Selenium and Zinc Biofortification of Pleurotus eryngii Mycelium and Fruiting Bodies as a Tool for Controlling Their Biological Activity

 ,
, 
 and
and
Abstract
:1. Introduction
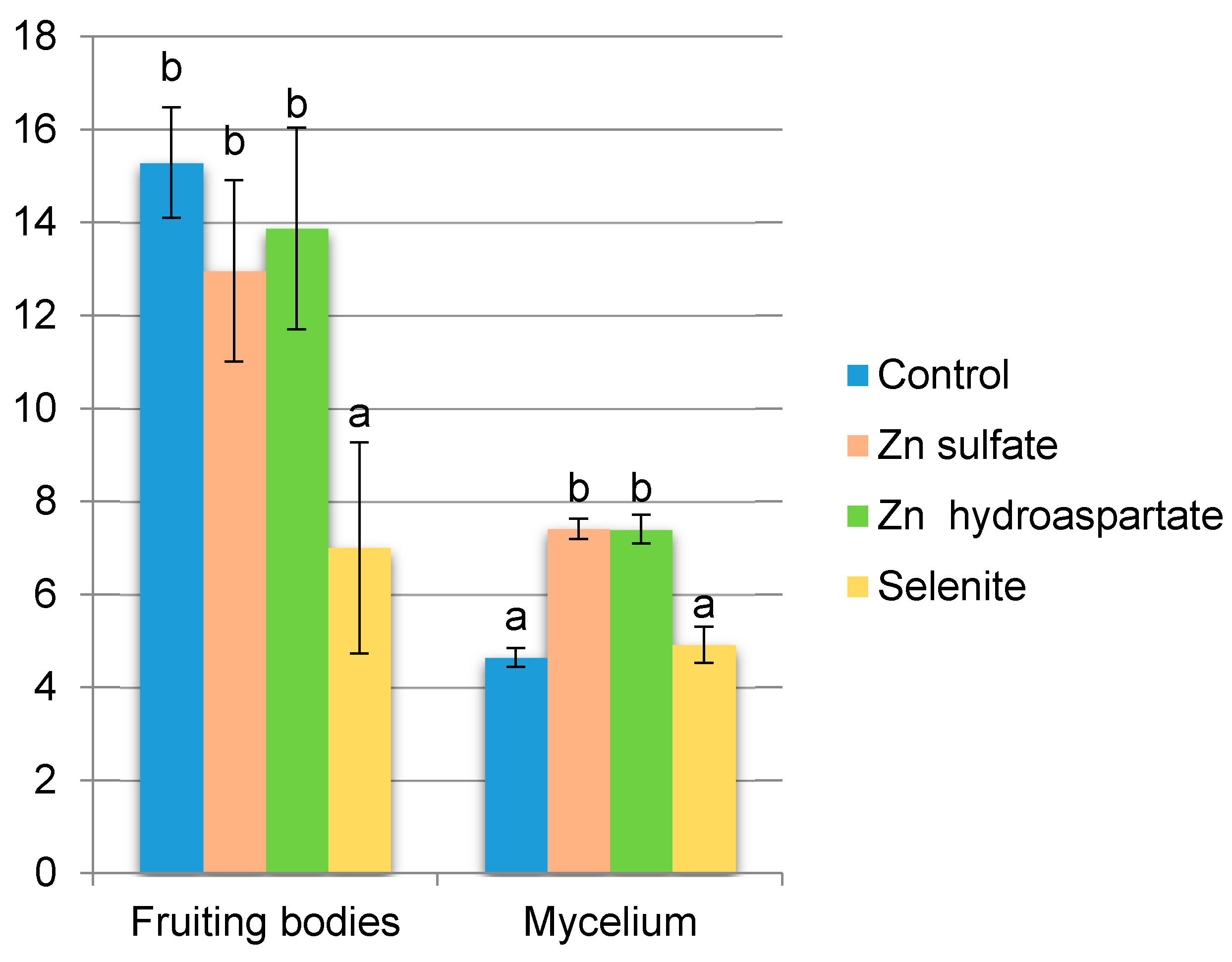
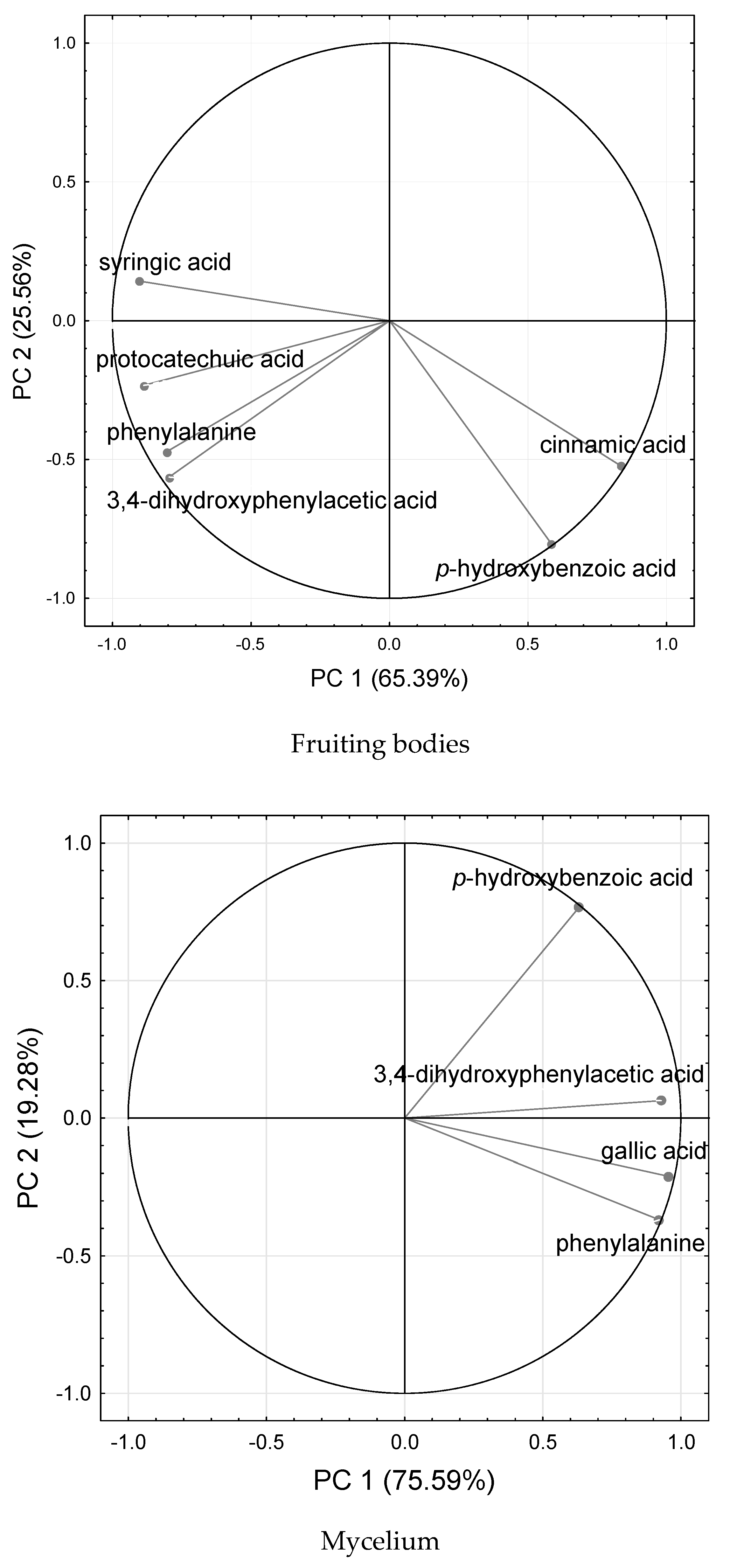
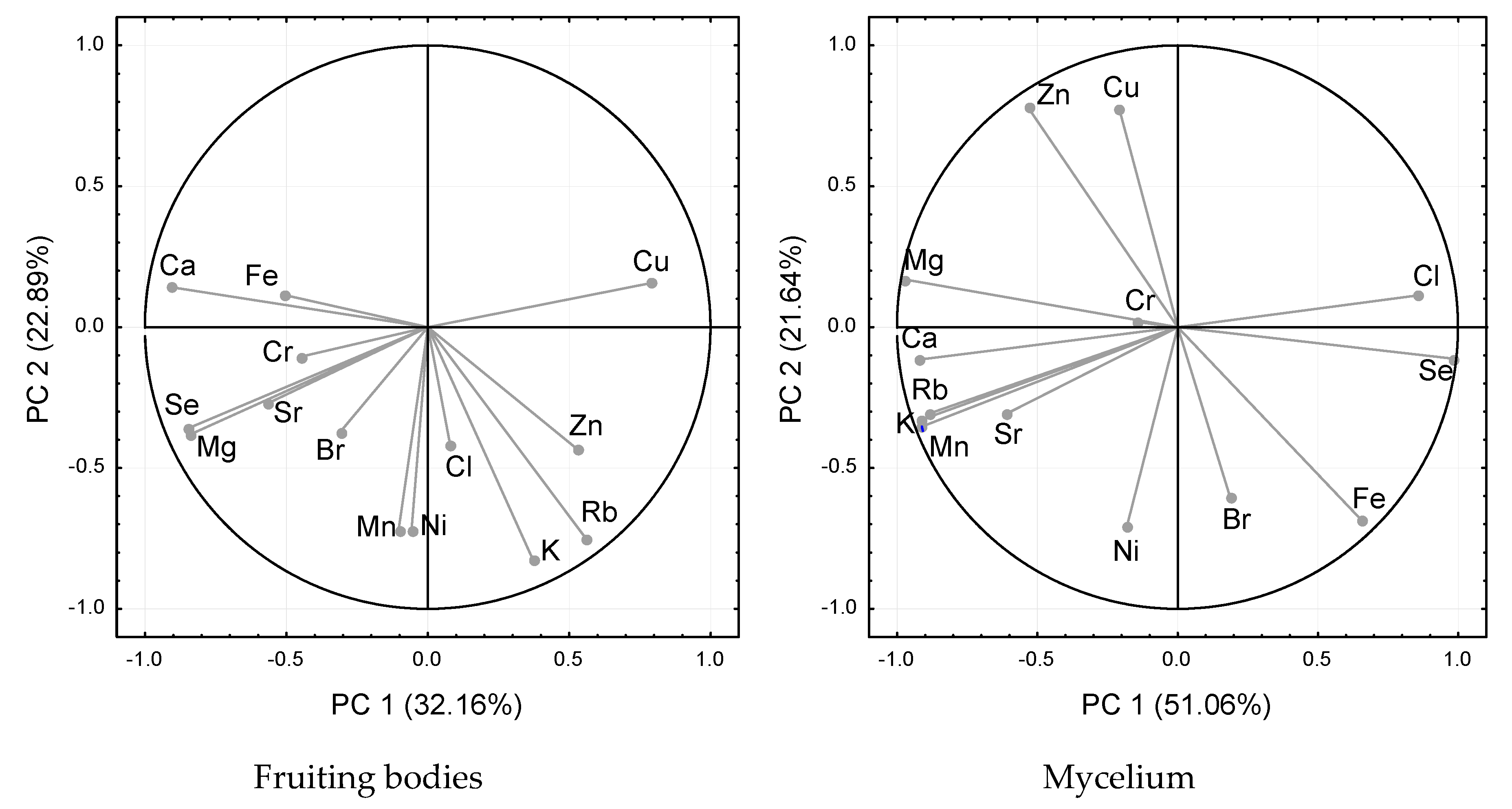
2. Results
2.1. Organic Compounds Total Phenolics and 2,2-Diphenyl-1-picrylhydrazyl ( DPPH) Scavenging Activity Analysis
2.2. Bioelements Analysis
3. Discussion
3.1. Organic Compound—Total Phenolics, Lovastatin, and DPPH˙ Scavenging Activity Analysis
3.2. Bioelements Analysis
4. Materials and Methods
4.1. Reagents and Standards
4.2. Pleurotus Eryngii Materials
4.3. In Vitro Cultures of Pleurotus eryngii
4.4. Fruiting Bodies of Pleurotus eryngii
4.5. Mushroom Extracts
4.6. Organic Compounds Analysis—Lovastatin
4.7. Organic Compounds Analysis—Phenolic Compounds
4.8. Scavenging Activity Analysis (%DPPH˙)
4.9. Total Phenol Content
4.10. Bioelements Analysis
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Vetvicka, V.; Gover, O.; Karpovsky, M.; Hayby, H.; Danay, O.; Ezov, N.; Hadar, Y.; Schwartz, B. Immune-modulating activities of glucans extracted from Pleurotus ostreatus and Pleurotus eryngii. J. Funct. Foods 2019, 54, 81–91. [Google Scholar] [CrossRef]
- Stamets, P. Growing Gourmet and Medicinal Mushrooms, 3rd ed.; Ten Speed Press: Berkeley, CA, USA, 2011; pp. 301–304. [Google Scholar]
- Akyüz, M.; Kirbag, S. Determination of culture process for obtaining basidiocarp of Pleurotus eryngii (DC. ex Fr.) Quel. var. ferulae Lanzi: A speciality mushroom. Indian J. Hortic. 2010, 67, 73–75. [Google Scholar]
- Sekara, A.; Kalisz, A.; Grabowska, A.; Siwulski, M. Auricularia spp.-mushrooms as Novel Food and therapeutic agents-a review. Sydowia 2015, 67, 1–10. [Google Scholar]
- Urbanelli, S.; Della Rosa, V.; Punelli, F.; Porretta, D.; Reverberi, M.; Fabbri, A.A.; Fanelli, C. DNA-fingerprinting (AFLP and RFLP) for genotypic identification in species of the Pleurotus eryngii complex. Appl. Microbiol. Biotechnol. 2007, 74, 592–600. [Google Scholar] [CrossRef]
