
Potential Industrial Production of a Well-Soluble, Alkaline-Stable, and Anti-Inflammatory Isoflavone Glucoside from 8-Hydroxydaidzein Glucosylated by Recombinant Amylosucrase of Deinococcus geothermalis

 ,
,  ,
,
Abstract
:1. Introduction
2. Results
2.1. Production of DgAS Protein in Recombinant Escherichia coli
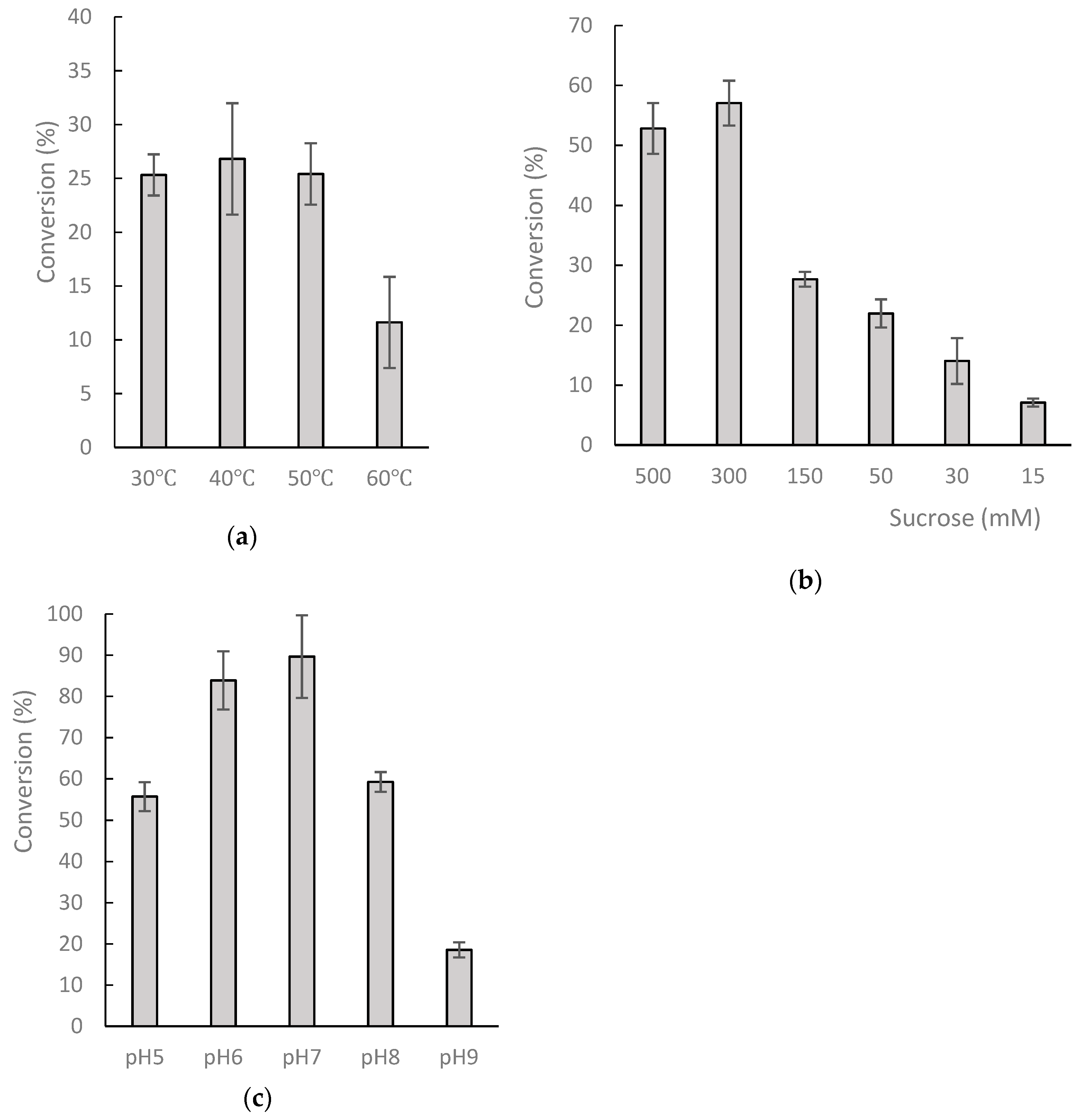
2.2. Biotransformation of 8-OHDe by Recombinant DgAS Protein
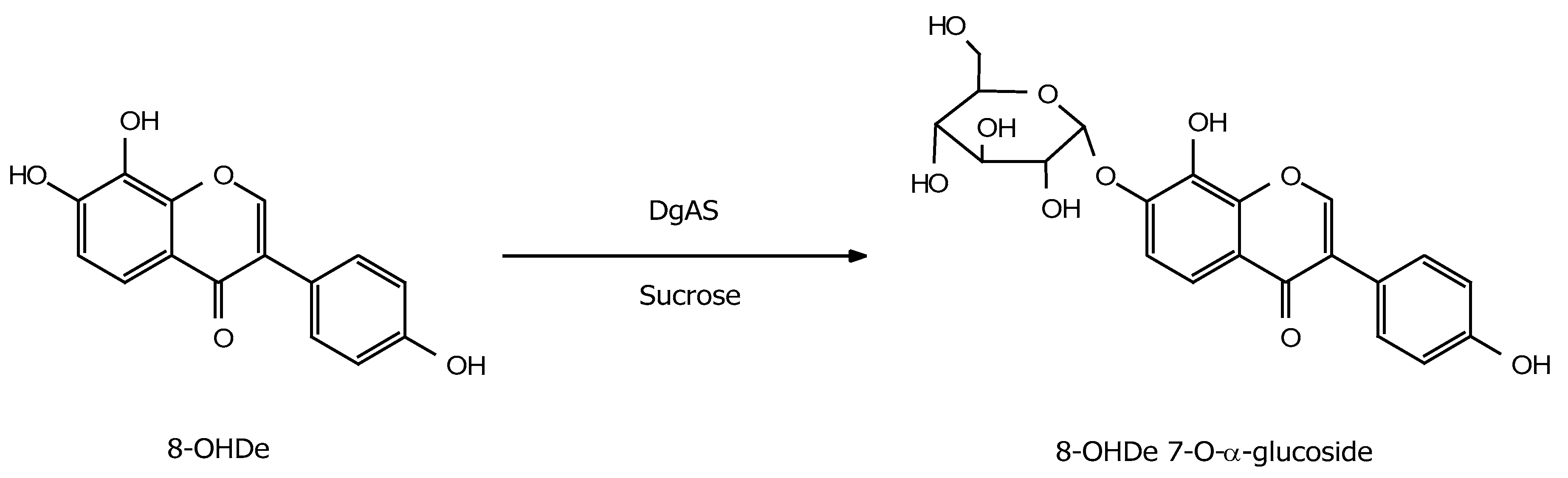
2.3. Identification of the Major Product
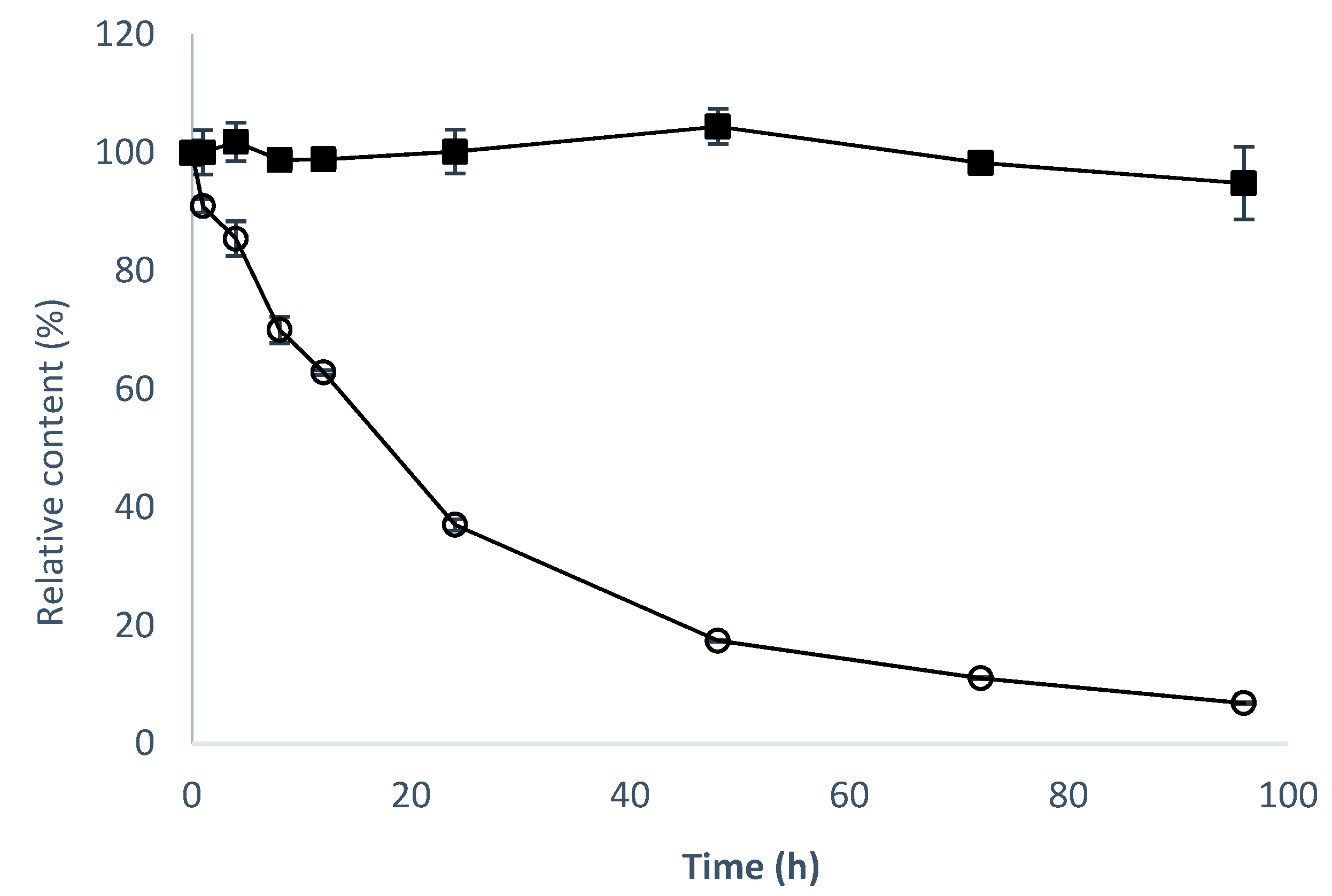
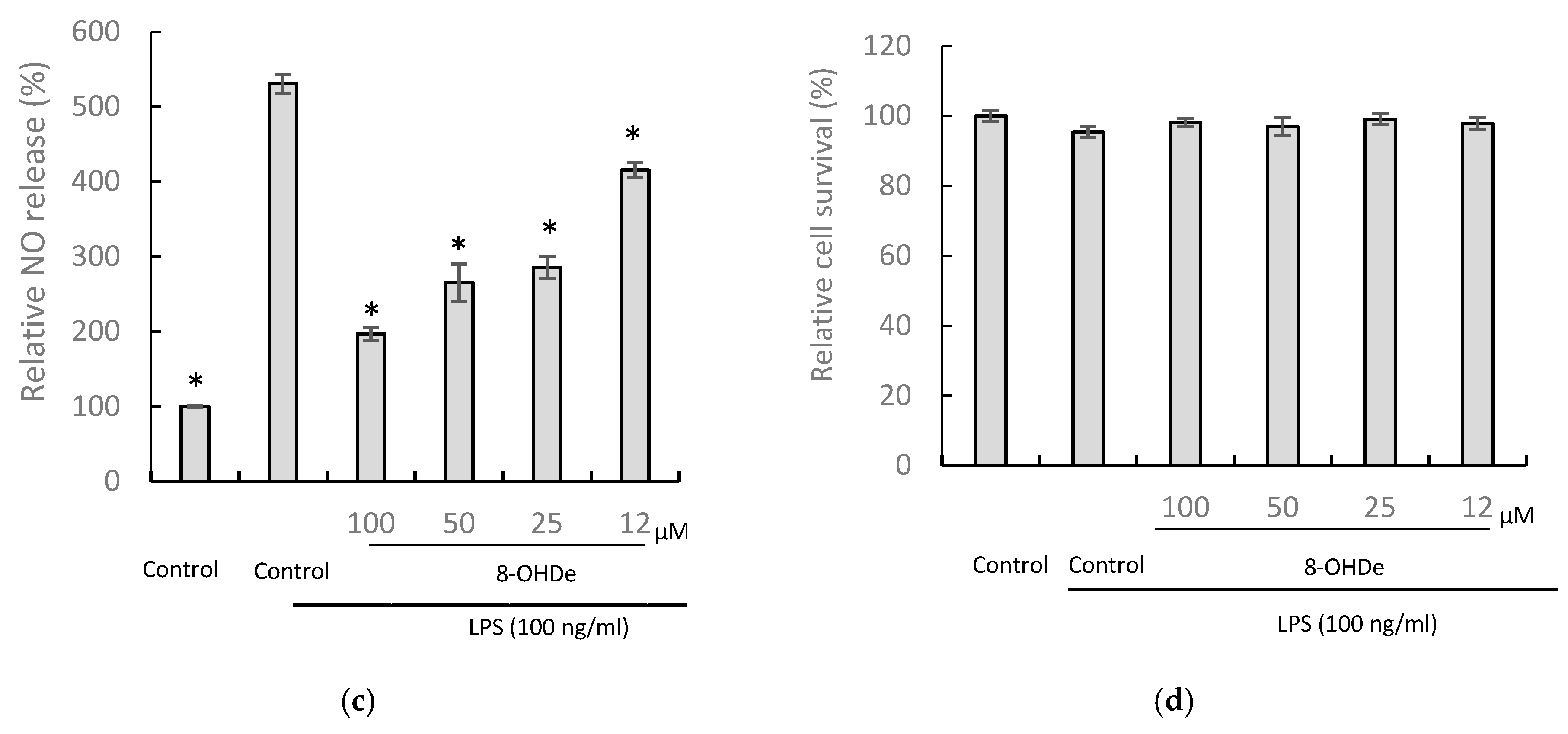
2.4. Solubility, Stability, and Anti-Inflammatory Activity of 8-OHDe 7-α-glucoside.
3. Discussion
4. Materials and Methods
4.1. Microorganisms, Animal Cells, and Chemicals
4.2. Preparation of Recombinant DgAS Enzyme
4.3. Biotransformation of 8-OHDe by the Purified DgAS Enzyme
4.4. UPLC
4.5. Purification and Identification of the Biotransformation Product
4.6. Determination of Aqueous Solubility and Stability
4.7. Determination of Anti-Inflammatory Activity
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Franke, A.A.; Custer, L.J.; Cerna, C.M.; Narala, K.K. Quantitation of phytoestrogens in legumes by HPLC. J. Agric. Food Chem. 1994, 42, 1905–1913. [Google Scholar] [CrossRef]
- Chang, T.S. Isolation, bioactivity, and production of ortho-hydroxydaidzein and ortho-hydroxygenistein. Int. J. Mol. Sci. 2014, 15, 5699–5716. [Google Scholar] [CrossRef] [PubMed]
