Antimicrobial Peptides as Anticancer Agents: Functional Properties and Biological Activities

 ,
,  ,
, 
Abstract
:
1. Introduction
2. The Classification of Antimicrobial Peptides
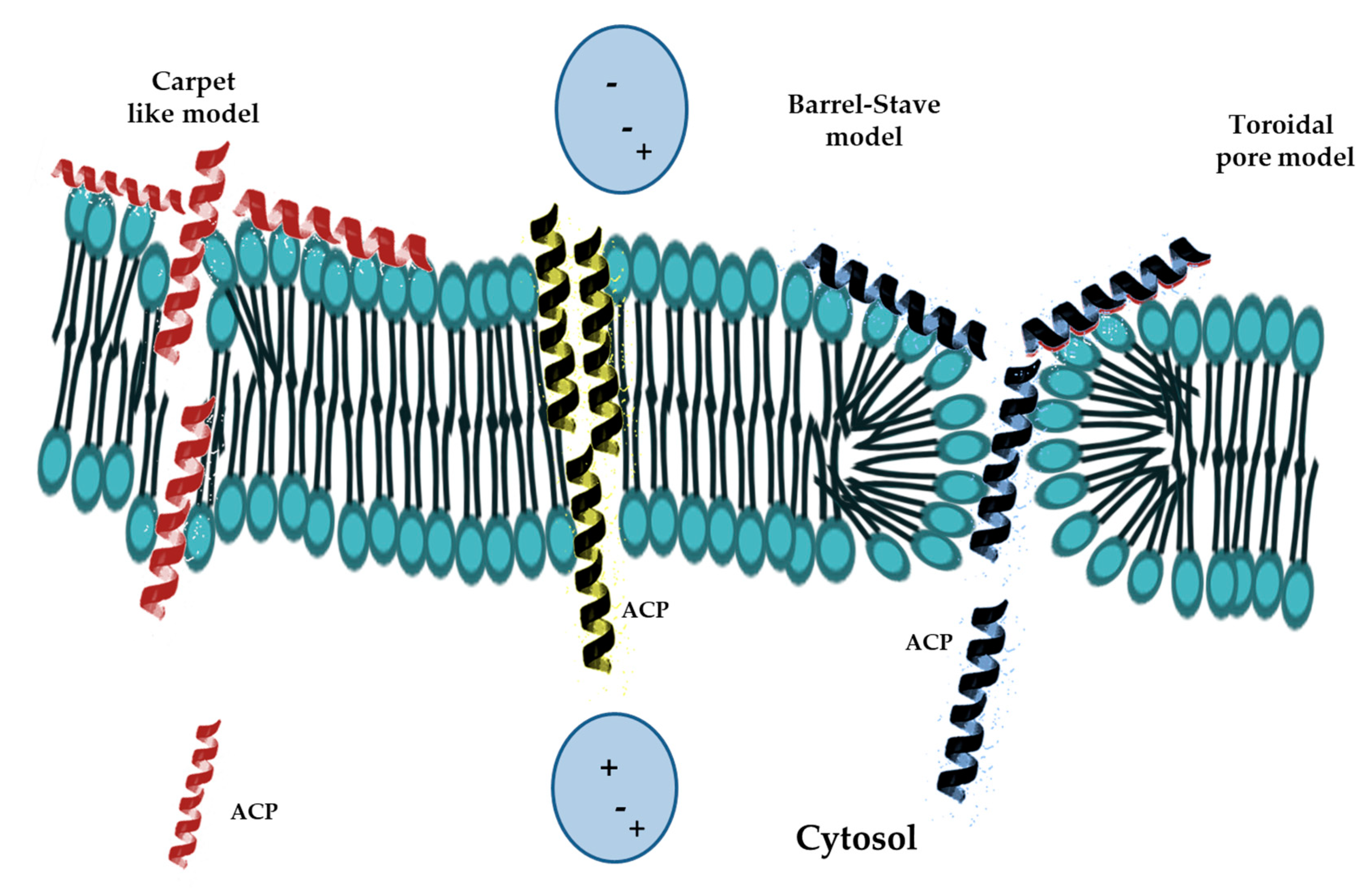
Mode of Action of Antimicrobial Peptides
3. Critical Issues, Possible Solutions for Clinical Use of AMPs
3.1. Antimicrobial Peptides as Anticancer Drugs
3.2. Examples of ACPs
3.2.1. Cathelicidins
3.2.2. Human Defensins
3.2.3. Bovine Lactoferricin (LfcinB)
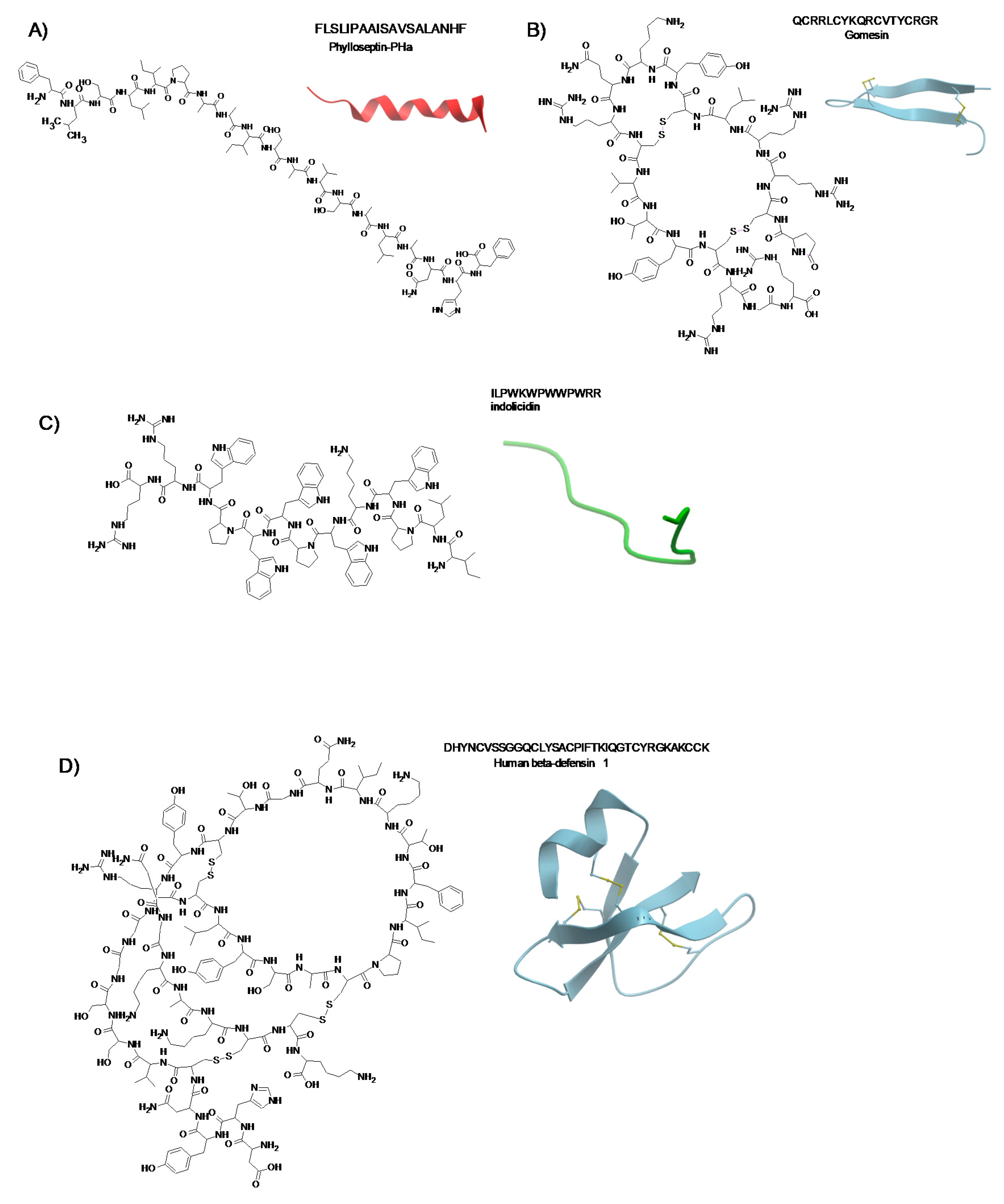
3.2.4. Gomesin
3.2.5. Mastoparan-C
3.2.6. Cecropin B1
3.2.7. Magainin 2
3.2.8. Buforin IIb
3.2.9. Brevinin-2R
3.2.10. Limnonectes fujianensis Brevinvin
3.2.11. Phylloseptin-PHa
3.2.12. Ranatuerin-2PLx
3.2.13. Dermaseptins
3.2.14. Chrysophsins
3.2.15. Ss-arasin
3.2.16. Turgencin A and Turgencin B
3.2.17. D-K6L9
3.2.18. KLA
3.2.19. Dusquetide
3.3. ACP for the Treatment of Hepatocellular Carcinoma
3.3.1. Tv1
3.3.2. SALL4 Peptide FFW
3.3.3. SP94
3.3.4. R-Tf-D-LP4
3.3.5. GG-8-6
3.3.6. β3 Peptide
3.3.7. CecropinXJ
3.3.8. GW-H1
3.3.9. Bombinin
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Raheem, N.; Straus, S.K. Mechanisms of action for antimicrobial peptides with antibacterial and antibiofilm functions. Front. Microbiol. 2019, 10, 2866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fleming, A.; Allison, V.D. Observations on a bacteriolytic substance (“lysozyme”) found in secretions and tissues. Br. J. Exp. Pathol. 1992, 3, 252–260. [Google Scholar]
- Dubos, R.J. Studies on a bactericidal agent extracted from a soil bacillus: I. Preparation of the agent. Its activity In Vitro. J. Exp. Med. 1939, 70, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubos, R.J. Studies on a bactericidal agent extracted from a soil bacillus: Ii. Protective effect of the bactericidal agent against experimental pneumococcus infections in mice. J. Exp. Med. 1939, 70, 11–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hotchkiss, R.D.; Dubos, R.J. Fractionation of the bactericidal agent from cultures of a soil bacillus. J. Biol. Chem. 1940, 132, 791–792. [Google Scholar]
- Van Epps, H.L. Rene dubos: Unearthing antibiotics. J. Exp. Med. 2006, 203, 259. [Google Scholar] [CrossRef] [Green Version]
- Hirsch, J.G. Phagocytin: A bactericidal substance from polymorphonuclear leucocytes. J. Exp. Med. 1956, 103, 589–611. [Google Scholar] [CrossRef]
- Zeya, H.I.; Spitznagel, J.K. Antibacterial and enzymic basic proteins from leukocyte lysosomes: Separation and identification. Science 1963, 142, 1085–1087. [Google Scholar] [CrossRef]
- Ganz, T.; Selsted, M.E.; Szklarek, D.; Harwig, S.S.; Daher, K.; Bainton, D.F.; Lehrer, R.I. Defensins. Natural peptide antibiotics of human neutrophils. J. Clin. Investig. 1985, 76, 1427–1435. [Google Scholar] [CrossRef]
- Guani-Guerra, E.; Santos-Mendoza, T.; Lugo-Reyes, S.O.; Teran, L.M. Antimicrobial peptides: General overview and clinical implications in human health and disease. Clin. Immunol. 2010, 135, 1–11. [Google Scholar] [CrossRef]
- Lehrer, R.I. Primate defensins. Nat. Rev. Microbiol. 2004, 2, 727–738. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Li, X.; Wang, Z. Apd3: The antimicrobial peptide database as a tool for research and education. Nucleic Acids Res. 2016, 44, D1087–D1093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waghu, F.H.; Barai, R.S.; Gurung, P.; Idicula-Thomas, S. Campr3: A database on sequences, structures and signatures of antimicrobial peptides. Nucleic Acids Res. 2016, 44, D1094–D1097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, X.; Dong, F.; Shi, C.; Liu, S.; Sun, J.; Chen, J.; Li, H.; Xu, H.; Lao, X.; Zheng, H. Dramp 2.0, an updated data repository of antimicrobial peptides. Sci. Data 2019, 6, 148. [Google Scholar] [CrossRef] [Green Version]
- Seebah, S.; Suresh, A.; Zhuo, S.; Choong, Y.H.; Chua, H.; Chuon, D.; Beuerman, R.; Verma, C. Defensins knowledgebase: A manually curated database and information source focused on the defensins family of antimicrobial peptides. Nucleic Acids Res. 2007, 35, D265–D268. [Google Scholar] [CrossRef] [Green Version]
- Qureshi, A.; Thakur, N.; Tandon, H.; Kumar, M. Avpdb: A database of experimentally validated antiviral peptides targeting medically important viruses. Nucleic Acids Res. 2014, 42, D1147–D1153. [Google Scholar] [CrossRef] [Green Version]
- Mehta, D.; Anand, P.; Kumar, V.; Joshi, A.; Mathur, D.; Singh, S.; Tuknait, A.; Chaudhary, K.; Gautam, S.K.; Gautam, A.; et al. Parapep: A web resource for experimentally validated antiparasitic peptide sequences and their structures. Database (Oxf.) 2014, 2014. [Google Scholar] [CrossRef]
- Tyagi, A.; Tuknait, A.; Anand, P.; Gupta, S.; Sharma, M.; Mathur, D.; Joshi, A.; Singh, S.; Gautam, A.; Raghava, G.P. Cancerppd: A database of anticancer peptides and proteins. Nucleic Acids Res. 2015, 43, D837–D843. [Google Scholar] [CrossRef] [Green Version]
- Bobone, S.; Stella, L. Selectivity of antimicrobial peptides: A complex interplay of multiple equilibria. Adv. Exp. Med. Biol. 2019, 1117, 175–214. [Google Scholar]
- Savini, F.; Loffredo, M.R.; Troiano, C.; Bobone, S.; Malanovic, N.; Eichmann, T.O.; Caprio, L.; Canale, V.C.; Park, Y.; Mangoni, M.L.; et al. Binding of an antimicrobial peptide to bacterial cells: Interaction with different species, strains and cellular components. Biochim. Biophys. Acta Biomembr. 2020, 1862, 183291. [Google Scholar] [CrossRef]
