Long Non-Coding RNAs in the Regulation of Gene Expression: Physiology and Disease
by
 ,
,
Juliane C. R. Fernandes
 ,
,
Stephanie M. Acuña
, 
Juliana I. Aoki
,
Lucile M. Floeter-Winter
and
Sandra M. Muxel
*
Laboratório de Fisiologia de Tripanossomatídeos, Instituto de Biociências, Departamento de Fisiologia, Universidade de São Paulo, São Paulo 05508-090, Brazil
*
Author to whom correspondence should be addressed.
Non-Coding RNA 2019, 5(1), 17; https://doi.org/10.3390/ncrna5010017
Submission received: 20 December 2018
/
Revised: 11 February 2019
/
Accepted: 12 February 2019
/
Published: 17 February 2019
(This article belongs to the Special Issue Non-Coding RNAs: Variations and Disease)
Abstract
:The identification of RNAs that are not translated into proteins was an important breakthrough, defining the diversity of molecules involved in eukaryotic regulation of gene expression. These non-coding RNAs can be divided into two main classes according to their length: short non-coding RNAs, such as microRNAs (miRNAs), and long non-coding RNAs (lncRNAs). The lncRNAs in association with other molecules can coordinate several physiological processes and their dysfunction may impact in several pathologies, including cancer and infectious diseases. They can control the flux of genetic information, such as chromosome structure modulation, transcription, splicing, messenger RNA (mRNA) stability, mRNA availability, and post-translational modifications. Long non-coding RNAs present interaction domains for DNA, mRNAs, miRNAs, and proteins, depending on both sequence and secondary structure. The advent of new generation sequencing has provided evidences of putative lncRNAs existence; however, the analysis of transcriptomes for their functional characterization remains a challenge. Here, we review some important aspects of lncRNA biology, focusing on their role as regulatory elements in gene expression modulation during physiological and disease processes, with implications in host and pathogens physiology, and their role in immune response modulation.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
The discrepancy of about 20,000 protein-coding genes and over 100,000 different transcripts identified in mammalian transcriptomes highlights the possibility of discovering a novel class of non-translated RNAs [1], beyond those already identified in the 1970s, as part of the translation machinery: ribosomal RNAs (rRNAs) [2] and transfer RNAs (tRNAs) [3]. The advent of next generation sequencing is providing considerable amounts of new information about genomic organization and gene expression regulation [4,5]. These data have been helping to revise our understanding of the current genome and gene annotations [5]. The transcriptome data from cellular lineages and human tissue samples showed that at least 60% of the genome is expressed as primary or processed transcripts, much more than previously predicted [6]. This analysis revealed that thousands of unannotated RNAs may act as non-coding regulatory elements in gene expression or originate as small RNAs [6].
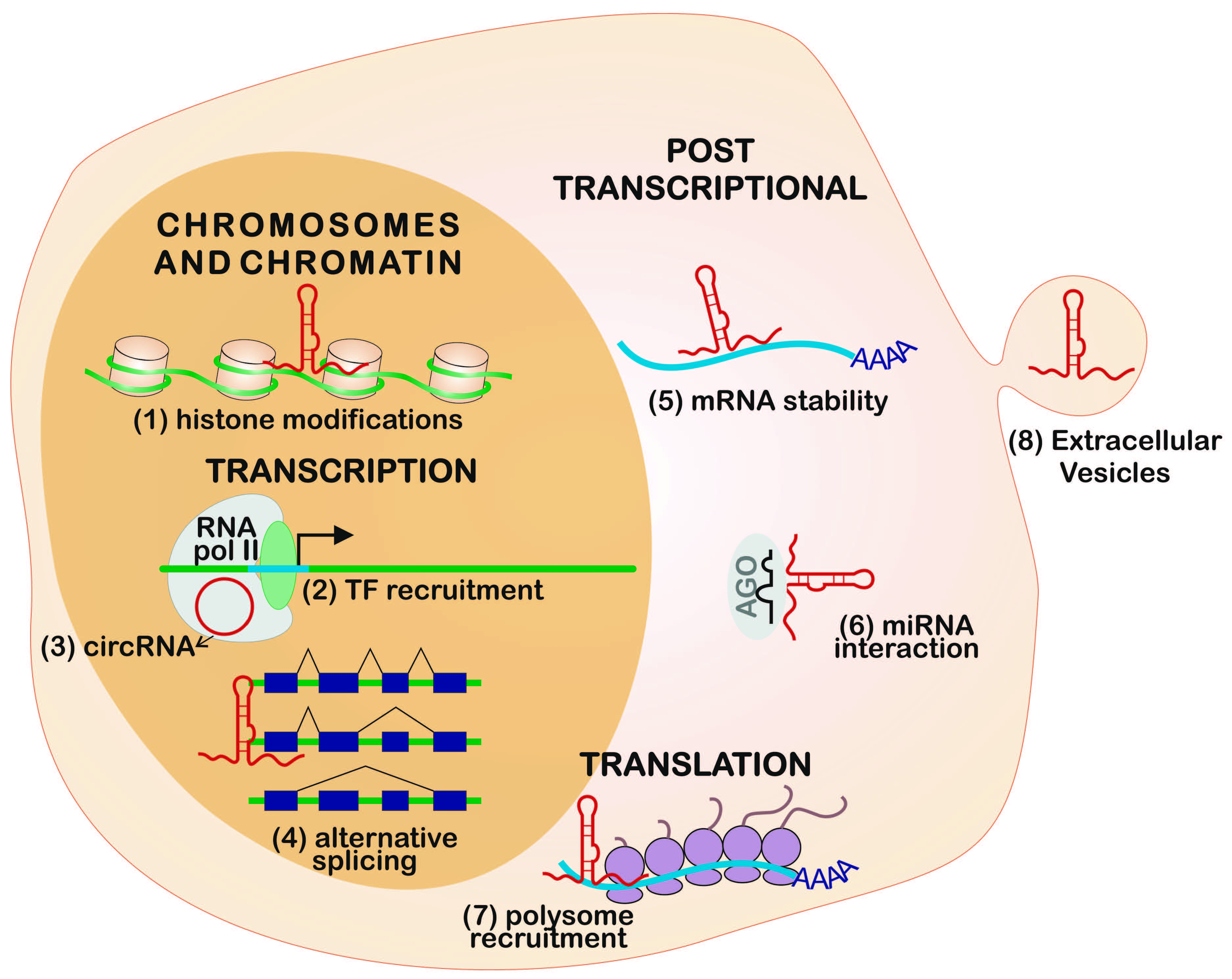
Non-coding RNAs (ncRNAs) can be grouped according to their length, localization, and/or function: long non-coding RNAs (lncRNAs), microRNAs (miRNAs), small interfering RNAs (siRNAs), small nucleolar RNAs (snoRNAs), small nuclear RNAs (snRNAs), and PIWI-interacting RNAs (piRNAs) [7,8]. The lncRNAs are larger than 200 nucleotides and can be subdivided according to their biogenesis loci: intergenic lncRNAs (lincRNAs) [9], intronic lncRNAs, antisense lncRNAs (aslncRNA or natural antisense transcripts, NATs) [10], bidirectional lncRNAs, and enhancer RNAs (eRNAs) (Figure 1A).
The lncRNA biogenesis is mostly similar to messenger RNA (mRNA), since this process is also mediated through RNA polymerase II. In addition, lncRNAs can appear with or without polyadenylation [11,12], alternative cleavage, alternative polyadenylation, and alternative splicing [13], leading to different isoforms from the same locus [11,14].
The lncRNAs genes share features with coding-genes promoters and can be regulated by some transcription factors (TFs), such as p53, nuclear factor-kappa B (NF-κB), Sox2, and Oct4 [15]. Some lncRNAs present unique requirements for their proper expression and functionality, which can be mediated by cMyc TF and Dicer, a key enzyme involved in miRNA processing [16]. The antisense lncRNAs originate from the complementary strand of protein-coding genes [10,17]; about 30% of human annotated genes have an antisense component that highly impacts gene profile expression [18]. Divergent transcription can occur when the RNA polymerase II is recruited in the antisense strand at an upstream site of the protein coding gene promoter, but only a fraction of them generate functional transcripts, called bidirectional lncRNAs [17,19,20]. Evaluation of lncRNAs in human and murine embryonic stem cells showed that over 60% were divergently transcribed and the changes are coordinated with cognate protein coding genes during differentiation [21]. Cap analysis of gene expression (CAGE) showed that eRNAs are bidirectionally transcribed and capped [22], but non-polyadenylated and non-spliced, depending on the integrator complex for the 3’ end cleavage of the transcript [23].
Circular RNAs (circRNAs) were predicted to be the most abundant isoform compared to the linear transcripts [24]. The circRNAs can be formed via regular splicing (intronic circRNAs) [25] and via non-canonical splicing, joining the splice donor at an upstream acceptor site (backsplicing) (Figure 1B) [24]. Some isoforms of circRNAs can be derived from circularization of an intronic fragment with their neighbor exons, forming the exon-intron-circRNA (elciRNAs), which can associate with RNA polymerase II and with U1 snRNA, increasing the transcription of their parental genes in cis [26].
The highly intricate mechanisms to regulates lncRNA degradation leads to specific expression in different cellular types and seems to be subjected to the same mechanisms correlated with mRNA decapping [27], or alternatively through nonsense-mediated decay (NMD) [28], a process that depends on ribosome recruiting and that will be described further in this review for the translational potential of lncRNAs.
Expression of lncRNAs can be detected from uni- to pluri-cellular eukaryotic organisms, although the processing and mechanisms of action can differ [29]. The most recent NONCODE database source points to over 100,000 lncRNAs in the human genome, but this number seems to be underestimated [1,30,31].
Cellular and temporal specificity drives the mechanism of action of lncRNAs and their simultaneous impact in diverse target genes [32,33,34]. Long non-coding RNA can regulates neighbor protein-coding genes expression and thus contribute to the mRNA and protein content in the cell [32,35]. For instance, in the analysis of T lymphocytes transcriptomes, 1500 lncRNAs were identified, showing that 50% appeared to be exclusively expressed in this cell population, whereas only 7% of the mRNAs are stage- or lineage-specific, suggesting lncRNAs as strong components in cell identity [36].
Similar to protein domains, lncRNAs can interact with nucleic acids or proteins through base-pairing or structural recognition, respectively, so a single lncRNA molecule is able to interact with diverse macromolecules [8]. This interaction is mediated by ribonucleoprotein complexes in which lncRNAs are associated with proteins [8]. A 100-ribonucleotide hairpin chain can interact simultaneously with more proteins than peptide domains of 100 amino acids interacting with other proteins. This can be indicative of possible evolutive advantages in regulation mediated by RNAs [37].
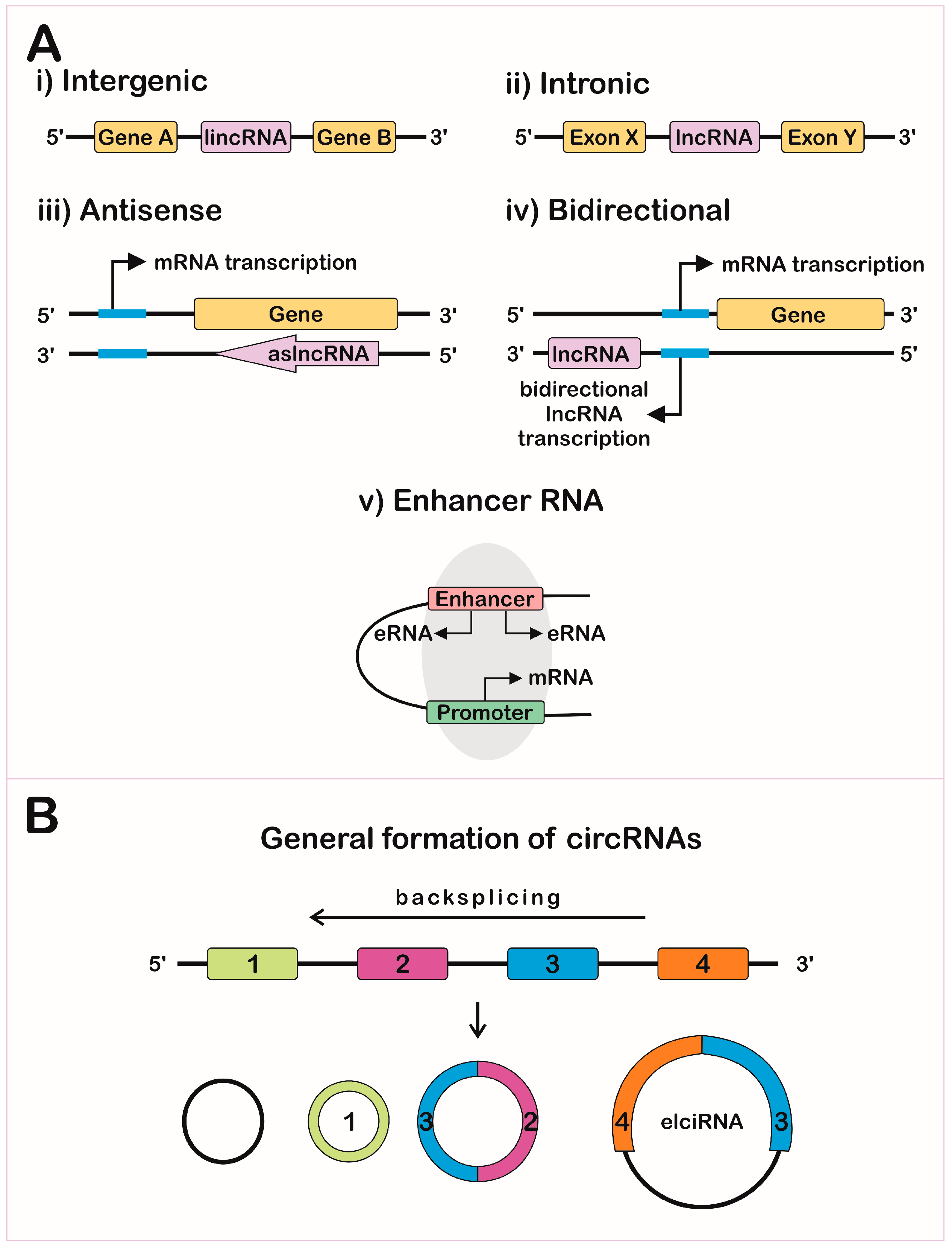
Long non-coding RNAs are found within the nucleus, nucleolus, cytoplasm, and even in the mitochondria [38,39], and its localization is a good indicator of their mode of action [1], as shown in Figure 2. The RNA-Seq obtained from these compartments showed a strong enrichment of lncRNAs in the nucleus, specifically associated with chromatin in several cell lines when compared to mRNAs [5]. Recent studies estimated that more than half of the expressed lncRNAs are in the cytoplasm, associated with polysome fractions, controlling stability, and translation of mRNAs [40]. Altogether, these data indicate that lncRNA localization depends on the motifs signatures: protein signal-peptides, nuclear-restricted lincRNA BMP/OP-responsive gene (BORG) [41], and Alu-related sequences in a more generally-spread nuclear retention mechanism [42].
