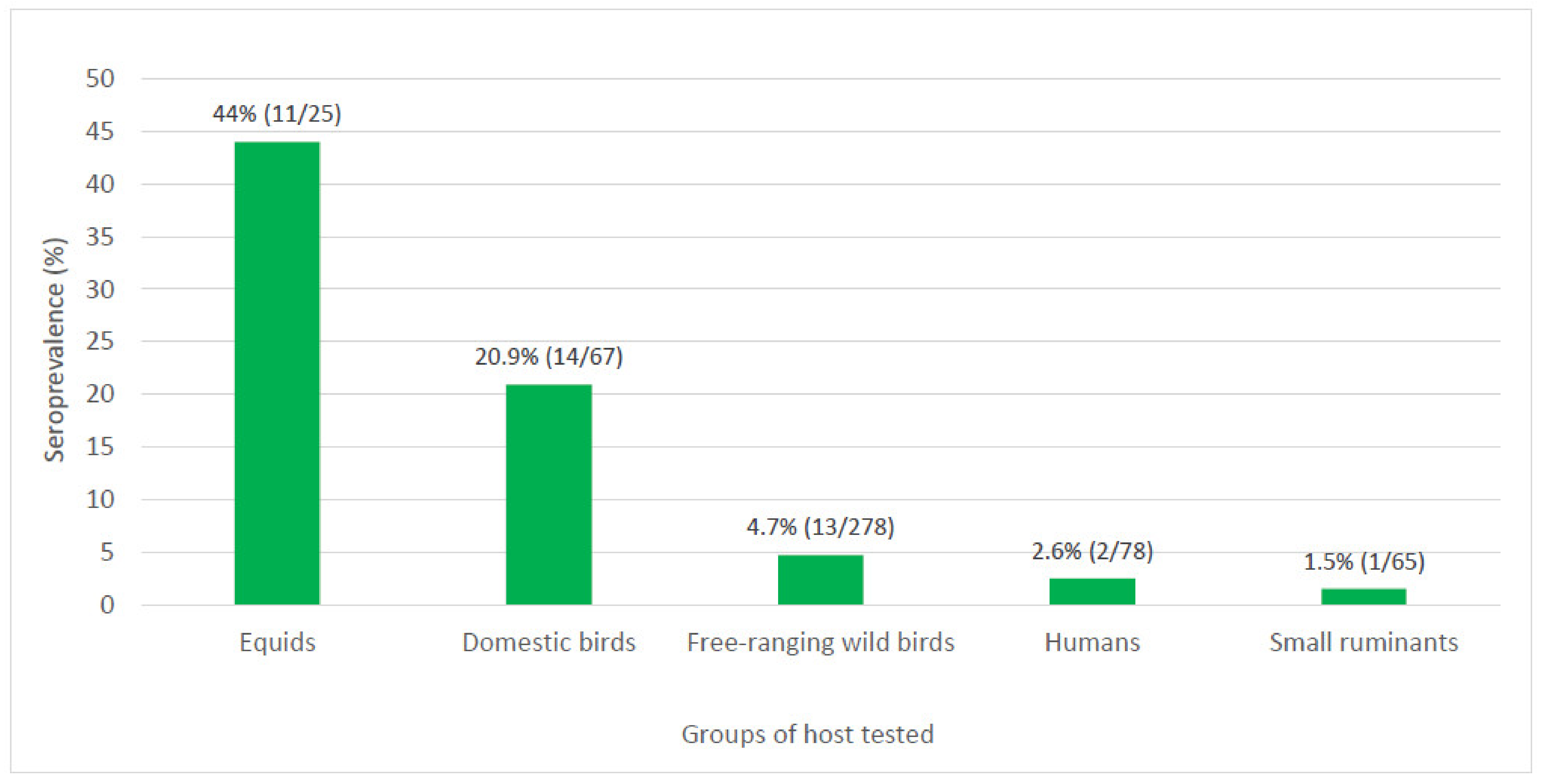
Except for ovine, all other vertebrate groups locally investigated, including humans, equids, domestic, and wild birds, and goats had at least one individual that tested seropositive for WNV by a highly specific serological test. Higher prevalence for WNV was observed in equids and birds when compared to humans and small ruminants, and the seroprevalence varied greatly among species, going from 44% in equids to 1.5% in small ruminants (
Figure 2). The variation of WNV exposure by host groups has been reported elsewhere. In a study conducted in the United States (USA) after the 1999 epidemic in New York City, seroprevalence in birds reached 50% when compared to 2.6% in humans [
27].
