
Chemo-Protective Potential of Cerium Oxide Nanoparticles against Fipronil-Induced Oxidative Stress, Apoptosis, Inflammation and Reproductive Dysfunction in Male White Albino Rats


 ,
,  , , ,
, , ,  ,
,  , ,
, ,  and
and 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Experimental Animals and Protocol
2.2. Oxidative Stress/Antioxidants Test
2.3. Quantitative Reverse Transcription-PCR Test (RT-qPCR)
2.4. Histopathological Assessment
2.5. Immunohistochemistry (IHC)
2.6. Enzyme-Linked Immunosorbent Assay (ELISA)
2.7. Method and Machine Precision
2.8. Statistical Analysis
3. Results Sections in Wrong Order–Experimental Is Last–Renumber Everything Affected
3.1. Quality Control
3.2. Antioxidant Status and Lipid Peroxidation in Testicular Tissue and Serum Testosterone Levels
3.3. Gene Expressions in Testicular Tissue
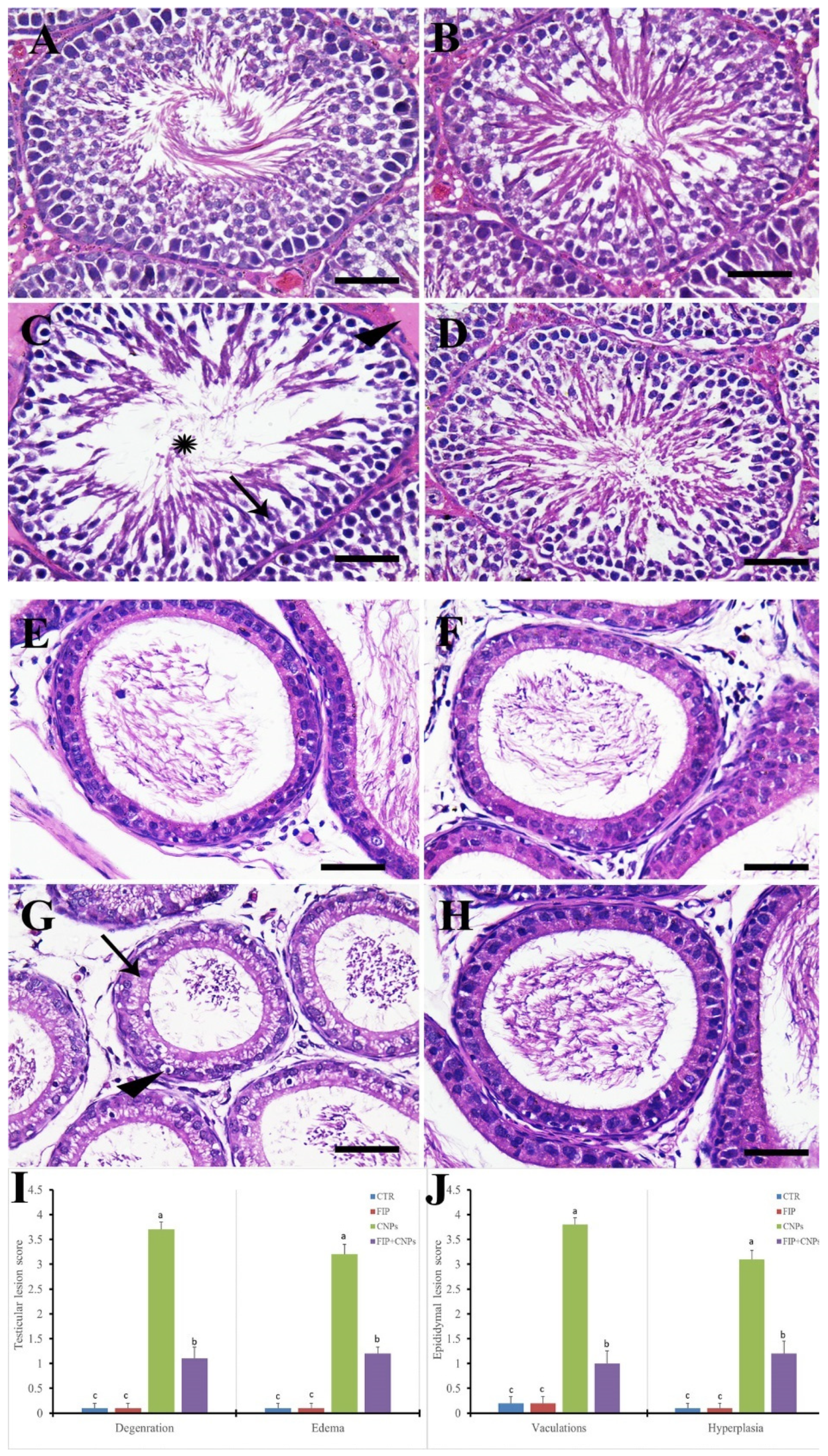
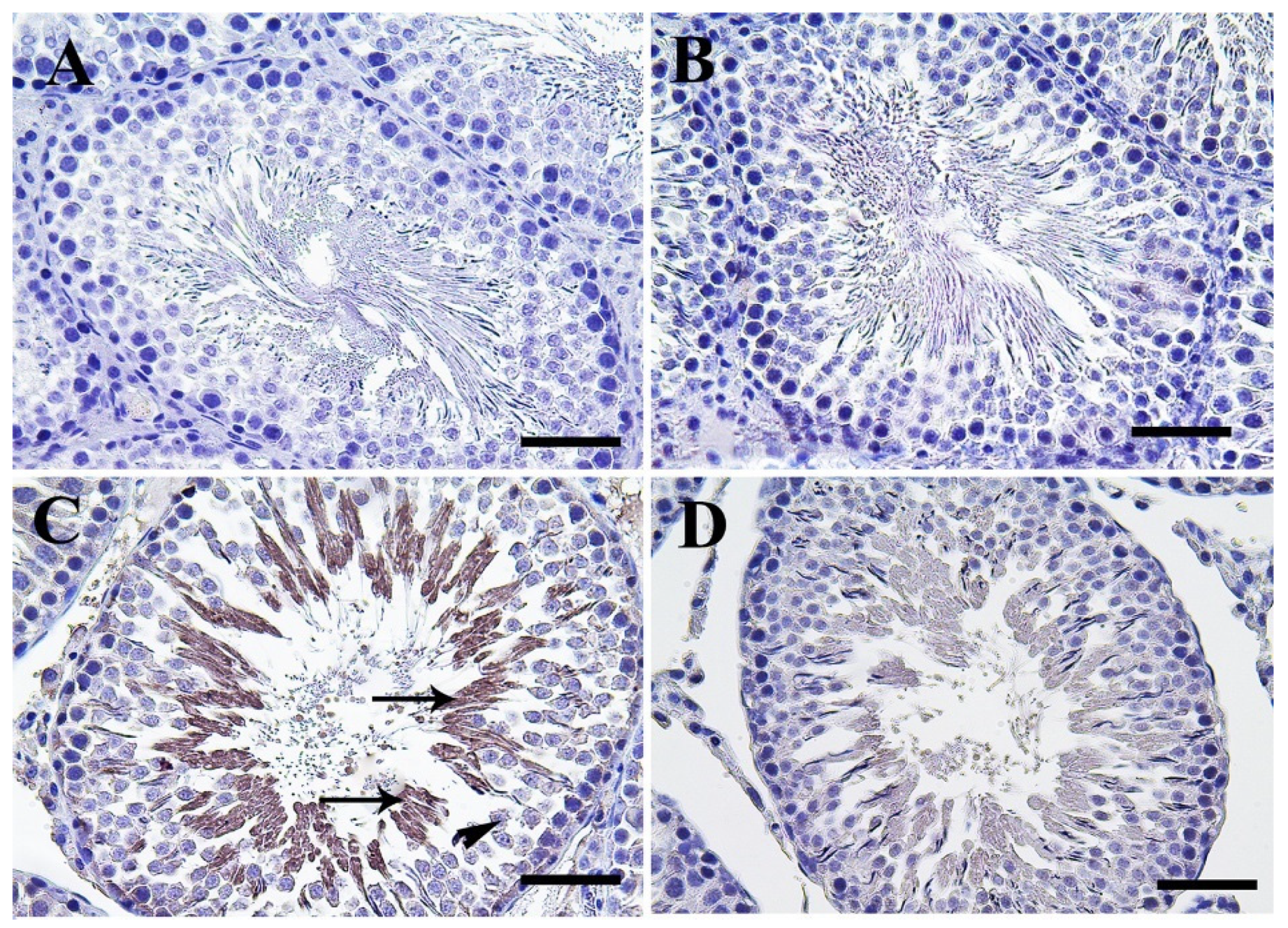
3.4. Histopathological Assessment of Testicular Tissue
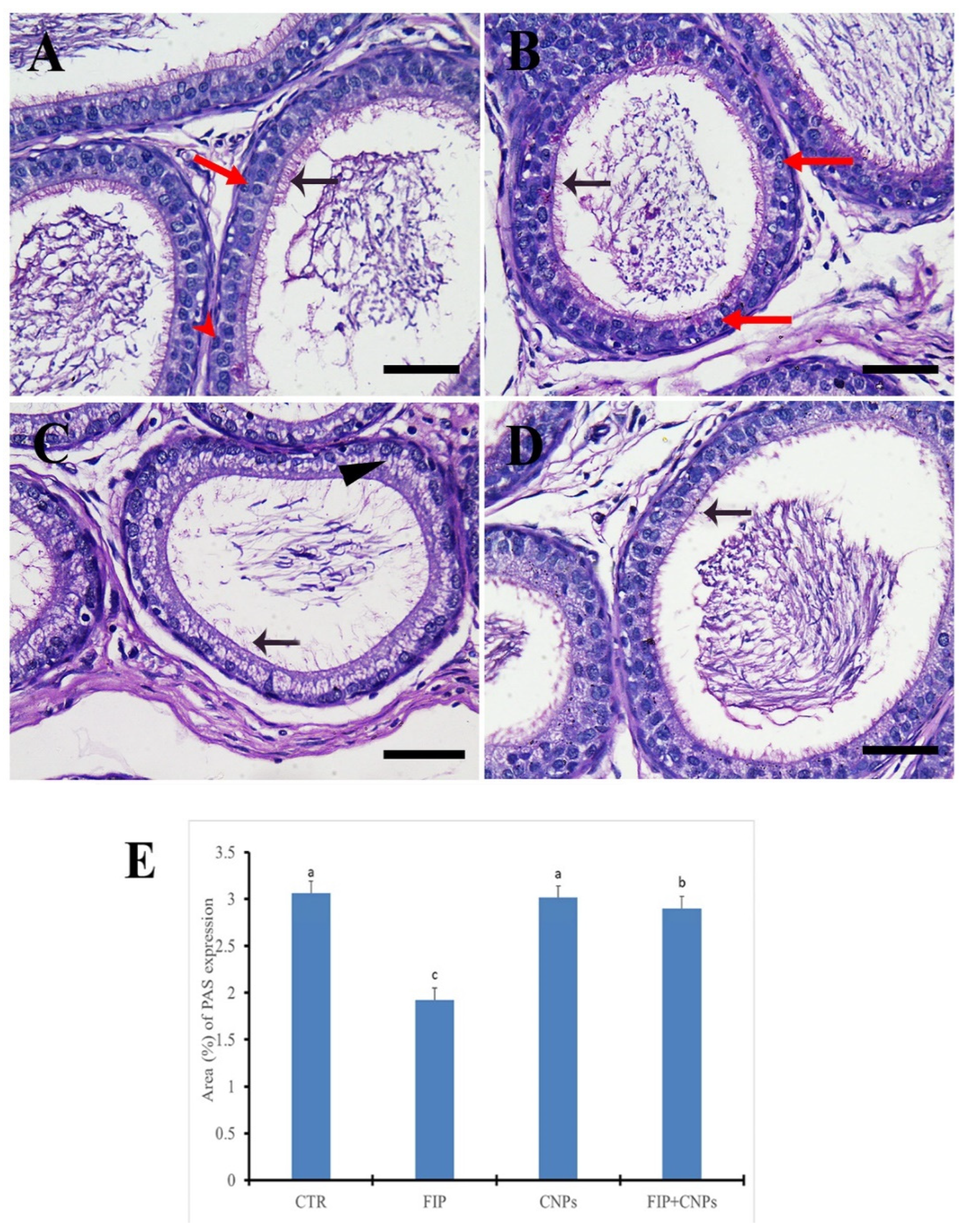
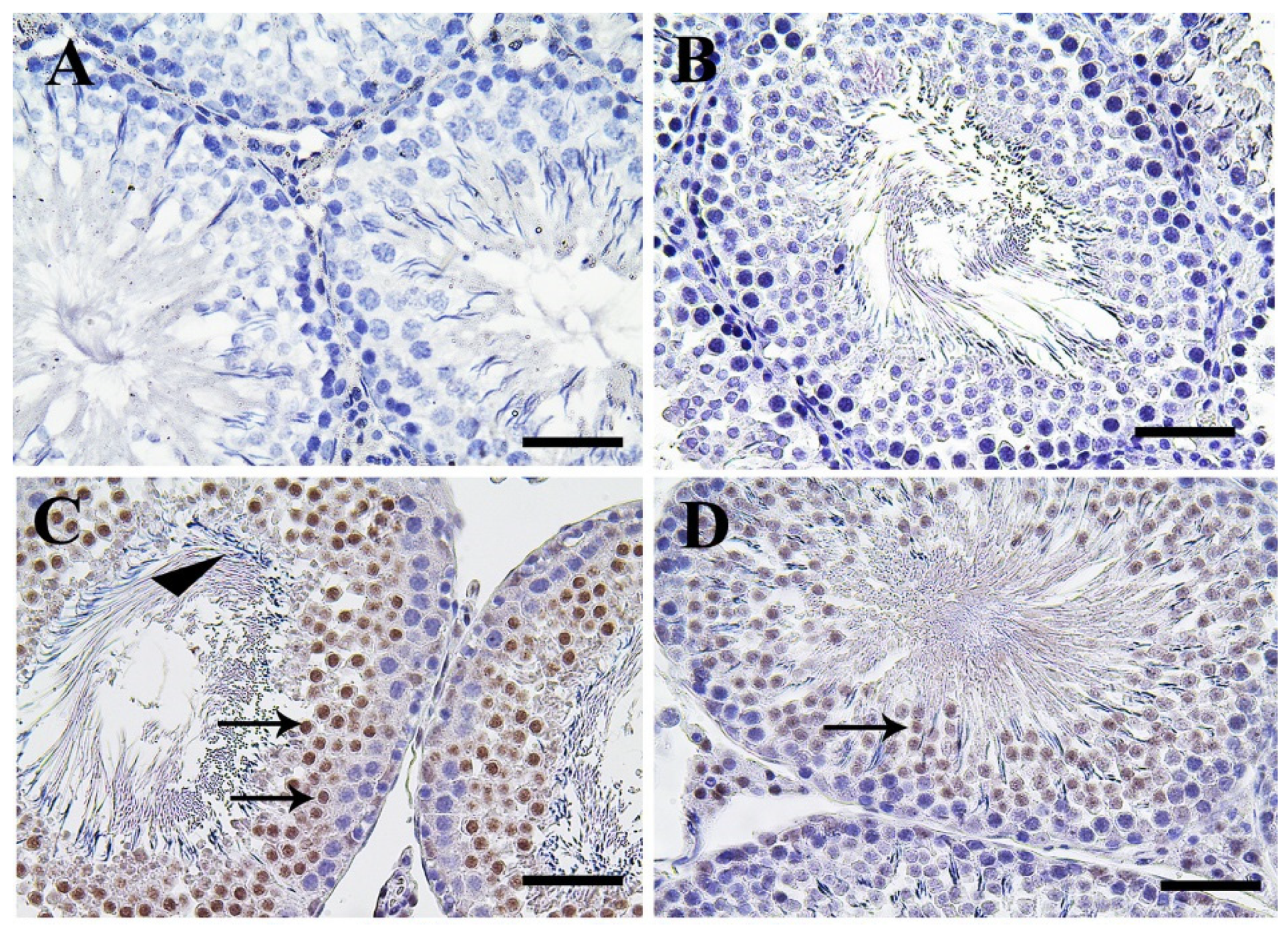
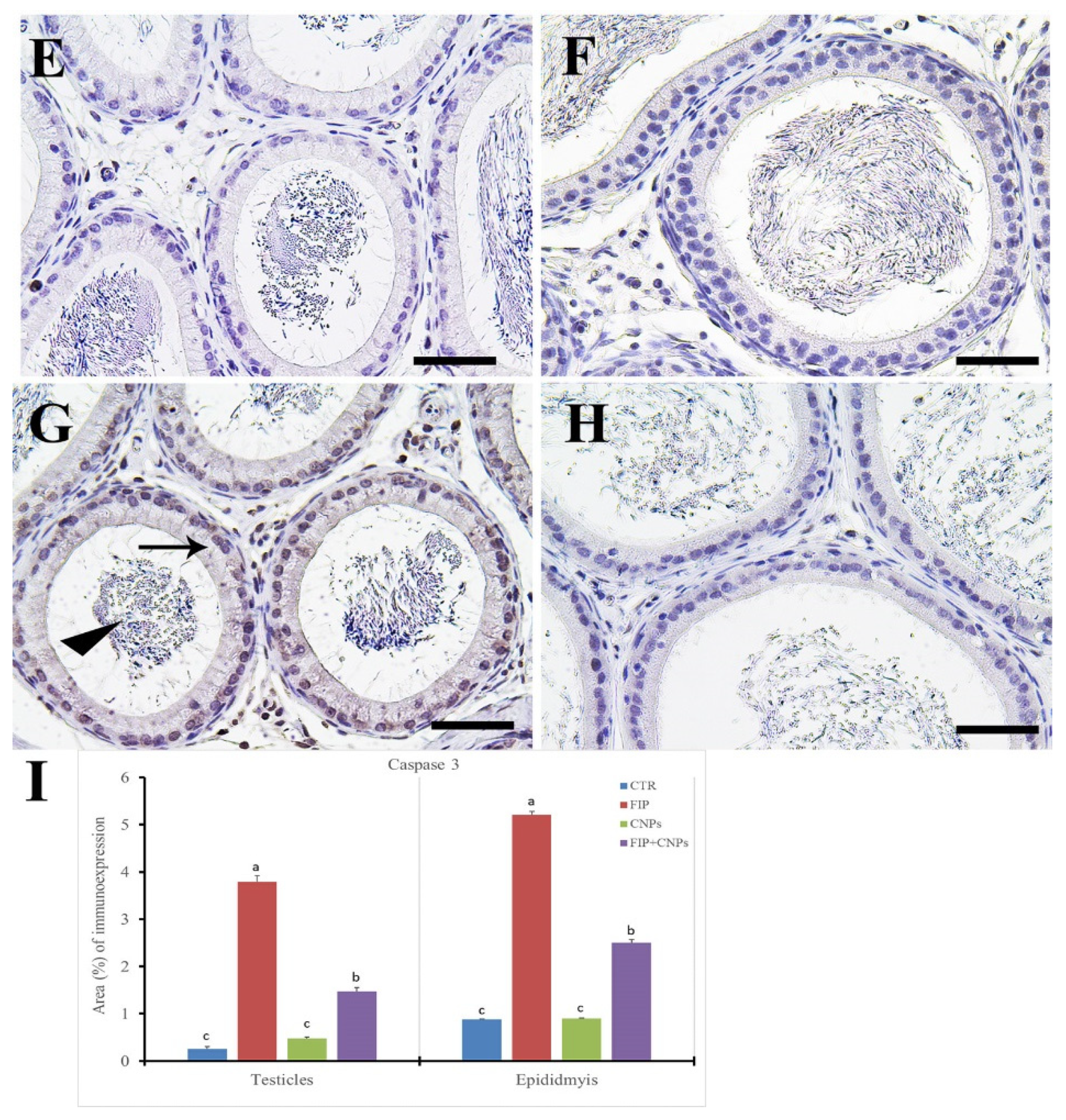
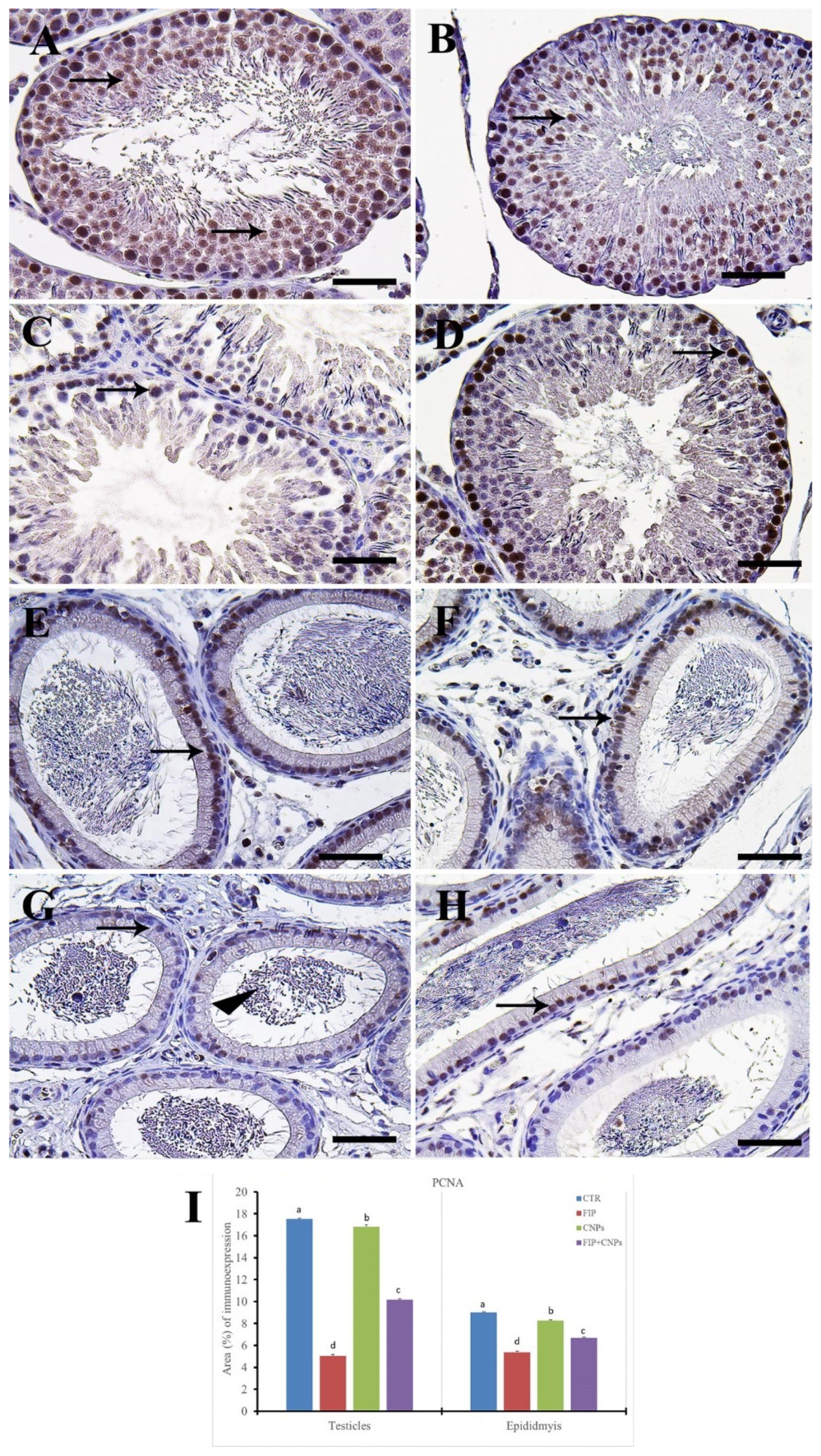
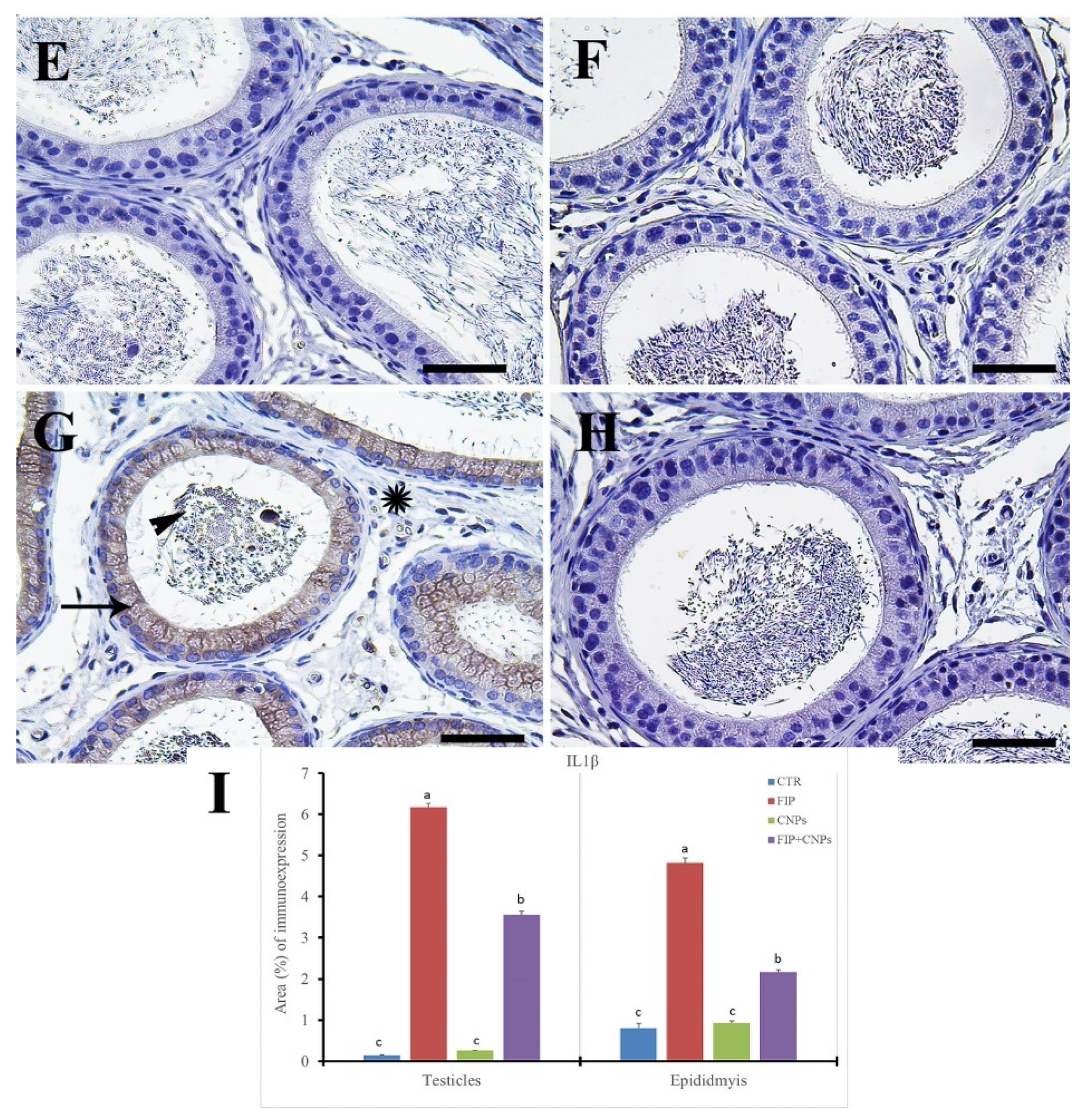
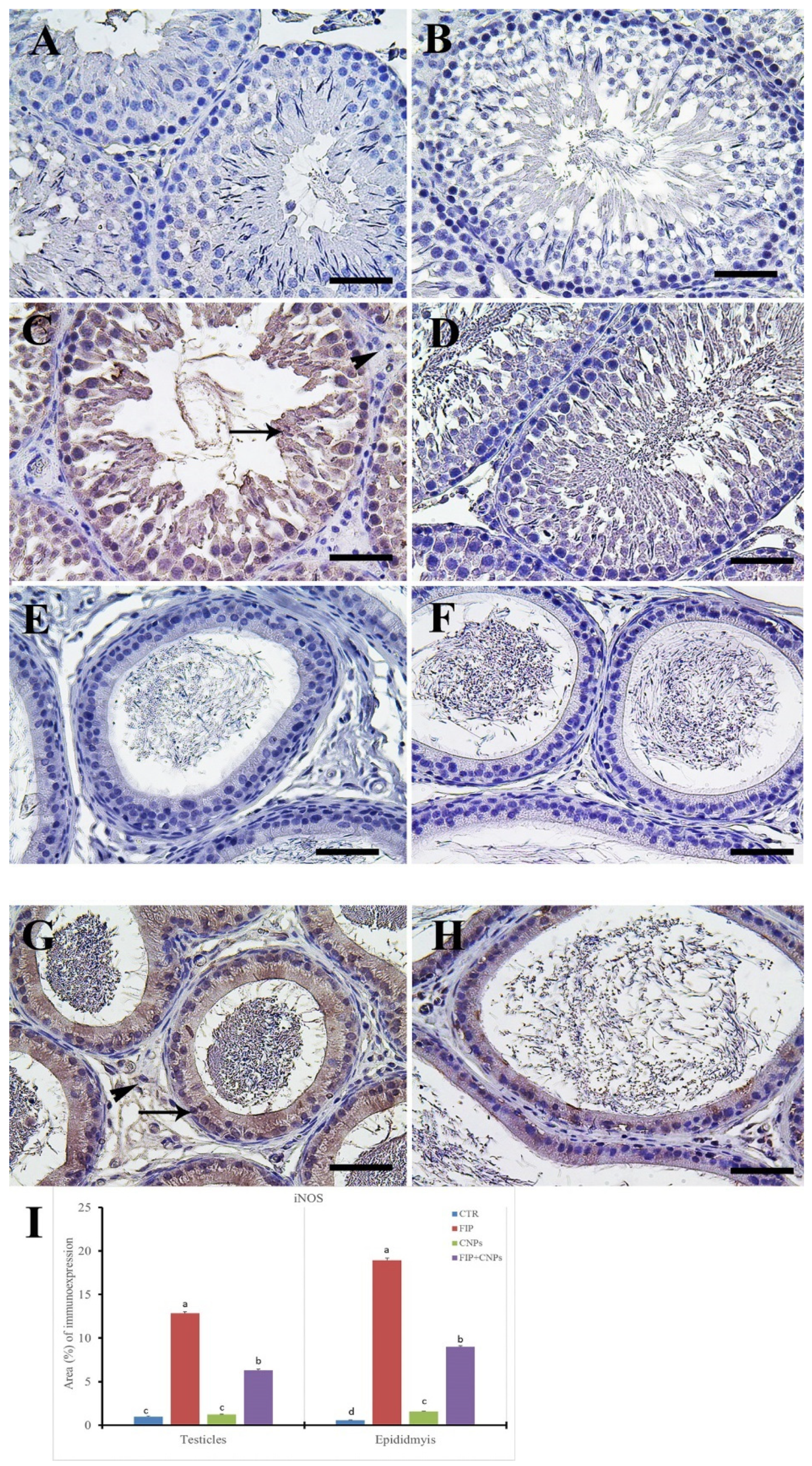
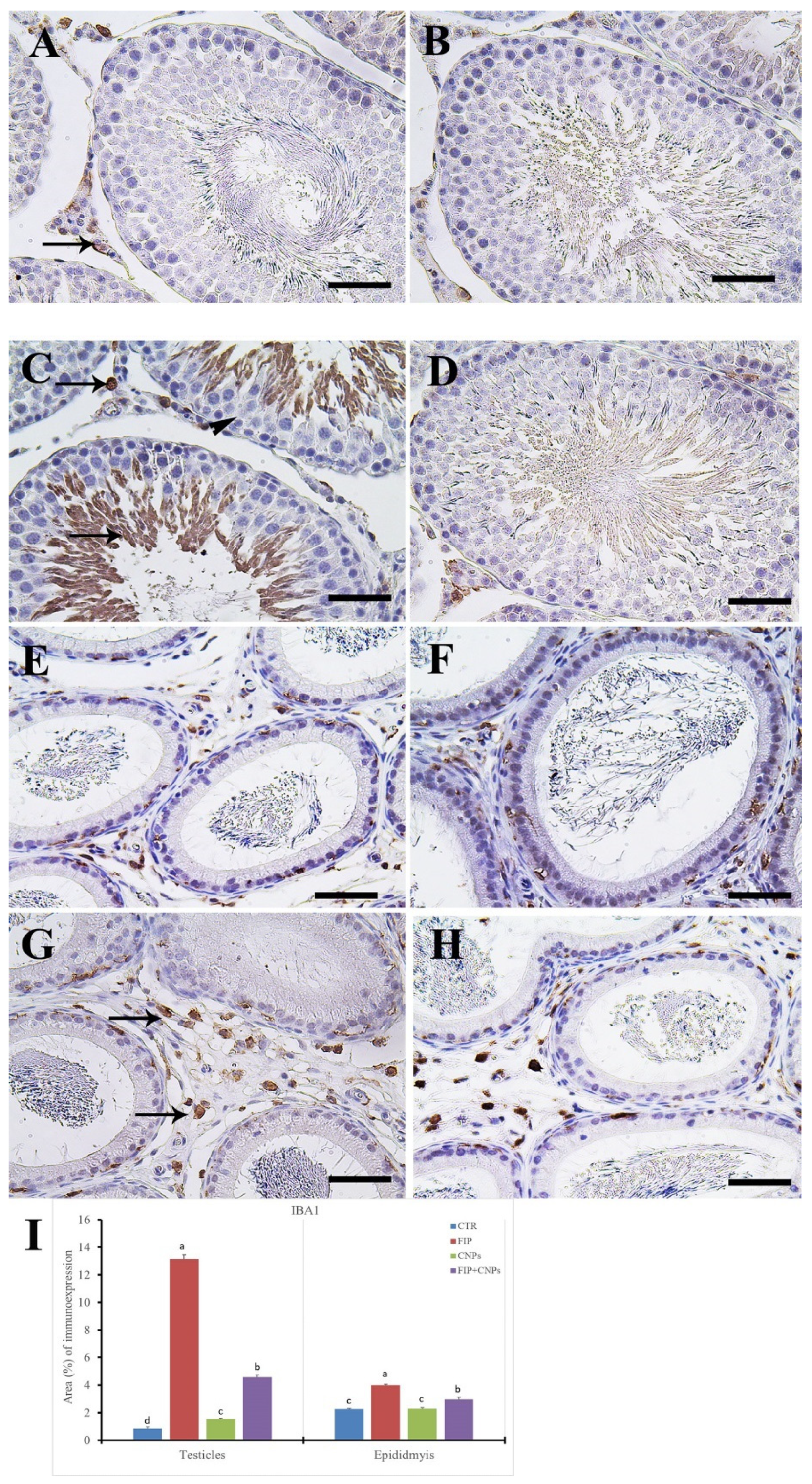
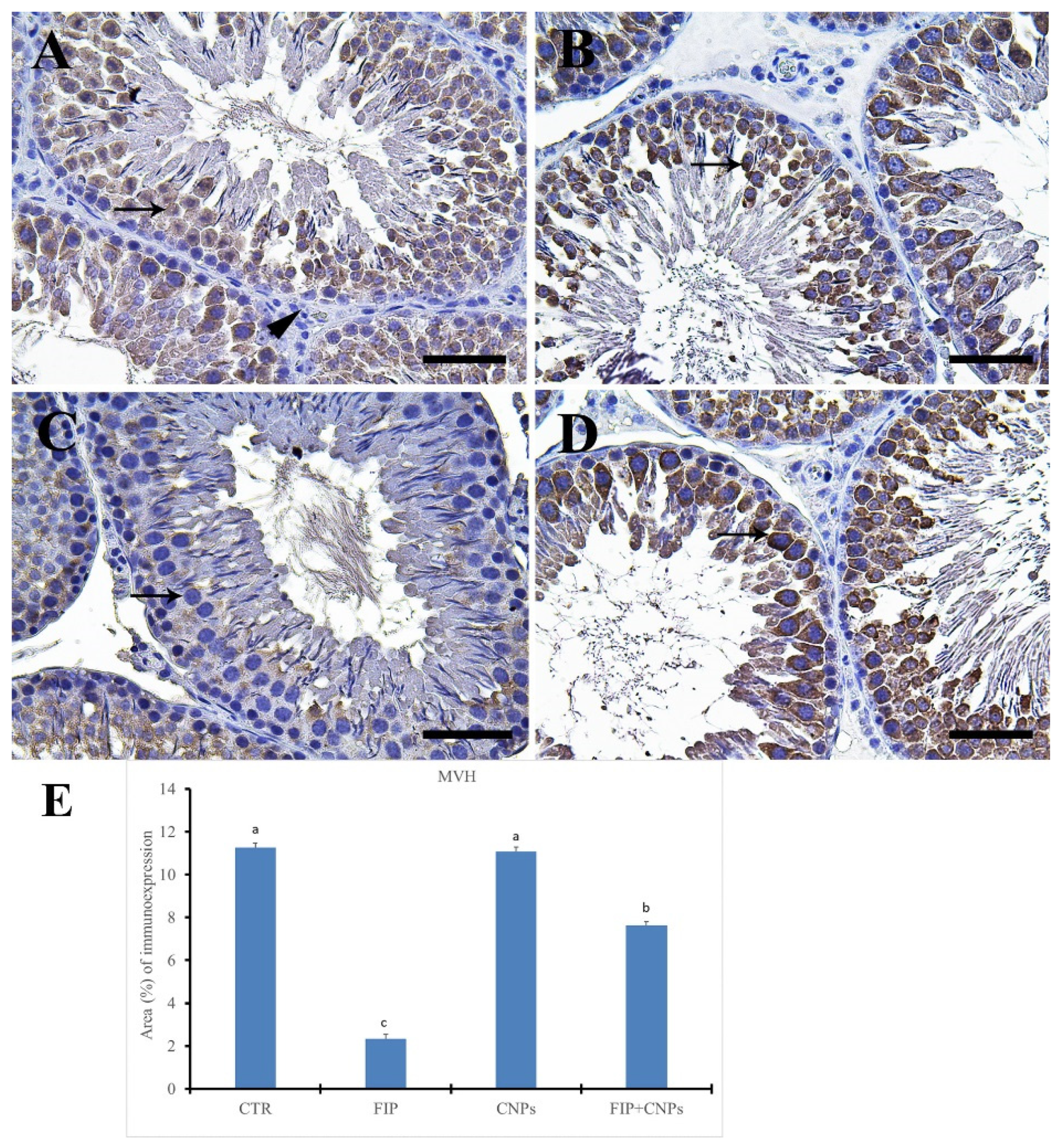
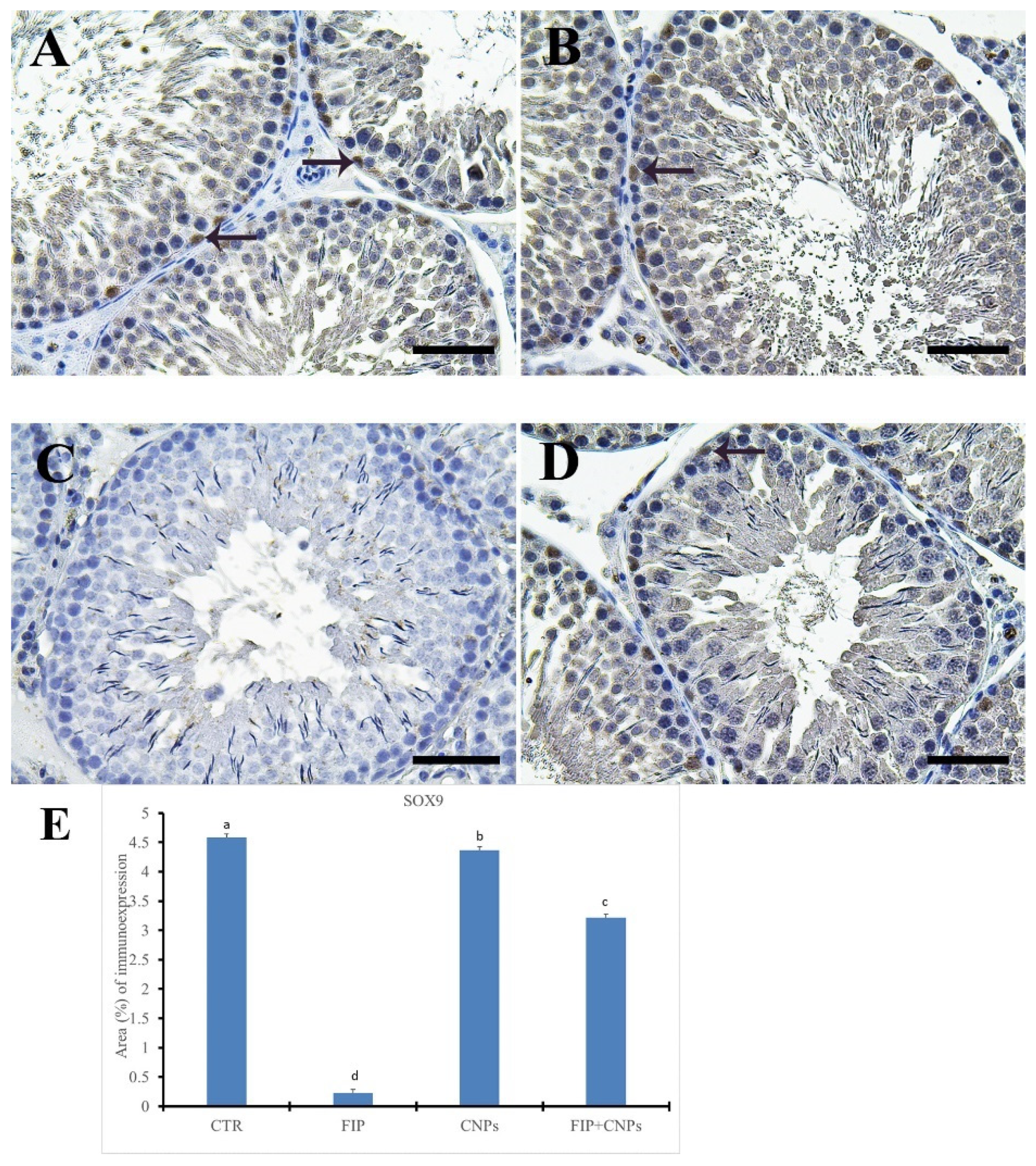
3.5. Immunohistochemistry Assessment of Testicular Tissue
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jensen, T.K.; Bonde, J.P.; Joffe, M. The influence of occupational exposure on male reproductive function. Occup. Med. 2006, 56, 544–553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.-Z.; Liu, S.-S.; Sun, Y.; Wu, J.-Y.; Zhou, Y.-L.; Zhang, J.-H. Beta-cypermethrin impairs reproductive function in male mice by inducing oxidative stress. Theriogenology 2009, 72, 599–611. [Google Scholar] [CrossRef] [PubMed]
- Vidau, C.; Brunet, J.-L.; Badiou, A.; Belzunces, L.P. Phenylpyrazole insecticides induce cytotoxicity by altering mechanisms involved in cellular energy supply in the human epithelial cell model Caco-2. Toxicol. In Vitro 2009, 23, 589–597. [Google Scholar] [CrossRef] [PubMed]
- Avudainayagam, S.; Megharaj, M.; Owens, G.; Kookana, R.S.; Chittleborough, D.; Naidu, R. Chemistry of chromium in soils with emphasis on tannery waste sites. In Reviews of Environmental Contamination and Toxicology; Springer: Berlin/Heidelberg, Germany, 2003; pp. 53–91. [Google Scholar]
- Gaines, T.B. Acute toxicity of pesticides. Toxicol. Appl. Pharmacol. 1969, 14, 515–534. [Google Scholar] [CrossRef]
- Kitulagodage, M. Impact of Fipronil, a New Generation Pesticide, on Avian Development and Health. Ph.D. Thesis, University of Wollongong, Wollongong, Australia, 2011. [Google Scholar]