Apart from lncRNA localization, secondary and tertiary structures of lncRNA are also important for their mechanism of action. In 2010, a group used a high-throughput approach describing the whole transcriptome structure of Saccharomyces cerevisiae at nucleotide resolution [43]. These conformations have been unveiled using diverse techniques, such as fragmentation sequencing (FragSeq), which is based on sequencing of fragments digested by single- or double-strand specific nucleases [44], which can be useful in the description of RNA molecular structure and in the identification of folding domains that mediate interaction with other macromolecules. This is also important in the understanding of lncRNA evolution once there is a low level of primary sequence conservation [45,46], but also conservation at the stem-loop structure level, maintaining the functionality of these molecules [47].
The lncRNA structural changes can also regulate the availability of recognition sites for RNA binding proteins through thermodynamic adjustments in the hairpin stability [48,49]. The most common RNA chemical modifications are the exchange of adenosine to inosine, catalyzed by adenosine deaminases, and the reversible modifications by N6-methyl-adenosine (m6A) methylation [50]. Besides regulating function, these modifications are essential for the recognition of the RNAs as endogenous and non-pathogenic molecules, whereas non-modified RNAs are capable of stimulating the immune response mediated by toll-like receptors (TLRs) [31]. Some tools can predict sites responsible for editing and the impact on structure and function, such as interaction with miRNAs [51].
2. Gene Expression Regulation Levels
The regulation of gene expression in eukaryotes is complex and compartmentalized [52]. It can occur in multiple steps, such as in the chromatin organization, transcription machinery recruitment, mRNA processing and its delivery to the cytoplasm, mRNA half-life, translation, and posttranslational processes, which can be interfered with by lncRNAs [31,53], as represented in Figure 2. These molecules can also be secreted within extracellular vesicles, modulating the gene expression in its environment [54].
2.1. Chromosome and Chromatin Structure
The idea that RNA can be a chromatin-associated structural component was corroborated by the description that, there, the amount of RNA is twice as high as the DNA associated with the chromatin structure [55]. Many studies identified several types of RNAs related to this function, such as snRNAs, and lncRNAs, such as the X inactivation-specific transcription (XIST), AIR, and H19, were associated with heterochromatin formation and imprinting [8]. Additionally, lncRNAs that are expressed only in embryonic stem cells interact directly with the chromatin, then modulate gene expression and the maintenance of pluripotency [56]. The lncRNA interaction with DNA can occur by sequence complementarity to a single-stranded fragment of DNA or allocation in the helix [31]. Additionally, eRNAs can execute their function by mediating chromosomal looping together with the mediator complex [57]. As such, lncRNAs are related to a general structuration of the genome, organizing nuclear architecture, and consequently, gene expression [58], as shown in Figure 2.
2.2. Transcription
At the transcriptional level, the promoter region of a lncRNA sequence, regardless of its synthesis, can act as an enhancer, characterizing a cis regulation [59]. The NAT asOct4-pg5 can indirectly regulate epigenetic markers through the RNA/DNA binding protein PURA (purine-rich element binding factor A), which reduces transcription from the protein-coding sense transcripts and simultaneously represses other NATs in a negative-feedback loop [60]. Some ncRNAs can interact directly with the transcription machinery, as shown by circRNAs that directly interact with RNA pol II, according to crosslink followed by immunoprecipitation assays (Figure 2) [31]. Additionally, eRNAs can bind to transcription factors, positioning them in specific promoters [61]. Other lncRNAs can regulate transcription, controlling DNA methyltransferases recruitment, TFs, zinc-finger proteins, and others transcription regulators [1].
2.3. Post-Transcriptional Regulation
2.3.1. Long Non-Coding RNA and MicroRNA Interplay
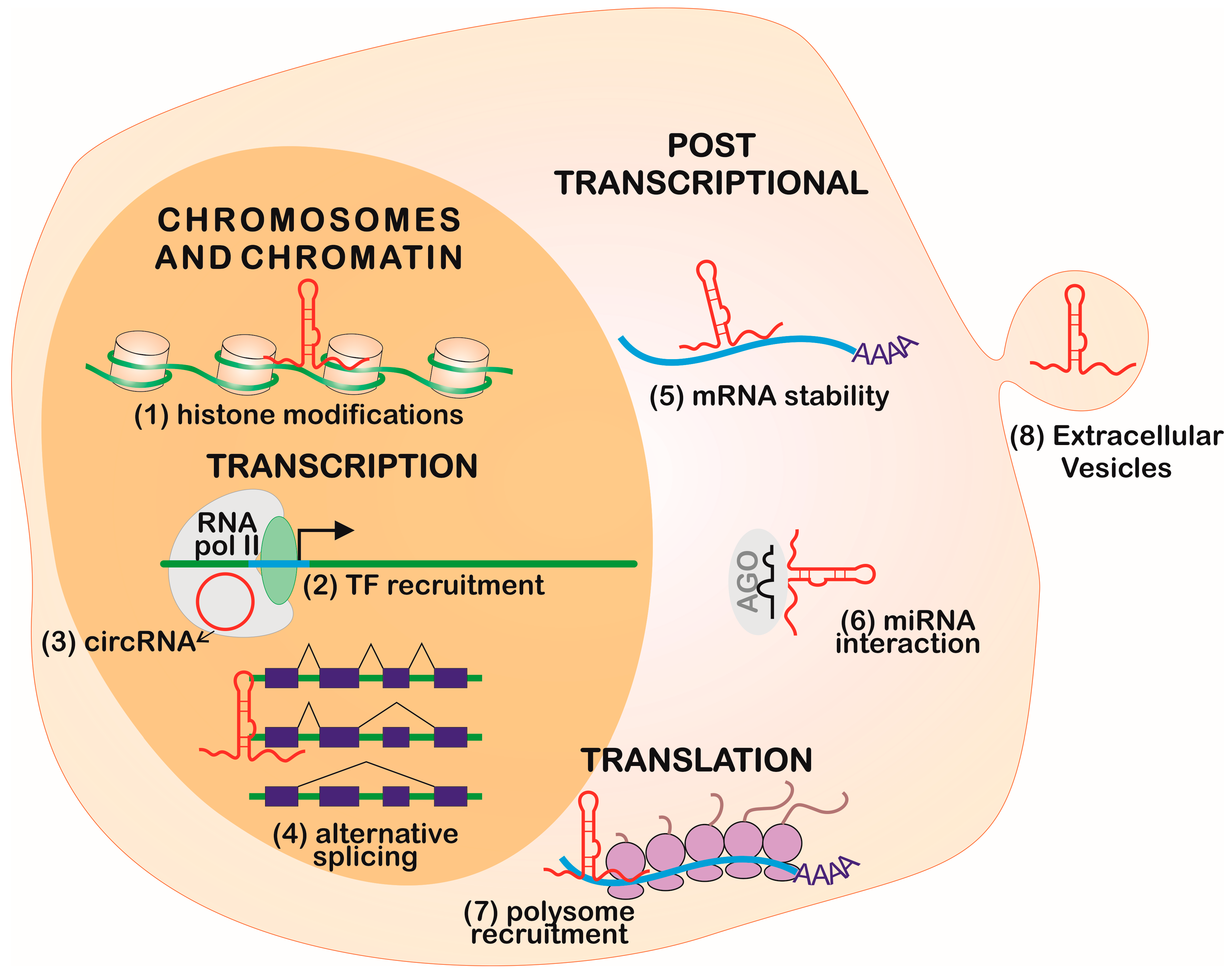
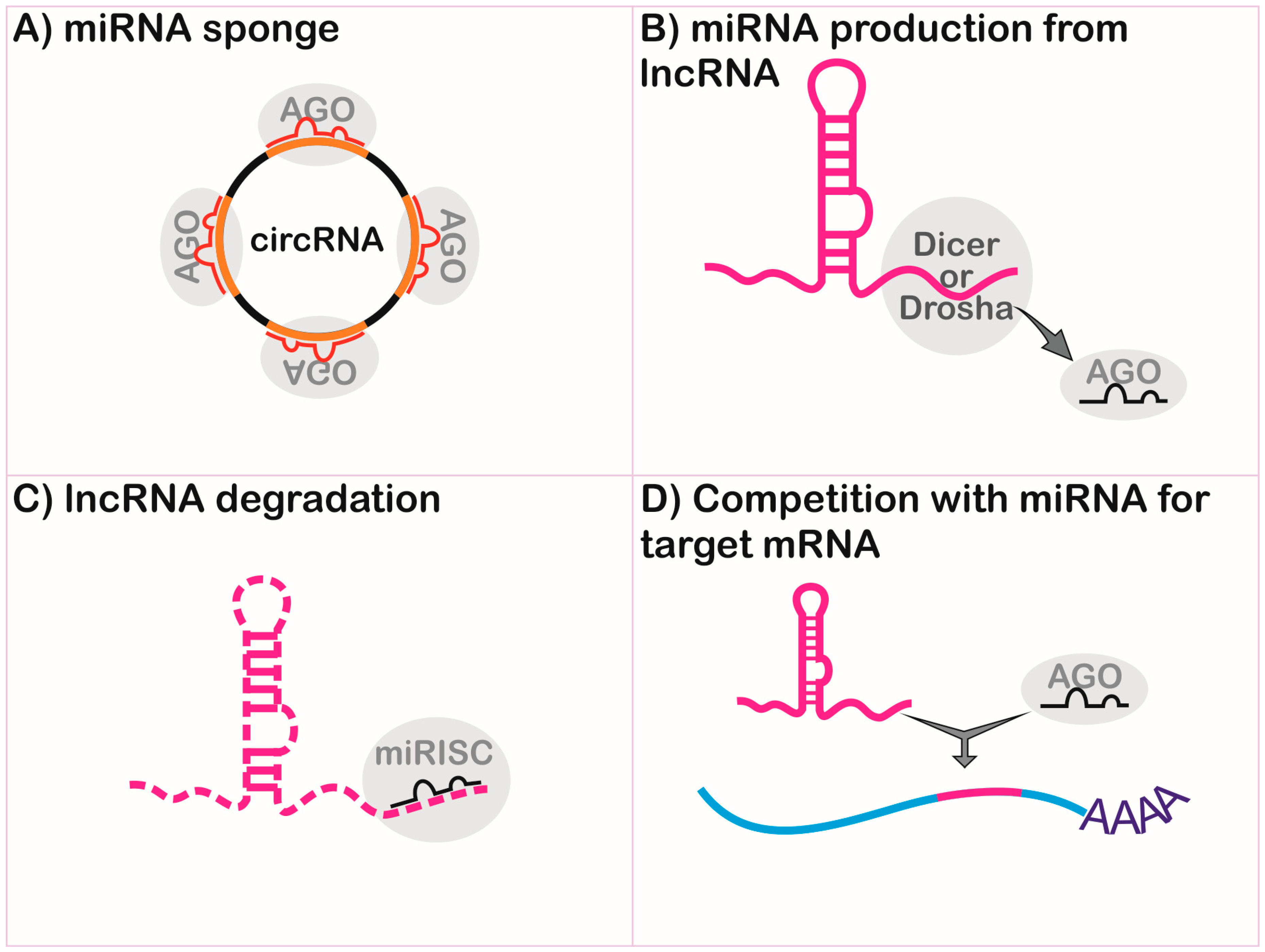
Different classes of ncRNAs can interact through sequence complementarity by executing coordinated functions. The most remarkable interplay occurs between lncRNAs and miRNAs in the regulation of gene expression (Figure 2 and Figure 3).
Long non-coding RNAs can be endogenous competitors RNAs (ceRNAs), also called miRNA sponges, by presenting binding sequences for miRNAs [62,63], and can impair the functional interaction of miRNA and mRNA by interference in the gene regulation, as shown in Figure 3A. Both linear and circular isoforms can exert this function [62]. The first description of competition between these molecules involved the naturally-expressed circular RNA sponge for miR-7 (ciRS-7), via miRNA-dependent binding to argonaute (AGO) proteins [62]. During myogenesis, the linear transcript named lnc-MG competes with miR-125b, controlling insulin-like growth factor 2 (IGF-2) levels [64]. In hepatitis C virus infection, lncRNA-ATB is activated by transforming growth factor-beta (TGF-β) and competes with miR-425-5p, upregulating the TGF-β type II receptor (TGF-βRII) and SMAD family member 2 (SMAD2), promoting liver fibrosis [65].
Long non-coding RNAs can be precursors of miRNAs (Figure 3B) and can regulate different points of miRNA biogenesis, acting on microprocessor activity to finish the primary transcript in a mechanism independent of polyadenylation [66]. For example, the lncRNA named LOC554202 originates miR-31 and is important in preventing metastasis in breast cancer [67]. The lncRNA deleted in lymphocytic leukemia 2 (DLEU2) harbors miR-15a/16.1 within its third intron and expression dysfunction is associated with lymphocytic leukemia [68,69]. The lncRNA colon cancer-associated transcript 2 (CCAT2) blocks the maturation of miR-145 by inhibition of Dicer cleavage and cytoplasm export [70]. The miR-221 and miR-222 are co-transcribed with lnc-Ang362, an angiotensin II-upregulated lncRNA in endothelial cells [71].
MicroRNAs can regulate the stability and half-life of lncRNA (Figure 3C) [72]. The miR-152 is a negative regulator of the XIST lncRNA in a repression feedback loop in glioblastoma cells [73]. Both miR-101 and miR-217 negatively regulate metastasis associated with lung adenocarcinoma transcript 1 (MALAT1) in carcinoma cells [74], and miR-449a inhibits the expression of nuclear enriched abundant transcript 1 (NEAT1) lncRNA in lung cancer [75].
Long non-coding RNAs can compete with miRNAs for the target site of mRNA (Figure 3D). This is demonstrated by antisense transcript for the β-site amyloid precursor protein cleaving enzyme 1 (BACE1-AS), which binds to its sense partner in the miR-485-5p recognition site, thus impairing its function, which is implicated in Alzheimer’s disease [76].