The infection by WNV in the brain tissue of the index case was confirmed by real-time RT-PCR, and 5 out of 11 (45%) WNV-seropositive equids had a confirmed history of staying at the PLI area at some point. These findings corroborate the role of the PLI area as a source of infection in Boa Viagem. Interestingly, the area where the index case was reported has a pond that used to be surrounded by trees and accessed by the equids after training. The area was recently cleared for settlement, and the niche impact for the surge of WNV horse clinical infection in Boa Viagem needs further investigation.
4.3. Potential Amplifying Hosts of WNV in Boa Viagem
Although the main amplifying bird species of WNV in Boa Viagem remains to be confirmed, multiplying the squared seroprevalence per species by their capture prevalence, we grossly speculated amplification capacity of the captured species in Boa Viagem. Because of the bias of bird sampling in Farm 1, which had as one of the targets a House sparrow communal nest site, we considered three scenarios, including both sites together (
Table 8), PLI alone (
Table S2), and Farm 1 alone (
Table S3). Combining results, stronger amplification capacity was observed for Orange-headed tanager and Rufous-bellied thrush and Ruddy ground dove. When considering only species with more than ten specimens captured, Ruddy ground dove and Sayaca tanager presented stronger amplification capacity.
Similar results were seen when analyzing only captures from the PLI area. While stronger amplification capacity was observed for Blue-winged parrotlet and Rufous-bellied thrush, among the most abundant species, Sayaca tanager and Ruddy ground dove presented the highest amplification capacity (
Table S2).
At Farm 1, Orange-headed tanager and Ruddy ground dove had the higher amplification capacity, and the House sparrow that was the most captured species presented 4.8% seroprevalence for WNV (
Table S3). Any of these aforementioned species are candidates for WNV amplification in Boa Viagem. It is possible that more than one of these species played a role in WNV amplification in Boa Viagem. In Louisiana, which has a humid subtropical climate, a variety of passerine bird species combined play an important role as amplifying hosts in the WNV transmission cycles [
45].
Of the potential amplifying hosts of WNV in Boa Viagem, only the Ruddy ground dove is nonpasserine, which is seen to be likely less competent to infect blood-feeding mosquitoes elsewhere. However, other studies have demonstrated that doves are being exposed to WNV and may contribute to the mass of WNV-infectious mosquitoes in some regions as Colorado in the USA [
42]. In Argentina, free-ranging specimens of White-tipped Dove (
Leptotila verreauxi) were found seropositive for WNV [
46], and in experimental infections, Picui ground doves presented the highest and longest viremia among three autochthonous bird species evaluated. These results suggest that Picui ground dove is a potential amplifying host for WNV in Argentina [
47].
Serological evidence of WNV infection has also been reported in Ruddy ground dove from Guatemala and Mexico [
41,
48]. In a study conducted in different regions of Brazil between 2008 and 2010, 24 Ruddy ground doves were tested, but no evidence of WNV infection was reported [
6]. In the present study, 2 (15.4%) out 13 individuals tested were seropositive for WNV and Ruddy ground dove presented the third highest amplification capacity in Boa Viagem (
Table 8). Considering the abovementioned evidence and its wide geographical distribution, Ruddy ground doves and other species of columbiformes should not be ruled out as potential amplifying hosts of WNV in Boa Viagem or elsewhere in the country.