- Zervakis, G.I.; Venturella, G.; Papadopoulou, K. Genetic polymorphism and taxonomic infrastructure of the Pleurotus eryngii species-complex as determined by RAPD analysis, isozyme profiles and ecomorphological characters. Microbiology 2001, 147, 3183–3194. [Google Scholar] [CrossRef] [Green Version]
- Stajić, M.; Vukojevic, J.; Duletic-Lausevic, S. Biology of Pleurotus eryngii and role in biotechnological processes: A review. Crit. Rev. Biotechnol. 2009, 29, 55–66. [Google Scholar] [CrossRef]
- Avni, S.; Ezov, N.; Hanani, H.; Yadid, I.; Karpovsky, M.; Hayby, H.; Gover, O.; Hadar, Y.; Schwartz, B.; Danay, O. Olive Mill Waste Enhances α-Glucan Content in the Edible Mushroom Pleurotus eryngii. Int. J. Mol. Sci. 2017, 18, 1564. [Google Scholar] [CrossRef]
- Carrasco, J.; Zied, D.C.; Pardo, J.E.; Preston, G.; Pardo-Giménez, A. Supplementation in mushroom crops and its impact on yield and quality. AMB Express 2018, 8, 146. [Google Scholar] [CrossRef] [Green Version]
- Cardoso, R.V.C.; Fernandes, A.; Oliveira, M.B.P.P.; Calhelha, R.C.; Barros, L.; Martins, A.; Ferreira, I.C. Development of nutraceutical formulations based on the mycelium of Pleurotus ostreatus and Agaricus bisporus. Food Funct. 2017, 8, 2155–2164. [Google Scholar] [CrossRef] [Green Version]
- Tello, I.; Montiel, E.; Romero, O.; Nava-García, E.; Leon, I. Comparative Mycelial Growth of Pleurotus djamor and Pleurotus ostreatus in Culture Media. J. Pure Appl. Microbiol. 2016, 10, 2563–2568. [Google Scholar] [CrossRef]
- Souilem, F.; Fernandes, Â.; Calhelha, R.C.; Barreira, J.; Barros, L.; Skhiri, F.; Ferreira, I.C. Bioactive compounds and anti-inflammatory activity of Pleurotus eryngii and Suillus bellinii: A comparison between fruiting bodies and mycelia. In Proceedings of the 9es Journées Scientifiques Internationales sur la Valorisation des Bioressources, Monastir, Tunisia, 3–6 May 2018. [Google Scholar]
- Nieto-López, C.; Márquez-Rocha, F.J.; Sánchez-Vazquez, J.E. Box Wilson factorial design methodology for the optimization of laccase production by Pleurotus djamor in liquid Culture. Curr. Res. Microbiol. Biotechnol. 2016, 2, 835–841. [Google Scholar]
- Niedzielski, P.; Mleczek, M.; Siwulski, M.; Rzymski, P.; Gąsecka, M.; Kozak, L. Supplementation of cultivated mushroom species with selenium: Bioaccumulation and speciation study. Eur. Food Res. Technol. 2015, 241, 419–426. [Google Scholar] [CrossRef]