- World Health Organization. The WHO Recommended Classification of Pesticides by Hazard and Guidelines to Classification 2009; World Health Organization: Geneva, Switzerland, 2010. [Google Scholar]
- Ohi, M.; Dalsenter, P.; Andrade, A.; Nascimento, A. Reproductive adverse effects of fipronil in Wistar rats. Toxicol. Lett. 2004, 146, 121–127. [Google Scholar] [CrossRef]
- Bencic, D.C.; Villeneuve, D.L.; Biales, A.D.; Blake, L.; Durhan, E.J.; Jensen, K.M.; Kahl, M.D.; Makynen, E.A.; Martinović-Weigelt, D.; Ankley, G.T. Effects of the insecticide fipronil on reproductive endocrinology in the fathead minnow. Environ. Toxicol. Chem. 2013, 32, 1828–1834. [Google Scholar] [CrossRef]
- Beshbishy, A.M.; Batiha, G.E.; Yokoyama, N.; Igarashi, I. Ellagic acid microspheres restrict the growth of Babesia and Theileria in vitro and Babesia microti in vivo. Parasites Vectors 2019, 12, 269. [Google Scholar] [CrossRef] [Green Version]
- Das, S.; Dowding, J.M.; Klump, K.E.; McGinnis, J.F.; Self, W.; Seal, S. Cerium oxide nanoparticles: Applications and prospects in nanomedicine. Nanomed. Nanotechnol. Biol. Med. 2013, 8, 1483–1508. [Google Scholar] [CrossRef]
- Culcasi, M.; Benameur, L.; Mercier, A.; Lucchesi, C.; Rahmouni, H.; Asteian, A.; Casano, G.; Botta, A.; Kovacic, H.; Pietri, S. EPR spin trapping evaluation of ROS production in human fibroblasts exposed to cerium oxide nanoparticles: Evidence for NADPH oxidase and mitochondrial stimulation. Chem. Biol. Interact. 2012, 199, 161–176. [Google Scholar] [CrossRef]
- Korsvik, C.; Patil, S.; Seal, S.; Self, W.T. Superoxide dismutase mimetic properties exhibited by vacancy engineered ceria nanoparticles. Chem. Commun. 2007, 10, 1056–1058. [Google Scholar] [CrossRef]
- Celardo, I.; Pedersen, J.Z.; Traversa, E.; Ghibelli, L. Pharmacological potential of cerium oxide nanoparticles. Nanoscale 2011, 3, 1411–1420. [Google Scholar] [CrossRef] [PubMed]
- Heckert, E.G.; Karakoti, A.S.; Seal, S.; Self, W.T. The role of cerium redox state in the SOD mimetic activity of nanoceria. Biomaterials 2008, 29, 2705–2709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobyliak, N.M.; Falalyeyeva, T.M.; Kuryk, O.G.; Beregova, T.V.; Bodnar, P.M.; Zholobak, N.M.; Shcherbakov, O.B.; Bubnov, R.V.; Spivak, M.Y. Antioxidative effects of cerium dioxide nanoparticles ameliorate age-related male infertility: Optimistic results in rats and the review of clinical clues for integrative concept of men health and fertility. EPMA J. 2015, 6, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caballero, M.; Ares, I.; Martinez, M.; Martinez-Larranaga, M.; Anadon, A.; Martinez, M. Fipronil induces CYP isoforms in rats. Food Chem. Toxicol. 2015, 83, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Pakzad, M.; Fouladdel, S.; Nili-Ahmadabadi, A.; Pourkhalili, N.; Baeeri, M.; Azizi, E.; Sabzevari, O.; Ostad, S.N.; Abdollahi, M. Sublethal exposures of diazinon alters glucose homostasis in Wistar rats: Biochemical and molecular evidences of oxidative stress in adipose tissues. Pestic. Biochem. Physiol. 2013, 105, 57–61. [Google Scholar] [CrossRef]
