
Efficacy of Phytochemicals Derived from Avicennia officinalis for the Management of COVID-19: A Combined In Silico and Biochemical Study

 , ,
, ,  , ,
, ,  and
and 
Abstract
:1. Introduction
2. Results
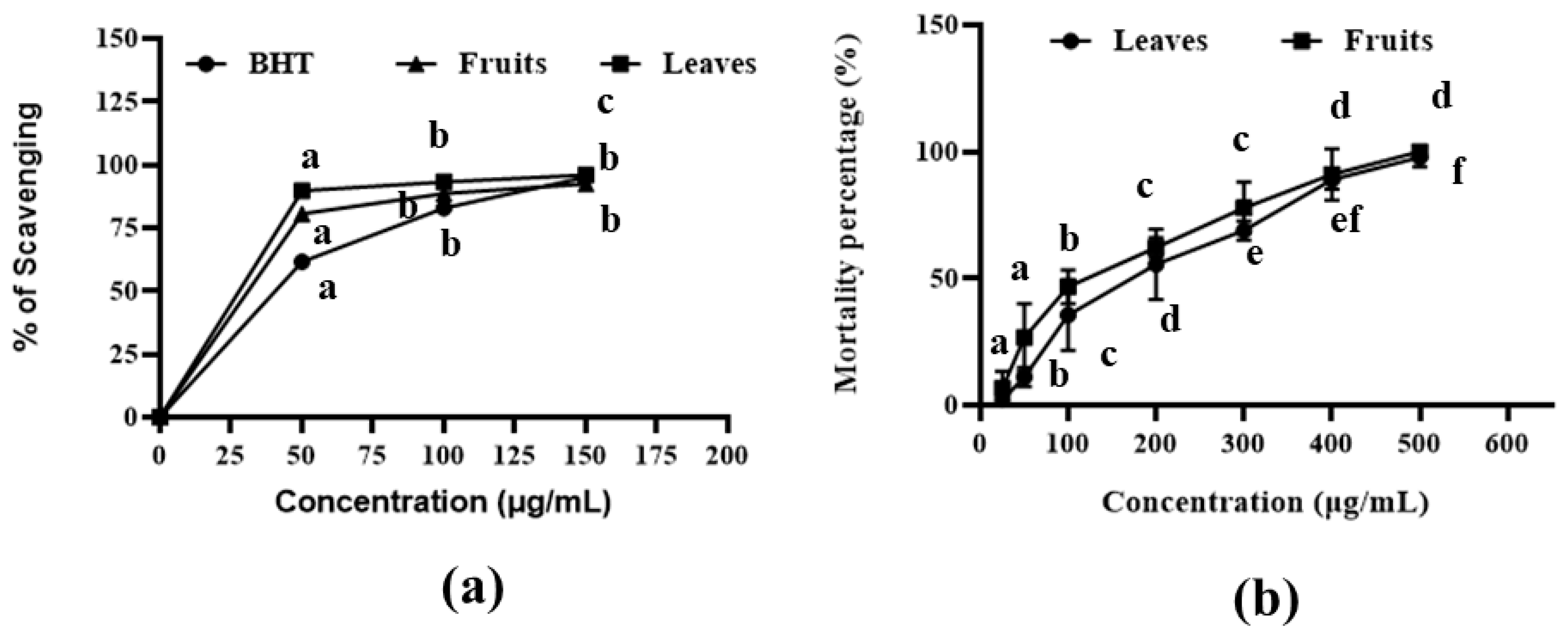
2.1. DPPH Antioxidant Scavenging Assay
2.2. Cytotoxic Assay
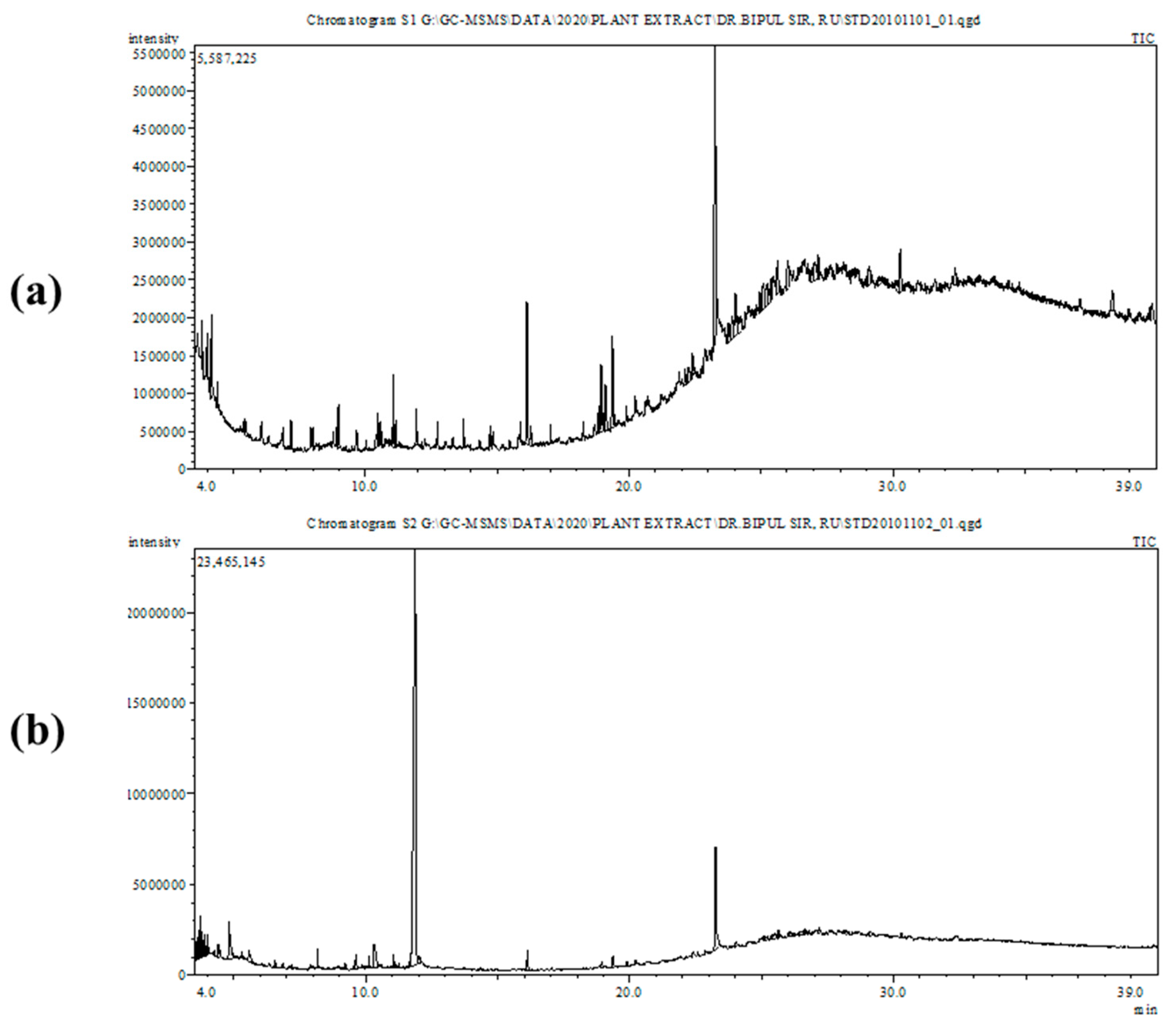
2.3. GC-MS Analysis
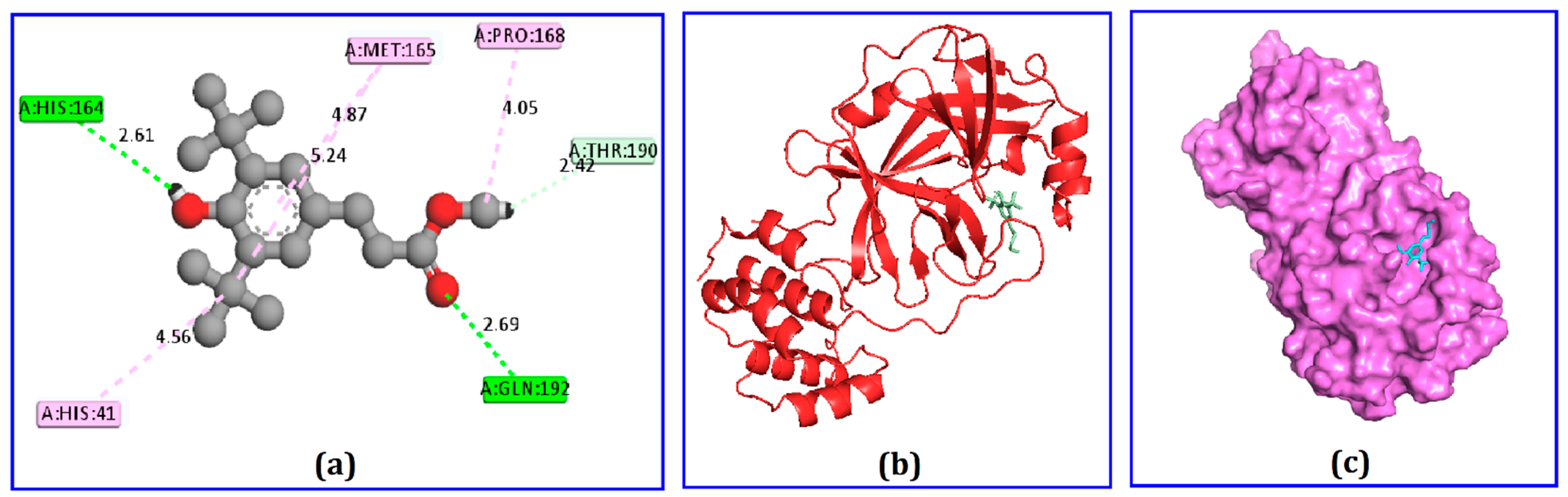
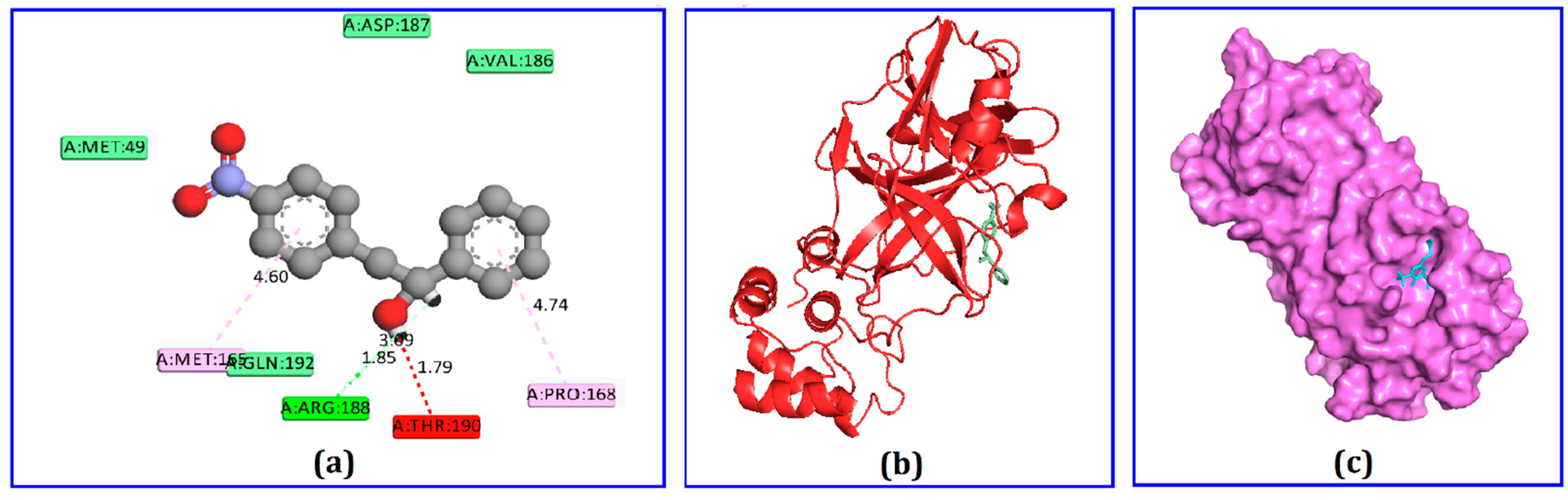
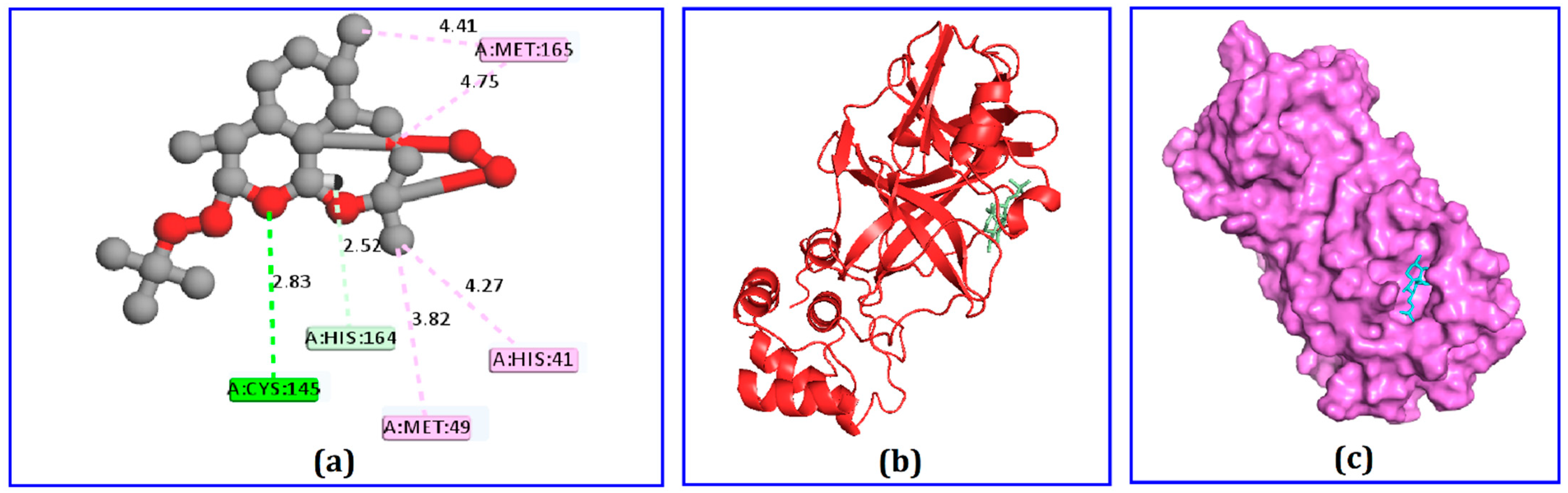
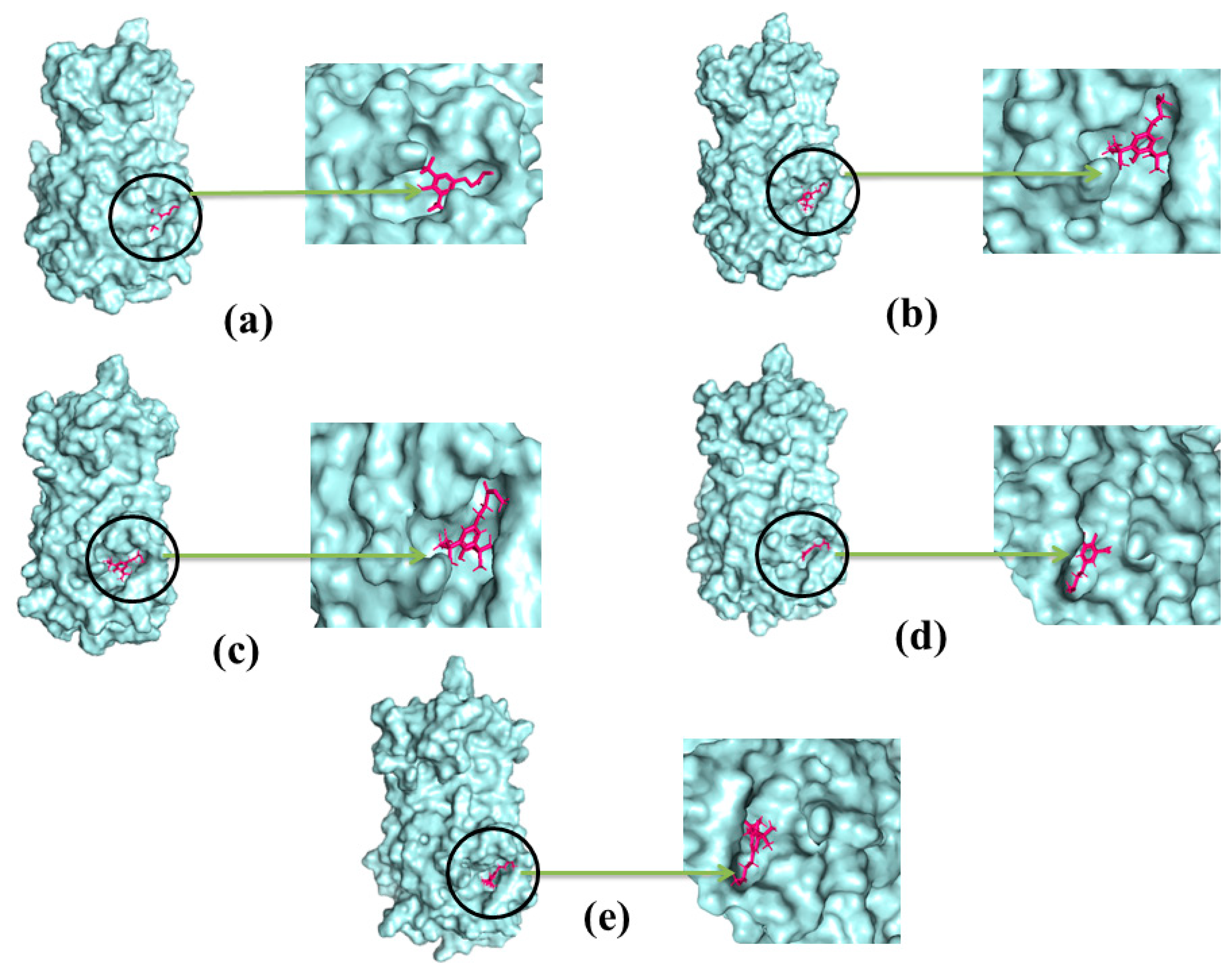
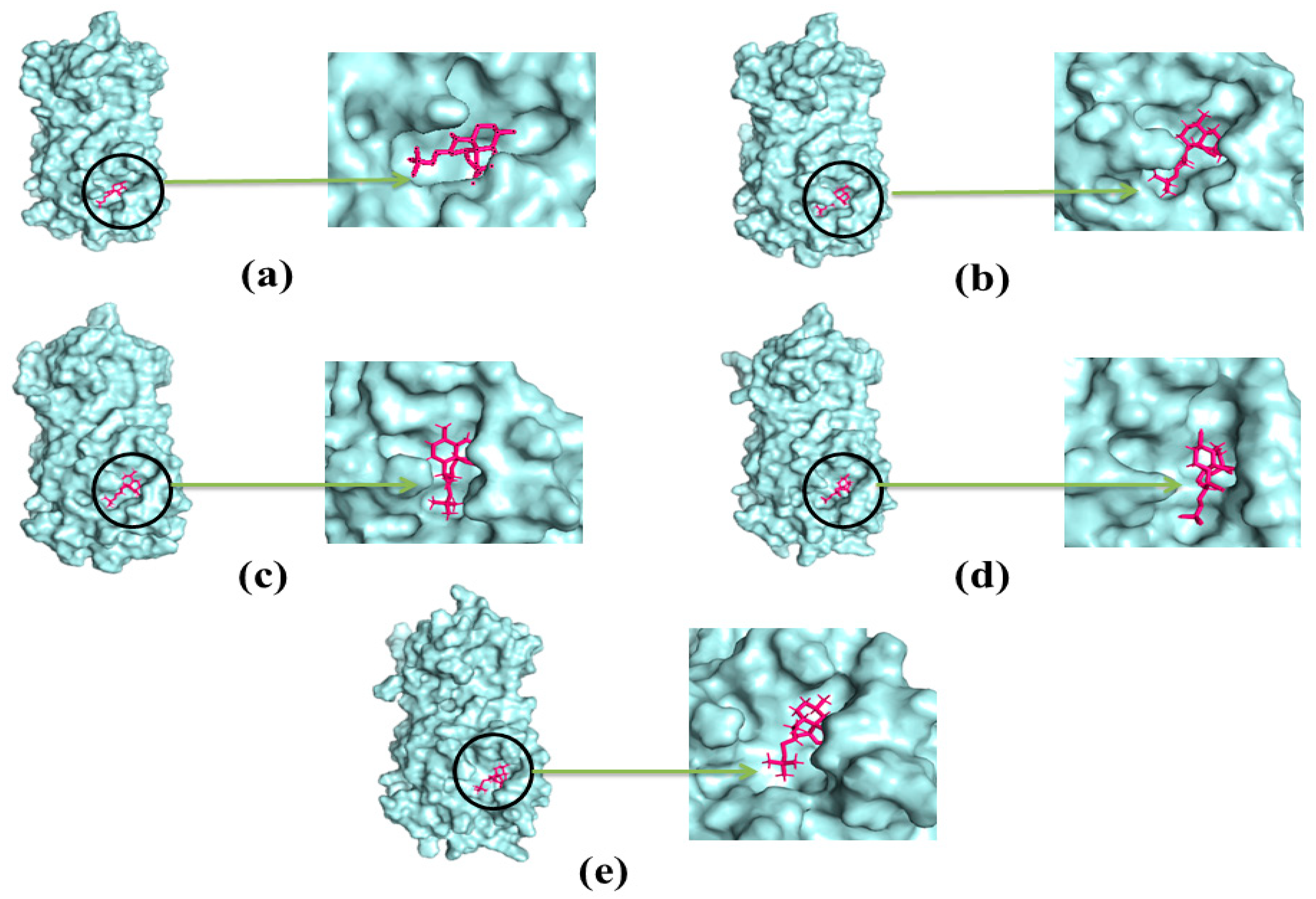
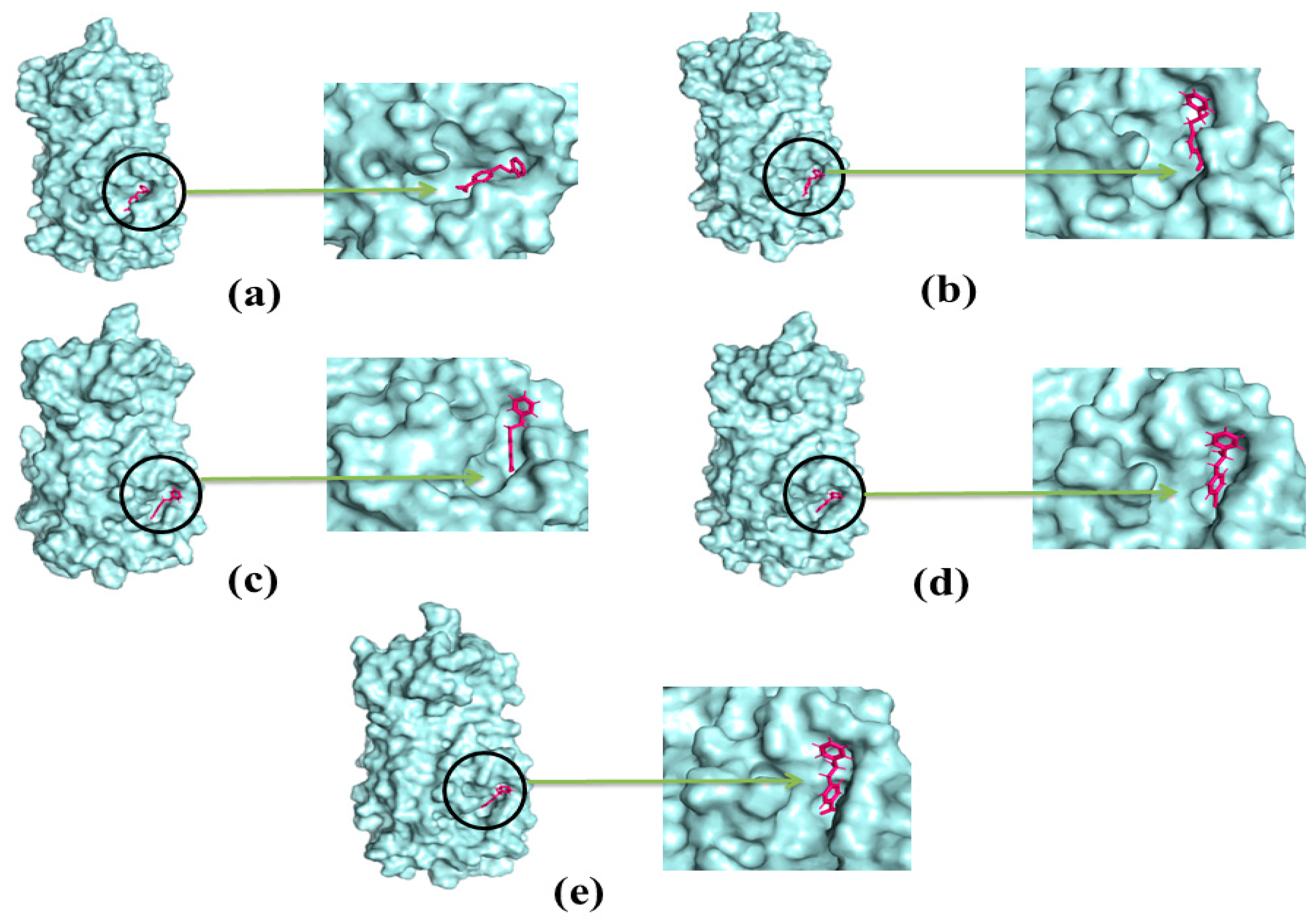
2.4. Docking Analysis
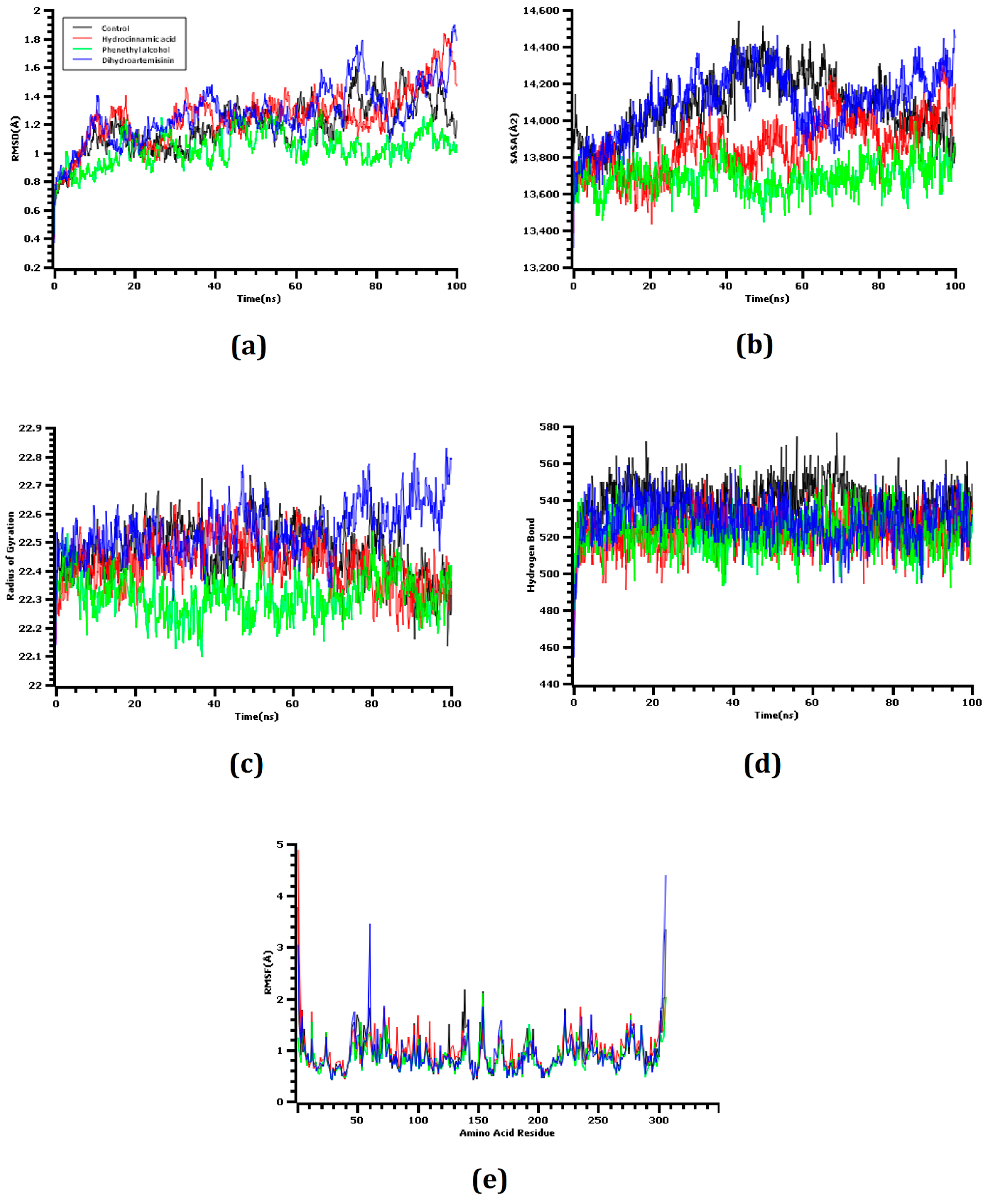
2.5. Molecular Dynamics
2.6. ADMET Analysis
3. Discussion
4. Materials and Methods
4.1. Sample Collection
4.2. Preparation of Plant Extract
4.3. Antioxidant Activity
4.4. Cytotoxicity Assay
4.5. Gas Chromatography-Mass Spectroscopy (GC-MS) Analysis
4.6. Ligand Preparation
4.7. Protein Preparation
4.8. Molecular Docking
4.9. Molecular Dynamics Simulation
4.10. ADMET (Absorption, Distribution, Metabolism, Toxicity) Analysis
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Lau, H.; Khosrawipour, V.; Kocbach, P.; Mikolajczyk, A.; Schubert, J.; Bania, J.; Khosrawipour, T. The positive impact of lockdown in Wuhan on containing the COVID-19 outbreak in China. J. Travel Med. 2020, 27, taaa037. [Google Scholar] [CrossRef] [Green Version]
- Nainu, F.; Abidin, R.S.; Bahar, M.A.; Frediansyah, A.; Emran, T.B.; Rabaan, A.A.; Dhama, K.; Harapnan, H. SARS-CoV-2 reinfection and implications for vaccine development. Hum. Vaccines Immunotherap. 2020, 16, 3061–3073. [Google Scholar] [CrossRef]
- Harapan, H.; Ryan, M.; Yohan, B.; Abidin, R.S.; Nainu, F.; Rakib, A.; Jahan, I.; Emran, T.B.; Ullah, I.; Panta, K.; et al. COVID-19 and dengue: Double punches for dengue-endemic countries in Asia. Rev. Med. Virol. 2021, 31, e2161. [Google Scholar] [CrossRef]
- Zhu, L.; Yang, P.; Zhao, Y.; Zhuang, Z.; Wang, Z.; Song, R.; Zhang, J.; Liu, C.; Gao, Q.; Xu, Q.; et al. Single-Cell Sequencing of Peripheral Mononuclear Cells Reveals Distinct Immune Response Landscapes of COVID-19 and Influenza Patients. Immunity 2020, 53, 685–696. [Google Scholar] [CrossRef]