Although Rufous-bellied thrush and the Band-tailed hornero presented high seroprevalence for WNV among all species with a sample size of
n > 5 (
Table 7), these species were not abundant, corresponding for less than 2% each of all birds captured. Evidence of these species acting as amplifying hosts of WNV worldwide remains unreported, and most evidence is restricted to eventual serological findings of WNV exposure in closely related species. Rufous hornero, which is a species of Furnariidae, as the Band-tailed hornero, has been exposed to WNV, and if proven to be competent amplifying host, could play role in WNV transmission in Argentina [
49]. Regarding the Rufous-bellied thrush, of four individuals sampled in Buenos Aires between 2012 and 2013, none of them were seropositive for WNV [
50]. In Brazil, an investigation for WNV conducted between 2008 and 2010, 18 Rufous-bellied thrush tested negative for WNV by molecular and serological methods [
6]. Between 2012 and 2013, free-ranging birds, including 13 individuals of Rufous-bellied thrush captured from green areas of the city of São Paulo, southeastern Brazil, tested negative for flaviviruses by real-time RT-PCR [
51].
Other WNV antibody-positive trushes and congeners have been eventually detected in Central and South America [
52,
53,
54]. Serological evidence of WNV infection was detected in pale-breasted thrush in Venezuela [
55]. In Guatemala, Clay-colored thrushes were abundant in a WNV transmission area but were not considered competent amplifying hosts of WNV [
41]. Similar results were seen in experimental infections conducted with the same species from Mexico [
56].
In the present study, the Orange-headed tanager presented the highest amplification capacity (
Table 8). However, only two individuals were captured in Boa Viagem, corresponding to less than 1% of all specimens captured. On the other hand, Sayaca tanager, which is another Thraupidae, was the fifth most captured species in Boa Viagem, and among 19 specimens tested at the PLI area, 10.5% were seropositive for WNV (
Table S2). Evidence of WNV exposure has not been commonly reported for these two species, but because its local occurrence and serological evidence of WNV infection, these two species of tanager are potential amplifying hosts of WNV in CE, and other close locations where it occurs. Because of its abundance at the PLI area, and in other parts of the country, Sayaca tanager may be of a particular interest. The potential participation of this species in flavivirus enzootic cycles of transmission in Brazil has already been investigated. Between 2012 and 2013, 29 free-ranging Sayaca tanager individuals collected in the city of São Paulo, southeastern Brazil, tested negative by real-time RT-PCR for flaviviruses [
51]. As part of an investigation for WNV in northeast Brazil, four Sayaca tanager individuals were tested, but no evidence of WNV infection was reported [
6].
The House sparrow, which is primarily an urban species, is also a potential amplifying host of WNV in Brazil. House sparrows are among the leaders of infected birds in the urban areas worldwide, and their behavior is believed to enhance this species’ involvement, as these birds also nest and roost communally [
57]. In experimental infection studies, House sparrows are strong amplifiers of WNV, regardless of the strain of virus evaluated [
58]. In the USA, where the dynamics of enzootic transmission of WNV has been more explored, House sparrow and House finch have been shown to be not only exposed countrywide but also are among the most competent amplifying species for WNV [
59].
In tropical ecosystems, which present a larger avian species diversity, data regarding the participation of House sparrow and other common species in WNV cycles of transmission remain scarce. In the present study, 3.9% of House sparrows tested were seropositive for WNV, but this species presented the second lowest amplification capacity among species with >5 individuals tested in Boa Viagem (
Table 7). However, the House sparrow’s great amplifying potential may not always be predicted by its seroprevalence.
The House sparrow was the most captured species during the investigation (
Figure S1), and it is the only one among all species captured in Boa Viagem known to die from infection with the New York 1999 strain of WNV [
34], which is the same lineage recently isolated in Brazil [
10]. The susceptibility of local sparrows to fatal infection by WNV in Boa Viagem remains unknown, but unnoticed mortality in this species could potentially contribute to a low detection rate for seropositive individuals. Notably, all 14 House sparrows captured and tested from the PLI area were seronegative for WNV. Between 2008 and 2010, eight individuals collected in southern Brazil tested negative for WNV by molecular and serological methods [
6].
Evidence of WNV circulation in Brazil has been reported since 2011, but potential amplifying bird species in the country remain unknown. In the most recent WNV investigation conducted with free-ranging bird of prey in the country, all samples from southern Brazil tested negative by real-time RT-PCR and PRNT [
60].