- Gąsecka, M.; Mleczek, M.; Siwulski, M.; Niedzielski, P. Phenolic composition and antioxidant properties of Pleurotus ostreatus and Pleurotus eryngii enriched with selenium and zinc. Eur. Food Res. Technol. 2016, 242, 723–732. [Google Scholar] [CrossRef] [Green Version]
- Poniedziałek, B.; Mleczek, M.; Niedzielski, P.; Siwulski, M.; Gąsecka, M.; Kozak, L.; Komosa, A.; Rzymski, P. Bio-enriched Pleurotus mushrooms for deficiency control and improved antioxidative protection of human platelets? Eur. Food Res. Technol. 2017, 243, 2187–2198. [Google Scholar] [CrossRef]
- Fasoranti, O. Nutrient contents and antioxidant properties of Pleurotus spp. cultivated on substrate fortified with Selenium. Curr. Res. Environ. Appl. Mycol. 2019, 9, 66–76. [Google Scholar] [CrossRef]
- Alam, N.; Yoon, K.N.; Lee, K.R.; Kim, H.Y.; Shin, P.G.; Cheong, J.C.; Yoo, Y.B.; Shim, M.J.; Lee, M.W.; Lee, T.S. Assessment of Antioxidant and Phenolic Compound Concentrations as well as Xanthine Oxidase and Tyrosinase Inhibitory Properties of Different Extracts of Pleurotus citrinopileatus Fruiting Bodies. Mycobiology 2011, 39, 12–19. [Google Scholar] [CrossRef] [Green Version]
- Shibu, M.A.; Agrawal, D.C.; Huang, C.-Y. Mushrooms: A Pandora Box of Cardioprotective Phytochemicals. In Medicinal and Aromatic Plants of the Middle-East; Springer: Berlin/Heidelberg, Germany, 2017; Volume 4, pp. 337–362. [Google Scholar]
- Priscilla, D.H.; Prince, P.S.M. Cardioprotective effect of gallic acid on cardiac troponin-T, cardiac marker enzymes, lipid peroxidation products and antioxidants in experimentally induced myocardial infarction in Wistar rats. Chem. Interact. 2009, 179, 118–124. [Google Scholar] [CrossRef]
- Carrasco-Gonzalez, J.A.; Serna-Saldívar, S.; Gutiérrez-Uribe, J.A. Mycochemical Changes Induced by Selenium Enrichment in P. ostreatus Fruiting Bodies. J. Agric. Food Chem. 2017, 65, 4074–4082. [Google Scholar] [CrossRef]
- Abidin, M.H.Z.; Abdullah, N.; Abidin, N.Z. Therapeutic properties of Pleurotus species (Oyster mushrooms) for Atherosclerosis: A review. Int. J. Food Prop. 2017, 20, 1251–1261. [Google Scholar] [CrossRef] [Green Version]