- Arabi, Y.M.; Murthy, S.; Webb, S. COVID-19: A novel coronavirus and a novel challenge for critical care. Intensive. Care. Med. 2020, 46, 833–836. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Lin, D.; Sun, X.; Curth, U.; Drosten, C.; Sauerhering, L.; Becker, S.; Rox, K.; Hilgenfeld, R. Crystal structure of SARS-CoV-2 main protease provides a basis for design of improved a-ketoamide inhibitors. Science 2020, 368, 409–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tallei, T.E.; Tumilaar, S.G.; Niode, N.J.; Fatimawali; Kepel, B.J.; Idroes, R.; Effendi, Y.; Sakib, S.A.; Emran, T.B. Potential of plant bioactive compounds as SARS-CoV-2 main protease (Mpro) and spike (S) glycoprotein inhibitors: A molecular docking study. Scientifica 2020, 2020, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, M.; Kleine-Weber, H.; Pöhlmann, S.A. Multibasic Cleavage Site in the Spike Protein of SARS-CoV-2 Is Essential for Infection of Human Lung Cells. Mol. Cell. 2020, 78, 779–784. [Google Scholar] [CrossRef] [PubMed]
- Prompetchara, E.; Ketloy, C.; Palaga, T. Immune responses in COVID-19 and potential vaccines: Lessons learned from SARS and MERS epidemic. Asian Pac. J. Allergy Immunol. 2020, 38, 1–9. [Google Scholar] [CrossRef]
- Islam, M.J.; Parves, M.R.; Mahmud, S.; Tithi, F.A.; Reza, M.A. Assessment of structurally and functionally high-risk nsSNPs impacts on human bone morphogenetic protein receptor type IA (BMPR1A) by computational approach. Comput. Biol. Chem. 2019, 80, 31–45. [Google Scholar] [CrossRef]
- Khan, M.A.; Mahmud, S.; Alam, A.S.M.R.U.; Rahman, M.E.; Ahmed, F.; Rahmatullah, M. Comparative molecular investigation of the potential inhibitors against SARS-CoV-2 main protease: A molecular docking study. J. Biomol. Struct. Dyn. 2020, 2020, 1–7. [Google Scholar] [CrossRef]
- Thirunavukkarasu, P.; Ramanathan, T.; Ramkumar, L.; Shanmugapriya, R.; Renugadevi, G. The antioxidant and free radical scavenging effect of Avicennia officinalis. J. Med. Plant Res. 2011, 5, 4754–4758. [Google Scholar] [CrossRef]
- Das, S.K.; Samantaray, D.; Mahapatra, A.; Pal, N.; Munda, R.; Thatoi, H. Pharmacological activities of leaf and bark extracts of a medicinal mangrove plant Avicennia officinalis L. Clin. Phytosci. 2018, 4, 13. [Google Scholar] [CrossRef]