- Komiyama, K.; Funayama, S.; Anraku, Y.; Mita, A.; Takahashi, Y.; Omura, S. Isolation of isoflavonoids possessing antioxidant activity from the fermentation broth of Streptomyces sp. J. Antibiot. 1989, 42, 1344–1349. [Google Scholar] [CrossRef]
- Kiriakidis, S.; Högemeier, O.; Starcke, S.; Dombrowski, F.; Hahne, J.C.; Pepper, M.; Jha, H.C.; Wernert, N. Novel tempeh (fermented soybean) isoflavones inhibit in vivo angiogenesis in the chicken chorioallantoic membrane assay. Br. J. Nutr. 2005, 93, 317–323. [Google Scholar] [CrossRef] [PubMed]
- Klus, K.; Barz, W. Formation of polyhydroxylated isoflavones from the soybean seed isoflavones daidzein and glycitein by bacteria isolated from tempe. Arch. Microbiol. 1995, 164, 428–434. [Google Scholar] [CrossRef] [PubMed]
- Esaki, H.; Onozaki, H.; Morimitsu, Y. Potent antioxidative isoflavones isolated from soybean fermented with Aspergillus saitoi. Biosci. Biotechnol. Biochem. 1998, 62, 740–746. [Google Scholar] [CrossRef] [PubMed]
- Esaki, H.; Kawakishi, S.; Morimitsu, Y.; Osawa, T. New potent antioxidative o-dihydroxyisoflavones in fermented Japanese soybean products. Biosci. Biotechnol. Biochem. 1999, 63, 1637–1639. [Google Scholar] [CrossRef]
- Hirota, A.; Taki, S.; Kawaii, S.; Yano, M.; Abe, N. 1,1-Diphenyl-2-picrylhydrazyl radical-scavenging compounds from soybean miso and antiproliferative activity of isoflavones from soybean miso toward the cancer cell lines. Biosci. Biotechnol. Biochem. 2000, 64, 1038–1040. [Google Scholar] [CrossRef]
- Lo, Y.L. A potential daidzein derivative enhances cytotoxicity of epirubicin on human colon adenocarcinoma Caco-2 cells. Int. J. Mol. Sci. 2012, 14, 158–176. [Google Scholar] [CrossRef]