We report here evidence of WNV exposure in different bird species collected in Boa Viagem. As discussed above, it is possible that some of these species that were seropositive for WNV in Boa Viagem may have potentially acted as amplifying hosts in the region. When considering only the species with at least a dozen individuals tested, for a more significant analysis, the seroprevalence observed among them is lower than commonly observed in avian surveys conducted during WNV outbreaks in the USA [
17]. The lower prevalence observed in species captured in Boa Viagem may reflect the low or diluted local enzootic circulation of WNV. No other clinical cases of WNV infection have been confirmed in Boa Viagem since the index case was reported in June of 2019.
4.4. Arrival of WNV to Boa Viagem
If WNV has silently circulated in birds of Boa Viagem for a while, or if its arrival is a recent event, remains unclear. Between 2012 and 2013, oropharyngeal and cloacal swabs from 529 wild birds from 89 species captured also in the northeast region of the country found no evidence of active circulation of WNV [
61]. On the other hand, hemagglutination-inhibition antibodies for WNV have been found in birds sampled after the human case reported in the neighbor state of Piauí (P Lima 2021, personal communication, 4 May). Hypothetically, WNV could have recently circulated in Boa Viagem region in low levels through temporary enzootic cycles of transmission resulting in sporadic spill-over events driven by unknown ecological factors. Ecological factors could include migration movements of potential amplifying birds and/or the increase of vector population density.
This hypothesis is corroborated by the recent report that the ancient strain NY99 was the cause of WNV infection in the equid epizootic occurred in southeast Brazil [
13]. The NY99 was the first strain of WNV detected in 1999 in the USA and between 2001 and 2003 went extinct after being displaced by other strains [
62]. A low level of transmission potentially due to local ecological conditions could be preventing large outbreaks and evolution of WNV in Brazil, as it has done in the USA. High biodiversity as dilution factor associated with immunity to other flaviviruses and potential limited vector competence in arrival sites could be restricting, at least for now, WNV amplification to isolated and temporary non-continuous cycles of transmission.
The arrival of WNV in Boa Viagem remains unknown, but WNV-infected migratory birds coming from different regions is one of the commonly discussed hypotheses for WNV introductions worldwide [
63]. Bird species in the order Charadriiformes, such as shorebirds and terns, are candidates for carrying WNV from North America to South America due to long-lasting high-level viremias, occasional persistent infectious viral loads in skin, and direct, long-distance flights [
63]. Another possibility would be the introduction of WNV to CE from other areas of Central and South America, including from other regions of Brazil.
The Eared dove (
Zenaida auriculata), which is locally called Avoante is commonly seen in flock gatherings during the rainy season, where it forms breeding colonies in the northeast region of Brazil. It has been classified as migratory species and moves in response to rain movements in the Caatinga, where it appears soon after March in southern CE [
64,
65,
66,
67]. Eared dove is occasionally hunted in Boa Viagem and the eight year old girl who was seropositive for WNV reported consumption of Eared dove meat between April and September 2019. In a study conducted in Argentina, 157 individuals of Eared dove tested negative for WNV antibodies [
50]. In the present study, the only individual that was sampled was seronegative for WNV (
Table S1); therefore, the exposure of Eared dove to WNV in Boa Viagem remains unknown and merits further investigation.
The findings presented here associated with evidence reported in at least ten other states from all regions of the country indicates that WNV is more spread than originally thought [
3,
4,
5,
6,
7,
8,
9,
10,
11,
12,
68,
69,
70]. For decades urban areas in Brazil have been heavily hit by Aedini-borne arboviruses, but less impacted by arboviruses transmitted by Culicine mosquitoes. With the recent increase of WNV reports in Brazil, this empty niche in urban and peri-urban areas should be constantly monitored. The large variety of flaviviruses circulating in Brazil, which could generate heterologous antibodies enough to prevent clinical infections by WNV, has been considered one of the main reasons for the absence of large neurological disorder outbreaks caused by WNV in the country. From 513 samples tested in the present study, seven (1.4%) samples were seropositive for SLEV and one (0.2%) for DENV-1. It is noteworthy that these findings are likely to be underestimated since these samples were found at random during differential diagnosis. No specific investigations for SLEV, DENV, and ILHV were conducted in Boa Viagem.