- Van De Donk, N.W.C.J.; Kamphuis, M.M.J.; Lokhorst, H.M.; Bloem, A.C. The cholesterol lowering drug lovastatin induces cell death in myeloma plasma cells. Leukemia 2002, 16, 1362–1371. [Google Scholar] [CrossRef] [Green Version]
- Golak-Siwulska, I.; Kałuzewicz, A.; Spiżewski, T.; Siwulski, M.; Sobieralski, K. Bioactive compounds and medicinal properties of Oyster mushrooms (Pleurotus sp.). Folia Hortic. 2018, 30, 191–201. [Google Scholar] [CrossRef] [Green Version]
- Xu, N.; Ren, Z.; Zhang, J.; Song, X.; Gao, Z.; Jing, H.; Li, S.; Wang, S.; Jia, L. Antioxidant and anti-hyperlipidemic effects of mycelia zinc polysaccharides by Pleurotus eryngii var. tuoliensis. Int. J. Boil. Macromol. 2017, 95, 204–214. [Google Scholar] [CrossRef] [PubMed]
- Svoboda, L.; Zimmermannová, K.; Kalač, P. Concentrations of mercury, cadmium, lead and copper in fruiting bodies of edible mushrooms in an emission area of a copper smelter and a mercury smelter. Sci. Total. Environ. 2000, 246, 61–67. [Google Scholar] [CrossRef]
- Brown, K.M.; Arthur, J.R. Selenium, selenoproteins and human health: A review. Public Health Nutr. 2001, 4, 593–599. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, P.R.; Berry, M.J. The influence of selenium on immune responses. Mol. Nutr. Food Res. 2008, 52, 1273–1280. [Google Scholar] [CrossRef]
- Greenough, M.A.; Volitakis, I.; Li, Q.-X.; Laughton, K.; Evin, G.; Ho, M.; Dalziel, A.H.; Camakaris, J.; Bush, I.A. Presenilins Promote the Cellular Uptake of Copper and Zinc and Maintain Copper Chaperone of SOD1-dependent Copper/Zinc Superoxide Dismutase Activity. J. Boil. Chem. 2011, 286, 9776–9786. [Google Scholar] [CrossRef] [Green Version]
- Plum, L.M.; Rink, L.; Haase, H. The Essential Toxin: Impact of Zinc on Human Health. Int. J. Environ. Res. Public Health 2010, 7, 1342–1365. [Google Scholar] [CrossRef] [Green Version]
- Yan, H.; Chang, H. Antioxidant and Antitumor Activities of Selenium- and Zinc-Enriched Oyster Mushroom in Mice. Boil. Trace Elem. Res. 2012, 150, 236–241. [Google Scholar] [CrossRef]
- Silva, M.D.C.S.D.; Da Luz, J.M.R.; Paiva, A.P.S.; Mendes, D.R.; Carvalho, A.A.C.; Naozuka, J.; Kasuya, M.C.M. Growth and Tolerance of Pleurotus ostreatus at Different Selenium Forms. J. Agric. Sci. 2019, 11, 151–158. [Google Scholar] [CrossRef]