- Hancock, R.E.; Lehrer, R. Cationic peptides: A new source of antibiotics. Trends Biotechnol. 1998, 16, 82–88. [Google Scholar] [CrossRef]
- Shai, Y. Mechanism of the binding, insertion and destabilization of phospholipid bilayer membranes by alpha-helical antimicrobial and cell non-selective membrane-lytic peptides. Biochim. Biophys. Acta 1999, 1462, 55–70. [Google Scholar] [CrossRef] [Green Version]
- Travkova, O.G.; Moehwald, H.; Brezesinski, G. The interaction of antimicrobial peptides with membranes. Adv. Colloid Interface Sci. 2017, 247, 521–532. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, A.; Siman-Tov, G.; Hall, G.; Bhalla, N.; Narayanan, A. Human antimicrobial peptides as therapeutics for viral infections. Viruses 2019, 11, 704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, D.; Lu, W. Defensins: A double-edged sword in host immunity. Front. Immunol. 2020, 11, 764. [Google Scholar] [CrossRef] [PubMed]
- Wilson, S.S.; Wiens, M.E.; Holly, M.K.; Smith, J.G. Defensins at the mucosal surface: Latest insights into defensin-virus interactions. J. Virol. 2016, 90, 5216–5218. [Google Scholar] [CrossRef] [Green Version]
- Lafferty, M.K.; Sun, L.; Christensen-Quick, A.; Lu, W.; Garzino-Demo, A. Human beta defensin 2 selectively inhibits hiv-1 in highly permissive ccr6(+)cd4(+) t cells. Viruses 2017, 9, 111. [Google Scholar] [CrossRef] [Green Version]
- Wodlej, C.; Riedl, S.; Rinner, B.; Leber, R.; Drechsler, C.; Voelker, D.R.; Choi, J.Y.; Lohner, K.; Zweytick, D. Interaction of two antitumor peptides with membrane lipids—Influence of phosphatidylserine and cholesterol on specificity for melanoma cells. PLoS ONE 2019, 14, e0211187. [Google Scholar] [CrossRef]
- Bevers, E.M.; Comfurius, P.; Zwaal, R.F. Regulatory mechanisms in maintenance and modulation of transmembrane lipid asymmetry: Pathophysiological implications. Lupus 1996, 5, 480–487. [Google Scholar] [CrossRef]
- Ran, S.; Downes, A.; Thorpe, P.E. Increased exposure of anionic phospholipids on the surface of tumor blood vessels. Cancer Res. 2002, 62, 6132–6140. [Google Scholar]
- Beloribi-Djefaflia, S.; Vasseur, S.; Guillaumond, F. Lipid metabolic reprogramming in cancer cells. Oncogenesis 2016, 5, e189. [Google Scholar] [CrossRef] [PubMed]
- Scocchi, M.; Mardirossian, M.; Runti, G.; Benincasa, M. Non-Membrane permeabilizing modes of action of antimicrobial peptides on bacteria. Curr. Top. Med. Chem. 2016, 16, 76–88. [Google Scholar] [CrossRef] [PubMed]
- Lei, J.; Sun, L.; Huang, S.; Zhu, C.; Li, P.; He, J.; Mackey, V.; Coy, D.H.; He, Q. The antimicrobial peptides and their potential clinical applications. Am. J. Transl. Res. 2019, 11, 3919–3931. [Google Scholar] [PubMed]
- Gkeka, P.; Sarkisov, L. Interactions of phospholipid bilayers with several classes of amphiphilic alpha-helical peptides: Insights from coarse-grained molecular dynamics simulations. J. Phys. Chem. B 2010, 114, 826–839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Housman, G.; Byler, S.; Heerboth, S.; Lapinska, K.; Longacre, M.; Snyder, N.; Sarkar, S. Drug resistance in cancer: An overview. Cancers 2014, 6, 1769–1792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoskin, D.W.; Ramamoorthy, A. Studies on anticancer activities of antimicrobial peptides. Biochim. Biophys. Acta 2008, 1778, 357–375. [Google Scholar] [CrossRef] [Green Version]
- Hicks, R.P. Antibacterial and anticancer activity of a series of novel peptides incorporating cyclic tetra-substituted c(α) amino acids. Bioorg. Med. Chem. 2016, 24, 4056–4065. [Google Scholar] [CrossRef]
- Kunda, N.K. Antimicrobial peptides as novel therapeutics for non-small cell lung cancer. Drug Discov. Today 2020, 25, 238–247. [Google Scholar] [CrossRef]
- Giuliani, A. Antimicrobial peptides: An overview of a promising class of therapeutics. Cent. Eur. J. Biol. 2007, 2, 1–33. [Google Scholar] [CrossRef]
- Zhang, C.; Yang, M.; Ericsson, A.C. Antimicrobial peptides: Potential application in liver cancer. Front. Microbiol. 2019, 10, 1257. [Google Scholar] [CrossRef]
- Diamond, G.; Beckloff, N.; Weinberg, A.; Kisich, K.O. The roles of antimicrobial peptides in innate host defense. Curr. Pharm. Des. 2009, 15, 2377–2392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piotrowska, U.; Sobczak, M.; Oledzka, E. Current state of a dual behaviour of antimicrobial peptides-therapeutic agents and promising delivery vectors. Chem. Biol. Drug Des. 2017, 90, 1079–1093. [Google Scholar] [CrossRef] [PubMed]
- Ramesh, S.; Cherkupally, P.; Govender, T.; Kruger, H.G.; Albericio, F.; de la Torre, B.G. Highly chemoselective ligation of thiol- and amino-peptides on a bromomaleimide core. Chem. Commun. 2016, 52, 2334–2337. [Google Scholar] [CrossRef] [PubMed]
- Roudi, R.; Syn, N.L.; Roudbary, M. Antimicrobial peptides as biologic and immunotherapeutic agents against cancer: A comprehensive overview. Front. Immunol. 2017, 8, 1320. [Google Scholar] [CrossRef] [Green Version]
- Costa, F.; Carvalho, I.F.; Montelaro, R.C.; Gomes, P.; Martins, M.C. Covalent immobilization of antimicrobial peptides (amps) onto biomaterial surfaces. Acta Biomater. 2011, 7, 1431–1440. [Google Scholar] [CrossRef] [Green Version]
- Bahar, A.A.; Ren, D. Antimicrobial peptides. Pharmaceuticals 2013, 6, 1543–1575. [Google Scholar] [CrossRef] [Green Version]
- Baxter, A.A.; Lay, F.T.; Poon, I.K.H.; Kvansakul, M.; Hulett, M.D. Tumor cell membrane-targeting cationic antimicrobial peptides: Novel insights into mechanisms of action and therapeutic prospects. Cell. Mol. Life Sci. 2017, 74, 3809–3825. [Google Scholar] [CrossRef]