- Chang, T.S.; Ding, H.Y.; Tai, S.S.K.; Wu, C.Y. Tyrosinase inhibitors isolated from soygerm koji fermented with Aspergillus oryzae BCRC 32288. Food Chem. 2007, 105, 1430–1438. [Google Scholar] [CrossRef]
- Chang, T.S. Two potent suicide substrates of mushroom tyrosinase: 7,8,4′-trihydroxyisoflavone and 5,7,8,4′-tetrahydroxyisoflavone. J. Agric. Food Chem. 2007, 55, 2010–2015. [Google Scholar] [CrossRef] [PubMed]
- Goh, M.J.; Park, J.S.; Bae, J.H.; Kim, D.H.; Kim, H.K.; Na, Y.J. Effects of ortho-dihydroxyisoflavone derivatives from Korean fermented soybean paste on melanogenesis in B16 melanoma cells and human skin equivalents. Phytother. Res. 2012, 26, 1107–1112. [Google Scholar] [CrossRef] [PubMed]
- Tai, S.S.K.; Lin, C.G.; Wu, M.H.; Chang, T.S. Evaluation of depigmenting activity by 8-hydroxydaidzein in mouse B16 melanoma cells and human volunteers. Int. J. Mol. Sci. 2009, 10, 4257–4266. [Google Scholar] [CrossRef] [PubMed]
- Fujita, T.; Funako, T.; Hayashi, H. 8-Hydroxydaidzein, an aldose reductase inhibitor from okara fermented with Aspergillus sp. HK-388. Biosci. Biotechnol. Biochem. 2004, 68, 1588–1590. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.S.; Ding, H.Y.; Yen, J.H.; Chen, S.F.; Lee, K.H.; Wu, M.J. Anti-inflammatory activity of 8-hydroxydaidzein in LPS-stimulated BV2 microglial cells via activation of Nrf2-antioxidant and attenuation of Akt/NF-κB-inflammatory signaling pathways, as well as inhibition of COX-2 activity. J. Agric. Food Chem. 2018, 66, 5790–5801. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.; Kang, Y.G.; Kim, J.H.; Kim, Y.J.; Lee, T.R.; Lee, J.; Kim, D.; Cho, J.Y. The antioxidant and anti-inflammatory activities of 8-hydroxydaidzein (8-HD) in activated macrophage-like RAW 264.7 cells. Int. J. Mol. Sci. 2018, 19, 1828. [Google Scholar] [CrossRef] [PubMed]
- Seo, M.H.; Kim, B.N.; Kim, K.R.; Lee, K.W.; Lee, C.H.; Oh, D.K. Production of 8-hydroxydaidzein from soybean extract by Aspergillus oryzae KACC 40247. Biosci. Biotechnol. Biochem. 2013, 77, 1245–1250. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.C.; Chang, C.W.; Lin, C.W.; Hsu, Y.C. Production of 8-hydroxydaidzein polyphenol using biotransformation by Aspergillus oryzae. Food Sci. Technol. Res. 2015, 21, 557–562. [Google Scholar] [CrossRef]
- Chang, T.S. 8-Hydroxydaidzein is unstable in alkaline solutions. J. Cosmet. Sci. 2009, 60, 353–357. [Google Scholar] [CrossRef]
- Chiang, C.M.; Wang, T.Y.; Yang, S.Y.; Wu, J.Y.; Chang, T.S. Production of new isoflavone glucosides from glycosylation of 8-hydroxydaidzein by glycosyltransferase from Bacillus subtilis ATCC 6633. Catalysts 2018, 8, 387. [Google Scholar] [CrossRef]
- Das, S.; Rosazza, J.P. Microbial and enzymatic transformation of flavonoids. J. Nat. Prod. 2006, 69, 499–508. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.; Chen, X.; Jassbi, A.R.; Xiao, J. Microbial biotransformation of bioactive flavonoids. Biotechnol. Adv. 2015, 33, 214–223. [Google Scholar] [CrossRef] [PubMed]