Some lncRNAs can exert multiple functions, as described for lncRNA H19, which acts as a molecular sponge for let-7 [77] and is a precursor of miR-675 [78], mediating muscular differentiation and regeneration [77,78]. The lnc-MD1 sequestrates both miR-133 and miR-135, allowing mastermind-like-1 and myocyte-specific enhancer factor 2C expression during myogenesis [79]. miRNA clusters can comprise functional lncRNA, such as the increased levels observed for the miR-99a, let7c, and miR-125b-2 cluster in leukemic cells lines, which comprise a lncRNA gene MIR99AHG in the intronic region of the hsa21 gene and the miR-125b-1, let-7a-2, and miR-100 cluster, forming MIR100HG lncRNA within the hsa11 locus [80].
2.3.2. Alternative Splicing
More than half of intron-containing genes in animals and plants can be alternatively spliced, generating different mature mRNA isoforms [81]. Diverse ncRNAs are involved in this process, such as the well-described uridine-rich small nuclear RNAs (U snRNAs) that act as a ribonucleoprotein complex in all splicing processes [82]. Recently, the lncRNAs NEAT1 and MALAT1 were observed colocalizing with nuclear speckles containing the splicing factor SC35 [83]. Also, MALAT1 regulates phosphorylation and activation of serine/arginine (SR) splicing factors [84]. From these observations, lncRNAs are described as interacting with splicing factors, composing duplexes of pre-mRNAs and antisense lncRNAs or chromatin remodeling, which can directly influence the transcriptional rate by RNA polymerase II and modulate splicing [85,86]. An example of this mechanism is described in different human cell lines in which the first apoptosis signal (Fas)-antisense lncRNA-SAF interacts with the human splicing factor SPF45, leading to exclusion of exon 6 during splicing of Fas and production of a soluble Fas protein, thus preventing Fas-FasL-mediated apoptosis [87].
2.3.3. Messenger RNA Stability
Long non-codin RNAs have properties capable of mediating the nonsense-mediated mRNA decay (NMD) pathway, in a miRNA-independent way. This function is exemplified by the complex formed by lncRNA half-STAU1-binding site RNAs (1/2-sbsRNAs) with the target mRNA, creating a double-stranded (ds) transactivation motif that interacts with the STAU1-dsRNA binding protein, leading to mRNA degradation [88]. Some NATs can also decrease the stability of the protein-coding sense transcript [89,90]. Alternatively, some aslncRNAs can increase pair stability, as observed for β-secretase-1 in Alzheimer’s disease [91]. A study demonstrated a tumor suppression role for the PDCD4 antisense transcript, as it stabilizes its cognate mRNA by forming a duplex structure that regulates the association with mRNA decay factors [92].
2.4. Translation
Translation processes can be facilitated or repressed by lncRNAs, as shown by the dopaminergic neuron-specific expression of Uchl1 (ubiquitin carboxy-terminal hydrolase L1), which is regulated by its antisense transcript (AS Uchl1), which recruits polysomes by the repetitive domain SINEB2 to promote a cap-independent translation [93]. lncRNA-p21 was demonstrated to repress translation of JunB and β-catenin mRNAs by recruiting translational repressors [94,95].
3. Physiological Conditions and Disease
The roles of lncRNAs in genome integrity and gene expression have demonstrated the relevance of these molecules for physiological and pathological conditions [96,97,98]. Here, we review the involvement of lncRNAs in chromosomal compensation, imprinting, chronic diseases, immune response process, and in some pathogens.
3.1. X Chromosome Dosage Compensation
A classic example of chromatin structure regulation mediated by lncRNA, described in the 1990s, is the dosage compensation of the X chromosome in females during embryogenesis, since one of them is inactivated equalizing the dosage as in XY males [99,100]. XIST is expressed exclusively from the inactivated X chromosome (Xi), producing a lncRNA that binds directly to polycomb repressive complexes 1 and 2 (PRC1 and PRC2, respectively) [76]. This mechanism leads to the formation of heterochromatin with the recruitment of histone methylases of the H3K27me3 type [101]. Additionally, a repressive nuclear environment is formed by the depletion of TFs, RNA polymerase II, splicing factors, and nascent RNAs, mainly in the pericentromeric region [102]. The specificity and recognition motifs of XIST to the Xi chromosome have been extensively studied, but it was not possible to determine consensus domains, suggesting that these lncRNAs can act through multiple interactions [103]. Colocalization studies demonstrated ligation mainly in repetitive regions of the X chromosome, which are not coding genes, and long interspaced elements (LINEs), abundant in the X chromosome [104]. Ectopic expression of XIST in autosomal chromosomes can lead to heterochromatin formation [105]. The female heterochromatinization of the X chromosome is complex and depends on nuclear architecture, epigenetic modifications, and non-coding RNAs [106].
In Drosophila melanogaster, the dosage compensation is performed via male X chromosome hyperactivation through histone acetylation [107,108,109]. This mechanism is also mediated by lncRNAs, the roX1 and roX2 that play redundant functions and act together with a complex of five proteins exclusive to males, and are essential for survival [110,111].
3.2. Imprinting
In mammals, epigenetic markers can regulate maternal or paternal chromosomes expression; this phenomenon of exclusive or differential expression is called imprinting and can be mediated by lncRNAs. The genes Air (antisense Igf2r RNA) and Kcnq1ot1 (lincRNA from Kcnq1 gene) are transcribed from the paternal chromosome and recruit G9a methylase for histone modification silencing exclusively in the maternal genes [112]. The regulation can occur in cis via Air binding to the promoter region of the paternal Igf2r gene, silencing its expression by inhibiting the recruitment of RNA pol II [113]. Besides the potential for chromatin silencing by lncRNAs, these molecules can act as enhancers, for example, the lncRNA HOTTIP (HOXA distal transcript antisense RNA) interacts directly with components of the histone activation machinery through H3K4me3 modifications [94].
3.3. Chronic Diseases
Besides all physiological mechanisms described, the dysregulation of the expression of lncRNAs can occur during chronic multifactorial diseases [114]. Most studies describe the function of lncRNA in cancer, correlating it with diverse TFs and molecules that regulate the cell cycle, as well as linking it with the processes of pluripotency and differentiation [56]. A larger number of human diseases involve lncRNAs and more than 900 lncRNAs, having an important role in complex diseases, such as cancer and cardiovascular and neurological diseases, according to the experimentally supported data in the lncRNA disease database [115].
Long non-coding RNAs are correlated with different aspects of complex diseases and differential expression was observed in patients with neurodegeneration, such as Alzheimer’s [116], Huntington’s [117], and Parkinson’s [118] diseases; schizophrenia [119,120]; and autism spectrum disorders [121,122]. Dysregulation is frequently described in cardiovascular diseases, such as during chronic heart failure [123], diabetic cardiomyopathy [124], atherosclerosis [125], and infarction [126]. The complexity of lncRNA regulation and specificity are being revealed as important determinants in diabetes mellitus [127].
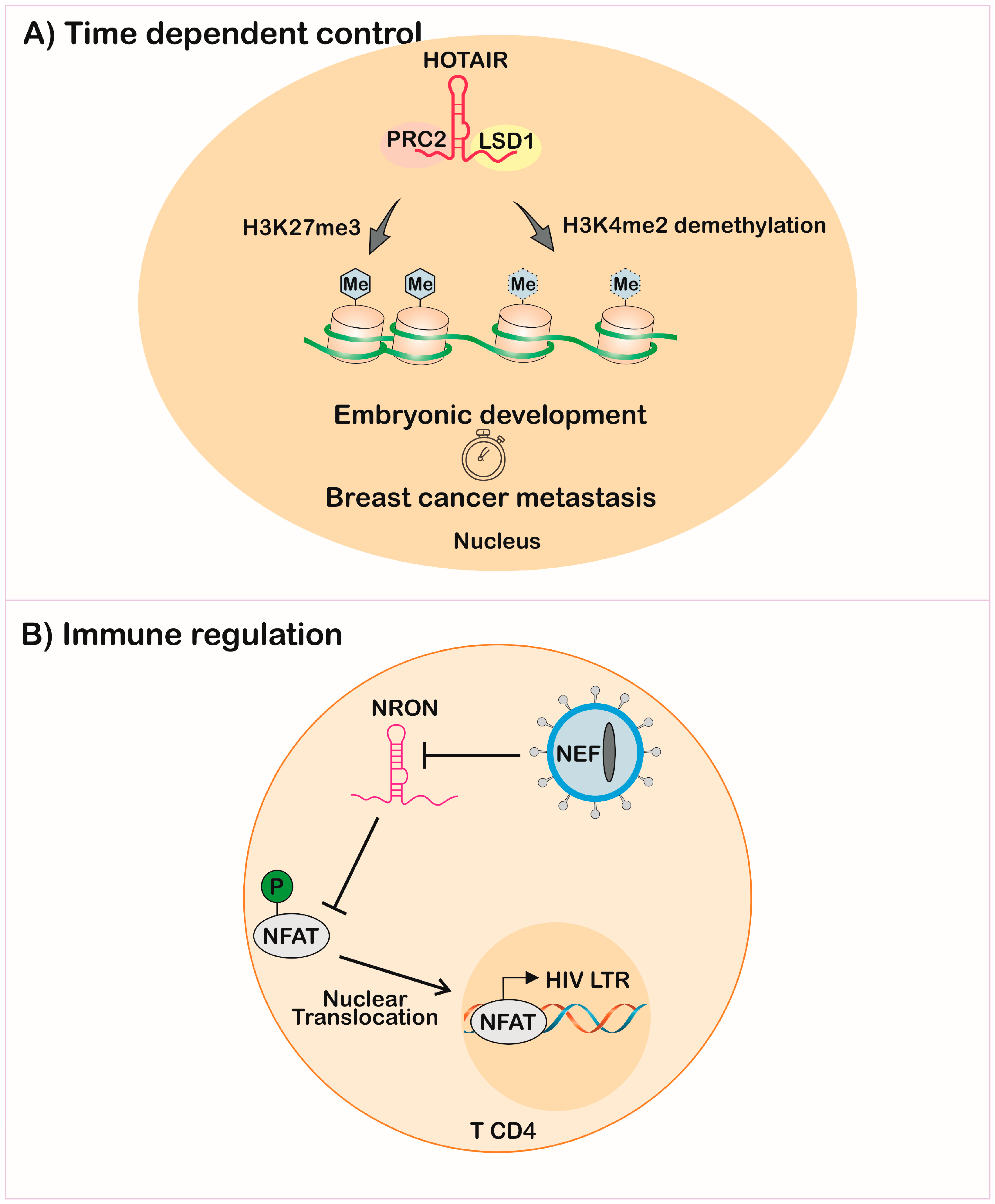
The same innate mechanisms of action of lncRNAs can lead to malignant transformation of the cell if expressed in moments distinct from the physiological ones. In mammals, the lncRNA HOTAIR (Hox transcript antisense intergenic RNA) has two chromatin modifier recruitment domains, ensuring H3K27 methylation by PRC2 and H3K4 demethylation by lysine-specific demethylase 1 (LSD1). This mechanism is important during embryonic development in regulating homeotic genes together with other HoxC cluster components [128,129]. However, the dysregulation of HOTAIR promotes transformation and metastasis in diverse cancer models [122,130] (Figure 4A).
The dysregulation of Foxp3 long intergenic non-coding RNA (FLICR) can decrease FoxP3 levels and; therefore, T regulatory cells (Treg) control, leading to autoimmunity [131], showing that lncRNA controls both pro- and anti-inflammatory processes, as next described for infectious diseases. Other autoimmune diseases may also involve lncRNAs, such as psoriasis [132], rheumatoid arthritis [133], and Chron’s disease. [134]
3.4. Immune Response Against Infectious Diseases
Complex relationships between host and pathogens often use gene expression control of the host cell in the evasion of the immune response. lncRNAs have been related to the fine-tuned regulation of inflammatory processes [135,136]. The nuclear architecture was found to be an essential component of trained immunity transcription of several genes mediated by lncRNA-driven chromatin labels, named immune gene-priming lncRNA (IPLs) [137]. This occurs in responses mediated, for example, by tumor-necrosis factor (TNF), which explain the simultaneous upregulation of several cytokines and chemokines [137]. The mechanism relies on trimethylation of histone 3 at lysine 4 (H3K4me3) on the promoters of trained immune genes, in which the modification is mediated by topologically-associated domain (TAD)-transcribed lncRNAs recruiting mixed lineage leukaemia (MLL) methyltransferases and other transcriptional regulators, leading to RNA polymerase II recruitment and activation [137].
The activation of TLR4 by lipopolysaccharide (LPS) recognition led to differential expression of 221 of 989 evaluated lncRNAs in monocytes [138]. Diverse lncRNAs are capable of interfering in NF-κB signaling, one of the main TFs mediating the inflammatory response. Ma et al. demonstrated that lncRNA-Tnfaip3 acts as a coregulator of NF-κB in murine macrophages [139]. lncRNA Lethe is induced through TNF-α signaling and inhibits transcription of NF-κB-dependent genes by association with RelA, such as Il6, Il8, and Nfkbia [140]. The lncRNA THRIL (TNF-α and hnRNPL related immunoregulatory lncRNA) is induced by TLR2 and regulates TNF-α expression [141]. Sequestration of the p50 subunit by the lncRNA PACER enables the formation of the activation dimer and increases Cox2 transcription [142]. Stimulation with Pam3CSK4 (TLR2), LPS (TLR4), and R848 (TLR7/8) mediates MyD88 and NF-κB transcription, inducing a co-expression of Cox2 and lncRNA-Cox2 [143]. Activation of the TLR-pathway also modulates miRNA [144]. The NAT AS-IL1α regulates IL-1α transcription and is expressed at low levels in resting macrophages and is induced by Listeria monocytogenes infection or TLR ligands followed by NF-κB activation [145]. Other regulated pathways by lncRNAs include the Janus kinase/signal transducers and activators of transcription (JAK-STAT) and mitogen-activated protein kinase (MAPK) pathways [146,147].
Similar to miRNA studies, an increasing number of dysregulation processes during infections have been showing molecular markers of lncRNAs associated with diverse pathogens [148,149,150]. lncRNAs are modulated during infection with Mycobacterium tuberculosis [115,151,152,153,154,155], Salmonella typhimurium [156,157], Escherichia coli [158], Helicobacter pylori [159,160], and Campylobacter oncisus [161].