- Bakshi, M.; Ghosh, S.; Chakraborty, D.; Hazra, S.; Chaudhuri, P. Assessment of potentially toxic metal (PTM) pollution in mangrove habitats using biochemical markers: A case study on Avicennia officinalis L. in and around Sundarban, India. Mar. Pollut. Bull. 2018, 133, 157–172. [Google Scholar] [CrossRef]
- Rakib, A.; Paul, A.; Ahmed, S.; Chy, M.N.U.; Sami, S.A.; Baral, S.K.; Majumder, M.; Tareq, A.T.; Amin, M.N.; Shahriar, A.; et al. Biochemical and computational approach of phytocompounds from Tinospora crispa in the management of COVID-19. Molecules 2020, 25, 3936. [Google Scholar] [CrossRef]
- Das, S.K.; Samantaray, D.; Sahoo, S.K.; Patra, J.K.; Samanta, L.; Thatoi, H. Bioactivity guided isolation and structural characterization of the antidiabetic and antioxidant compound from bark extract of Avicennia officinalis L. S. Afr. J. Bot. 2019, 125, 109–115. [Google Scholar] [CrossRef]
- Teodoro, A.J. Bioactive compounds of food: Their role in the prevention and treatment of diseases. Oxid. Med. Cell. Longev. 2019, 2019, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Rege, A.A.; Ambaye, R.Y.; Deshmukh, R.A. In-vitro testing of anti-HIV activity of some medicinal plants. Indian J. Nat. Prod. Resour. 2010, 1, 193–199. [Google Scholar]
- Sartini, S.; Permana, A.D.; Mitra, S.; Tareq, A.M.; Salim, E.; Ahmad, I.; Harapan, H.; Emran, T.B.; Nainu, F. Current State and Promising Opportunities on Pharmaceutical Approaches in the Treatment of Polymicrobial Diseases. Pathogens 2021, 10, 245. [Google Scholar] [CrossRef]
- Duong, N.T.; Vinh, P.D.; Thuong, P.T.; Hoai, N.T.; Thanh, L.N.; Bach, T.T.; Nam, N.H.; Anh, N.H. Xanthine oxidase inhibitors from Archidendron clypearia (Jack.) I.C. Nielsen: Results from systematic screening of Vietnamese medicinal plants. Asian Pac. J. Trop. Med. 2017, 10, 549–556. [Google Scholar] [CrossRef]
- Floris, S.; Fais, A.; Rosa, A.; Piras, A.; Marzouki, H.; Medda, R.; González-Paramás, A.M.; Kumar, A.; Santos-Buelga, C.; Era, B. Phytochemical composition and the cholinesterase and xanthine oxidase inhibitory properties of seed extracts from the: Washingtonia filifera palm fruit. RSC Adv. 2019, 9, 21278–21287. [Google Scholar] [CrossRef] [Green Version]