- Muñoz, A.H.S.; Kubachka, K.; Wrobel, K.; Corona, J.F.G.; Yathavakilla, S.K.V.; Caruso, J.A.; Wrobel, K. Se-Enriched Mycelia ofPleurotus ostreatus: Distribution of Selenium in Cell Walls and Cell Membranes/Cytosol. J. Agric. Food Chem. 2006, 54, 3440–3444. [Google Scholar] [CrossRef]
- Kim, M.-Y.; Seguin, P.; Ahn, J.-K.; Kim, J.-J.; Chun, S.C.; Kim, E.-H.; Seo, S.-H.; Kang, E.-Y.; Kim, S.-L.; Park, Y.-J.; et al. Phenolic Compound Concentration and Antioxidant Activities of Edible and Medicinal Mushrooms from Korea. J. Agric. Food Chem. 2008, 56, 7265–7270. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Shah, N.P. Effects of Various Heat Treatments on Phenolic Profiles and Antioxidant Activities of Pleurotus eryngii Extracts. J. Food Sci. 2013, 78, 1122–1129. [Google Scholar] [CrossRef] [PubMed]
- Chirinang, P.; Intarapichet, K.O. Amino acids and antioxidant properties of the oyster mushrooms, Pleurotus ostreatus and Pleurotus sajor-caju. Sci. Asia 2009, 35, 326–331. [Google Scholar] [CrossRef]
- Reis, F.S.; Barros, L.; Martins, A.; Ferreira, I.C. Chemical composition and nutritional value of the most widely appreciated cultivated mushrooms: An inter-species comparative study. Food Chem. Toxicol. 2012, 50, 191–197. [Google Scholar] [CrossRef] [Green Version]
- Atli, B.; Yamaç, M. Screening of medicinal higher Basidiomycetes mushrooms from Turkey for lovastatin production. Int. J. Med. Mushrooms 2012, 14, 149–159. [Google Scholar] [CrossRef]
- Zhang, Q.; Chen, L.; Guo, K.; Zheng, L.; Liu, B.; Yu, W.; Guo, C.; Liu, Z.; Chen, Y.; Tang, Z. Effects of Different Selenium Levels on Gene Expression of a Subset of Selenoproteins and Antioxidative Capacity in Mice. Boil. Trace Elem. Res. 2013, 154, 255–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.-H.; Lee, H.-S.; Kwon, H.-J.; Patnaik, B.B.; Nam, K.-W.; Han, Y.S.; Bang, I.-S.; Han, M.-D. Effects of different selenium levels on growth and regulation of laccase and versatile peroxidase in white-rot fungus, Pleurotus eryngii. World J. Microbiol. Biotechnol. 2014, 30, 2101–2109. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, B.; Zhang, D.; Wu, Y. Selenium uptake, tolerance and reduction in Flammulina velutipes supplied with selenite. PeerJ 2016, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaur, G.; Kalia, A.; Sodhi, H.S. Selenium biofortification of Pleurotus species and its effect on yield, phytochemical profiles, and protein chemistry of fruiting bodies. J. Food Biochem. 2018, 42. [Google Scholar] [CrossRef]