However, additional factors as recent arrival, local ecological characteristics, and vector and vertebrate host competence may also be involved. Heterologous antibodies for other flaviviruses were not enough to prevent the arrival and spread of DENV in the 1980s in a yellow fever endemic state, neither of ZIKV in a dengue-hyperendemic country in 2015. The impact of cross-reactivity observed among flaviviruses also influences serological diagnostic results. The interpretation of serological tests in flavivirus diagnosis is complex and requires careful evaluation [
17]. With the establishment of WNV in Brazil, where at least ten other flaviviruses of medical importance circulate, the serological diagnostic of flaviviruses, including WNV, tends to become even more challenging. For that reason, we used a conservative threshold for detection of neutralizing antibodies (90%), and we considered monotypic serologic responses to be the most reliable, as these samples reacted with just one of all viruses employed in the tests, with no indication of crossreaction.
4.5. Seroconversion and Potential Maintenance of WNV in Boa Viagem
At least four local domestic birds had four-fold increase in PRNT
90 titers between September and November of 2019 (
Table 5). Moreover, according to the results two more seroconversions occurred among the studied domestic birds. The seroconversions observed in domestic birds reported here are suggestive of recent exposure to WNV. However, stronger evidence relies on a single female helmeted guineafowl that neutralizing antibody titers went from <10 to 160 (
Table 5). The seroconversion observed in the other birds would not be enough to confirm the recent circulation of WNV. Some individuals that are sequentially infected by a heterologous flavivirus species can boost antibody levels against the original virus, resulting in a phenomenon known as ‘original antigenic sin’ which can lead to incorrect diagnoses [
71]. Samples collected in November of 2019 were not tested for other flaviviruses.
Moreover, except for the index horse case, all vertebrate and mosquito samples that were tested for WNV by real-time RT-PCR were negative. To be more likely detected using direct diagnostic methods in sera of terminal hosts, blood sampling must be performed during the short period of viremia of WNV, which may peak before the onset of neurological clinical signs. A recent study conducted in Brazil, reported genetic evidence of WNV in equine red blood cells by portable nanopore sequencing, suggesting whole blood as a potential alternative sample for the diagnostic of WNV in horses [
12].