- Draper, H.; Hadley, M. Malondialdehyde determination as index of lipid Peroxidation. In Methods in Enzymology; Elsevier: Cambridge, MA, USA, 1990; Volume 186, pp. 421–431. [Google Scholar]
- Davis, K.L.; Martin, E.; Turko, I.V.; Murad, F. Novel effects of nitric oxide. Annu. Rev. Pharmacol. Toxicol. 2001, 41, 203–236. [Google Scholar] [CrossRef] [PubMed]
- Nishikimi, M.; Appaji, N.; Yagi, K. The occurrence of superoxide anion in the reaction of reduced phenazine methosulfate and molecular oxygen. Biochem. Biophys. Res. Commun. 1972, 46, 849–854. [Google Scholar] [CrossRef]
- Paglia, D.E.; Valentine, W.N. Studies on the quantitative and qualitative characterization of erythrocyte glutathione peroxidase. J. Lab. Clin. Med. 1967, 70, 158–169. [Google Scholar]
- Bancroft, J.D.; Layton, C. The hematoxylin and eosin, connective and mesenchymal tissues with their stains. In Bancroft’s Theory and Practice of Histological Techniques, 7th ed.; Kim, S., Ed.; Churchill Livingstone: Philadelphia, PA, USA, 2013; pp. 173–186. [Google Scholar]
- Layton, C.; Bancroft, J.D. Carbohydrates. In Bancroft’s Theory and Practice of Histological Techniques, 7th ed.; Kim, S., Ed.; Churchill Livingstone: Philadelphia, PA, USA, 2013; pp. 215–238. [Google Scholar]
- Gibson-Corley, K.N.; Olivier, A.K.; Meyerholz, D.K. Principles for valid histopathologic scoring in research. Vet. Pathol. 2013, 50, 1007–1015. [Google Scholar] [CrossRef] [Green Version]
- Johnsen, S.G. Testicular biopsy score count—A method for registration of spermatogenesis in human testes: Normal values and results in 335 hypogonadal males. Horm. Res. Paediatr. 1970, 1, 2–25. [Google Scholar] [CrossRef]
- Noreldin, A.E.; Elewa, Y.H.A.; Kon, Y.; Warita, K.; Hosaka, Y.Z. Immunohistochemical localization of osteoblast activating peptide in the mouse kidney. Acta Histochem. 2018, 120, 323–328. [Google Scholar] [CrossRef] [PubMed]
- Sysel, A.M.; Valli, V.E.; Nagle, R.B.; Bauer, J.A. Immunohistochemical quantification of the vitamin B12 transport protein (TCII), cell surface receptor (TCII-R) and Ki-67 in human tumor xenografts. Anticancer Res. 2013, 33, 4203–4212. [Google Scholar] [PubMed]
- Vis, A.N.; Kranse, R.; Nigg, A.L.; van der Kwast, T.H. Quantitative analysis of the decay of immunoreactivity in stored prostate needle biopsy sections. Am. J. Clin. Pathol. 2000, 113, 369–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ermer, J. Method validation in pharmaceutical analysis. In MS Applications in Drug Development Reference Materials for Chemical Analysis; Ermer, J., Miller, J.H.M., Eds.; Wiley-VCH GmbH and Co KGaA: Weinheim, Germany, 2005; Volume 83. [Google Scholar]