The measurement of lncRNAs after infection of T-cell lines with the human immunodeficiency virus (HIV) showed regulation of host-lncRNA. A previous study also linked HIV infection with upregulation of lncRNA NEAT1 [162]. The regulation of the non-coding repressor of NFAT (nuclear factor of activated T-cells) named NRON was decreased by the HIV Nef-early replication protein and increased by HIV Vpu-late replication, showing that viruses’ proteins can modulate lncRNA expression on host cells. NRON was shown to be an immune-response subversion molecule, once it impairs expression of genes with NFAT-dependent promoters, through impairment of its nuclear translocation [163]. This model showed that HIV downregulated lncRNA, promoting the NFAT translocation and, consequently, its replication through the long terminal repeat (LTR) expression, evidenced by a higher number of virions in the supernatant of NRON knockdown cells [164] (Figure 4B).
The first report on modulation of lncRNA expression during a protozoan parasite infection was recently reported. Human foreskin fibroblasts infected with Toxoplasma gondii presented 996 lncRNAs differentially expressed in comparison with non-infected cells [165]. Additionally, co-expression networks revealed that these molecules are correlated with modulated mRNAs involved in immune response, mainly proinflammatory cytokines [165]. High or low virulent strains of T. gondii also modulated lncRNAs during infection of mouse bone marrow-derived macrophages [166], corroborating the immune response modulation through lncRNA profile regulation.
Little is known about the mechanism of these lncRNAs during infections, and many models, mainly parasitic and protozoan infections, have not yet been investigated, providing an interesting field of research.
4. Long Non-Coding RNA Expression in Pathogens
Some studies have provided new insights into pathogen genome and transcriptome data. These data describe pathogens with tightly-tuned regulation, whereas the regulatory elements themselves are still largely uncharacterized.
Thousands of lncRNAs were identified within the Schistosoma mansoni [167] and Schistosoma japonicum [168] genomes. Expression analysis revealed specificity along different stages of the parasite, indicating an important modulatory function of lncRNA in the lifecycle of a parasite [167], as well as sex-specific- or drug-resistant-related lncRNAs [169].
The first study in pathogenic fungus was on Cryptococcus neoformans lncRNA RZE1, which participates in the morphological transition from yeast to hypha, essential for pathogenesis [170].
On the side of the protozoans, NATs were described for Plasmodium falciparum [171,172], Trypanosoma brucei [173], Leishmania major [174], Leishmania infantum [175], Giardia lamblia [176,177], and Trichomonas vaginalis [178], although no consensus exists on labelling these transcripts as lncRNAs, as many studies retain the ncRNA definition.
Several lncRNAs were identified in the malaria parasite P. falciparum, which differs in expression during different stages of the life cycle [179,180], with some associated with the telomere-associated repetitive elements (TARE) region [181]. These molecules were associated with the regulation of expression of virulence-related genes [182], especially with the complex regulation of the monoallelic expression of the var genes family [183].
The first evidence of ncRNAs in Leishmania showed RNA polymerase II-dependent transcription from subtelomeric tandem repeats, 3´-end processing by polyadenylation, expression of sense and antisense transcripts in a tightly regulated manner, cytosolic localization, and potential association with a small RNP complex [175,184]. The identification of ncRNAs through computational analysis was described in L. major and L. donovani localized in untranslated regions (UTRs), suggesting UTR transcripts as a common feature in Leishmania [185]. Around 11,243 ncRNAs of different classes were identified along the L. braziliensis genome by algorithm prediction and confirmed by RNA-seq analysis, suggesting that they are real transcripts [186]. Leishmania amazonensis promastigotes express a ncRNA among the top five transcripts differentially expressed in the comparison of wild type parasites with parasite arginase knockout, an enzyme essential to parasite survival and differentiation [148,187,188], indicating that this ncRNA may has a role in the gene expression modulation in different metabolic contexts, such as arginase activity [189].
The transcriptome of T. brucei revealed ncRNAs without open reading frames (ORFs) or having the potential to codify small peptides [190], but no study has demonstrated functionality for such transcripts.
lncRNA description in eukaryotic pathogens is poorly annotated, probably due to insufficient genome investigation and annotations, mostly because of complex genomic arrangements. However, the characterization of these molecules could improve the knowledge of fungi and parasite biology, genomic organization, and transcriptional expression and regulation, possibly providing potential drug targets.
The lncRNA function in virus replication has been extensively studied, as reviewed by Chavez-Calvillo [191], showing that lncRNA encoded by Gammaherpesvirus, a subfamily of Herpesviridae, is directly involved in the scope of the immune response and viral lytic reactivation, promoting virus genes expression and DNA amplification in the assembly and release of progeny virions [192], as exemplified by Epstein–Barr virus (EBV)-encoded RNA1 (EBER1) [193], EBV-encoded RNA1 and 2 (EBER1 and EBER2, respectively) [194], and Kaposi’s sarcoma-associated herpesvirus (KSHV) polyadenylated nuclear (PAN) RNA [195].
5. Novel Perspectives
lncRNAs comprise a field of study that has gained visibility and has become attractive to the scientific community. Findings have evidenced the complexity of mechanisms of action of these molecules and that their functions surpass those initially described. Many complicating factors have been added to the study of these molecules, including the protein coding potential of some molecules annotated as non-coding. This has attracted those interested in gene expression regulation. The clinical importance of these findings is being exploited to find novel biomarkers and drug targets due to both the specificity of lncRNA expression and the potential for plasma localization when secreted in extracellular vesicles (EVs).
5.1. Long Non-Coding RNAs as Biomarkers
The importance of understanding lncRNA mechanisms extends beyond the description of gene regulation and can be used as diagnostic markers or drug targets as well as prognostic markers. One potential diagnosis marker is the prostate cancer-associated transcript 1 (PCAT-1), which can be identified in the urine of prostate cancer patients [196]. Still on prognosis, the lncRNAs named CAT104, LINC01234, and STXBP5-AS1 have clinical significance in predicting survival of breast cancer [197]. lncRNAs have been proposed as therapeutic targets, and difficulties reside within complete understanding of their mechanism of action [198,199].
Many cancer models were demonstrated to release EVs containing different sets of ncRNAs, such as miRNAs and lncRNAs (Figure 2). Some specific transcripts are abundant in EVs, indicating that they may play a role in neighboring cells. This was shown in a colorectal cancer model where miRNA [200], lncRNA [201], and circRNA [202] were selectively exported to EVs and were shown to be not simply correlated with cytosolic RNA pool levels. During hypoxia, hepatocellular cancer cells contained the linc-RoR (regulator of reprogramming), thus decreasing miR-145 and hypoxia-inducible factor 1 alpha (HIF-1α) in recipient cells [203]. This feature demonstrates the potential for lncRNAs as biomarkers, as shown in liver cancer [204].
5.2. Micropeptides
The postulate that lncRNAs do not codify proteins was questioned by the description of functional peptides codified by small ORFs. Once the presence of initiation codons followed by stop codons of translation is frequent in a genome, it was necessary to apply appropriate techniques to the identification of peptides codified by small ORFs [205]. Some studies demonstrated the translation and functionality of peptides from the sequencing of RNA fragments bound to ribosomes, followed by the identification of micropeptides through mass spectrometry together with an evolutive analysis, demonstrating the conservation of these sequences [206,207]. This discovery evidenced the importance of verifying the translation potential in these molecules. The products are called micropeptides, small peptides (sPEPs), or peptides of small ORFs (smORFs), and their role in physiological and pathological processes demonstrated their biological importance.
Some studies detailed mechanisms of regulation of muscular function, characterized by contraction and relaxing mechanisms, by micropeptides codified by lncRNAs. Myoregulin (MLN) interacts with the sarcoendoplasmic reticulum calcium transport ATPase SERCA, preventing calcium uptake (Ca2+) to the sarcoplasmic reticulum in skeletal muscle and; therefore, knockout mice with this gene have a higher potential of muscular function [208]. The micropeptide DWORF increases SERCA activity by withdrawing the inhibitors phospholamban, sarcolipin, and myoregulin. Knockout mice manipulated by CRISPR/Cas9 showed a reduction in SERCA activity and retarded relaxation of muscle fibers [209]. The lncRNA LINC00961 produces a polypeptide named SPAR (small regulatory polypeptide of amino acid response) that inhibits mTORC1 activation, which is important in muscle regeneration [210,211].
6. Concluding Remarks
The central dogma of biology changed with the discovery of functional ncRNA molecules, such as rRNAs and tRNAs. After many observations were initially treated with distrust by the scientific community, the importance of these ncRNAs was demonstrated by their functionality, mainly in gene expression regulation.
The study of ncRNAs involved in gene expression regulation started with the discovery of small RNAs capable of binding to the 3’ UTR region of mRNAs, thus controlling protein production. The well-known miRNAs are associated with physiological processes, chronic pathologies, and infectious diseases. The emergence of long RNAs without an apparent ORF soon confirmed the presence of lncRNAs, involved in some of the same processes as miRNAs. In this context, many studies rapidly evaluated the regulatory functions shared by these molecules in signaling or metabolic pathways. Many studies demonstrated the interaction and regulation between these molecules and provided a new perspective on the ncRNA world. lncRNAs revealed their complexity when they were described as able to interact simultaneously with diverse macromolecules, controlling different points of gene expression flux. We encourage the study of different subsets of ncRNA in different models given the importance of their coregulation.
However, the annotation of these lncRNAs as non-coding is controversial. New studies have demonstrated that some can be translated into micropeptides that exert biological functions. The complexity of these molecules involves the capacity to interact with many types of molecules within the cell, and then the possibility of translation into peptides, increasing the regulatory capacity. Both forms are active and seem to be part of a regulatory network not yet completely described. We still need to understand how cellular components, or the molecule itself, are capable of controlling the timing of its translation.
In describing the variety of lncRNAs classes, the complexity exceeds the description of their chromosomal loci, used as the default in classification. lncRNAs can also be related to their ability to interact with different molecules, defining their involvement in the regulation of gene expression. The ability to acquire secondary structures mediates these interactions, such as the formation of circular structures, highlighting the multifunctional characteristics of these molecules. For much of the transcriptomic data deposited in databanks, no attempt has yet been made to analyze lncRNA regulation. Therefore, groups with interest in this area are encouraged to use pipelines developed to perform this analysis [212], for prospecting and further validating their functionality in different models.
Author Contributions
J.C.R.F., S.M.M., and J.I.A. wrote the original article; J.C.R.F. and S.M.A. designed the figures; S.M.M., J.I.A., J.C.R.F., S.M.A., and L.M.F.-W. revised and approved the final version of this review.