- Shimoda, K.; Hamada, H.; Hamada, H. Synthesis of xylooligosaccharides of daidzein and their anti-oxidant and anti-allergic activities. Int. J. Mol. Sci. 2011, 12, 5616–5625. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, P.; Sangwan, R.S.; Sangwan, N.S. Plant secondary metabolism linked glycosyltransferases: an update on expanding knowledge and scopes. Biotechnol. Adv. 2016, 34, 716–739. [Google Scholar] [CrossRef] [PubMed]
- Hofer, B. Recent developments in the enzymatic O-glycosylation of flavonoids. Appl. Microbiol. Biotechnol. 2016, 100, 4269–4281. [Google Scholar] [CrossRef]
- Claire, M.; Isabelle, A.; Magali, R.S. GH13 amylosucrases and GH70 branching sucrases, atypical enzymes in their respective families. Cell. Mol. Life Sci. 2016, 73, 2661–2679. [Google Scholar]
- Stephane, E.; Sophie, M.; Kais, J.; Isabelle, A.; Pierre, M.; Magali, R.S.; Gabrielle, P.V. Cloning, purification and characterization of a thermostable amylosucrase from Deinococcus geothermalis. FEMS Microbiol. Lett. 2008, 285, 25–32. [Google Scholar]
- Seo, D.H.; Jung, J.H.; Ha, S.J.; Cho, H.K.; Jung, D.H.; Kim, T.J.; Baek, N.I.; Yoo, S.H.; Park, C.S. High-yield enzymatic bioconversion of hydroquinone to α-arbutin, a powerful skin lightening agent, by amylosucrase. Appl. Microbiol. Biotechnol. 2012, 94, 1189–1197. [Google Scholar] [CrossRef]
- Lee, H.S.; Kim, T.S.; Parajuli, P.; Pandey, R.P.; Sohng, J.K. Sustainable production of dihydroxybenzene glucosides using immobilized amylosucrase from Deinococcus geothermalis. J. Microbiol. Biotechnol. 2018, 28, 1447–1456. [Google Scholar]