- Stajic, M.; Brceski, I.; Vukojević, J.; Duletic-Lausevic, S. Ability of selenium absorption by mycelia of Pleurotus eryngii (DC.: Fr.) Quel., depending on selenium source in medium. Int. J. Med. Mushrooms 2005, 7, 467–468. [Google Scholar]
- Gogavekar, S.S.; Rokade, S.A.; Ranveer, R.C.; Ghosh, J.S.; Kalyani, D.C.; Sahoo, A.K. Important nutritional constituents, flavour components, antioxidant and antibacterial properties of Pleurotus sajor-caju. J. Food Sci. Tech. 2014, 51, 1483–1491. [Google Scholar] [CrossRef] [Green Version]
- Mattila, P.; Salo-Väänänen, P.; Könkö, K.; Aro, H.; Jalava, T. Basic Composition and Amino Acid Contents of Mushrooms Cultivated in Finland. J. Agric. Food Chem. 2002, 50, 6419–6422. [Google Scholar] [CrossRef]
- Poursaeid, N.; Azadbakht, A.; Balali, G.R. Improvement of Zinc Bioaccumulation and Biomass Yield in the Mycelia and Fruiting Bodies of Pleurotus florida Cultured on Liquid Media. Appl. Biochem. Biotechnol. 2015, 175, 3387–3396. [Google Scholar] [CrossRef]
- Vieira, P.A.F.; Gontijo, D.C.; Vieira, B.C.; Fontes, E.A.; De Assunção, L.S.; Leite, J.; Oliveira, M.G.D.A.; Kasuya, M.C.M. Antioxidant activities, total phenolics and metal contents in Pleurotus ostreatus mushrooms enriched with iron, zinc or lithium. LWT 2013, 54, 421–425. [Google Scholar] [CrossRef]
- Kalač, P. A review of trace element concentrations in edible mushrooms. Food Chem. 2000, 69, 273–281. [Google Scholar] [CrossRef]
- Siwulski, M.; Budzyńska, S.; Rzymski, P.; Gąsecka, M.; Niedzielski, P.; Kalač, P.; Mleczek, M. The effects of germanium and selenium on growth, metalloid accumulation and ergosterol content in mushrooms: Experimental study in Pleurotus ostreatus and Ganoderma lucidum. Eur. Food Res. Technol. 2019, 245, 1799–1810. [Google Scholar] [CrossRef] [Green Version]
- U.S. Food and Drug Administration. Available online: http://www.fda.gov/ (accessed on 28 September 2012).