- Uddin, M.Z.; Paul, A.; Rakib, A.; Sami, S.A.; Mahmud, S.; Rana, M.S.; Hossain, S.; Tareq, A.M.; Dutta, M.; Emran, T.B.; et al. Chemical Profiles and Pharmacological Properties with In Silico Studies on Elatostema papillosum Wedd. Molecules 2021, 26, 809. [Google Scholar] [CrossRef] [PubMed]
- Barua, N.; Aziz, M.A.I.; Tareq, A.M.; Sayeed, M.A.; Alam, N.; ul Alam, N.; Uddin, M.A.; Lyzu, C.; Emran, T.B. In vivo and in vitro evaluation of pharmacological activities of Adenia trilobata (Roxb.). Biochem. Biophys. Rep. 2020, 23, 100772. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.S.; Mahmud, S.; Sultana, R.; Dong, W. Identification and in silico molecular modelling study of newly isolated Bacillus subtilis SI-18 strain against S9 protein of Rhizoctonia solani. Arab. J. Chem. 2020, 13, 8600–8612. [Google Scholar] [CrossRef]
- Rakib, A.; Sami, S.A.; Mimi, N.J.; Chowdhury, M.M.; Eva, T.A.; Nainu, F.; Paul, A.; Shahriar, A.; Tareq, A.M.; Emon, N.U.; et al. Immunoinformatics-guided design of an epitope-based vaccine against severe acute respiratory syndrome coronavirus 2 spike glycoprotein. Comput. Biol. Med. 2020, 124, 103967. [Google Scholar] [CrossRef] [PubMed]
- Mukhtar, M.; Arshad, M.; Ahmad, M.; Pomerantz, R.J.; Wigdahl, B.; Parveen, Z. Antiviral potentials of medicinal plants. Virus Res. 2008, 131, 111–120. [Google Scholar] [CrossRef]
- Palombo, E.A. Traditional medicinal plant extracts and natural products with activity against oral bacteria: Potential application in the prevention and treatment of oral diseases. Evid. Based Complement. Altern. Med. 2011, 2011, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Annan, K.; Sarpong, K.; Asare, C.; Dickson, R.; Amponsah, K.; Gyan, B.; Ofori, M.; Gbedema, S.Y. In vitro anti-plasmodial activity of three herbal remedies for malaria in Ghana: Adenia cissampeloides (Planch.) Harms., Termina liaivorensis A. Chev, and Elaeis guineensis Jacq. Pharmacogn. Res. 2012, 4, 225–229. [Google Scholar] [CrossRef] [Green Version]
- Guha, B.; Arman, M.; Islam, M.N.; Tareq, S.M.; Rahman, M.M.; Sakib, S.A.; Mutsuddy, R.; Tareq, A.M.; Emran, T.B.; Alqahtani, A.M. Unveiling pharmacological studies provide new insights on Mangifera longipes and Quercus gomeziana. Saudi J. Biol. Sci. 2021, 28, 183–190. [Google Scholar] [CrossRef]