- Wang, G. Improved methods for classification, prediction, and design of antimicrobial peptides. Methods Mol. Biol. 2015, 1268, 43–66. [Google Scholar]
- Falanga, A.; Galdiero, M.; Morelli, G.; Galdiero, S. Membranotropic peptides mediating viral entry. Pept. Sci. 2018, 110, e24040. [Google Scholar] [CrossRef]
- Sitaram, N.; Nagaraj, R. Interaction of antimicrobial peptides with biological and model membranes: Structural and charge requirements for activity. Biochim. Biophys. Acta 1999, 1462, 29–54. [Google Scholar] [CrossRef] [Green Version]
- Ebenhan, T.; Gheysens, O.; Kruger, H.G.; Zeevaart, J.R.; Sathekge, M.M. Antimicrobial peptides: Their role as infection-selective tracers for molecular imaging. BioMed Res. Int. 2014, 2014, 867381. [Google Scholar] [CrossRef] [PubMed]
- Ruan, Y.; Shen, T.; Wang, Y.; Hou, M.; Li, J.; Sun, T. Antimicrobial peptide ll-37 attenuates lta induced inflammatory effect in macrophages. Int. Immunopharmacol. 2013, 15, 575–580. [Google Scholar] [CrossRef] [PubMed]
- Shang, D.; Zhang, Q.; Dong, W.; Liang, H.; Bi, X. The effects of lps on the activity of trp-containing antimicrobial peptides against gram-negative bacteria and endotoxin neutralization. Acta Biomater. 2016, 33, 153–165. [Google Scholar] [CrossRef]
- Deslouches, B.; Di, Y.P. Antimicrobial peptides with selective antitumor mechanisms: Prospect for anticancer applications. Oncotarget 2017, 8, 46635–46651. [Google Scholar] [CrossRef] [Green Version]
- Hancock, R.E. Peptide antibiotics. Lancet 1997, 349, 418–422. [Google Scholar] [CrossRef]
- Borrelli, A.; Tornesello, A.L.; Tornesello, M.L.; Buonaguro, F.M. Cell penetrating peptides as molecular carriers for anti-cancer agents. Molecules 2018, 23, 295. [Google Scholar] [CrossRef] [Green Version]
- Last, N.B.; Schlamadinger, D.E.; Miranker, A.D. A common landscape for membrane-active peptides. Protein Sci. 2013, 22, 870–882. [Google Scholar] [CrossRef] [Green Version]
- Hancock, R.E.; Chapple, D.S. Peptide antibiotics. Antimicrob. Agents Chemother. 1999, 43, 1317–1323. [Google Scholar] [CrossRef] [Green Version]
- Teixeira, V.; Feio, M.J.; Bastos, M. Role of lipids in the interaction of antimicrobial peptides with membranes. Prog. Lipid Res. 2012, 51, 149–177. [Google Scholar] [CrossRef]
- Haney, E.F.; Nathoo, S.; Vogel, H.J.; Prenner, E.J. Induction of non-lamellar lipid phases by antimicrobial peptides: A potential link to mode of action. Chem. Phys. Lipids 2010, 163, 82–93. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Chau, J.K.; Perry, N.A.; de Boer, L.; Zaat, S.A.; Vogel, H.J. Serum stabilities of short tryptophan- and arginine-rich antimicrobial peptide analogs. PLoS ONE 2010, 5, e12684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uggerhoj, L.E.; Poulsen, T.J.; Munk, J.K.; Fredborg, M.; Sondergaard, T.E.; Frimodt-Moller, N.; Hansen, P.R.; Wimmer, R. Rational design of α-helical antimicrobial peptides: Do’s and don’ts. ChemBioChem 2015, 16, 242–253. [Google Scholar] [CrossRef] [PubMed]
- Braunstein, A.; Papo, N.; Shai, Y. In Vitro activity and potency of an intravenously injected antimicrobial peptide and its dl amino acid analog in mice infected with bacteria. Antimicrob. Agents Chemother. 2004, 48, 3127–3129. [Google Scholar] [CrossRef] [Green Version]
- Jia, F.; Wang, J.; Peng, J.; Zhao, P.; Kong, Z.; Wang, K.; Yan, W.; Wang, R. D-Amino acid substitution enhances the stability of antimicrobial peptide polybia-cp. Acta Biochim. Biophys. Sin. 2017, 49, 916–925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelly, G.J.; Kia, A.F.; Hassan, F.; O’Grady, S.; Morgan, M.P.; Creaven, B.S.; McClean, S.; Harmey, J.H.; Devocelle, M. Polymeric prodrug combination to exploit the therapeutic potential of antimicrobial peptides against cancer cells. Org. Biomol. Chem. 2016, 14, 9278–9286. [Google Scholar] [CrossRef] [PubMed]
- Spinks, C.B.; Zidan, A.S.; Khan, M.A.; Habib, M.J.; Faustino, P.J. Pharmaceutical characterization of novel tenofovir liposomal formulations for enhanced oral drug delivery: In Vitro pharmaceutics and caco-2 permeability investigations. Clin. Pharmacol. 2017, 9, 29–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iachetta, G.; Falanga, A.; Molino, Y.; Masse, M.; Jabes, F.; Mechioukhi, Y.; Laforgia, V.; Khrestchatisky, M.; Galdiero, S.; Valiante, S. Gh625-Liposomes as tool for pituitary adenylate cyclase-activating polypeptide brain delivery. Sci. Rep. 2019, 9, 9183. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.H.; Lan, C.H.; Wu, K.L.; Wu, Y.M.; Jane, W.N.; Hsiao, M.; Wu, H.C. Hepatocellular carcinoma-targeted nanoparticles for cancer therapy. Int. J. Oncol. 2018, 52, 389–401. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Yuan, D.; Sun, M.; Ping, Q. Effect of ligand density and peg modification on octreotide-targeted liposome via somatostatin receptor in vitro and In Vivo. Drug Deliv. 2016, 23, 3562–3572. [Google Scholar] [CrossRef] [Green Version]
- Domalaon, R.; Findlay, B.; Ogunsina, M.; Arthur, G.; Schweizer, F. Ultrashort cationic lipopeptides and lipopeptoids: Evaluation and mechanistic insights against epithelial cancer cells. Peptides 2016, 84, 58–67. [Google Scholar] [CrossRef]
- Dabrowska, K.; Kazmierczak, Z.; Majewska, J.; Miernikiewicz, P.; Piotrowicz, A.; Wietrzyk, J.; Lecion, D.; Hodyra, K.; Nasulewicz-Goldeman, A.; Owczarek, B.; et al. Bacteriophages displaying anticancer peptides in combined antibacterial and anticancer treatment. Future Microbiol. 2014, 9, 861–869. [Google Scholar] [CrossRef] [PubMed]