- Kim, M.D.; Jung, D.H.; Seo, D.H.; Jung, J.H.; Seo, E.J.; Baek, N.I.; Yoo, S.H.; Park, C.S. Acceptor specificity of amylosucrase from Deinococcus radiopugnans and its application for the synthesis of rutin derivatives. J. Microbiol. Biotechnol. 2016, 26, 1845–1854. [Google Scholar] [CrossRef]
- Kim, E.R.; Rha, C.S.; Jung, Y.S.; Choi, J.M.; Kim, G.T.; Jung, D.H.; Kim, T.J.; Seo, D.H.; Kim, D.O. Enzymatic modification of daidzin using heterologous expressed amylosucrase in Bacillus subtilis. Food Sci. Biotechnol. 2018, 28, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.K.; Kim, H.H.; Seo, D.H.; Jung, J.H.; Park, J.H.; Baek, N.I.; Kim, M.J.; Yoo, S.H.; Cha, J.; Kim, Y.R.; et al. Biosynthesis of catechin glycosides using recombinant amylosucrase from Deinococcus geothermalis DSM 11300. Enz. Microbial Tech. 2011, 49, 246–253. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.H.; Park, Y.D.; Park, H.; Moon, K.O.; Ha, K.T.; Baek, N.I.; Park, C.S.; Joo, M.; Cha, J. Synthesis and biological evaluation of a novel baicalein glycoside as an anti-inflammatory agent. Eur. J. Pharm. 2014, 744, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Hamalainen, M.; Nieminen, R.; Vuorela, P.; Heinonen, M.; Moilanen, E. Anti-inflammatory effects of flavonoids: genistein, kaempferol, quercetin, and daidzein inhibit STAT-1 and NF-κB activations, whereas flavone, isorhamnetin, naringenin, and pelargonidin inhibit only NF-κB activation along with their inhibitory effect on iNOS expression and NO production in activated macrophages. Med. Infla. 2007. [Google Scholar] [CrossRef]