- Guevara, A.P.; Vargas, C.; Sakurai, H.; Fujiwara, Y.; Hashimoto, K.; Maoka, T.; Kozuka, M.; Ito, Y.; Tokuda, H.; Nishino, H. An antitumor promoter from Moringa oleifera Lam. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 1999, 440, 181–188. [Google Scholar] [CrossRef]
- Poojary, S. Role of Bioinformatics, Computational Biology and Computer Technologies in Combating COVID-19 Virus-a Review. Int. J. Biotech. Trends Technol. 2020, 10, 26–30. [Google Scholar] [CrossRef]
- Mahmud, S.; Uddin, M.A.R.; Zaman, M.; Sujon, K.M.; Rahman, M.E.; Shehab, M.N.; Islam, A.; Alom, M.W.; Amin, A.; Akash, A.S.; et al. Molecular docking and dynamics study of natural compound for potential inhibition of main protease of SARS-CoV-2. J. Biomol. Struct. Dyn. 2020, 2020, 1–9. [Google Scholar] [CrossRef]
- Bappy, S.S.; Sultana, S.; Adhikari, J.; Mahmud, S.; Khan, M.A.; Kibria, K.M.K.; Rahman, M.M.; Shibly, A.Z. Extensive immunoinformatics study for the prediction of novel peptide-based epitope vaccine with docking confirmation against envelope protein of Chikungunya virus: A computational biology approach. J. Biomol. Struct. Dyn. 2021, 39, 1139–1154. [Google Scholar] [CrossRef]
- Ghosh, R.; Chakraborty, A.; Biswas, A.; Chowdhuri, S. Evaluation of green tea polyphenols as novel corona virus (SARS CoV-2) main protease (Mpro) inhibitors–an in silico docking and molecular dynamics simulation study. J. Biomol. Struct. Dyn. 2020, 2020, 1–13. [Google Scholar] [CrossRef]
- Rakib, A.; Sami, S.A.; Islam, M.A.; Ahmed, S.; Faiz, F.B.; Khanam, B.H.; Marma, K.K.S.; Rahman, M.; Uddin, M.M.N.; Nainu, F.; et al. Epitope-Based Immunoinformatics Approach on Nucleocapsid Protein of Severe Acute Respiratory Syndrome-Coronavirus-2. Molecules 2020, 25, 5088. [Google Scholar] [CrossRef]
- Chowdhury, K.H.; Chowdhury, M.R.; Mahmud, S.; Tareq, A.M.; Hanif, N.B.; Banu, N.; Reza, A.S.M.A.; Emran, T.B.; Simal-Gandara, J. Drug Repurposing Approach against Novel Coronavirus Disease (COVID-19) through Virtual Screening Targeting SARS-CoV-2 Main Protease. Biology 2020, 10, 2. [Google Scholar] [CrossRef]
- Ashfaq, U.A.; Mumtaz, A.; ul Qamar, T.; Fatima, T. MAPS Database: Medicinal plant Activities, Phytochemical and Structural Database. Bioinformation 2013, 9, 993–995. [Google Scholar] [CrossRef] [PubMed]