- International Programme on Chemical Safety (IPCS). Harmonization Project Document 11: Guidance Document on Evaluating And Expressing Uncertainty in Hazard Characterization; World Health Organization: Geneva, Switzerland, 2014; pp. 1–146. [Google Scholar]
- Badgujar, P.C.; Pawar, N.N.; Chandratre, G.A.; Telang, A.; Sharma, A. Fipronil induced oxidative stress in kidney and brain of mice: Protective effect of vitamin E and vitamin C. Pestic. Biochem. Physiol. 2015, 118, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Abdelaziz, A.S.; Kamel, M.A.; Ahmed, A.I.; Shalaby, S.I.; El-Darier, S.M.; Magdy Beshbishy, A.; Batiha, G.E.; Alomar, S.Y.; Khodeer, D.M. Chemotherapeutic potential of Epimedium brevicornum extract: The cGMP-specific PDE5 inhibitor as anti-infertility agent following long-term administration of tramadol in male rats. Antibiotics 2020, 9, 318. [Google Scholar] [CrossRef]
- Circu, M.L.; Aw, T.Y. Reactive oxygen species, cellular redox systems, and apoptosis. Free Radic. Biol. Med. 2010, 48, 749–762. [Google Scholar] [CrossRef] [Green Version]
- Pierini, D.; Bryan, N.S. Nitric oxide availability as a marker of oxidative stress. In Advanced Protocols in Oxidative Stress III; Springer: Berlin/Heidelberg, Germany, 2015; pp. 63–71. [Google Scholar]
- Sen, S.; Chakraborty, R. The role of antioxidants in human health. In Oxidative Stress: Diagnostics, Prevention, and Therapy; ACS Publications: Washington, DC, USA, 2011; pp. 1–37. [Google Scholar]
- Bhattacharjee, R.; Sil, P.C. The protein fraction of Phyllanthus niruri plays a protective role against acetaminophen induced hepatic disorder via its antioxidant properties. Phytother. Res. 2006, 20, 595–601. [Google Scholar] [CrossRef]
- Kavlock, R.; Cummings, A. Mode of action: Inhibition of androgen receptor function—Vinclozolin-induced malformations in reproductive development. Crit. Rev. Toxicol. 2005, 35, 721–726. [Google Scholar] [CrossRef]
- Haider, S.G. Cell biology of Leydig cells in the testis. Int. Rev. Cytol. 2004, 233, 181–241. [Google Scholar]
- Kasahara, E.; Miyoshi, M.; Konaka, R.; Hiramoto, K.; Sasaki, J.; Tokuda, M.; Nakano, Y.; Inoue, M. Role of oxidative stress in germ cell apoptosis induced by di (2-ethylhexyl) phthalate. Biochem. J. 2002, 365, 849–856. [Google Scholar] [CrossRef] [Green Version]
- Gao, H.-B.; Tong, M.-H.; Hu, Y.-Q.; You, H.-Y.; Guo, Q.-S.; Ge, R.-S.; Hardy, M.P. Mechanisms of glucocorticoid-induced Leydig cell apoptosis. Mol. Cell. Endocrinol. 2003, 199, 153–163. [Google Scholar] [CrossRef]