Sample Availability: 1 mg of 8-OHDe-7-α-glucoside for each request is available from the authors. |








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Aqueous-Solubility (mg/L) | Fold |
|---|---|---|
| 8-OHDe | 47.3 | 1 |
| 8-OHDe-7-O-α-glucoside | 185.4 | 3.92 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, T.-S.; Wang, T.-Y.; Yang, S.-Y.; Kao, Y.-H.; Wu, J.-Y.; Chiang, C.-M. Potential Industrial Production of a Well-Soluble, Alkaline-Stable, and Anti-Inflammatory Isoflavone Glucoside from 8-Hydroxydaidzein Glucosylated by Recombinant Amylosucrase of Deinococcus geothermalis. Molecules 2019, 24, 2236. https://doi.org/10.3390/molecules24122236
Chang T-S, Wang T-Y, Yang S-Y, Kao Y-H, Wu J-Y, Chiang C-M. Potential Industrial Production of a Well-Soluble, Alkaline-Stable, and Anti-Inflammatory Isoflavone Glucoside from 8-Hydroxydaidzein Glucosylated by Recombinant Amylosucrase of Deinococcus geothermalis. Molecules. 2019; 24(12):2236. https://doi.org/10.3390/molecules24122236
Chicago/Turabian StyleChang, Te-Sheng, Tzi-Yuan Wang, Szu-Yi Yang, Yu-Han Kao, Jiumn-Yih Wu, and Chien-Min Chiang. 2019. "Potential Industrial Production of a Well-Soluble, Alkaline-Stable, and Anti-Inflammatory Isoflavone Glucoside from 8-Hydroxydaidzein Glucosylated by Recombinant Amylosucrase of Deinococcus geothermalis" Molecules 24, no. 12: 2236. https://doi.org/10.3390/molecules24122236