- Rakib, A.; Nain, Z.; Islam, M.A.; Sami, S.A.; Mahmud, S.; Islam, A.; Ahmed, S.; Siddiqui, A.B.F.; Babu, S.M.O.F.; Hossain, P.; et al. A molecular modelling approach for identifying antiviral selenium-containing heterocyclic compounds that inhibit the main protease of SARS-CoV-2: An in silico investigation. Brief. Bioinform. 2021, 22, 1476–1498. [Google Scholar] [CrossRef]
- Mahmud, S.; Uddin, M.A.R.; Paul, G.K.; Shimu, M.S.S.; Islam, S.; Rahman, E.; Islam, A.; Islam, M.S.; Promi, M.M.; Emran, T.B.; et al. Virtual screening and molecular dynamics simulation study of plant derived compounds to identify potential inhibitor of main protease from SARS-CoV-2. Brief. Bioinform. 2021, 22, 1402–1414. [Google Scholar] [CrossRef]
- Thuy, B.T.P.; My, T.T.A.; Hai, N.T.T.; Hieu, L.T.; Hoa, T.T.; Thi Phuong Loan, H.; Triet, N.T.; Anh, T.T.V.; Quy, P.T.; Tat, P.V.; et al. Investigation into SARS-CoV-2 Resistance of Compounds in Garlic Essential Oil. ACS Omega 2020, 5, 8312–8320. [Google Scholar] [CrossRef]
- Kar, P.; Sharma, N.R.; Singh, B.; Sen, A.; Roy, A. Natural compounds from Clerodendrum spp. as possible therapeutic candidates against SARS-CoV-2: An in silico investigation. J. Biomol. Struct. Dyn. 2020, 2020, 1–12. [Google Scholar] [CrossRef]
- Ying, T.C.; Ibrahim, Z.; Abd Rahman, M.B.; Tejo, B.A. Structure-based design of peptide inhibitors for protein arginine deiminase type iv (PAD4). Encycl. Bioinform. Comput. Biol. ABC Bioinform. 2018, 1, 729–740. [Google Scholar] [CrossRef]
- Mahmud, S.; Parves, M.R.; Riza, Y.M.; Sujon, K.M.; Ray, S.; Tithi, F.A.; Zaoti, Z.F.; Alam, S.; Absar, N. Exploring the potent inhibitors and binding modes of phospholipase A2 through in silico investigation. J. Biomol. Struct. Dyn. 2020, 38, 4221–4231. [Google Scholar] [CrossRef]
- Cob-Calan, N.N.; Chi-Uluac, L.A.; Ortiz-Chi, F.; Cerqueda-García, D.; Navarrete-Vázquez, G.; Ruiz-Sánchez, E.; Hernández-Núñez, E. Molecular docking and dynamics simulation of protein β-tubulin and antifungal cyclic lipopeptides. Molecules 2019, 24, 3387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, J.; Cheng, F.; Xu, Y.; Li, W.; Tang, Y. Estimation of ADME properties with substructure pattern recognition. J. Chem. Inf. Model. 2010, 50, 1034–1041. [Google Scholar] [CrossRef]
- Sanguinetti, M.C.; Tristani-Firouzi, M. hERG potassium channels and cardiac arrhythmia. Nature 2006, 440, 463–469. [Google Scholar] [CrossRef]