A complementary approach to investigate active infection of WNV would be testing by real-time RT-PCR plasma samples from free-ranging wild birds. As amplifying hosts, passerines present higher-level and longer viremia making their plasma or serum samples more suitable for direct diagnostic methods for WNV. Because of the low volume of blood (<~200 microliters) drawn for most specimens of birds captured in Boa Viagem, the evaluation of WNV exposure in these animals had to be restricted to the investigation of antibodies. In the present study, an additional subset of tissue samples was collected from individuals that unfortunately died during netting and sampling, and will be further investigated. Future investigations would ideally assess not only antibodies, but also viremia and/or antigen of WNV, preferentially but not restricted to the species that were seropositive for WNV in the present study.
4.6. Potential Screening Methods for WNV Serological Surveys
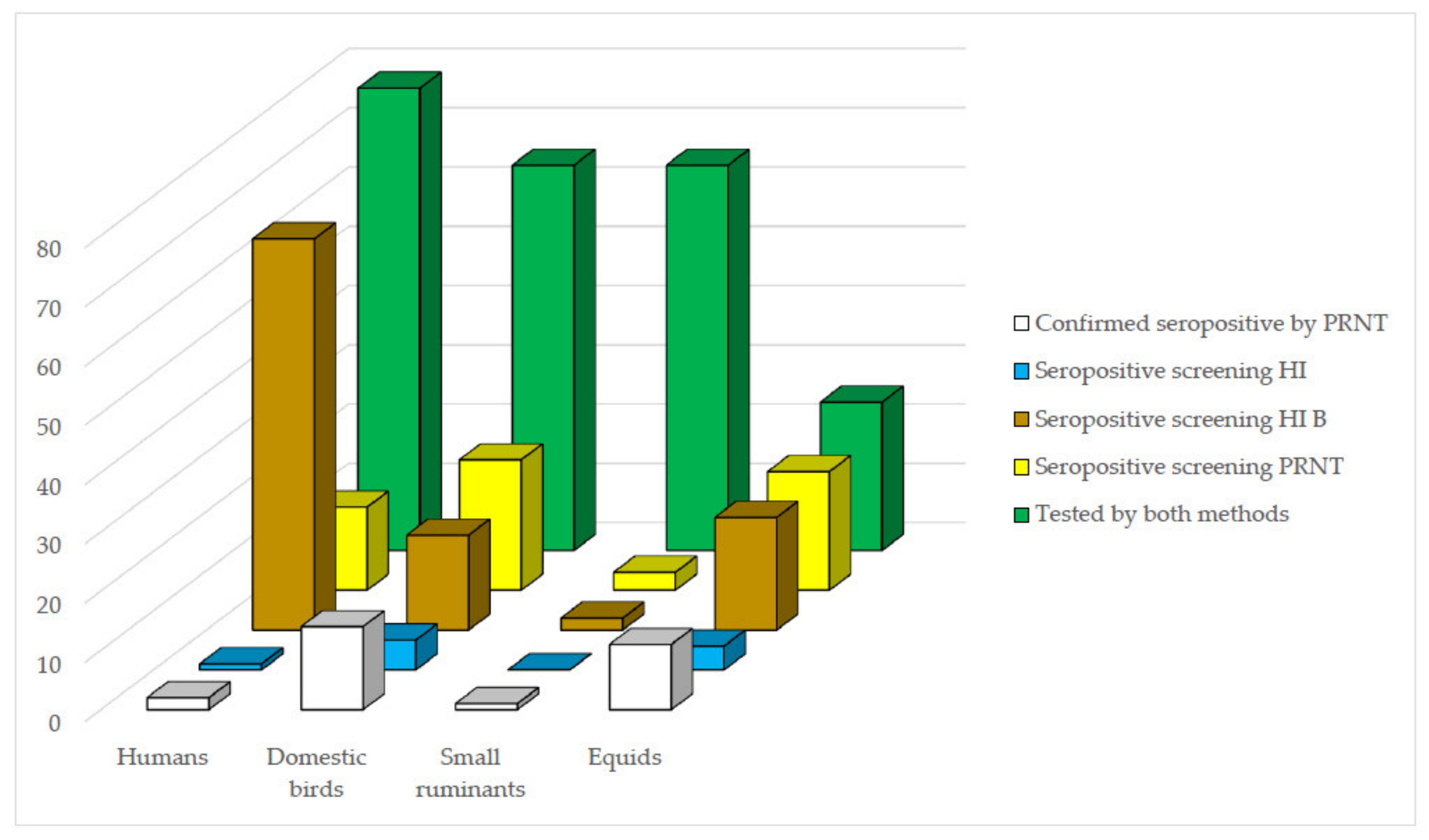
Regarding the potential use of HI as a screening method for confirmatory PRNT
90 for future WNV-specific antibody investigations in Brazil, an optimal screening test should detect all positive samples even at the cost of detecting some false positives. The comparisons between HI and PRNT
90 results demonstrate that the value of HI as a screening method for PRNT
90 for WNV was in general limited, varied according to the vertebrate group tested, and was greatly improved by the use of HI B criterion of seropositivity in most groups. While for equid, domestic bird, and small ruminant samples, the efficiency of HI as a screening method was improved by the use of HI B as selection criterion, for human samples, no much difference was observed between the two criteria of selection (
Figure 3). These results were below expectations particularly for human samples and indicates that screening HI B criterion should be preferred in future similar WNV surveys in domestic animals.
The use of HI as a screening method demonstrated to be particularly informative to assess the local arbovirus diversity. In the present study, grossly 86% of the human residents of Boa Viagem presented HI titer ≥20 to at least one flavivirus. When the same group of samples was tested for alphaviruses and orthobunyaviruses, 24.3% and 0% were reactive, respectively (
Table S1). Among domestic bird samples, similar results were observed. These HI results demonstrated that local hosts were more exposed to flaviviruses than alphaviruses or orthobunyaviruses combined. Moreover, various samples from domestic birds and humans were seropositive by HI to the alphavirus Mayaro virus, and small ruminants and equids were seropositive to the orthobunyaviruses Maguari, Tacaiuma, and Caraparu viruses. Combining HI and PRNT results, at least eight other arboviruses could be silently circulating in Boa Viagem, and this evidences merit further investigation.