Funding
This work was supported by grants from the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq, http://www.cnpq.br: 406351/2018-0.) and Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP, http://www.fapesp.br: 2014/50717-1, 2016/19815-2, 2018/24693-9). FAPESP fellowship: J.C.R.F. (2017/21906-9), S.M.A. (2017/23519-2), and J.I.A. (2016/03273-6). L.M.F.-W. received a research fellowship from CNPq (307587/2014-2).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Long, Y.; Wang, X.; Youmans, D.T.; Cech, T.R. How do lncRNAs regulate transcription? Sci. Adv. 2017, 3, eaao2110. [Google Scholar] [CrossRef] [PubMed]
- Blin, N.; Stephenson, E.C.; Stafford, D.W. Isolation and some properties of a mammalian ribosomal DNA. Chromosoma 1976, 58, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Stephenson, M.L.; Zamecnik, P.C. Inhibition of Rous sarcoma viral RNA translation by a specific oligodeoxyribonucleotide. Proc. Natl. Acad. Sci. USA 1978, 75, 285–288. [Google Scholar] [CrossRef] [PubMed]
- Küpper, H.; Sekiya, T.; Rosenberg, M.; Egan, J.; Landy, A. A Rho-dependent termination site in the gene coding for tyrosine tRNA su3 of Escherichia Coli. Nature 1978, 272, 423–428. [Google Scholar] [CrossRef] [PubMed]
- Derrien, T.; Johnson, R.; Bussotti, G.; Tanzer, A.; Djebali, S.; Tilgner, H.; Guernec, G.; Martin, D.; Merkel, A.; Knowles, D.G.; et al. The gencode v7 catalog of human long noncoding RNAs: Analysis of their gene structure, evolution, and expression. Genome Res. 2012, 22, 1775–1789. [Google Scholar] [CrossRef] [PubMed]
- Djebali, S.; Davis, C.A.; Merkel, A.; Dobin, A.; Lassmann, T.; Mortazavi, A.; Tanzer, A.; Lagarde, J.; Lin, W.; Schlesinger, F.; et al. Landscape of transcription in human cells. Nature 2012, 489, 101–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ulitsky, I.; Bartel, D.P. Lincrnas: Genomics, evolution, and mechanisms. Cell 2013, 154, 26–46. [Google Scholar] [CrossRef]
- Rinn, J.L.; Chang, H.Y. Genome regulation by long noncoding RNAs. Ann. Rev. Biochem. 2012, 81, 145–166. [Google Scholar] [CrossRef]
- Cabili, M.N.; Trapnell, C.; Goff, L.; Koziol, M.; Tazon-Vega, B.; Regev, A.; Rinn, J.L. Integrative annotation of human large intergenic noncoding rnas reveals global properties and specific subclasses. Genes Dev. 2011, 25, 1915–1927. [Google Scholar] [CrossRef]
- Li, K.; Ramchandran, R. Natural antisense transcript: A concomitant engagement with protein-coding transcript. Oncotarget. 2010, 1, 447–452. [Google Scholar] [CrossRef]
- Quinn, J.J.; Chang, H.Y. Unique features of long non-coding RNA biogenesis and function. Nat. Rev. Genet. 2016, 17, 47–62. [Google Scholar] [CrossRef]
- Hangauer, M.J.; Vaughn, I.W.; McManus, M.T. Pervasive transcription of the human genome produces thousands of previously unidentified long intergenic noncoding RNAs. PLoS Genet. 2013, 9, e1003569. [Google Scholar] [CrossRef]
- Niemczyk, M.; Ito, Y.; Huddleston, J.; Git, A.; Abu-Amero, S.; Caldas, C.; Moore, G.E.; Stojic, L.; Murrell, A. Imprinted chromatin around DIRAS3 regulates alternative splicing of GNG12-AS1, a long noncoding RNA. Am. J. Hum. Genet. 2013, 93, 224–235. [Google Scholar] [CrossRef]
- Ziegler, C.; Kretz, M. The more the merrier-complexity in long non-coding RNA loci. Front. Endocrinol. (Lausanne) 2017, 8. [Google Scholar] [CrossRef]
- Guttman, M.; Amit, I.; Garber, M.; French, C.; Lin, M.F.; Feldser, D.; Huarte, M.; Zuk, O.; Carey, B.W.; Cassady, J.P.; et al. Chromatin signature reveals over a thousand highly conserved large non-coding RNAs in mammals. Nature 2009, 458, 223–227. [Google Scholar] [CrossRef] [Green Version]
- Zheng, G.X.; Do, B.T.; Webster, D.E.; Khavari, P.A.; Chang, H.Y. Dicer-microRNA-Myc circuit promotes transcription of hundreds of long noncoding RNAs. Nat. Struct. Mol. Biol. 2014, 21, 585–590. [Google Scholar] [CrossRef] [Green Version]
- Uesaka, M.; Nishimura, O.; Go, Y.; Nakashima, K.; Agata, K.; Imamura, T. Bidirectional promoters are the major source of gene activation-associated non-coding rnas in mammals. BMC Genom. 2014, 15, 35. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, X.S.; Liu, Q.-R.; Wei, L. Genome-wide in silico identification and analysis of cis natural antisense transcripts (cis-NATS) in ten species. Nucleic Acids Res. 2006, 34, 3465–3475. [Google Scholar] [CrossRef]
- Seila, A.C.; Calabrese, J.M.; Levine, S.S.; Yeo, G.W.; Rahl, P.B.; Flynn, R.A.; Young, R.A.; Sharp, P.A. Divergent transcription from active promoters. Science 2008, 322, 1849–1851. [Google Scholar] [CrossRef]
- Core, L.J.; Waterfall, J.J.; Lis, J.T. Nascent RNA sequencing reveals widespread pausing and divergent initiation at human promoters. Science 2008, 322, 1845–1848. [Google Scholar] [CrossRef]
- Sigova, A.A.; Mullen, A.C.; Molinie, B.; Gupta, S.; Orlando, D.A.; Guenther, M.G.; Almada, A.E.; Lin, C.; Sharp, P.A.; Giallourakis, C.C.; et al. Divergent transcription of long noncoding RNA/mRNA gene pairs in embryonic stem cells. Proc. Natl. Acad. Sci. USA 2013, 110, 2876–2881. [Google Scholar] [CrossRef] [Green Version]
- Andersson, R.; Gebhard, C.; Miguel-Escalada, I.; Hoof, I.; Bornholdt, J.; Boyd, M.; Chen, Y.; Zhao, X.; Schmidl, C.; Suzuki, T.; et al. An atlas of active enhancers across human cell types and tissues. Nature 2014, 507, 455–461. [Google Scholar] [CrossRef] [Green Version]
- Lai, F.; Gardini, A.; Zhang, A.; Shiekhattar, R. Integrator mediates the biogenesis of enhancer RNAs. Nature 2015, 525, 399–403. [Google Scholar] [CrossRef] [Green Version]
- Salzman, J.; Gawad, C.; Wang, P.L.; Lacayo, N.; Brown, P.O. Circular RNAs are the predominant transcript isoform from hundreds of human genes in diverse cell types. PLoS ONE 2012, 7, e30733. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, X.O.; Chen, T.; Xiang, J.F.; Yin, Q.F.; Xing, Y.H.; Zhu, S.; Yang, L.; Chen, L.L. Circular intronic long noncoding RNAs. Mol. Cell 2013, 51, 792–806. [Google Scholar] [CrossRef]
- Li, Z.; Huang, C.; Bao, C.; Chen, L.; Lin, M.; Wang, X.; Zhong, G.; Yu, B.; Hu, W.; Dai, L.; et al. Exon-intron circular rnas regulate transcription in the nucleus. Nat. Struct. Mol. Biol. 2015, 22, 256–264. [Google Scholar] [CrossRef]
- Geisler, S.; Lojek, L.; Khalil, A.M.; Baker, K.E.; Coller, J. Decapping of long noncoding RNAs regulates inducible genes. Mol. Cell 2012, 45, 279–291. [Google Scholar] [CrossRef]
- Wery, M.; Descrimes, M.; Vogt, N.; Dallongeville, A.S.; Gautheret, D.; Morillon, A. Nonsense-mediated decay restricts lncRNA levels in yeast unless blocked by double-stranded RNA structure. Mol. Cell 2016, 61, 379–392. [Google Scholar] [CrossRef]
- Beck, Z.T.; Xing, Z.; Tran, E.J. LncRNAs: Bridging environmental sensing and gene expression. RNA Biol. 2016, 13, 1189–1196. [Google Scholar] [CrossRef]
- Zhao, Y.; Li, H.; Fang, S.; Kang, Y.; Wu, W.; Hao, Y.; Li, Z.; Bu, D.; Sun, N.; Zhang, M.Q.; et al. Noncode 2016: An informative and valuable data source of long non-coding RNAs. Nucleic Acids Res. 2016, 44, D203–D208. [Google Scholar] [CrossRef]
- Chen, Y.G.; Satpathy, A.T.; Chang, H.Y. Gene regulation in the immune system by long noncoding RNAs. Nat. Immunol. 2017, 18, 962–972. [Google Scholar] [CrossRef]
- Goff, L.A.; Groff, A.F.; Sauvageau, M.; Trayes-Gibson, Z.; Sanchez-Gomez, D.B.; Morse, M.; Martin, R.D.; Elcavage, L.E.; Liapis, S.C.; Gonzalez-Celeiro, M.; et al. Spatiotemporal expression and transcriptional perturbations by long noncoding RNAs in the mouse brain. Proc. Natl. Acad Sci. USA 2015, 112, 6855–6862. [Google Scholar] [CrossRef] [Green Version]
- Iyer, M.K.; Niknafs, Y.S.; Malik, R.; Singhal, U.; Sahu, A.; Hosono, Y.; Barrette, T.R.; Prensner, J.R.; Evans, J.R.; Zhao, S.; et al. The landscape of long noncoding RNAs in the human transcriptome. Nat. Genet. 2015, 47, 199–208. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.J.; Horlbeck, M.A.; Cho, S.W.; Birk, H.S.; Malatesta, M.; He, D.; Attenello, F.J.; Villalta, J.E.; Cho, M.Y.; Chen, Y.; et al. Crispri-based genome-scale identification of functional long noncoding RNA loci in human cells. Science 2017, 355, eaah7111. [Google Scholar] [CrossRef]
- Joung, J.; Engreitz, J.M.; Konermann, S.; Abudayyeh, O.O.; Verdine, V.K.; Aguet, F.; Gootenberg, J.S.; Sanjana, N.E.; Wright, J.B.; Fulco, C.P.; et al. Genome-scale activation screen identifies a lncRNA locus regulating a gene neighbourhood. Nature 2017, 548, 343–346. [Google Scholar] [CrossRef] [Green Version]
- Hu, G.; Tang, Q.; Sharma, S.; Yu, F.; Escobar, T.M.; Muljo, S.A.; Zhu, J.; Zhao, K. Expression and regulation of intergenic long noncoding RNAs during t cell development and differentiation. Nat. Immunol. 2013, 14, 1190–1198. [Google Scholar] [CrossRef]
- Cech, T.R.; Steitz, J.A. The noncoding RNA revolution—trashing old rules to forge new ones. Cell 2014, 157, 77–94. [Google Scholar] [CrossRef]
- Rackham, O.; Shearwood, A.-M.J.; Mercer, T.R.; Davies, S.M.K.; Mattick, J.S.; Filipovska, A. Long noncoding RNAs are generated from the mitochondrial genome and regulated by nuclear-encoded proteins. RNA 2011, 17, 2085–2093. [Google Scholar] [CrossRef]
- Cabili, M.N.; Dunagin, M.C.; McClanahan, P.D.; Biaesch, A.; Padovan-Merhar, O.; Regev, A.; Rinn, J.L.; Raj, A. Localization and abundance analysis of human lncRNAs at single-cell and single-molecule resolution. Genome Biol. 2015, 16, 20. [Google Scholar] [CrossRef]
- Carlevaro-Fita, J.; Rahim, A.; Guigó, R.; Vardy, L.A.; Johnson, R. Cytoplasmic long noncoding RNAs are frequently bound to and degraded at ribosomes in human cells. RNA 2016, 22, 867–882. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Gunawardane, L.; Niazi, F.; Jahanbani, F.; Chen, X.; Valadkhan, S. A novel RNA motif mediates the strict nuclear localization of a long noncoding RNA. Mol. Cell. Biol. 2014, 34, 2318–2329. [Google Scholar] [CrossRef]
- Lubelsky, Y.; Ulitsky, I. Sequences enriched in alu repeats drive nuclear localization of long RNAs in human cells. Nature 2018, 555, 107–111. [Google Scholar] [CrossRef]
- Kertesz, M.; Wan, Y.; Mazor, E.; Rinn, J.L.; Nutter, R.C.; Chang, H.Y.; Segal, E. Genome-wide measurement of RNA secondary structure in yeast. Nature 2010, 467, 103–107. [Google Scholar] [CrossRef] [Green Version]
- Underwood, J.G.; Uzilov, A.V.; Katzman, S.; Onodera, C.S.; Mainzer, J.E.; Mathews, D.H.; Lowe, T.M.; Salama, S.R.; Haussler, D. Fragseq: Transcriptome-wide RNA structure probing using high-throughput sequencing. Nat. Methods 2010, 7, 995–1001. [Google Scholar] [CrossRef]
- Pegueroles, C.; Gabaldón, T. Secondary structure impacts patterns of selection in human lncRNAs. BMC Biol. 2016, 14, 60. [Google Scholar] [CrossRef]
- Smith, M.A.; Gesell, T.; Stadler, P.F.; Mattick, J.S. Widespread purifying selection on RNA structure in mammals. Nucleic Acids Res. 2013, 41, 8220–8236. [Google Scholar] [CrossRef] [Green Version]
- Diederichs, S. The four dimensions of noncoding RNA conservation. Trends Genet. 2014, 30, 121–123. [Google Scholar] [CrossRef]
- Liu, N.; Dai, Q.; Zheng, G.; He, C.; Parisien, M.; Pan, T. N(6)-methyladenosine-dependent RNA structural switches regulate RNA-protein interactions. Nature 2015, 518, 560–564. [Google Scholar] [CrossRef]
- Zhou, K.I.; Parisien, M.; Dai, Q.; Liu, N.; Diatchenko, L.; Sachleben, J.R.; Pan, T. N(6)-methyladenosine modification in a long noncoding RNA hairpin predisposes its conformation to protein binding. J. Mol. Biol. 2016, 428, 822–833. [Google Scholar] [CrossRef]
- Liu, N.; Pan, T. N6-methyladenosine-encoded epitranscriptomics. Nat. Struct. Mol. Biol. 2016, 23, 98–102. [Google Scholar] [CrossRef]
- Gong, J.; Liu, C.; Liu, W.; Xiang, Y.; Diao, L.; Guo, A.-Y.; Han, L. Lncediting: A database for functional effects of RNA editing in lncRNAs. Nucleic Acids Res. 2017, 45, D79–D84. [Google Scholar] [CrossRef]
- Singer, R.H.; Green, M.R. Compartmentalization of eukaryotic gene expression: Causes and effects. Cell 1997, 91, 291–294. [Google Scholar] [CrossRef]
- Kopp, F.; Mendell, J.T. Functional classification and experimental dissection of long noncoding RNAs. Cell 2018, 172, 393–407. [Google Scholar] [CrossRef]