- Banerjee, T.; Valacchi, G.; Ziboh, V.A. Inhibition of TNFα expression by amentoflavone through suppression κB activation in A549 cells of NF-κB. Mol. Cell. Biochem. 2002, 95616, 105–110. [Google Scholar] [CrossRef] [PubMed]
- Mariswamy, Y.; Gnaraj, W.E.; Johnson, M. Chromatographic finger print analysis of steroids in Aerva lanata L by HPTLC technique. Asian Pac. J. Trop. Biomed. 2011, 1, 428–433. [Google Scholar] [CrossRef] [Green Version]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Jahan, I.; Tona, M.R.; Sharmin, S.; Sayeed, M.A.; Tania, F.Z.; Paul, A.; Chy, M.; Uddin, N.; Rakib, A.; Emran, T.B. GC-MS phytochemical profiling, pharmacological properties, and in silico studies of Chukrasia velutina leaves: A novel source for bioactive agents. Molecules 2020, 25, 3536. [Google Scholar] [CrossRef]
- Achakzai, J.K.; Anwar Panezai, M.; Kakar, M.A.; Kakar, A.M.; Kakar, S.; Khan, J.; Khan, N.Y.; Khilji, I.; Tareen, A.K. In Vitro Anticancer MCF-7, Anti-Inflammatory, and Brine Shrimp Lethality Assay (BSLA) and GC-MS Analysis of Whole Plant Butanol Fraction of Rheum ribes (WBFRR). Biomed Res. Int. 2019, 2019, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Obaidullah, A.J.; Alanazia, M.A.; Alsaifa, N.A.; Mahdi, W.A.; Fantoukh, O.I.; Tareq, A.M.; Sami, S.A.; Alqahtani, A.M.; Emran, T.B. Deeper Insights on Cnesmone javanica Blume Leaves Extract: Chemical Profiles, Biological Attributes, Network Pharmacology and Molecular Docking. Plants 2021, 10, 728. [Google Scholar] [CrossRef]
- Kim, S.; Thiessen, P.A.; Bolton, E.E.; Chen, J.; Fu, G.; Gindulyte, A.; Han, L.; He, J.; He, S.; Shoemaker, B.A. PubChem substance and compound databases. Nucleic Acids Res. 2016, 44, D1202–D1213. [Google Scholar] [CrossRef]
- Hanwell, M.D.; Curtis, D.E.; Lonie, D.C.; Vandermeerschd, T.; Zurek, E.; Hutchison, G.R. Avogadro: An advanced semantic chemical editor, visualization, and analysis platform. J. Cheminform. 2012, 4, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halgren, T.A. Merck Molecular Force Field. J. Comput. Chem. 1996, 17, 490–519. [Google Scholar] [CrossRef]
- Protein Data Bank RCSB PDB: Homepage. RCSB PDB. Available online: https://www.rcsb.org/ (accessed on 1 January 2000).