- Khan, S.; Jan, M.; Kumar, D.; Telang, A. Firpronil induced spermotoxicity is associated with oxidative stress, DNA damage and apoptosis in male rats. Pestic. Biochem. Physiol. 2015, 124, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Skommer, J.; Wlodkowic, D.; Deptala, A. Larger than life: Mitochondria and the Bcl-2 family. Leuk. Res. 2007, 31, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Sen, N.; Das, B.; Ganguly, A.; Mukherjee, T.; Tripathi, G.; Bandyopadhyay, S.; Rakshit, S.; Sen, T.; Majumder, H. Camptothecin induced mitochondrial dysfunction leading to programmed cell death in unicellular hemoflagellate Leishmania donovani. Cell Death Differ. 2004, 11, 924. [Google Scholar] [CrossRef] [PubMed]
- Puka-Sundvall, M.; Gajkowska, B.; Cholewinski, M.; Blomgren, K.; Lazarewicz, J.W.; Hagberg, H. Subcellular distribution of calcium and ultrastructural changes after cerebral hypoxia-ischemia in immature rats. Dev. Brain Res. 2000, 125, 31–41. [Google Scholar] [CrossRef]
- Chan, W.Y.; Lorke, D.E.; Tiu, S.C.; Yew, D.T. Proliferation and apoptosis in the developing human neocortex. Anat. Rec. 2002, 267, 261–276. [Google Scholar] [CrossRef]
- Sharpe, R.; Turner, K.; McKinnell, C.; Groome, N.; Atanassova, N.; Millar, M.; Buchanan, D.; Cooke, P.S. Inhibin B levels in plasma of the male rat from birth to adulthood: Effect of experimental manipulation of Sertoli cell number. J. Androl. 1999, 20, 94–101. [Google Scholar]
- Meachem, S.; Nieschlag, E.; Simoni, M. Inhibin B in male reproduction: Pathophysiology and clinical relevance. Eur. J. Endocrinol. 2001, 145, 561–571. [Google Scholar] [CrossRef] [Green Version]
- Adeyemi, O.S.; Shittu, E.O.; Akpor, O.B.; Rotimi, D.; Batiha, G.E. Silver nanoparticles restrict microbial growth by promoting oxidative stress and DNA damage. EXCLI J. 2020, 19, 492–500. [Google Scholar]
- Lambright, C.; Ostby, J.; Bobseine, K.; Wilson, V.; Hotchkiss, A.; Mann, P.; Gray, L., Jr. Cellular and molecular mechanisms of action of linuron: An antiandrogenic herbicide that produces reproductive malformations in male rats. Toxicol. Sci. 2000, 56, 389–399. [Google Scholar] [CrossRef] [Green Version]
- Saradha, B.; Mathur, P. Effect of environmental contaminants on male reproduction. Environ. Toxicol. Pharmacol. 2006, 21, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Zeng, L.; Kong, X.-T.; Su, J.-W.; Xia, T.-L.; Na, Y.-Q.; Guo, Y.-L. Evaluation of germ-cell kinetics in infertile patients with proliferating cell nuclear antigen proliferating index. Asian J. Androl. 2001, 3, 63–66. [Google Scholar] [PubMed]