- Hewson, C.; Capraro, D.; Burdach, J.; Whitaker, N.; Morris, K.V. Extracellular vesicle associated long non-coding RNAs functionally enhance cell viability. Non-Coding RNA Res. 2016, 1, 3–11. [Google Scholar] [CrossRef] [Green Version]
- Paul, J.; Duerksen, J.D. Chromatin-associated rna content of heterochromatin and euchromatin. Mol. Cell. Biochem. 1975, 9, 9–16. [Google Scholar] [CrossRef]
- Guttman, M.; Donaghey, J.; Carey, B.W.; Garber, M.; Grenier, J.K.; Munson, G.; Young, G.; Lucas, A.B.; Ach, R.; Bruhn, L.; et al. Lincrnas act in the circuitry controlling pluripotency and differentiation. Nature 2011, 477, 295–300. [Google Scholar] [CrossRef]
- Lai, F.; Orom, U.A.; Cesaroni, M.; Beringer, M.; Taatjes, D.J.; Blobel, G.A.; Shiekhattar, R. Activating RNAs associate with mediator to enhance chromatin architecture and transcription. Nature 2013, 494, 497–501. [Google Scholar] [CrossRef]
- Khalil, A.M.; Guttman, M.; Huarte, M.; Garber, M.; Raj, A.; Rivea Morales, D.; Thomas, K.; Presser, A.; Bernstein, B.E.; van Oudenaarden, A.; et al. Many human large intergenic noncoding RNAs associate with chromatin-modifying complexes and affect gene expression. Proc. Natl. Acad. Sci. USA 2009, 106, 11667–11672. [Google Scholar] [CrossRef] [Green Version]
- Engreitz, J.M.; Haines, J.E.; Perez, E.M.; Munson, G.; Chen, J.; Kane, M.; McDonel, P.E.; Guttman, M.; Lander, E.S. Local regulation of gene expression by lncRNA promoters, transcription and splicing. Nature 2016, 539, 452–455. [Google Scholar] [CrossRef] [Green Version]
- Hawkins, P.G.; Morris, K.V. Transcriptional regulation of Oct4 by a long non-coding RNA antisense to Oct4-pseudogene 5. Transcription 2010, 1, 165–175. [Google Scholar] [CrossRef]
- Azofeifa, J.G.; Allen, M.A.; Hendrix, J.R.; Read, T.; Rubin, J.D.; Dowell, R.D. Enhancer RNA profiling predicts transcription factor activity. Genome Res. 2018. [Google Scholar] [CrossRef]
- Hansen, T.B.; Jensen, T.I.; Clausen, B.H.; Bramsen, J.B.; Finsen, B.; Damgaard, C.K.; Kjems, J. Natural RNA circles function as efficient microRNA sponges. Nature 2013, 495, 384–388. [Google Scholar] [CrossRef]
- Furió-Tarí, P.; Tarazona, S.; Gabaldón, T.; Enright, A.J.; Conesa, A. Spongescan: A web for detecting microrna binding elements in lncRNA sequences. Nucleic Acids Res. 2016, 44, W176–W180. [Google Scholar] [CrossRef]
- Zhu, M.; Liu, J.; Xiao, J.; Yang, L.; Cai, M.; Shen, H.; Chen, X.; Ma, Y.; Hu, S.; Wang, Z.; et al. Lnc-mg is a long non-coding RNA that promotes myogenesis. Nat. Commun. 2017, 8, 14718. [Google Scholar] [CrossRef] [Green Version]
- Fu, N.; Niu, X.; Wang, Y.; Du, H.; Wang, B.; Du, J.; Li, Y.; Wang, R.; Zhang, Y.; Zhao, S.; et al. Role of lncrna-activated by transforming growth factor beta in the progression of hepatitis C virus-related liver fibrosis. Discov. Med. 2016, 22, 29–42. [Google Scholar]
- Dhir, A.; Dhir, S.; Proudfoot, N.J.; Jopling, C.L. Microprocessor mediates transcriptional termination of long noncoding RNA transcripts hosting microRNAs. Nat. Struct. Mol. Biol. 2015, 22, 319–327. [Google Scholar] [CrossRef] [Green Version]
- Augoff, K.; McCue, B.; Plow, E.F.; Sossey-Alaoui, K. Mir-31 and its host gene lncRNA loc554202 are regulated by promoter hypermethylation in triple-negative breast cancer. Mol. Cancer 2012, 11, 5. [Google Scholar] [CrossRef]
- Calin, G.A.; Dumitru, C.D.; Shimizu, M.; Bichi, R.; Zupo, S.; Noch, E.; Aldler, H.; Rattan, S.; Keating, M.; Rai, K.; et al. Frequent deletions and down-regulation of micro-RNA genes mir15 and mir16 at 13q14 in chronic lymphocytic leukemia. Proc. Natl. Acad. Sci. USA 2002, 99, 15524–15529. [Google Scholar] [CrossRef]
- Lerner, M.; Harada, M.; Lovén, J.; Castro, J.; Davis, Z.; Oscier, D.; Henriksson, M.; Sangfelt, O.; Grandér, D.; Corcoran, M.M. Dleu2, frequently deleted in malignancy, functions as a critical host gene of the cell cycle inhibitory microRNAs mir-15a and mir-16-1. Exp. Cell Res. 2009, 315, 2941–2952. [Google Scholar] [CrossRef]
- Yu, Y.; Nangia-Makker, P.; Farhana, L.; Majumdar, A.P.N. A novel mechanism of lncRNA and miRNA interaction: Ccat2 regulates mir-145 expression by suppressing its maturation process in colon cancer cells. Mol. Cancer 2017, 16, 155. [Google Scholar] [CrossRef]
- Leung, A.; Trac, C.; Jin, W.; Lanting, L.; Akbany, A.; Saetrom, P.; Schones, D.E.; Natarajan, R. Novel long noncoding RNAs are regulated by angiotensin II in vascular smooth muscle cells. Circ. Res. 2013, 113, 266–278. [Google Scholar] [CrossRef]
- Yoon, J.-H.; Abdelmohsen, K.; Gorospe, M. Functional interactions among microRNAs and long noncoding RNAs. Semin. Cell Dev. Biol. 2014, 34, 9–14. [Google Scholar] [CrossRef] [Green Version]
- Yao, Y.; Ma, J.; Xue, Y.; Wang, P.; Li, Z.; Liu, J.; Chen, L.; Xi, Z.; Teng, H.; Wang, Z.; et al. Knockdown of long non-coding RNA xist exerts tumor-suppressive functions in human glioblastoma stem cells by up-regulating mir-152. Cancer Lett. 2015, 359, 75–86. [Google Scholar] [CrossRef]
- Wang, X.; Li, M.; Wang, Z.; Han, S.; Tang, X.; Ge, Y.; Zhou, L.; Zhou, C.; Yuan, Q.; Yang, M. Silencing of long noncoding RNA malat1 by mir-101 and mir-217 inhibits proliferation, migration, and invasion of esophageal squamous cell carcinoma cells. J. Biol. Chem. 2015, 290, 3925–3935. [Google Scholar] [CrossRef]
- You, J.; Zhang, Y.; Liu, B.; Li, Y.; Fang, N.; Zu, L.; Li, X.; Zhou, Q. MicroRNA-449a inhibits cell growth in lung cancer and regulates long noncoding RNA nuclear enriched abundant transcript 1. Ind. J. Cancer 2014, 51 3, 77–81. [Google Scholar] [CrossRef]
- Faghihi, M.A.; Zhang, M.; Huang, J.; Modarresi, F.; Van der Brug, M.P.; Nalls, M.A.; Cookson, M.R.; St-Laurent, G., 3rd; Wahlestedt, C. Evidence for natural antisense transcript-mediated inhibition of microRNA function. Genome Biol. 2010, 11, R56. [Google Scholar] [CrossRef]
- Kallen, A.N.; Zhou, X.B.; Xu, J.; Qiao, C.; Ma, J.; Yan, L.; Lu, L.; Liu, C.; Yi, J.S.; Zhang, H.; et al. The imprinted h19 lncRNA antagonizes let-7 microRNAs. Mol. Cell 2013, 52, 101–112. [Google Scholar] [CrossRef]
- Dey, B.K.; Pfeifer, K.; Dutta, A. The h19 long noncoding rna gives rise to microRNAs mir-675-3p and mir-675-5p to promote skeletal muscle differentiation and regeneration. Genes Dev. 2014, 28, 491–501. [Google Scholar] [CrossRef]
- Cesana, M.; Cacchiarelli, D.; Legnini, I.; Santini, T.; Sthandier, O.; Chinappi, M.; Tramontano, A.; Bozzoni, I. A long noncoding RNA controls muscle differentiation by functioning as a competing endogenous RNA. Cell 2011, 147, 358–369. [Google Scholar] [CrossRef]
- Emmrich, S.; Streltsov, A.; Schmidt, F.; Thangapandi, V.R.; Reinhardt, D.; Klusmann, J.H. LincRNAs monc and mir100hg act as oncogenes in acute megakaryoblastic leukemia. Mol. Cancer 2014, 13, 171. [Google Scholar] [CrossRef]
- Kan, Z.; Rouchka, E.C.; Gish, W.R.; States, D.J. Gene structure prediction and alternative splicing analysis using genomically aligned ests. Genome Res. 2001, 11, 889–900. [Google Scholar] [CrossRef]
- Will, C.L.; Luhrmann, R. Spliceosome structure and function. Cold Spring Harb. Perspect. Biol. 2011, 3. [Google Scholar] [CrossRef]
- Hutchinson, J.N.; Ensminger, A.W.; Clemson, C.M.; Lynch, C.R.; Lawrence, J.B.; Chess, A. A screen for nuclear transcripts identifies two linked noncoding RNAs associated with Sc35 splicing domains. BMC Genom. 2007, 8, 39. [Google Scholar] [CrossRef]
- Tripathi, V.; Ellis, J.D.; Shen, Z.; Song, D.Y.; Pan, Q.; Watt, A.T.; Freier, S.M.; Bennett, C.F.; Sharma, A.; Bubulya, P.A.; et al. The nuclear-retained noncoding RNA MALAT1 regulates alternative splicing by modulating SR splicing factor phosphorylation. Mol. Cell 2010, 39, 925–938. [Google Scholar] [CrossRef]
- Gonzalez, I.; Munita, R.; Agirre, E.; Dittmer, T.A.; Gysling, K.; Misteli, T.; Luco, R.F. A lncRNA regulates alternative splicing via establishment of a splicing-specific chromatin signature. Nat. Struct. Mol. Biol. 2015, 22, 370–376. [Google Scholar] [CrossRef]
- Romero-Barrios, N.; Legascue, M.F.; Benhamed, M.; Ariel, F.; Crespi, M. Splicing regulation by long noncoding RNAs. Nucleic Acids Res. 2018, 46, 2169–2184. [Google Scholar] [CrossRef] [Green Version]
- Villamizar, O.; Chambers, C.B.; Riberdy, J.M.; Persons, D.A.; Wilber, A. Long noncoding RNA SAF and splicing factor 45 increase soluble FAS and resistance to apoptosis. Oncotarget 2016, 7, 13810–13826. [Google Scholar] [CrossRef]
- Gong, C.; Maquat, L.E. LncRNAs transactivate Stau1-mediated mRNA decay by duplexing with 3′ UTRS via Alu elements. Nature 2011, 470, 284–288. [Google Scholar] [CrossRef]
- Kerin, T.; Ramanathan, A.; Rivas, K.; Grepo, N.; Coetzee, G.A.; Campbell, D.B. A noncoding RNA antisense to Moesin at 5p14.1 in autism. Sci. Transl. Med. 2012, 4, 128ra140. [Google Scholar] [CrossRef]
- Johnsson, P.; Ackley, A.; Vidarsdottir, L.; Lui, W.O.; Corcoran, M.; Grander, D.; Morris, K.V. A pseudogene long-noncoding-RNA network regulates Pten transcription and translation in human cells. Nat. Struct. Mol. Biol. 2013, 20, 440–446. [Google Scholar] [CrossRef]
- Faghihi, M.A.; Modarresi, F.; et al. Expression of a noncoding RNA is elevated in alzheimer’s disease and drives rapid feed-forward regulation of beta-secretase. Nat. Med. 2008, 14, 723–730. [Google Scholar] [CrossRef]
- Jadaliha, M.; Gholamalamdari, O.; Tang, W.; Zhang, Y.; Petracovici, A.; Hao, Q.; Tariq, A.; Kim, T.G.; Holton, S.E.; Singh, D.K.; et al. A natural antisense lncRNA controls breast cancer progression by promoting tumor suppressor gene mRNA stability. PLoS Genet. 2018, 14, e1007802. [Google Scholar] [CrossRef]
- Carrieri, C.; Cimatti, L.; Biagioli, M.; Beugnet, A.; Zucchelli, S.; Fedele, S.; Pesce, E.; Ferrer, I.; Collavin, L.; Santoro, C.; et al. Long non-coding antisense RNA controls Uchl1 translation through an embedded Sineb2 repeat. Nature 2012, 491, 454–457. [Google Scholar] [CrossRef]
- Wang, K.C.; Yang, Y.W.; Liu, B.; Sanyal, A.; Corces-Zimmerman, R.; Chen, Y.; Lajoie, B.R.; Protacio, A.; Flyn, R.A.; Gupta, R.A.; et al. A long noncoding RNA maintains active chromatin to coordinate homeotic gene expression. Nature 2011, 472, 120–124. [Google Scholar] [CrossRef] [Green Version]
- Rashid, F.; Shah, A.; Shan, G. Long non-coding RNAs in the cytoplasm. Genom. Proteom. Bioinform. 2016, 14, 73–80. [Google Scholar] [CrossRef]
- Salviano-Silva, A.; Lobo-Alves, S.C.; Almeida, R.C.; Malheiros, D.; Petzl-Erler, M.L. Besides pathology: Long non-coding RNA in cell and tissue homeostasis. Non-Coding RNA 2018, 4, 3. [Google Scholar] [CrossRef]
- Kapranov, P.; Cheng, J.; Dike, S.; Nix, D.A.; Duttagupta, R.; Willingham, A.T.; Stadler, P.F.; Hertel, J.; Hackermuller, J.; Hofacker, I.L.; et al. Rna maps reveal new RNA classes and a possible function for pervasive transcription. Science 2007, 316, 1484–1488. [Google Scholar] [CrossRef]
- Carninci, P.; Kasukawa, T.; Katayama, S.; Gough, J.; Frith, M.C.; Maeda, N.; Oyama, R.; Ravasi, T.; Lenhard, B.; Wells, C.; et al. The transcriptional landscape of the mammalian genome. Science 2005, 309, 1559–1563. [Google Scholar] [CrossRef]
- Brockdorff, N.; Ashworth, A.; Kay, G.F.; McCabe, V.M.; Norris, D.P.; Cooper, P.J.; Swift, S.; Rastan, S. The product of the mouse xist gene is a 15 kb inactive X-specific transcript containing no conserved Orf and located in the nucleus. Cell 1992, 71, 515–526. [Google Scholar] [CrossRef]
- Brown, C.J.; Hendrich, B.D.; Rupert, J.L.; Lafrenière, R.G.; Xing, Y.; Lawrence, J.; Willard, H.F. The human xist gene: Analysis of a 17 kb inactive X-specific rna that contains conserved repeats and is highly localized within the nucleus. Cell 1992, 71, 527–542. [Google Scholar] [CrossRef]
- Schoeftner, S.; Sengupta, A.K.; Kubicek, S.; Mechtler, K.; Spahn, L.; Koseki, H.; Jenuwein, T.; Wutz, A. Recruitment of prc1 function at the initiation of X inactivation independent of prc2 and silencing. EMBO J. 2006, 25, 3110–3122. [Google Scholar] [CrossRef]
- Chaumeil, J.; Le Baccon, P.; Wutz, A.; Heard, E. A novel role for xist RNA in the formation of a repressive nuclear compartment into which genes are recruited when silenced. Genes Dev. 2006, 20, 2223–2237. [Google Scholar] [CrossRef]