- Chemistry Collection: Basic Chemistry User Guide, Pipeline Pilot; Accelrys Software Inc.: San Diego, CA, USA, 2011.
- Krieger, E.; Vriend, G. YASARA View–molecular graphics for all devices from smartphones to workstations. Bioinformatics 2014, 30, 2981–2982. [Google Scholar] [CrossRef] [Green Version]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [Green Version]
- Huey, R.; Morris, G.M.; Forli, S. Using AutoDock 4 and AutoDock Vina with AutoDockTools: A Tutorial. Scripps Res. Inst. Mol. 2012, 16, 2785–2791. [Google Scholar] [CrossRef] [Green Version]
- Tumilaar, S.G.; Siampa, J.P.; Fatimawali; Kepel, B.J.; Niode, N.J.; Idroes, R.; Rakib, A.; Emran, T.B.; Tallei, T.E. Potential of leaf extract of Pangium edule Reinw as HIV-1 protease inhibitor: A computational biology approach. J. Appl. Pharm. Sci. 2021, 11, 101–110. [Google Scholar] [CrossRef]
- DeLano, W.L. The PyMOL Molecular Graphics System, Version 1.1. 2002. Available online: https://pymol.org/2/ (accessed on 19 January 2017).
- Krieger, E.; Elmar, G.V.; Spronk, C. YASARA–Yet Another Scientific Artificial Reality Application. 2013. Available online: http://www.yasara.org/ (accessed on 18 August 2004).
- Case, D.A.; Cheatham, T.E.; Darden, T.; Gohlke, H.; Luo, R.; Merz, K.M.; Onufriev, A.; Simmerling, C.; Wang, B.; Woods, R.J. The Amber biomolecular simulation programs. J. Comput. Chem. 2005, 26, 1668–1688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krieger, E.; Vriend, G. New ways to boost molecular dynamics simulations. J. Comput. Chem. 2015, 36, 996–1007. [Google Scholar] [CrossRef]
- Krieger, E.; Nielsen, J.E.; Spronk, C.A.E.M.; Vriend, G. Fast empirical pKa prediction by Ewald summation. J. Mol. Graph. Model. 2006, 4, 481–486. [Google Scholar] [CrossRef]
- Swargiary, A.; Mahmud, S.; Saleh, M.A. Screening of phytochemicals as potent inhibitor of 3-chymotrypsin and papain-like proteases of SARS-CoV2: An in silico approach to combat COVID-19. J. Biomol. Struct. Dyn. 2020, 2020, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Lou, C.; Sun, L.; Li, J.; Cai, Y.; Wang, Z.; Li, W.; Liu, G.; Tang, Y. AdmetSAR 2.0: Web-service for prediction and optimization of chemical ADMET properties. Bioinformatics 2019, 35, 1067–1069. [Google Scholar] [CrossRef]
- Cheng, F.; Li, W.; Zhou, Y.; Shen, J.; Wu, Z.; Liu, G.; Lee, P.W.; Tang, Y. AdmetSAR: A comprehensive source and free tool for assessment of chemical ADMET properties. J. Chem. Inf. Model. 2012, 52, 3099–3105. [Google Scholar] [CrossRef] [PubMed]
- Pires, D.E.V.; Blundell, T.L.; Ascher, D.B. pkCSM: Predicting small-molecule pharmacokinetic and toxicity properties using graph-based signatures. J. Med. Chem. 2015, 58, 4066–4072. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hit Compounds | Docking Scores (Kcal/mol) |
|---|---|
| Hydrocinnamic acid | −7.5 |
| Phenethyl alcohol | −7.3 |
| Dihydroartemisinin | −7.0 |
| Compound | Amino Acid | Bond Type | Distance (Å) |
|---|---|---|---|
| Hydrocinnamic acid | His164 Gln192 Thr190 Pro168 Met165 His41 | H H H A A PA | 2.60 2.69 2.41 4.04 5.23 4.55 |
| Phenethyl alcohol | Arg188 Arg187 Val186 Thr190 Gln192 Met49 Met165 Pro168 | H H H H H H PA PA | 2.18 2.01 2.76 2.52 2.33 2.48 4.47 4.58 |
| Dihydroartemisinin | Cys145 His164 Met165 Met49 His41 | H H A A PA | 2.82 2.51 4.75 4.40 4.26 |
| Complex | Residue | Bond Type | Distance (Å) |
|---|---|---|---|
| Hydrocinnamic acid | Gln192 Gln189 Ala191 Pro168 Met165 | H H H A A | 2.81 2.33 2.30 4.35 5.31 |
| Phenethyl alcohol | Glu166 Thr190 Asp187 Leu167 Pro168 Met165 | H H H PA PA A | 1.87 2.59 2.92 4.89 4.25 4.52 |
| Dihydroartemisinin | His41 Gln189 Met165 Leu167 Pro168 Cys44 Met49 | H H A A PA PA PA | 2.07 2.12 4.55 4.77 4.95 4.17 4.44 |
| Properties | Hydrocinnamic Acid | Phenethyl Alcohol | Dihydroartemisinin |
|---|---|---|---|
| Molecular Weight | 292.41 | 243.46 | 356.45 |
| LogP | 4.09 | 2.87 | 3.941 |
| Surface Area | 127.28 | 104.66 | 149.825 |
| Blood Brain Barrier | 0.8341 (+) | 0.8805 (+) | 0.9342 (+) |
| Human Intestinal Absorption | 0.9346 (+) | 0.9803 (+) | 0.9352 (+) |
| P-Glycoprotein Inhibitor | 0.8815 (-) | 0.89978 (-) | 0.8302 (-) |
| AMES Toxicity | 0.8612 (-) | 0.6173 (-) | 0.6536 (-) |
| HERG Inhibition | 0.9582 (WI) | 0.6933 (WI) | 0.9563 (WI) |
| Hepatotoxicity | No | No | No |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mahmud, S.; Paul, G.K.; Afroze, M.; Islam, S.; Gupt, S.B.R.; Razu, M.H.; Biswas, S.; Zaman, S.; Uddin, M.S.; Khan, M.; et al. Efficacy of Phytochemicals Derived from Avicennia officinalis for the Management of COVID-19: A Combined In Silico and Biochemical Study. Molecules 2021, 26, 2210. https://doi.org/10.3390/molecules26082210
Mahmud S, Paul GK, Afroze M, Islam S, Gupt SBR, Razu MH, Biswas S, Zaman S, Uddin MS, Khan M, et al. Efficacy of Phytochemicals Derived from Avicennia officinalis for the Management of COVID-19: A Combined In Silico and Biochemical Study. Molecules. 2021; 26(8):2210. https://doi.org/10.3390/molecules26082210
Chicago/Turabian StyleMahmud, Shafi, Gobindo Kumar Paul, Mirola Afroze, Shirmin Islam, Swagota Briti Ray Gupt, Mamudul Hasan Razu, Suvro Biswas, Shahriar Zaman, Md. Salah Uddin, Mala Khan, and et al. 2021. "Efficacy of Phytochemicals Derived from Avicennia officinalis for the Management of COVID-19: A Combined In Silico and Biochemical Study" Molecules 26, no. 8: 2210. https://doi.org/10.3390/molecules26082210