- Yamada, M.; Ohsawa, K.; Imai, Y.; Kohsaka, S.; Kamitori, S. X-ray structures of the microglia/macrophage-specific protein Iba1 from human and mouse demonstrate novel molecular conformation change induced by calcium binding. J. Mol. Biol. 2006, 364, 449–457. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, S.; Suzuki, K.; Watanabe, M.; Matsuda, A.; Tone, S.; Koike, T. Upregulation of a new microglial gene, mrf-1, in response to programmed neuronal cell death and degeneration. J. Neurosci. 1998, 18, 6358–6369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Utans, U.; Arceci, R.J.; Yamashita, Y.; Russell, M.E. Cloning and characterization of allograft inflammatory factor-1: A novel macrophage factor identified in rat cardiac allografts with chronic rejection. J. Clin. Investig. 1995, 95, 2954–2962. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.-W.; Ahren, B.; Östenson, C.-G.; Cintra, A.; Bergman, T.; Möller, C.; Fuxe, K.; Mutt, V.; Jörnvall, H.; Efendic, S. Identification, isolation, and characterization of daintain (allograft inflammatory factor 1), a macrophage polypeptide with effects on insulin secretion and abundantly present in the pancreas of prediabetic BB rats. Proc. Natl. Acad. Sci. USA 1997, 94, 13879–13884. [Google Scholar] [CrossRef] [Green Version]
- Ito, D.; Imai, Y.; Ohsawa, K.; Nakajima, K.; Fukuuchi, Y.; Kohsaka, S. Microglia-specific localisation of a novel calcium binding protein, Iba1. Mol. Brain Res. 1998, 57, 1–9. [Google Scholar] [CrossRef]
- Iida, H.; Doiguchi, M.; Yamashita, H.; Sugimachi, S.; Ichinose, J.; Mori, T.; Shibata, Y. Spermatid-specific expression of Iba1, an ionized calcium binding adapter molecule-1, in rat testis. Biol. Reprod. 2001, 64, 1138–1146. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, Y.; Ohsawa, K.; Kanazawa, H.; Kohsaka, S.; Imai, Y. Iba1 is an actin-cross-linking protein in macrophages/microglia. Biochem. Biophys. Res. Commun. 2001, 286, 292–297. [Google Scholar] [CrossRef]
- Ohsawa, K.; Imai, Y.; Kanazawa, H.; Sasaki, Y.; Kohsaka, S. Involvement of Iba1 in membrane ruffling and phagocytosis of macrophages/microglia. J. Cell Sci. 2000, 113, 3073–3084. [Google Scholar]
- Wilson, M.J.; Jeyasuria, P.; Parker, K.L.; Koopman, P. The transcription factors steroidogenic factor-1 and SOX9 regulate expression of Vanin-1 during mouse testis development. J. Biol. Chem. 2005, 280, 5917–5923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toyooka, Y.; Tsunekawa, N.; Takahashi, Y.; Matsui, Y.; Satoh, M.; Noce, T. Expression and intracellular localization of mouse Vasa-homologue protein during germ cell development. Mech. Dev. 2000, 93, 139–149. [Google Scholar] [CrossRef]
- Nassar, A.; Salim, Y.; Eid, K.; Shaheen, H.M.; Saati, A.A.; Hetta, H.F.; Elmistekawy, A.; Batiha, G.E. Ameliorative effects of honey, propolis, pollen, and royal jelly mixture against chronic toxicity of sumithion insecticide in white albino rats. Molecules 2020, 25, 2633. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds and all data generated and/or analyzed during the present study are available from the corresponding author upon reasonable request. |