- Hao, X.; Yan, Q.; Zhao, J.; Wang, W.; Huang, Y.; Chen, Y. Tat modification of α-helical anticancer peptides to improve specificity and efficacy. PLoS ONE 2015, 10, e0138911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tornesello, A.L.; Tagliamonte, M.; Tornesello, M.L.; Buonaguro, F.M.; Buonaguro, L. Nanoparticles to improve the efficacy of peptide-based cancer vaccines. Cancers 2020, 12, 1049. [Google Scholar] [CrossRef] [PubMed]
- Pazos, E.; Sleep, E.; Rubert Perez, C.M.; Lee, S.S.; Tantakitti, F.; Stupp, S.I. Nucleation and growth of ordered arrays of silver nanoparticles on peptide nanofibers: Hybrid nanostructures with antimicrobial properties. J. Am. Chem. Soc. 2016, 138, 5507–5510. [Google Scholar] [CrossRef]
- Makowski, M.; Silva, I.C.; Pais do Amaral, C.; Goncalves, S.; Santos, N.C. Advances in lipid and metal nanoparticles for antimicrobial peptide delivery. Pharmaceutics 2019, 11, 588. [Google Scholar] [CrossRef] [Green Version]
- Mao, J.; Liu, S.; Ai, M.; Wang, Z.; Wang, D.; Li, X.; Hu, K.; Gao, X.; Yang, Y. A novel melittin nano-liposome exerted excellent anti-hepatocellular carcinoma efficacy with better biological safety. J. Hematol. Oncol. 2017, 10, 71. [Google Scholar] [CrossRef] [Green Version]
- Wang, A.Z.; Langer, R.; Farokhzad, O.C. Nanoparticle delivery of cancer drugs. Annu. Rev. Med. 2012, 63, 185–198. [Google Scholar] [CrossRef]
- Banerjee, K.; Ravishankar Rai, V.; Umashankar, M. Effect of peptide-conjugated nanoparticles on cell lines. Prog. Biomater. 2019, 8, 11–21. [Google Scholar] [CrossRef] [Green Version]
- Farkhani, S.M.; Valizadeh, A.; Karami, H.; Mohammadi, S.; Sohrabi, N.; Badrzadeh, F. Cell penetrating peptides: Efficient vectors for delivery of nanoparticles, nanocarriers, therapeutic and diagnostic molecules. Peptides 2014, 57, 78–94. [Google Scholar] [CrossRef]
- Niemirowicz, K.; Prokop, I.; Wilczewska, A.Z.; Wnorowska, U.; Piktel, E.; Watek, M.; Savage, P.B.; Bucki, R. Magnetic nanoparticles enhance the anticancer activity of cathelicidin ll-37 peptide against colon cancer cells. Int. J. Nanomed. 2015, 10, 3843–3853. [Google Scholar] [CrossRef] [Green Version]
- Piktel, E.; Prokop, I.; Wnorowska, U.; Krol, G.; Ciesluk, M.; Niemirowicz, K.; Savage, P.B.; Bucki, R. Ceragenin csa-13 as free molecules and attached to magnetic nanoparticle surfaces induce caspase-dependent apoptosis in human breast cancer cells via disruption of cell oxidative balance. Oncotarget 2018, 9, 21904–21920. [Google Scholar] [CrossRef] [PubMed]
- Lombardi, L.; Falanga, A.; Del Genio, V.; Galdiero, S. A new hope: Self-Assembling peptides with antimicrobial activity. Pharmaceutics 2019, 11, 166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Felicio, M.R.; Silva, O.N.; Goncalves, S.; Santos, N.C.; Franco, O.L. Peptides with dual antimicrobial and anticancer activities. Front. Chem. 2017, 5, 5. [Google Scholar] [CrossRef] [Green Version]
- Chuang, C.M.; Monie, A.; Wu, A.; Mao, C.P.; Hung, C.F. Treatment with ll-37 peptide enhances antitumor effects induced by cpg oligodeoxynucleotides against ovarian cancer. Hum. Gene Ther. 2009, 20, 303–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okumura, K.; Itoh, A.; Isogai, E.; Hirose, K.; Hosokawa, Y.; Abiko, Y.; Shibata, T.; Hirata, M.; Isogai, H. C-Terminal domain of human cap18 antimicrobial peptide induces apoptosis in oral squamous cell carcinoma sas-h1 cells. Cancer Lett. 2004, 212, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Mader, J.S.; Mookherjee, N.; Hancock, R.E.; Bleackley, R.C. The human host defense peptide ll-37 induces apoptosis in a calpain- and apoptosis-inducing factor-dependent manner involving bax activity. Mol. Cancer Res. 2009, 7, 689–702. [Google Scholar] [CrossRef] [Green Version]
- Xhindoli, D.; Pacor, S.; Guida, F.; Antcheva, N.; Tossi, A. Native oligomerization determines the mode of action and biological activities of human cathelicidin ll-37. Biochem. J. 2014, 457, 263–275. [Google Scholar] [CrossRef]
- Cheng, M.; Ho, S.; Yoo, J.H.; Tran, D.H.; Bakirtzi, K.; Su, B.; Tran, D.H.; Kubota, Y.; Ichikawa, R.; Koon, H.W. Cathelicidin suppresses colon cancer development by inhibition of cancer associated fibroblasts. Clin. Exp. Gastroenterol. 2015, 8, 13–29. [Google Scholar]
- Xu, N.; Wang, Y.S.; Pan, W.B.; Xiao, B.; Wen, Y.J.; Chen, X.C.; Chen, L.J.; Deng, H.X.; You, J.; Kan, B.; et al. Human α-defensin-1 inhibits growth of human lung adenocarcinoma xenograft in nude mice. Mol. Cancer Ther. 2008, 7, 1588–1597. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.S.; Li, D.; Shi, H.S.; Wen, Y.J.; Yang, L.; Xu, N.; Chen, X.C.; Chen, X.; Chen, P.; Li, J.; et al. Intratumoral expression of mature human neutrophil peptide-1 mediates antitumor immunity in mice. Clin. Cancer Res. 2009, 15, 6901–6911. [Google Scholar] [CrossRef] [Green Version]