- Sarma, K.; Levasseur, P.; Aristarkhov, A.; Lee, J.T. Locked nucleic acids (lnas) reveal sequence requirements and kinetics of xist RNA localization to the X chromosome. Proc. Natl. Acad. Sci. USA 2010, 107, 22196–22201. [Google Scholar] [CrossRef]
- Chow, J.C.; Ciaudo, C.; Fazzari, M.J.; Mise, N.; Servant, N.; Glass, J.L.; Attreed, M.; Avner, P.; Wutz, A.; Barillot, E.; et al. Line-1 activity in facultative heterochromatin formation during X chromosome inactivation. Cell 2010, 141, 956–969. [Google Scholar] [CrossRef]
- Lee, J.T.; Jaenisch, R. Long-range cis effects of ectopic X-inactivation centres on a mouse autosome. Nature 1997, 386, 275–279. [Google Scholar] [CrossRef]
- Wutz, A. Gene silencing in X-chromosome inactivation: Advances in understanding facultative heterochromatin formation. Nat. Rev. Genet. 2011, 12, 542–553. [Google Scholar] [CrossRef]
- Akhtar, A.; Becker, P.B. Activation of transcription through histone h4 acetylation by Mof, an acetyltransferase essential for dosage compensation in drosophila. Mol. Cell 2000, 5, 367–375. [Google Scholar] [CrossRef]
- Lucchesi, J.C. Dosage compensation indrosophila and the “Complex” World of transcriptional regulation. BioEssays 1996, 18, 541–547. [Google Scholar] [CrossRef]
- Smith, E.R.; Pannuti, A.; Gu, W.; Steurnagel, A.; Cook, R.G.; Allis, C.D.; Lucchesi, J.C. The drosophila Msl complex acetylates histone h4 at lysine 16, a chromatin modification linked to dosage compensation. Mol. Cell. Biol. 2000, 20, 312–318. [Google Scholar] [CrossRef]
- Franke, A.; Baker, B.S. The rox1 and rox2 rnas are essential components of the compensasome, which mediates dosage compensation in drosophila. Mol. Cell 1999, 4, 117–122. [Google Scholar] [CrossRef]
- Meller, V.H.; Rattner, B.P. The rox genes encode redundant male-specific lethal transcripts required for targeting of the Msl complex. EMBO J. 2002, 21, 1084–1091. [Google Scholar] [CrossRef]
- Pandey, R.R.; Mondal, T.; Mohammad, F.; Enroth, S.; Redrup, L.; Komorowski, J.; Nagano, T.; Mancini-DiNardo, D.; Kanduri, C. Kcnq1ot1 antisense noncoding rna mediates lineage-specific transcriptional silencing through chromatin-level regulation. Mol. Cell 2008, 32, 232–246. [Google Scholar] [CrossRef]
- Latos, P.A.; Pauler, F.M.; Koerner, M.V.; Şenergin, H.B.; Hudson, Q.J.; Stocsits, R.R.; Allhoff, W.; Stricker, S.H.; Klement, R.M.; Warczok, K.E.; et al. Airn transcriptional overlap, but not its lncRNA products, induces imprinted IGFgf2R silencing. Science (N.Y.) 2012, 338, 1469–1472. [Google Scholar] [CrossRef]
- Cipolla, G.A.; de Oliveira, J.C.; Salviano-Silva, A.; Lobo-Alves, S.C.; Lemos, D.S.; Oliveira, L.C.; Jucoski, T.S.; Mathias, C.; Pedroso, G.A.; Zambalde, E.P.; et al. Long non-coding RNAs in multifactorial diseases: Another layer of complexity. Non-Coding RNA 2018, 4, 13. [Google Scholar] [CrossRef]
- Chen, G.; Wang, Z.; Wang, D.; Qiu, C.; Liu, M.; Chen, X.; Zhang, Q.; Yan, G.; Cui, Q. LncRNA disease: A database for long-non-coding RNA-associated diseases. Nucleic Acids Res. 2012, 41, D983–D986. [Google Scholar] [CrossRef]
- Magistri, M.; Velmeshev, D.; Makhmutova, M.; Faghihi, M.A. Transcriptomics profiling of Alzheimer’s disease reveal neurovascular defects, altered amyloid-β homeostasis, and deregulated expression of long noncoding RNAs. J. Alzheimers Dis. 2015, 48, 647–665. [Google Scholar] [CrossRef]
- Johnson, R.; Richter, N.; Jauch, R.; Gaughwin, P.M.; Zuccato, C.; Cattaneo, E.; Stanton, L.W. Human accelerated region 1 noncoding rna is repressed by rest in huntington’s disease. Physiol. Genom. 2010, 41, 269–274. [Google Scholar] [CrossRef]
- Ni, Y.; Huang, H.; Chen, Y.; Cao, M.; Zhou, H.; Zhang, Y. Investigation of long non-coding RNA expression profiles in the substantia Nigra of Parkinson’s disease. Cell. Mol. Neurobiol. 2017, 37, 329–338. [Google Scholar] [CrossRef]
- Cui, X.; Niu, W.; Kong, L.; He, M.; Jiang, K.; Chen, S.; Zhong, A.; Zhang, Q.; Li, W.; Lu, J.; et al. Long noncoding RNA as an indicator differentiating schizophrenia from major depressive disorder and generalized anxiety disorder in nonpsychiatric hospital. Biomark. Med. 2017, 11, 221–228. [Google Scholar] [CrossRef]
- Sakamoto, K.; Crowley, J.J. A comprehensive review of the genetic and biological evidence supports a role for microrna-137 in the etiology of schizophrenia. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2018, 177, 242–256. [Google Scholar] [CrossRef]
- Wilkinson, B.; Campbell, D.B. Contribution of long noncoding RNAs to autism spectrum disorder risk. Int. Rev. Neurobiol. 2013, 113, 35–59. [Google Scholar] [CrossRef]
- Ziats, M.N.; Rennert, O.M. Aberrant expression of long noncoding rnas in autistic brain. J. Mol. Neurosci. MN 2013, 49, 589–593. [Google Scholar] [CrossRef]
- Kumarswamy, R.; Bauters, C.; Volkmann, I.; Maury, F.; Fetisch, J.; Holzmann, A.; Lemesle, G.; de Groote, P.; Pinet, F.; Thum, T. Circulating long noncoding rna, lipcar, predicts survival in patients with heart failure. Circ. Res 2014, 114, 1569–1575. [Google Scholar] [CrossRef]
- Zhou, X.; Zhang, W.; Jin, M.; Chen, J.; Xu, W.; Kong, X. LncRNA MIAT functions as a competing endogenous RNA to upregulate DAPK2 by sponging miR-22-3p in diabetic cardiomyopathy. Cell Death Dis. 2017, 8, e2929. [Google Scholar] [CrossRef]
- Kim, D.K.; Zhang, L.; Dzau, V.J.; Pratt, R.E. H19, a developmentally regulated gene, is reexpressed in RAT vascular smooth muscle cells after injury. J. Clin. Investig. 1994, 93, 355–360. [Google Scholar] [CrossRef]
- Yan, Y.; Zhang, B.; Liu, N.; Qi, C.; Xiao, Y.; Tian, X.; Li, T.; Liu, B. Circulating long noncoding RNA UCA1 as a novel biomarker of acute myocardial infarction. Biomed. Res. Int. 2016, 2016, 8079372. [Google Scholar] [CrossRef]
- He, X.; Ou, C.; Xiao, Y.; Han, Q.; Li, H.; Zhou, S. LncRNAs: Key players and novel insights into diabetes mellitus. Oncotarget 2017, 8, 71325–71341. [Google Scholar] [CrossRef]
- Li, L.; Liu, B.; Wapinski, O.L.; Tsai, M.C.; Qu, K.; Zhang, J.; Carlson, J.C.; Lin, M.; Fang, F.; Gupta, R.A.; et al. Targeted disruption of hotair leads to homeotic transformation and gene derepression. Cell Rep. 2013, 5, 3–12. [Google Scholar] [CrossRef]
- Amandio, A.R.; Necsulea, A.; Joye, E.; Mascrez, B.; Duboule, D. Hotair is dispensible for mouse development. PLoS Genet. 2016, 12, e1006232. [Google Scholar] [CrossRef]
- Gupta, R.A.; Shah, N.; Wang, K.C.; Kim, J.; Horlings, H.M.; Wong, D.J.; Tsai, M.C.; Hung, T.; Argani, P.; Rinn, J.L.; et al. Long non-coding RNA Hotair reprograms chromatin state to promote cancer metastasis. Nature 2010, 464, 1071–1076. [Google Scholar] [CrossRef]
- Zemmour, D.; Pratama, A.; Loughhead, S.M.; Mathis, D.; Benoist, C. Flicr, a long noncoding RNA, modulates FOXP3 expression and autoimmunity. Proc. Natl. Acad. Sci. USA 2017, 114, E3472–E3480. [Google Scholar] [CrossRef]
- Ahn, R.; Gupta, R.; Lai, K.; Chopra, N.; Arron, S.T.; Liao, W. Network analysis of psoriasis reveals biological pathways and roles for coding and long non-coding RNAs. BMC Genom. 2016, 17, 841. [Google Scholar] [CrossRef]
- Jiang, H.; Qin, X.J.; Li, W.P.; Ma, R.; Wang, T.; Li, Z.Q. LncRNAs expression in adjuvant-induced arthritis rats reveals the potential role of lncRNAs contributing to rheumatoid arthritis pathogenesis. Gene 2016, 593, 131–142. [Google Scholar] [CrossRef]
- Chen, D.; Liu, J.; Zhao, H.Y.; Chen, Y.P.; Xiang, Z.; Jin, X. Plasma long noncoding RNA expression profile identified by microarray in patients with Crohn’s disease. World J. Gastroenterol. 2016, 22, 4716–4731. [Google Scholar] [CrossRef]
- Chew, C.L.; Conos, S.A.; Unal, B.; Tergaonkar, V. Noncoding RNAs: Master regulators of inflammatory signaling. Trends Mol. Med. 2018, 24, 66–84. [Google Scholar] [CrossRef]
- Mathy, N.W.; Chen, X.-M. Long non-coding rnas (lncrnas) and their transcriptional control of inflammatory responses. J. Biol. Chem. 2017, 292, 12375–12382. [Google Scholar] [CrossRef]
- Fanucchi, S.; Fok, E.T.; Dalla, E.; Shibayama, Y.; Borner, K.; Chang, E.Y.; Stoychev, S.; Imakaev, M.; Grimm, D.; Wang, K.C.; et al. Immune genes are primed for robust transcription by proximal long noncoding rnas located in nuclear compartments. Nat. Gent. 2018. [Google Scholar] [CrossRef]
- Ilott, N.E.; Heward, J.A.; Roux, B.; Tsitsiou, E.; Fenwick, P.S.; Lenzi, L.; Goodhead, I.; Hertz-Fowler, C.; Heger, A.; Hall, N.; et al. Corrigendum: Long non-coding rnas and enhancer rnas regulate the lipopolysaccharide-induced inflammatory response in human monocytes. Nat. Commun. 2015, 6, 6814. [Google Scholar] [CrossRef]
- Ma, S.; Ming, Z.; Gong, A.-Y.; Wang, Y.; Chen, X.; Hu, G.; Zhou, R.; Shibata, A.; Swanson, P.C.; Chen, X.-M. A long noncoding RNA, lincRNA-Tnfaip3, acts as a coregulator of NF-κB to modulate inflammatory gene transcription in mouse macrophages. FASEB J. 2017, 31, 1215–1225. [Google Scholar] [CrossRef]
- Rapicavoli, N.A.; Qu, K.; Zhang, J.; Mikhail, M.; Laberge, R.M.; Chang, H.Y. A mammalian pseudogene lncrna at the interface of inflammation and anti-inflammatory therapeutics. eLife 2013, 2, e00762. [Google Scholar] [CrossRef]
- Li, Z.; Chao, T.C.; Chang, K.Y.; Lin, N.; Patil, V.S.; Shimizu, C.; Head, S.R.; Burns, J.C.; Rana, T.M. The long noncoding RNA THRIL regulates TNFα expression through its interaction with hnRNPL. Proc. Natl. Acad. Sci. USA 2014, 111, 1002–1007. [Google Scholar] [CrossRef]
- Krawczyk, M.; Emerson, B.M. P50-associated COX-2 extragenic RNA (PACER) activates COX-2 gene expression by occluding repressive NF-κB complexes. eLife 2014, 3. [Google Scholar] [CrossRef]
- Carpenter, S.; Aiello, D.; Atianand, M.K.; Ricci, E.P.; Gandhi, P.; Hall, L.L.; Byron, M.; Monks, B.; Henry-Bezy, M.; Lawrence, J.B.; et al. A long noncoding RNA mediates both activation and repression of immune response genes. Science 2013, 341, 789–792. [Google Scholar] [CrossRef]
- Muxel, S.M.; Acuna, S.M.; Aoki, J.I.; Zampieri, R.A.; Floeter-Winter, L.M. Toll-like receptor and miRNA-Let-7e expression alter the inflammatory response in leishmania amazonensis-infected macrophages. Front. Immunol. 2018, 9, 2792. [Google Scholar] [CrossRef]
- Chan, J.; Atianand, M.; Jiang, Z.; Carpenter, S.; Aiello, D.; Elling, R.; Fitzgerald, K.A.; Caffrey, D.R. Cutting edge: A natural antisense transcript, AS-IL1α, controls inducible transcription of the proinflammatory cytokine IL-1α. J. Immunol. 2015, 195, 1359–1363. [Google Scholar] [CrossRef]
- Peng, X.; Gralinski, L.; Armour, C.D.; Ferris, M.T.; Thomas, M.J.; Proll, S.; Bradel-Tretheway, B.G.; Korth, M.J.; Castle, J.C.; Biery, M.C.; et al. Unique signatures of long noncoding RNA expression in response to virus infection and altered innate immune signaling. mBio 2010, 1. [Google Scholar] [CrossRef]
- Collier, S.P.; Collins, P.L.; Williams, C.L.; Boothby, M.R.; Aune, T.M. Cutting edge: Influence of Tmevpg1, a long intergenic noncoding RNA, on the expression of ifng by Th1 cells. J. Immunol. 2012, 189, 2084–2088. [Google Scholar] [CrossRef]
- Muxel, S.M.; Laranjeira-Silva, M.F.; Zampieri, R.A.; Floeter-Winter, L.M. Leishmania (leishmania) amazonensis induces macrophage miR-294 and miR-721 expression and modulates infection by targeting NOS2 and L-arginine metabolism. Sci. Rep. 2017, 7, 44141. [Google Scholar] [CrossRef]
- Duval, M.; Cossart, P.; Lebreton, A. Mammalian microRNAs and long noncoding RNAs in the host-bacterial pathogen crosstalk. Semin. Cell Dev. Biol. 2017, 65, 11–19. [Google Scholar] [CrossRef] [Green Version]
- Zur Bruegge, J.; Einspanier, R.; Sharbati, S. A long journey ahead: Long non-coding RNAs in bacterial infections. Front. Cell. Infect. Microbiol. 2017, 7, 95. [Google Scholar] [CrossRef]
- He, J.; Ou, Q.; Liu, C.; Shi, L.; Zhao, C.; Xu, Y.; Kong, S.K.; Loo, J.F.C.; Li, B.; Gu, D. Differential expression of long non-coding RNAs in patients with tuberculosis infection. Tuberculosis 2017, 107, 73–79. [Google Scholar] [CrossRef]