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Gene Description | Accession Number | Sequences (5′ > 3′) |
|---|---|---|---|
| IBP | Inhibin subunit beta B (IBP), mRNA | NM_080771.1 | F: GGGTCCGCCTGTACTTCTTC R: CACCTTGACCCGTACCTTCC |
| Ar | Androgen receptor (Ar), mRNA | NM_012502.1 | F: CAGGGACCACGTTTTACCCA R:TTTCCGGAGACGACACGATG |
| Caspase-3 | Cysteine aspartate specific protease-3 | NM_001284409.1 | F: AGTTGGACCCACCTTGTGAG R: AGTCTGCAGCTCCTCCACAT |
| BCl-2 | B-cell lymphoma-2 like protein 4 | NM_007527.3 | F: CACCAGCTCTGAACAGATCATGA R: TCAGCCCATCTTCTTCCAGATGGT |
| IL-1β | Interleukin 1 beta, mRNA | NM_031512.2 | F: CAGGATGAGGACCCAAGCAC R: GTCGTCATCATCCCACGAGT |
| NOS | Nitric oxide synthase 2 | NM_012611.3 | F: GGAGAAAACCCCAGGTGCTATT R: TCGATGGAGTCACATGCAGC |
| Score | Definition and Criteria |
|---|---|
| 10 | Complete spermatogenesis with many mature spermatozoa |
| 9 | Many spermatozoa, with a disorganized germinal epithelium that showed sloughing into lumen |
| 8 | Presence of few spermatozoa (<5 to 10/seminiferous tubule) |
| 7 | Absence of spermatozoa, but many spermatids are present |
| 6 | Absence of spermatozoa, with few spermatids (<5/seminiferous tubule) |
| 5 | Absence of spermatozoa, and spermatid, with the presence of several spermatocytes |
| 4 | Absence of spermatozoa, and spermatid, with the presence of few spermatocytes (<5/seminiferous tubule) |
| 3 | Spermatogonia are the only cell present |
| 2 | Sertoli cells only present with absence of germ cells |
| 1 | No cells visualized in the tubular section |
| Antibody | Source | Dilution | Antigen Retrieval | Heating Condition |
|---|---|---|---|---|
| Rabbit polyclonal anti-active caspase 3 | (#9662, Cell Signaling Technology, Danvers, MA, USA) | 1:300 | 10 mM citrate buffer (pH 6.0) | 105 °C, 20 min |
| Rabbit polyclonal anti-iNOS | (ab15323, Abcam, Cambridge, UK) | 1:100 | 10 mM citrate buffer (pH 6.0) | 105 °C, 20 min |
| Rabbit polyclonal anti-Iba1 | (019-19741, Wako Osaka, Japan) | 1:1200 | 10 mM citrate buffer (pH 6.0) | 105 °C, 20 min |
| Rabbit polyclonal anti-MVH | (ab13840, Abcam, Cambridge, UK) | 1:200 | 10 mM citrate buffer (pH 6.0) | 105 °C, 20 min |
| Rabbit polyclonal anti-Sox9 | (AB5535, Merck Millipore, Burlington, MA, USA) | 1:1000 | 10 mM citrate buffer (pH 6.0) | 105 °C, 20 min |
| Goat polyclonal anti-IL1β | (AB-401-NA, R&D Systems, Minneapolis, MN, USA) | 1:200 | 10 mM citrate buffer (pH 6.0) | 105 °C, 20 min |
| Goat polyclonal anti-PCNA | (sc-9857, Santa Cruz Biotechnology, Santa Cruz, CA, USA) | 1:2000 | Dako, 105 °C, 20 min | 105 °C, 20 min |