- Phan, T.K.; Lay, F.T.; Poon, I.K.; Hinds, M.G.; Kvansakul, M.; Hulett, M.D. Human β-defensin 3 contains an oncolytic motif that binds pi(4,5)p2 to mediate tumour cell permeabilisation. Oncotarget 2016, 7, 2054–2069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eliassen, L.T.; Berge, G.; Leknessund, A.; Wikman, M.; Lindin, I.; Lokke, C.; Ponthan, F.; Johnsen, J.I.; Sveinbjornsson, B.; Kogner, P.; et al. The antimicrobial peptide, lactoferricin b, is cytotoxic to neuroblastoma cells In Vitro and inhibits xenograft growth In Vivo. Int. J. Cancer 2006, 119, 493–500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furlong, S.J.; Ridgway, N.D.; Hoskin, D.W. Modulation of ceramide metabolism in t-leukemia cell lines potentiates apoptosis induced by the cationic antimicrobial peptide bovine lactoferricin. Int. J. Oncol. 2008, 32, 537–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domingues, T.M.; Riske, K.A.; Miranda, A. Revealing the lytic mechanism of the antimicrobial peptide gomesin by observing giant unilamellar vesicles. Langmuir 2010, 26, 11077–11084. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, L.; Wu, Y.; Wang, L.; Ma, C.; Xi, X.; Bininda-Emonds, O.R.P.; Shaw, C.; Chen, T.; Zhou, M. Evaluation of the bioactivity of a mastoparan peptide from wasp venom and of its analogues designed through targeted engineering. Int. J. Biol. Sci. 2018, 14, 599–607. [Google Scholar] [CrossRef]
- Wu, J.M.; Jan, P.S.; Yu, H.C.; Haung, H.Y.; Fang, H.J.; Chang, Y.I.; Cheng, J.W.; Chen, H.M. Structure and function of a custom anticancer peptide, cb1a. Peptides 2009, 30, 839–848. [Google Scholar] [CrossRef]
- Li, X.; Shen, B.; Chen, Q.; Zhang, X.; Ye, Y.; Wang, F.; Zhang, X. Antitumor effects of cecropin b-lhrh’ on drug-resistant ovarian and endometrial cancer cells. BMC Cancer 2016, 16, 251. [Google Scholar] [CrossRef] [Green Version]
- Baker, M.A.; Maloy, W.L.; Zasloff, M.; Jacob, L.S. Anticancer efficacy of magainin2 and analogue peptides. Cancer Res. 1993, 53, 3052–3057. [Google Scholar]
- Lehmann, J.; Retz, M.; Sidhu, S.S.; Suttmann, H.; Sell, M.; Paulsen, F.; Harder, J.; Unteregger, G.; Stockle, M. Antitumor activity of the antimicrobial peptide magainin ii against bladder cancer cell lines. Eur. Urol. 2006, 50, 141–147. [Google Scholar] [CrossRef]
- Shin, S.Y.; Kang, J.H.; Jang, S.Y.; Kim, Y.; Kim, K.L.; Hahm, K.S. Effects of the hinge region of cecropin a(1-8)-magainin 2(1-12), a synthetic antimicrobial peptide, on liposomes, bacterial and tumor cells. Biochim. Biophys. Acta 2000, 1463, 209–218. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.S.; Park, C.B.; Kim, J.M.; Jang, S.A.; Park, I.Y.; Kim, M.S.; Cho, J.H.; Kim, S.C. Mechanism of anticancer activity of buforin iib, a histone h2a-derived peptide. Cancer Lett. 2008, 271, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Ghavami, S.; Asoodeh, A.; Klonisch, T.; Halayko, A.J.; Kadkhoda, K.; Kroczak, T.J.; Gibson, S.B.; Booy, E.P.; Naderi-Manesh, H.; Los, M. Brevinin-2r(1) semi-selectively kills cancer cells by a distinct mechanism, which involves the lysosomal-mitochondrial death pathway. J. Cell. Mol. Med. 2008, 12, 1005–1022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Lyu, P.; Xie, S.; Qin, H.; Pu, W.; Xu, H.; Chen, T.; Shaw, C.; Ge, L.; Kwok, H.F. Lfb: A novel antimicrobial brevinin-like peptide from the skin secretion of the fujian large headed frog, limnonectes fujianensi. Biomolecules 2019, 9, 242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Du, Q.; Ma, C.; Xi, X.; Wang, L.; Zhou, M.; Burrows, J.F.; Chen, T.; Wang, H. Structure-Activity relationship of an antimicrobial peptide, phylloseptin-pha: Balance of hydrophobicity and charge determines the selectivity of bioactivities. Drug Des. Dev. Ther. 2019, 13, 447–458. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Zhang, L.; Ma, C.; Zhang, Y.; Xi, X.; Wang, L.; Zhou, M.; Burrows, J.F.; Chen, T. A novel antimicrobial peptide, ranatuerin-2plx, showing therapeutic potential in inhibiting proliferation of cancer cells. Biosci. Rep. 2018, 38. [Google Scholar] [CrossRef] [Green Version]
- Bartels, E.J.H.; Dekker, D.; Amiche, M. Dermaseptins, multifunctional antimicrobial peptides: A review of their pharmacology, effectivity, mechanism of action, and possible future directions. Front. Pharmacol. 2019, 10, 1421. [Google Scholar] [CrossRef] [Green Version]
- Dos Santos, C.; Hamadat, S.; Le Saux, K.; Newton, C.; Mazouni, M.; Zargarian, L.; Miro-Padovani, M.; Zadigue, P.; Delbe, J.; Hamma-Kourbali, Y.; et al. Studies of the antitumor mechanism of action of dermaseptin b2, a multifunctional cationic antimicrobial peptide, reveal a partial implication of cell surface glycosaminoglycans. PLoS ONE 2017, 12, e0182926. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Xi, X.; Ma, C.; Chen, X.; Zhou, M.; Burrows, J.F.; Chen, T.; Wang, L. A novel dermaseptin isolated from the skin secretion of phyllomedusa tarsius and its cationicity-enhanced analogue exhibiting effective antimicrobial and anti-proliferative activities. Biomolecules 2019, 9, 628. [Google Scholar] [CrossRef] [Green Version]
- Hsu, J.C.; Lin, L.C.; Tzen, J.T.; Chen, J.Y. Characteristics of the antitumor activities in tumor cells and modulation of the inflammatory response in raw264.7 cells of a novel antimicrobial peptide, chrysophsin-1, from the red sea bream (Chrysophrys major). Peptides 2011, 32, 900–910. [Google Scholar] [CrossRef]
- Anju, A.; Smitha, C.K.; Preetha, K.; Boobal, R.; Rosamma, P. Molecular characterization, recombinant expression and bioactivity profile of an antimicrobial peptide, ss-arasin from the indian mud crab, Scylla serrata. Fish Shellfish Immunol. 2019, 88, 352–358. [Google Scholar] [CrossRef]