- Lavelli, V.; Proserpio, C.; Gallotti, F.; Laureati, M.; Pagliarini, E. Circular reuse of bio-resources: The role of Pleurotus spp. in the development of functional foods. Food Funct. 2018, 9, 1353–1372. [Google Scholar] [CrossRef] [Green Version]
- Estrada, A.E.R.; Lee, H.-J.; Beelman, R.B.; Jimenez-Gasco, M.D.M.; Royse, D.J. Enhancement of the antioxidants ergothioneine and selenium in Pleurotus eryngii var. eryngii basidiomata through cultural practices. World J. Microbiol. Biotechnol. 2009, 25, 1597–1607. [Google Scholar] [CrossRef]
- Kała, K.; Krakowska, A.; Sułkowska-Ziaja, K.; Szewczyk, A.; Reczyński, W.; Opoka, W.; Muszyńska, B. Kinetics of extracted bioactive components from mushrooms in artificial digestive juices. Int. J. Food Prop. 2017, 20, 1796–1817. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Treatment | Phenylalanine | Gallic Acid | Protocatechuic Acid | 3,4-Dihydroxy Phenylacetic Acid | p-Hydroxybenzoic Acid | Syringic Acid | Cinnamic Acid | Lovastatin | Total Phenolics | DPPH˙ Scavenging Activity |
|---|---|---|---|---|---|---|---|---|---|---|
| Fruiting Bodies | ||||||||||
| Control | 176 ± 0.85 d1 | n.d. | 0.42 ± 0.01 b | 99.30 ± 0.38 d | 2.44 ± 0.03 b | 0.09 ± 0.01 b | 0.92 ± 0.01 b | n.d. | 212.76 ± 14.04 a,b | 33.15 ± 1.03 a |
| Selenite | 141 ± 0.69 c | n.d. | 0.40 ± 0.02 b | 97.01 ± 0.27 c | 2.35 ± 0.03 a | 0.09 ± 0.01 b | 0.94 ± 0.07 a | n.d. | 226.44 ± 25.03 b | 39.22 ± 0.47 b |
| Zn sulfate | 137 ± 1.28 b | n.d. | 0.36 ± 0.02 a | 92.11 ± 0.89 b | 3.34 ± 0.03 c | 0.06 ± 0.01 a | 1.61 ± 0.05 d | n.d. | 263.04 ± 10.29 c | 41.56 ± 0.75 b |
| Zn hydroaspartate | 121 ± 0.46 a | n.d. | 0.36 ± 0.01 a | 84.20 ± 0.29 a | 2.31 ± 0.02 a | 0.07 ± 0.01 a | 0.82 ± 0.01 a | n.d. | 183.72 ± 14.04 a | 33.03 ± 2.21 a |
| Mycelium | ||||||||||
| Control | 257 ± 10.4 a | 0.58 ± 0.09 a | n.d. | 6.75 ± 0.10 a | 0.26 ± 0.02 b | n.d. | n.d. | 27.02 ± 0.07 d | 283.68 ± 12.96 a | 54.6 ± 0.25 b |
| Selenite | 622 ± 13.4 b | 1.14 ± 0.09 b | n.d. | n.d | 0.17 ± 0.01 a | n.d. | n.d. | 2.26 ± 0.09 c | 257.16 ± 3.11 a | 56.5 ± 1.53 b |
| Zn sulfate | 2233 ± 88.9 d | 10.42 ± 0.22 d | n.d. | 15.78 ± 0.26 c | 0.25 ± 0.02 b | n.d. | n.d. | 0.11 ± 0.03 a | 318.06 ± 3.42 a | 39.9 ± 1.37 a |
| Zn hydroaspartate | 1531 ± 45.1 c | 8.87 ± 0.26 c | n.d. | 8.34 ± 0.07 b | 0.28 ± 0.05 b | n.d. | n.d. | 0.69 ± 0.04 b | 317.52 ± 3.81 a | 41.6 ± 0.15 a |
| Organic Compounds | Fruiting Bodies | Mycelium | ||
|---|---|---|---|---|
| Zn | Se | Zn | Se | |
| Phenylalanine | 0.301 * | −0.045 | 0.766 ** | −0.479 |
| Gallic acid | nd | nd | 0.890 *** | −0.612 * |
| Protocatechuic acid | −0.123 | 0.357 | nd | nd |
| 3,4-Dihydroxyphenylacetic acid | 0.223 | 0.397 | 0.575 * | −0.824 *** |
| p-Hydroxybenzoic acid | 0.772 *** | −0.349 | 0.564 * | −0.842 *** |
| Syringic acid | −0.273 | 0.526 * | nd | nd |
| Cinnamic acid | 0.567 * | −0.322 | nd | nd |
| Lovastatin | nd | nd | −0.578 * | −0.176 |
| Total phenolics | −0.738 *** | 0.0896 | 0.195 | −0.513 * |
| 2,2-Diphenyl-1-picrylhydrazyl (DPPH) scavenging activity | −0.708 ** | 0.3555 | −0.957 *** | 0.711 *** |
| Treatment | K | Mg | Ca | Zn | Fe | Cl | Rb | Cu | Se | Mn | Ni | Cr | Br | Sr |