| Rabbit polyclonal anti-inhibin alpha | (CM 171 A, Biocare Medical, Concord, CA, USA) | 1:100 | 10 mM citrate buffer (pH 6.0) | 105 °C, 20 min |
| Group | Pancreatic Tissue | Serum | |||
|---|---|---|---|---|---|
| MDA (nmol/g) | NO (µmol/g) | SOD (U/g) | GPx (U/g) | Testosterone (ng/mL) | |
| CTR | 0.32 ± 0.03 c | 43.07 ± 1.03 b | 7.33 ± 0.28 b | 29.29 ± 1.02 ab | 7.51 + 0.27 a |
| FIP | 0.68 ± 0.03 a | 79.31 ± 0.89 a | 4.16 ± 0.08 c | 15.30 ± 0.47 c | 2.92 + 0.15 c |
| CeNPs | 0.36 ± 0.02 c | 32.32 ± 1.51 c | 7.57 ± 0.40 ab | 27.55 ± 0.59 b | 6.92 + 0.15 ab |
| FIP+CeNPs | 0.44 ± 0.001 b | 29.34 ± 0.56 c | 8.12 ± 0.23 a | 36.05 ± 2.24 a | 6.44 + 0.23 b |
| Sig p-value | 0.001 | 0.031 | 0.002 | 0.002 | 0.027 |
| Intra-Assay | 3.15 | 3.45 | 2.50 | 5.36 | 6.53 |
| Inter-Assay | 8.32 | 9.54 | 7.91 | 8.74 | 10.12 |
| Group | Casp3 | BCL-2 | IL-1β | NOS | Ar | IBP |
|---|---|---|---|---|---|---|
| CTR | 1.00 ± 0.25 b | 1.00 ± 0.25 b | 1.00 ± 0.25 a | 1.00 ± 0.25 b | 1.00 ± 0.25 a | 1.00 ± 0.25 b |
| FIP | 1.8 ± 0.45 a | 0.04 ± 0.01 c | 1.53 ± 0.38 a | 1.50 ± 0.37 b | 0.36 ± 0.09 b | 0.28 ± 0.07 c |
| CeNPs | 0.78 ± 0.19 c | 1.25 ± 0.31 ab | 0.25 ± 0.06 b | 3.09 ± 0.77 a | 1.37 ± 0.34 a | 1.60 ± 0.40 ab |
| FIP+CeNPs | 0.08 ± 0.02 d | 1.90 ± 0.30 a | 0.01 ± 0.002 c | 0.019 ± 0.005 c | 1.21 ± 0.30 a | 1.96 ± 0.49 a |
| Sig p value | 0.002 | 0.001 | 0.005 | 0.003 | 0.027 | 0.038 |
| Groups | Johnsen’s Score |
|---|---|
| CTR | 9.01 ± 0.54 b |
| FIP | 2.1 ± 0.29 a |
| CeNPs | 9.21 ± 0.73 b |
| FIP+CeNPs | 6.96 ± 0.24 b |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saleh, H.; Nassar, A.M.K.; Noreldin, A.E.; Samak, D.; Elshony, N.; Wasef, L.; Elewa, Y.H.A.; Hassan, S.M.A.; Saati, A.A.; Hetta, H.F.; et al. Chemo-Protective Potential of Cerium Oxide Nanoparticles against Fipronil-Induced Oxidative Stress, Apoptosis, Inflammation and Reproductive Dysfunction in Male White Albino Rats. Molecules 2020, 25, 3479. https://doi.org/10.3390/molecules25153479
Saleh H, Nassar AMK, Noreldin AE, Samak D, Elshony N, Wasef L, Elewa YHA, Hassan SMA, Saati AA, Hetta HF, et al. Chemo-Protective Potential of Cerium Oxide Nanoparticles against Fipronil-Induced Oxidative Stress, Apoptosis, Inflammation and Reproductive Dysfunction in Male White Albino Rats. Molecules. 2020; 25(15):3479. https://doi.org/10.3390/molecules25153479
Chicago/Turabian StyleSaleh, Hamida, Atef M. K. Nassar, Ahmed E. Noreldin, Dalia Samak, Norhan Elshony, Lamiaa Wasef, Yaser H. A. Elewa, Shaimaa M. A. Hassan, Abdullah A. Saati, Helal F. Hetta, and et al. 2020. "Chemo-Protective Potential of Cerium Oxide Nanoparticles against Fipronil-Induced Oxidative Stress, Apoptosis, Inflammation and Reproductive Dysfunction in Male White Albino Rats" Molecules 25, no. 15: 3479. https://doi.org/10.3390/molecules25153479