- Hansen, I.K.O.; Isaksson, J.; Poth, A.G.; Hansen, K.O.; Andersen, A.J.C.; Richard, C.S.M.; Blencke, H.M.; Stensvag, K.; Craik, D.J.; Haug, T. Isolation and characterization of antimicrobial peptides with unusual disulfide connectivity from the colonial ascidian synoicum turgens. Mar. Drugs 2020, 18, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papo, N.; Seger, D.; Makovitzki, A.; Kalchenko, V.; Eshhar, Z.; Degani, H.; Shai, Y. Inhibition of tumor growth and elimination of multiple metastases in human prostate and breast xenografts by systemic inoculation of a host defense-like lytic peptide. Cancer Res. 2006, 66, 5371–5378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mai, J.C.; Mi, Z.; Kim, S.H.; Ng, B.; Robbins, P.D. A proapoptotic peptide for the treatment of solid tumors. Cancer Res. 2001, 61, 7709–7712. [Google Scholar] [PubMed]
- Haug, B.E.; Camilio, K.A.; Eliassen, L.T.; Stensen, W.; Svendsen, J.S.; Berg, K.; Mortensen, B.; Serin, G.; Mirjolet, J.F.; Bichat, F.; et al. Discovery of a 9-mer cationic peptide (ltx-315) as a potential first in class oncolytic peptide. J. Med. Chem. 2016, 59, 2918–2927. [Google Scholar] [CrossRef] [Green Version]
- Sveinbjornsson, B.; Camilio, K.A.; Haug, B.E.; Rekdal, O. Ltx-315: A first-in-class oncolytic peptide that reprograms the tumor microenvironment. Future Med. Chem. 2017, 9, 1339–1344. [Google Scholar] [CrossRef] [Green Version]
- Heulot, M.; Jacquier, N.; Aeby, S.; Le Roy, D.; Roger, T.; Trofimenko, E.; Barras, D.; Greub, G.; Widmann, C. The anticancer peptide tat-rasgap317-326 exerts broad antimicrobial activity. Front. Microbiol. 2017, 8, 994. [Google Scholar] [CrossRef]
- Raman, K.; Kuberan, B. Chemical tumor biology of heparan sulfate proteoglycans. Curr. Chem. Biol. 2010, 4, 20–31. [Google Scholar]
- Chan, S.C.; Hui, L.; Chen, H.M. Enhancement of the cytolytic effect of anti-bacterial cecropin by the microvilli of cancer cells. Anticancer Res. 1998, 18, 4467–4474. [Google Scholar]
- Jin, G.; Weinberg, A. Human antimicrobial peptides and cancer. Semin. Cell Dev. Biol. 2019, 88, 156–162. [Google Scholar] [CrossRef]
- Niyonsaba, F.; Iwabuchi, K.; Someya, A.; Hirata, M.; Matsuda, H.; Ogawa, H.; Nagaoka, I. A cathelicidin family of human antibacterial peptide ll-37 induces mast cell chemotaxis. Immunology 2002, 106, 20–26. [Google Scholar] [CrossRef]
- Carretero, M.; Escamez, M.J.; Garcia, M.; Duarte, B.; Holguin, A.; Retamosa, L.; Jorcano, J.L.; Rio, M.D.; Larcher, F. In Vitro and In Vivo wound healing-promoting activities of human cathelicidin ll-37. J. Investig. Dermatol. 2008, 128, 223–236. [Google Scholar] [CrossRef] [Green Version]
- Xhindoli, D.; Pacor, S.; Benincasa, M.; Scocchi, M.; Gennaro, R.; Tossi, A. The human cathelicidin ll-37—A pore-forming antibacterial peptide and host-cell modulator. Biochim. Biophys. Acta 2016, 1858, 546–566. [Google Scholar] [CrossRef]
- Ren, S.X.; Cheng, A.S.; To, K.F.; Tong, J.H.; Li, M.S.; Shen, J.; Wong, C.C.; Zhang, L.; Chan, R.L.; Wang, X.J.; et al. Host immune defense peptide ll-37 activates caspase-independent apoptosis and suppresses colon cancer. Cancer Res. 2012, 72, 6512–6523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szyk, A.; Wu, Z.; Tucker, K.; Yang, D.; Lu, W.; Lubkowski, J. Crystal structures of human α-defensins hnp4, hd5, and hd6. Protein Sci. 2006, 15, 2749–2760. [Google Scholar] [CrossRef] [PubMed]
- Chavakis, T.; Cines, D.B.; Rhee, J.S.; Liang, O.D.; Schubert, U.; Hammes, H.P.; Higazi, A.A.; Nawroth, P.P.; Preissner, K.T.; Bdeir, K. Regulation of neovascularization by human neutrophil peptides (α-defensins): A link between inflammation and angiogenesis. FASEB J. 2004, 18, 1306–1308. [Google Scholar] [CrossRef] [PubMed]
- Bellamy, W.; Takase, M.; Yamauchi, K.; Wakabayashi, H.; Kawase, K.; Tomita, M. Identification of the bactericidal domain of lactoferrin. Biochim. Biophys. Acta 1992, 1121, 130–136. [Google Scholar] [CrossRef]
- Frank, M.G.; Weber, M.D.; Watkins, L.R.; Maier, S.F. Stress sounds the alarmin: The role of the danger-associated molecular pattern hmgb1 in stress-induced neuroinflammatory priming. Brain Behav. Immun. 2015, 48, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanner, J.D.; Deplazes, E.; Mancera, R.L. The biological and biophysical properties of the spider peptide gomesin. Molecules 2018, 23, 1733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaspar, D.; Veiga, A.S.; Castanho, M.A. From antimicrobial to anticancer peptides. A review. Front. Microbiol. 2013, 4, 294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Azevedo, R.A.; Figueiredo, C.R.; Ferreira, A.K.; Matsuo, A.L.; Massaoka, M.H.; Girola, N.; Auada, A.V.; Farias, C.F.; Pasqualoto, K.F.; Rodrigues, C.P.; et al. Mastoparan induces apoptosis in b16f10-nex2 melanoma cells via the intrinsic mitochondrial pathway and displays antitumor activity In Vivo. Peptides 2015, 68, 113–119. [Google Scholar] [CrossRef]
- Bessalle, R.; Kapitkovsky, A.; Gorea, A.; Shalit, I.; Fridkin, M. All-d-magainin: Chirality, antimicrobial activity and proteolytic resistance. FEBS Lett. 1990, 274, 151–155. [Google Scholar] [PubMed] [Green Version]
- Liu, S.; Yang, H.; Wan, L.; Cai, H.W.; Li, S.F.; Li, Y.P.; Cheng, J.Q.; Lu, X.F. Enhancement of cytotoxicity of antimicrobial peptide magainin ii in tumor cells by bombesin-targeted delivery. Acta Pharmacol. Sin. 2011, 32, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Ding, X.; Li, W.; Lu, T.; Ma, C.; Xi, X.; Wang, L.; Zhou, M.; Burden, R.; Chen, T. Discovery of two skin-derived dermaseptins and design of a tat-fusion analogue with broad-spectrum antimicrobial activity and low cytotoxicity on healthy cells. PeerJ 2018, 6, e5635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mechlia, M.B.; Belaid, A.; Castel, G.; Jallet, C.; Mansfield, K.L.; Fooks, A.R.; Hani, K.; Tordo, N. Dermaseptins as potential antirabies compounds. Vaccine 2019, 37, 4694–4700. [Google Scholar] [CrossRef]