The use of PRNT
90 also as a preliminary screening method proved to be useful and effective to exclude seronegative samples. With this approach we were able to reduce in more than 70% the number of samples to be confirmed by endpoint PRNT
90, minimizing the limitation of the use of this assay through adaption for a larger scale use. However, due to the inherent limitations of the PRNT, which includes cell culture production, high level biosafety facilities and highly trained personnel, its use has been restricted to confirmatory tests in reference laboratories. The use of MAC-ELISA as screening method for PRNT could also be an option, but it would need further evaluations. In the present study, from all 12 human samples that were reactive or borderline for WNV by MAC-ELISA, none of them were confirmed to seropositive for WNV by PRNT
90. (
Table S1).
Considering the technical limitations of the abovementioned methods, a surrogate screening test for the serological investigation of WNV in Brazil must be continuously pursued. The suitable screening method would be ideally a fast and qualitative kit for the detection of WNV-specific neutralizing antibodies in competitive ELISA or chromatographic immunoassay platforms, as it has been recently and shortly developed worldwide for the diagnostic of severe acute respiratory syndrome coronavirus 2 during the coronavirus disease pandemic. Few options with similar approach are already available for the investigation of flaviviruses including epitope-blocking ELISA and the VecTest-inhibition assay (VIA). Epitope-blocking ELISA has a good sensibility and specificity for animal samples, but it has demonstrated some limitations for human samples [
72,
73,
74]. VIA, which is a field-deployable rapid test has a high sensitivity for a variety of flaviviruses among a population of equines from Brazil and merits further evaluation [
75].
4.7. Environmental Aspects of WNV Circulation in Brazil
WNV has been reported in different environments worldwide. Evidence of WNV circulation has been reported from temperate and low-average temperature countries to warm and tropical areas [
1]. Although only with data of countrywide ecological studies, we could better evaluate the influence of climate and ecological conditions for the occurrence of WNV in Brazil, the dry and hot conditions observed in the region of Boa Viagem may have had some influence for the local WNV spillover event. Temperature is being related to rates of WNV transmission, and dry weather has already been related to outbreaks of human WNV infections [
76]. High temperature and low rainfall enhanced the probability of chicken seroconversions, which occurred in both urban and rural sites in Guatemala [
41].
Boa Viagem region has a semiarid hot tropical climate with average annual temperature of 29 °C and average rainfall of 717.7 mm concentrated from January to April. Notably, the first human clinical infection by WNV in Brazil was detected in 2014 in the municipality of Aroeiras do Itaim, which has similar climate conditions and is located in the same biome (
caatinga) in the neighbor state of Piauí, which is roughly 300 Km distant from Boa Viagem [
8].
Landscapes where WNV has been detected worldwide vary from wetlands to semiarid regions [
77,
78]. Similar pattern seems to be occurring in Brazil. Besides the clinical infection of WNV in the state of Piauí [
8], the first recognized outbreak of WNV in equines in Brazil occurred at the beginning of the dry season, in an area influenced by the humid Atlantic rainforest biome [
9]. Moreover, the first evidences of WNV in Brazil were reported in equids from the Brazilian Pantanal, which is one of world’s largest wetlands with prolonged flooding and dry seasons [
3,
4,
5,
6,
7]. Drought leads to close contact between avian hosts and mosquitoes around remaining water sources and therefore facilitates the epizootic cycling of WNV within these populations [
79]. Because of the reduced flushing of
Culex mosquitoes during drought events, standing water pools become richer in the organic material that
Culex spp. need in order to thrive. Such water areas might be attractive for several bird species also, which might increase the bird–mosquito interaction [
80,
81]. In a study conducted in the USA, a model predicted that without drought nor warmth there would have been 43% fewer cases of WNV in 2018 [
82]. Based on climate model predictions for climate change and potentially greater drought occurrence in the future, the frequency and relative risk of WNV outbreaks could increase [
76]. This can be particularly important in tropical areas where enzootic transmission cycles have already been identified countrywide, as in Brazil.



 ,
, 




{kind=link}
{kind=link}
{kind=link}