| Growth Substrate | ||||||||||||||
| 449 ± 19 | 49.8 ± 1.6 | 153.6 ± 2.3 | 3.43 ± 0.08 | 13.6 ± 0.46 | 1.22 ± 0.33 | 0.33 ± 0.04 | 0.27 ± 0.08 | 0.02 ± 0.02 | 4.27 ± 0.11 | 1.24 ± 0.15 | 1.40 ± 0.25 | 0.14 ± 0.06 | 0.84 ± 0.06 | |
| Fruiting Bodies | ||||||||||||||
| Control | 2328 ± 235 c1 | 117 ± 7 a,b | 17.4 ± 1.6 a | 4.94 ± 0.12 b | 3.85 ± 0.22 a | 1.98 ± 0.36 a | 1.64 ± 0.13 c | 1.42 ± 0.06 c | 0.05 ± 0.05 a | 0.90 ± 0.04 a | 0.49 ± 0.05 b | 0.12 ± 0.02 a | 0.10 ± 0.03 a | 0.08 ± 0.02 a |
| Selenite | 2070 ± 129 b,c | 235 ± 11 c | 52.8 ± 4.7 d | 4.48 ± 0.35 a | 5.11 ± 0.97 b | 1.89 ± 0.39 a | 1.34 ± 0.06 a,b | 0.92 ± 0.03 a | 1.36 ± 0.08 b | 0.92 ± 0.08 a | 0.50 ± 0.11 b | 0.16 ± 0.07 a | 0.12 ± 0.06 a | 0.14 ± 0.09 a |
| Zn sulfate | 1753 ± 240 a | 110 ± 2 a | 40.1 ± 0.3 c | 4.39 ± 0.29 a | 4.98 ± 0.37 b | 1.85 ± 0.29 a | 1.24 ± 0.14 a | 1.23 ± 0.23 b | 0.03 ± 0.03 a | 0.86 ± 0.11 a | 0.38 ± 0.05 a | 0.13 ± 0.03 a | 0.08 ± 0.03 a | 0.07 ± 0.02 a |
| Zn hydroaspartate | 2034 ± 122 b | 121 ± 3 b | 30.5 ± 6.7 b | 5.45 ± 0.25 c | 5.56 ± 1.06 b | 1.86 ± 0.18 a | 1.44 ± 0.08 b | 1.14 ± 0.09 b | 0.03 ± 0.03 a | 0.87 ± 0.04 a | 0.43 ± 0.06 b | 0.11 ± 0.04 a | 0.13 ± 0.03 a | 0.09 ± 0.03 a |
| Mycelium | ||||||||||||||
| Control | 872 ± 62 c | 175 ± 7.1 b | 61.8 ± 7.1 b | 15.1 ± 0.88 b | 31.4 ± 0.36 c | 17.46 ± 4.75 a,b | 0.29 ± 0.05 c | 0.29 ± 0.07 a | 2.1 ± 0.03 b | 9.98 ± 0.41 d | 0.63 ± 0.13 b | 0.14 ± 0.06 a | 0.26 ± 0.09 a | 0.20 ± 0.00 a |
| Selenite | 432 ± 9 a | 79 ± 1.5 a | 29.1 ± 6.5 a | 10.9 ± 0.54 a | 33.46 ± 0.88 d | 31.64 ± 2.62 c | 0.13 ± 0.03 a | 0.35 ± 0.07 a,b | 18.79 ± 0.44 b | 4.46 ± 0.17 a | 0.46 ± 0.05 a | 0.14 ± 0.02 a | 0.24 ± 0.04 a | 0.14 ± 0.05 a |
| Zn sulfate | 739 ± 46 b | 192 ± 8.2 c | 54.7 ± 5.9 b | 289.5 ± 1.80 d | 22.81 ± 0.74 b | 21.95 ± 5.50 b | 0.24 ± 0.03 b | 0.46 ± 0.06 c | 0.23 ± 0.08 a | 7.82 ± 0.13 b | 0.45 ± 0.08 a | 0.17 ± 0.07 a | 0.19 ± 0.02 a | 0.18 ± 0.03 a |
| Zn hydroaspartate | 729 ± 29 b | 193 ± 0.4 c | 61.8 ± 6.7 b | 178.3 ± 2.43 d | 20.74 ± 1.28 a | 16.09 ± 1.46 a | 0.25 ± 0.00 b,c | 0.41 ± 0.03 b,c | 0.36 ± 0.05 a | 8.48 ± 0.21 c | 0.41 ± 0.07 a | 0.12 ± 0.06 a | 0.19 ± 0.02 a | 0.18 ± 0.03 a |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zięba, P.; Kała, K.; Włodarczyk, A.; Szewczyk, A.; Kunicki, E.; Sękara, A.; Muszyńska, B. Selenium and Zinc Biofortification of Pleurotus eryngii Mycelium and Fruiting Bodies as a Tool for Controlling Their Biological Activity. Molecules 2020, 25, 889. https://doi.org/10.3390/molecules25040889
Zięba P, Kała K, Włodarczyk A, Szewczyk A, Kunicki E, Sękara A, Muszyńska B. Selenium and Zinc Biofortification of Pleurotus eryngii Mycelium and Fruiting Bodies as a Tool for Controlling Their Biological Activity. Molecules. 2020; 25(4):889. https://doi.org/10.3390/molecules25040889
Chicago/Turabian StyleZięba, Piotr, Katarzyna Kała, Anna Włodarczyk, Agnieszka Szewczyk, Edward Kunicki, Agnieszka Sękara, and Bożena Muszyńska. 2020. "Selenium and Zinc Biofortification of Pleurotus eryngii Mycelium and Fruiting Bodies as a Tool for Controlling Their Biological Activity" Molecules 25, no. 4: 889. https://doi.org/10.3390/molecules25040889