- Cichon, T.; Smolarczyk, R.; Matuszczak, S.; Barczyk, M.; Jarosz, M.; Szala, S. D-k6l 9 peptide combination with il-12 inhibits the recurrence of tumors in mice. Arch. Immunol. Ther. Exp. 2014, 62, 341–351. [Google Scholar] [CrossRef] [Green Version]
- Jakel, C.E.; Meschenmoser, K.; Kim, Y.; Weiher, H.; Schmidt-Wolf, I.G. Efficacy of a proapoptotic peptide towards cancer cells. In Vivo 2012, 26, 419–426. [Google Scholar]
- Horton, K.L.; Kelley, S.O. Engineered apoptosis-inducing peptides with enhanced mitochondrial localization and potency. J. Med. Chem. 2009, 52, 3293–3299. [Google Scholar] [CrossRef]
- Bahmani, T.; Sharifzadeh, S.; Tamaddon, G.H.; Farzadfard, E.; Zare, F.; Fadaie, M.; Alizadeh, M.; Hadi, M.; Ranjbaran, R.; Mosleh-Shirazi, M.A.; et al. Mitochondrial targeted peptide (klaklak)2, and its synergistic radiotherapy effects on apoptosis of radio resistant human monocytic leukemia cell line. J. Biomed. Phys. Eng. 2019. [Google Scholar] [CrossRef]
- Bouchet, S.; Tang, R.; Fava, F.; Legrand, O.; Bauvois, B. The cngrc-gg-d(klaklak)2 peptide induces a caspase-independent, ca2+-dependent death in human leukemic myeloid cells by targeting surface aminopeptidase n/cd13. Oncotarget 2016, 7, 19445–19467. [Google Scholar] [CrossRef]
- Kudrimoti, M.; Curtis, A.; Azawi, S.; Worden, F.; Katz, S.; Adkins, D.; Bonomi, M.; Elder, J.; Sonis, S.T.; Straube, R.; et al. Dusquetide: A novel innate defense regulator demonstrating a significant and consistent reduction in the duration of oral mucositis in preclinical data and a randomized, placebo-controlled phase 2a clinical study. J. Biotechnol. 2016, 239, 115–125. [Google Scholar] [CrossRef] [Green Version]
- Kudrimoti, M.; Curtis, A.; Azawi, S.; Worden, F.; Katz, S.; Adkins, D.; Bonomi, M.; Scott, Z.; Elder, J.; Sonis, S.T.; et al. Dusquetide: Reduction in oral mucositis associated with enduring ancillary benefits in tumor resolution and decreased mortality in head and neck cancer patients. Biotechnol. Rep. 2017, 15, 24–26. [Google Scholar] [CrossRef] [PubMed]
- Nilius, B.; Owsianik, G. The transient receptor potential family of ion channels. Genome Biol. 2011, 12, 218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Boustany, C.; Bidaux, G.; Enfissi, A.; Delcourt, P.; Prevarskaya, N.; Capiod, T. Capacitative calcium entry and transient receptor potential canonical 6 expression control human hepatoma cell proliferation. Hepatology 2008, 47, 2068–2077. [Google Scholar] [CrossRef] [PubMed]
- Anand, P.; Filipenko, P.; Huaman, J.; Lyudmer, M.; Hossain, M.; Santamaria, C.; Huang, K.; Ogunwobi, O.O.; Holford, M. Antitumor effects of tv1 venom peptide in liver cancer. BioRxiv 2019, 518340. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.H.; Jobichen, C.; Chia, C.S.B.; Chan, T.H.M.; Tang, J.P.; Chung, T.X.Y.; Li, J.; Poulsen, A.; Hung, A.W.; Koh-Stenta, X.; et al. Targeting cancer addiction for sall4 by shifting its transcriptome with a pharmacologic peptide. Proc. Natl. Acad. Sci. USA 2018, 115, E7119–E7128. [Google Scholar] [CrossRef] [Green Version]
- Lo, A.; Lin, C.T.; Wu, H.C. Hepatocellular carcinoma cell-specific peptide ligand for targeted drug delivery. Mol. Cancer Ther. 2008, 7, 579–589. [Google Scholar] [CrossRef] [Green Version]
- Pittala, S.; Krelin, Y.; Shoshan-Barmatz, V. Targeting liver cancer and associated pathologies in mice with a mitochondrial vdac1-based peptide. Neoplasia 2018, 20, 594–609. [Google Scholar] [CrossRef]
- Chen, J.T.; Ma, R.; Sun, S.C.; Zhu, X.F.; Xu, X.L.; Mu, Q. Synthesis and biological evaluation of cyclopeptide gg-8-6 and its analogues as anti-hepatocellular carcinoma agents. Bioorg. Med. Chem. 2018, 26, 609–622. [Google Scholar] [CrossRef]
- Wang, S.; Zhu, J.; Liu, Y. A novel anti-adhesion peptide (β3) inhibits hepatocellular carcinoma activity In Vitro and In Vivo. Oncol. Lett. 2016, 12, 4744–4748. [Google Scholar] [CrossRef] [Green Version]
- Xia, L.; Wu, Y.; Ma, J.I.; Yang, J.; Zhang, F. The antibacterial peptide from bombyx mori cecropinxj induced growth arrest and apoptosis in human hepatocellular carcinoma cells. Oncol. Lett. 2016, 12, 57–62. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.L.; Li, J.H.; Yu, C.Y.; Lin, C.J.; Chiu, P.H.; Chen, P.W.; Lin, C.C.; Chen, W.J. Novel cationic antimicrobial peptide gw-h1 induced caspase-dependent apoptosis of hepatocellular carcinoma cell lines. Peptides 2012, 36, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.; Zhou, C.; Hou, X.; Liu, Y.; Wang, Z.; Peng, X.; Zhang, Z.; Wang, R.; Kong, D. Molecular characterization and bioactivity evaluation of two novel bombinin peptides from the skin secretion of oriental fire-bellied toad, bombina orientalis. Amino Acids 2018, 50, 241–253. [Google Scholar] [CrossRef] [PubMed]
- Ashley, C.E.; Carnes, E.C.; Epler, K.E.; Padilla, D.P.; Phillips, G.K.; Castillo, R.E.; Wilkinson, D.C.; Wilkinson, B.S.; Burgard, C.A.; Kalinich, R.M.; et al. Delivery of small interfering rna by peptide-targeted mesoporous silica nanoparticle-supported lipid bilayers. ACS Nano 2012, 6, 2174–2188. [Google Scholar] [CrossRef] [PubMed]
- Ashley, C.E.; Carnes, E.C.; Phillips, G.K.; Durfee, P.N.; Buley, M.D.; Lino, C.A.; Padilla, D.P.; Phillips, B.; Carter, M.B.; Willman, C.L.; et al. Cell-Specific delivery of diverse cargos by bacteriophage ms2 virus-like particles. ACS Nano 2011, 5, 5729–5745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Hu, Y.; Xiao, J.; Liu, G.; Li, X.; Zhao, Y.; Tan, H.; Shi, H.; Cheng, D. Investigation of sp94 peptide as a specific probe for hepatocellular carcinoma imaging and therapy. Sci. Rep. 2016, 6, 33511. [Google Scholar] [CrossRef] [PubMed]
- Shoshan-Barmatz, V.; De Pinto, V.; Zweckstetter, M.; Raviv, Z.; Keinan, N.; Arbel, N. Vdac, a multi-functional mitochondrial protein regulating cell life and death. Mol. Asp. Med. 2010, 31, 227–285. [Google Scholar] [CrossRef] [PubMed]
- Shoshan-Barmatz, V.; Mizrachi, D.; Keinan, N. Oligomerization of the mitochondrial protein vdac1: From structure to function and cancer therapy. Prog. Mol. Biol. Transl. Sci. 2013, 117, 303–334. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
| AMP Name | Amino Acid Sequence | Structure Class | Net Charge | Source | Tumor Target | Mechanism | Ref. |
|---|---|---|---|---|---|---|---|
| Cathelicidins LL37 hCAP18 | LLGDFFRKSKEKIGKEFKRIVQRIKDFLRNLVPRTES | unknown | 6 | Human | HTC/STC | MP/Apoptosis | [84,85,86,87,88] |
| α-Defensin-1 HNP-1 | ACYCRIPACIAGERRYGTCIYQGRLWAFCC | beta | 3 | Human | HTC/STC | Apoptosis | [89,90] |
| Human b-defensin-3 (hBD3) | GIINTLQKYYCRVRGGRCAVLSCLPKEEQIGKCSTRGRKCCRRKK | mixed structure | 11 | Human | HTC/STC | MP | [91] |
| Lactoferricin B(LfcinB) | FKCRRWQWRMKKLG APSITCVRRAF | beta | 8 | Bovine | HTC/STC | MP/Apoptosis | [92,93] |
| Gomesin | * ZCRRLCYKQRCVTYCRGR | beta | 6 | Spider | STC | MP | [94] |
| Mastoparan-C (MP-C) | LNLKALLAVAKKIL | helix | 4 | Venom | STC | Apoptosis | [95] |
| Cecropin B | KWKVFKKIEKMGRNIRNGIVKAGPAIAVLGEAKAL | unknown | 8 | Silk moth | HTC/STC | MP/Apoptosis | [96,97] |
| Magainin 2 | GIGKFLHSAKKFGKAFVGEIMNS | helix | 3 | Frog | HTC/STC | MP | [98,99] |
| CA-MA-2 | KWKLFKKIPKFLHSAKKF | helix | 8 | Hybrid | STC | MP | [100] |
| BuforinIIb | RAGLQFPVG[RLLR]3 | unstructured | 7 | Frog | HTC/STC | Apoptosis | [101] |
| Brevenin-2R | KFALGKVNAKLQSLNAKSLKQSGCC | helix | 5 | Frog | STC | LDP | [102] |
| LFB | GLFSVVKGVLKGVGKNVSGSLLDQLKCKISGGC | unknown | 4 | Frog | STC | MP | [103] |
| Phylloseptin-PHa | FLSLIPAAISAVSALANHF | helix | 2 | frog | STC | MP | [104] |
| Ranatuerin-2PLx | GIMDTVKNAAKNLAGQLLDKLKCSITAC | helix | 2 | frog | STC | Apoptosis | [105] |
| Dermaseptin-PS1 | ALWKTMLKKLGTVALHAGKAALGAVADTISQ | helix | 5 | frog | STC/ ICD | MP | [106,107] |
| Dermaseptin (DPT9) | GLWSKIKDAAKTAGKAALGFVNEMV | helix | 2 | Phyllomedusatarsius | STC | MP | [108] |
| chrysophsin-1 | FFGWLIKGAIHAGKAIHGLI | helix | 6 | Red sea bream | HTC/STC | MP | [109] |
| Ss-arasin | SPRVRRRYGRPFGGRPFVGGQFGGRPGCVCIRSPCPCANYG | bridge | 8 | Indian mud crab | STC | uncharacterized | [110] |
| Turgencin A and B | GIKEMLCNMACAQTVCKKSGGPLCDTCQAACKALG | helix | 3 | Synoicum turgens | STC | uncharacterized | [111] |
| D-K6L9 | LKLLKKLLKKLLKLL | helix | 3 | Engineered | STC | MP | [112] |
| KLA | RRQRRTSKLMKRGGKLAKLAKKLAKLAK(KLAKLAK)2 | unknown | 19 | Engineered | STC | MP | [113] |
| LTX-315 | K-K-W-W-K-K-W-Dip-K | unknown | 5 | Engineered | HTC/STC | MP/ICD | [114,115] |
| TAT-RasGAP317-326 | RRRQRRKKRGGGDTRLNTVWMW | unknown | 8 | Engineered | STC | MP | [116] |
| Peptide Sequence | Clinical Stages | Indications | Identifier Number |
|---|---|---|---|
| LL37: LLGDFFRKSKEKIGKEFFJVQRIKDFLRNLVPRTES | Phase II | Melanoma | NCT02225366 |
| LTX315: KKWWKK-Dip-K-NH2 | Phase I Phase I Phase II | Solid tumors | NCT01058616 NCT01986426 NCT03725605 |
| SGX942 Dusquetide:RIVPA | Phase III | Head and neck cancer | NCT03237325 |
| AMP Name | Amino Acid Sequence | Structure Class | Net Charge | Source | Mechanism | Ref. |
|---|---|---|---|---|---|---|
| Tv1 | TRICCGCYWNGSKDVCSQSCC | mixed | 2 | Venom | Apoptosis | [142,143,144] |
| SALL4 peptide FFW | RRKFAKFQWI | mixed | 4 | Engineered | Silencing tumor-suppressor gene | [145] |
| SP94 | SFSIIHTPILPL | unknown | 0 | Human | Apoptosis | [68,146] |
| R-Tf-D-LP4 | KWTWKNSNGATWALNVATELKKEWTWSHRPYIAH | unknown | 5 | Human | Apoptosis | [147] |
| GG-8-6 | cyclo-VLPILLVL | cyclic | 0 | Plant | Apoptosis | [148] |
| β3 | DLYYLMDLSYSMKGGDLYYL MDLSYSMKGGDLYYLMDLSYSMK | unknown | 3 | Engineered | Anti adhesion activity | [149] |
| CecropinX | RWKIFKKIEKMGRNIRDGIVKAGPAIEVLGSAKAIGK | unkonown | 10 | Silk moth | Apoptosis | [150] |
| GW-H1 | GYNYAKKLANLAKKPANALW | helix | 4 | Engineered | Apoptosis | [151] |
| Bombinin | GIGSAILSAGKSIIKGLAKGLAEHF IIGPVLGLVGKALGGLL | helix unkonown | 31 | Fire-bellied toad | Apoptosis | [152] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tornesello, A.L.; Borrelli, A.; Buonaguro, L.; Buonaguro, F.M.; Tornesello, M.L. Antimicrobial Peptides as Anticancer Agents: Functional Properties and Biological Activities. Molecules 2020, 25, 2850. https://doi.org/10.3390/molecules25122850
Tornesello AL, Borrelli A, Buonaguro L, Buonaguro FM, Tornesello ML. Antimicrobial Peptides as Anticancer Agents: Functional Properties and Biological Activities. Molecules. 2020; 25(12):2850. https://doi.org/10.3390/molecules25122850
Chicago/Turabian StyleTornesello, Anna Lucia, Antonella Borrelli, Luigi Buonaguro, Franco Maria Buonaguro, and Maria Lina Tornesello. 2020. "Antimicrobial Peptides as Anticancer Agents: Functional Properties and Biological Activities" Molecules 25, no. 12: 2850. https://doi.org/10.3390/molecules25122850