- Yang, X.; Yang, J.; Wang, J.; Wen, Q.; Wang, H.; He, J.; Hu, S.; He, W.; Du, X.; Liu, S.; et al. Microarray analysis of long noncoding RNA and mrna expression profiles in human macrophages infected with Mycobacterium tuberculosis. Sci. Rep. 2016, 6, 38963. [Google Scholar] [CrossRef]
- Yi, Z.; Li, J.; Gao, K.; Fu, Y. Identifcation of differentially expressed long non-coding RNAs in CD4+ T cells response to latent tuberculosis infection. J. Infect. 2014, 69, 558–568. [Google Scholar] [CrossRef]
- Fu, Y.; Xu, X.; Xue, J.; Duan, W.; Yi, Z. Deregulated lncRNAs in B cells from patients with active tuberculosis. PLoS ONE 2017, 12, e0170712. [Google Scholar] [CrossRef]
- Jiang, T.T.; Wei, L.L.; Shi, L.Y.; Chen, Z.L.; Wang, C.; Liu, C.M.; Li, Z.J.; Li, J.C. Microarray expression profile analysis of mRNAs and long non-coding RNAs in pulmonary tuberculosis with different traditional chinese medicine syndromes. BMC Complement. Altern. Med. 2016, 16, 472. [Google Scholar] [CrossRef]
- Westermann, A.J.; Förstner, K.U.; Amman, F.; Barquist, L.; Chao, Y.; Schulte, L.N.; Müller, L.; Reinhardt, R.; Stadler, P.F.; Vogel, J. Dual RNA-seq unveils noncoding RNA functions in host–pathogen interactions. Nature 2016, 529, 496–501. [Google Scholar] [CrossRef]
- Gomez, J.A.; Wapinski, O.L.; Yang, Y.W.; Bureau, J.F.; Gopinath, S.; Monack, D.M.; Chang, H.Y.; Brahic, M.; Kirkegaard, K. The nest long ncRNA controls microbial susceptibility and epigenetic activation of the interferon-γ locus. Cell 2013, 152, 743–754. [Google Scholar] [CrossRef]
- Yang, R.; Huang, F.; Fu, J.; Dou, B.; Xu, B.; Miao, L.; Liu, W.; Yang, X.; Tan, C.; Chen, H.; et al. Differential transcription profiles of long non-coding RNAs in primary human brain microvascular endothelial cells in response to meningitic Escherichia coli. Sci. Rep-Uk 2016, 6, 38903. [Google Scholar] [CrossRef]
- Zhu, H.; Wang, Q.; Yao, Y.; Fang, J.; Sun, F.; Ni, Y.; Shen, Y.; Wang, H.; Shao, S. Microarray analysis of long non-coding RNA expression profiles in human gastric cells and tissues with Helicobacter Pylori infection. BMC Med. Genom. 2015, 8, 84. [Google Scholar] [CrossRef]
- Yang, L.; Long, Y.; Li, C.; Cao, L.; Gan, H.; Huang, K.; Jia, Y. Genome-wide analysis of long noncoding RNA profile in human gastric epithelial cell response to Helicobacter pylori. Jpn. J. Infect. Dis. 2015, 68, 63–66. [Google Scholar] [CrossRef]
- Kaakoush, N.O.; Deshpande, N.P.; Man, S.M.; Burgos-Portugal, J.A.; Khattak, F.A.; Raftery, M.J.; Wilkins, M.R.; Mitchell, H.M. Transcriptomic and proteomic analyses reveal key innate immune signatures in the host response to the gastrointestinal pathogen campylobacter concisus. Infect. Immun. 2015, 83, 832–845. [Google Scholar] [CrossRef]
- Zhang, Q.; Chen, C.Y.; Yedavalli, V.S.; Jeang, K.T. Neat1 long noncoding RNA and paraspeckle bodies modulate HIV-1 posttranscriptional expression. mBio 2013, 4, e00596-12. [Google Scholar] [CrossRef]
- Willingham, A.T.; Orth, A.P.; Batalov, S.; Peters, E.C.; Wen, B.G.; Aza-Blanc, P.; Hogenesch, J.B.; Schultz, P.G. A strategy for probing the function of noncoding RNAs finds a repressor of NFAT. Science 2005, 309, 1570–1573. [Google Scholar] [CrossRef]
- Imam, H.; Bano, A.S.; Patel, P.; Holla, P.; Jameel, S. The lncRNA NRON modulates HIV-1 replication in a NFAT-dependent manner and is differentially regulated by early and late viral proteins. Sci. Rep. 2015, 5, 8639. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Huang, L.; Wei, Q.; Zhang, Y.; Zhang, S.; Zhang, W.; Cai, L.; Liang, S. Microarray analysis of long non-coding RNA expression profiles uncovers a toxoplasma-induced negative regulation of host immune signaling. Parasit. Vectors 2018, 11, 174. [Google Scholar] [CrossRef]
- Menard, K.L.; Haskins, B.E.; Colombo, A.P.; Denkers, E.Y. Toxoplasma gondii manipulates expression of host long noncoding RNA during intracellular infection. Sci. Rep. 2018, 8, 15017. [Google Scholar] [CrossRef]
- Vasconcelos, E.J.R.; daSilva, L.F.; Pires, D.S.; Lavezzo, G.M.; Pereira, A.S.A.; Amaral, M.S.; Verjovski-Almeida, S. The Schistosoma mansoni genome encodes thousands of long non-coding RNAs predicted to be functional at different parasite life-cycle stages. Sci. Rep. 2017, 7, 10508. [Google Scholar] [CrossRef]
- Liao, Q.; Zhang, Y.; Zhu, Y.; Chen, J.; Dong, C.; Tao, Y.; He, A.; Liu, J.; Wu, Z. Identification of long noncoding RNAs in Schistosoma mansoni and Schistosoma japonicum. Exp. Parasitol. 2018, 191, 82–87. [Google Scholar] [CrossRef]
- Oliveira, V.F.; Moares, L.A.G.; Mota, E.A.; Jannotti-Passos, L.K.; Coelho, P.M.Z.; Mattos, A.C.A.; Couto, F.F.B.; Caffrey, B.E.; Marsico, A.; Guerra-Sa, R. Identification of 170 new long noncoding RNAs in Schistosoma mansoni. BioMed Res. Int. 2018, 2018, 1264697. [Google Scholar] [CrossRef]
- Chacko, N.; Zhao, Y.; Yang, E.; Wang, L.; Cai, J.J.; Lin, X. The lncRNA RZE1 controls cryptococcal morphological transition. PLoS Genet. 2015, 11, e1005692. [Google Scholar] [CrossRef]
- Patankar, S.; Munasinghe, A.; Shoaibi, A.; Cummings, L.M.; Wirth, D.F. Serial analysis of gene expression in Plasmodium Falciparum reveals the global expression profile of erythrocytic stages and the presence of anti-sense transcripts in the malarial parasite. Mol. Biol. Cell 2001, 12, 3114–3125. [Google Scholar] [CrossRef]
- Militello, K.T.; Patel, V.; Chessler, A.D.; Fisher, J.K.; Kasper, J.M.; Gunasekera, A.; Wirth, D.F. RNA polymerase II synthesizes antisense RNA in Plasmodium falciparum. RNA 2005, 11, 365–370. [Google Scholar] [CrossRef]
- Liniger, M.; Bodenmuller, K.; Pays, E.; Gallati, S.; Roditi, I. Overlapping sense and antisense transcription units in Trypanosoma brucei. Mol. Microbiol. 2001, 40, 869–878. [Google Scholar] [CrossRef]
- Belli, S.I.; Monnerat, S.; Schaff, C.; Masina, S.; Noll, T.; Myler, P.J.; Stuart, K.; Fasel, N. Sense and antisense transcripts in the histone H1 (his-1) locus of Leishmania major. Int. J. Parasitol. 2003, 33, 965–975. [Google Scholar] [CrossRef]
- Dumas, C.; Chow, C.; Muller, M.; Papadopoulou, B. A novel class of developmentally regulated noncoding RNAs in Leishmania. Eukaryot. Cell 2006, 5, 2033–2046. [Google Scholar] [CrossRef]
- von Allmen, N.; Bienz, M.; Hemphill, A.; Muller, N. Quantitative assessment of sense and antisense transcripts from genes involved in antigenic variation (VSP genes) and encystation (CWP 1 gene) of Giardia Lamblia clone GS/M-83-h7. Parasitology 2005, 130, 389–396. [Google Scholar] [CrossRef]
- Teodorovic, S.; Walls, C.D.; Elmendorf, H.G. Bidirectional transcription is an inherent feature of Giardia Lamblia promoters and contributes to an abundance of sterile antisense transcripts throughout the genome. Nucleic Acids Res. 2007, 35, 2544–2553. [Google Scholar] [CrossRef]
- Woehle, C.; Kusdian, G.; Radine, C.; Graur, D.; Landan, G.; Gould, S.B. The parasite trichomonas vaginalis expresses thousands of pseudogenes and long non-coding RNAs independently from functional neighbouring genes. BMC Genom. 2014, 15, 906. [Google Scholar] [CrossRef]
- Broadbent, K.M.; Broadbent, J.C.; Ribacke, U.; Wirth, D.; Rinn, J.L.; Sabeti, P.C. Strand-specific RNA sequencing in Plasmodium falciparum malaria identifies developmentally regulated long non-coding RNA and circular RNA. BMC Genom. 2015, 16, 454. [Google Scholar] [CrossRef]
- Liao, Q.; Shen, J.; Liu, J.; Sun, X.; Zhao, G.; Chang, Y.; Xu, L.; Li, X.; Zhao, Y.; Zheng, H.; et al. Genome-wide identification and functional annotation of Plasmodium falciparum long noncoding RNAs from RNA-seq data. Parasitol. Res. 2014, 113, 1269–1281. [Google Scholar] [CrossRef]
- Broadbent, K.M.; Park, D.; Wolf, A.R.; Van Tyne, D.; Sims, J.S.; Ribacke, U.; Volkman, S.; Duraisingh, M.; Wirth, D.; Sabeti, P.C.; et al. A global transcriptional analysis of Plasmodium falciparum malaria reveals a novel family of telomere-associated lncRNAs. Genome Biol. 2011, 12, R56. [Google Scholar] [CrossRef]
- Vembar, S.S.; Scherf, A.; Siegel, T.N. Noncoding RNAs as emerging regulators of Plasmodium falciparum virulence gene expression. Curr. Opin. Microbiol. 2014, 20, 153–161. [Google Scholar] [CrossRef]
- Guizetti, J.; Barcons-Simon, A.; Scherf, A. Trans-acting GC-rich non-coding RNA at var expression site modulates gene counting in malaria parasite. Nucleic Acids Res. 2016, 44, 9710–9718. [Google Scholar] [CrossRef]
- Pawar, H.; Pai, K.; Patole, M.S. A novel protein coding potential of long intergenic non-coding RNAs (lincRNAs) in the Kinetoplastid Protozoan parasite Leishmania major. Acta Trop. 2017, 167, 21–25. [Google Scholar] [CrossRef]
- Freitas Castro, F.; Ruy, P.C.; Nogueira Zeviani, K.; Freitas Santos, R.; Simoes Toledo, J.; Kaysel Cruz, A. Evidence of putative non-coding RNAs from Leishmania untranslated regions. Mol. Biochem. Parasitol. 2017, 214, 69–74. [Google Scholar] [CrossRef]
- Torres, F.; Arias-Carrasco, R.; Caris-Maldonado, J.C.; Barral, A.; Maracaja-Coutinho, V.; De Queiroz, A.T.L. Leishdb: A database of coding gene annotation and non-coding RNAs in Leishmania braziliensis. Database (Oxford) 2017, 2017. [Google Scholar] [CrossRef]
- Laranjeira-Silva, M.F.; Zampieri, R.A.; Muxel, S.M.; Beverley, S.M.; Floeter-Winter, L.M. Leishmania amazonensis arginase compartmentalization in the glycosome is important for parasite infectivity. PLoS ONE 2012, 7, e34022. [Google Scholar] [CrossRef]
- Muxel, S.M.; Aoki, J.I.; Fernandes, J.C.R.; Laranjeira-Silva, M.F.; Zampieri, R.A.; Acuna, S.M.; Muller, K.E.; Vanderlinde, R.H.; Floeter-Winter, L.M. Arginine and polyamines fate in Leishmania infection. Front. Microbiol. 2018, 8, 2682. [Google Scholar] [CrossRef]
- Aoki, J.I.; Muxel, S.M.; Zampieri, R.A.; Laranjeira-Silva, M.F.; Muller, K.E.; Nerland, A.H.; Floeter-Winter, L.M. RNA-seq transcriptional profiling of Leishmania amazonensis reveals an arginase-dependent gene expression regulation. PLoS Negl. Trop. Dis. 2017, 11, e0006026. [Google Scholar] [CrossRef]
- Kolev, N.G.; Franklin, J.B.; Carmi, S.; Shi, H.; Michaeli, S.; Tschudi, C. The transcriptome of the human pathogen Trypanosoma brucei at single-nucleotide resolution. PLoS Pathog. 2010, 6, e1001090. [Google Scholar] [CrossRef]
- Cao, S.; Moss, W.; O’Grady, T.; Concha, M.; Strong, M.J.; Wang, X.; Yu, Y.; Baddoo, M.; Zhang, K.; Fewell, C.; et al. New noncoding lytic transcripts derived from the Epstein-Barr virus latency origin of replication, oriP, are hyperedited, bind the paraspeckle protein, NONO/p54nrb, and support viral lytic transcription. J. Virol. 2015, 89, 7120–7132. [Google Scholar] [CrossRef]
- Chavez-Calvillo, G.; Martin, S.; Hamm, C.; Sztuba-Solinska, J. The structure-to-function relationships of γherpesvirus-encoded long non-coding RNAs and their contributions to viral pathogenesis. Non-Coding RNA 2018, 4, 24. [Google Scholar] [CrossRef]
- Baglio, S.R.; van Eijndhoven, M.A.; Koppers-Lalic, D.; Berenguer, J.; Lougheed, S.M.; Gibbs, S.; Leveille, N.; Rinkel, R.N.; Hopmans, E.S.; Swaminathan, S.; et al. Sensing of latent EBV infection through exosomal transfer of 5′pppRNA. Proc. Natl. Acad. Sci. USA 2016, 113, E587–E596. [Google Scholar] [CrossRef] [Green Version]
- Lerner, M.R.; Andrews, N.C.; Miller, G.; Steitz, J.A. Two small rnas encoded by Epstein-Barr virus and complexed with protein are precipitated by antibodies from patients with systemic lupus erythematosus. Proc. Natl. Acad. Sci. USA 1981, 78, 805–809. [Google Scholar] [CrossRef]
- Rossetto, C.C.; Pari, G.S. Kaposi’s sarcoma-associated herpesvirus noncoding polyadenylated nuclear RNA interacts with virus- and host cell-encoded proteins and suppresses expression of genes involved in immune modulation. J. Virol. 2011, 85, 13290–13297. [Google Scholar] [CrossRef]
- Prensner, J.R.; Iyer, M.K.; Balbin, O.A.; Dhanasekaran, S.M.; Cao, Q.; Brenner, J.C.; Laxman, B.; Asangani, I.A.; Grasso, C.S.; Kominsky, H.D.; et al. Transcriptome sequencing across a prostate cancer cohort identifies PCAT-1, an unannotated lincRNA implicated in disease progression. Nat. Biotechnol. 2011, 29, 742–749. [Google Scholar] [CrossRef] [Green Version]
- Guo, L.; Zhao, Y.; Yang, S.; Zhang, H.; Wu, Q.; Chen, F. An integrated evolutionary analysis of miRNA-lncRNA in mammals. Mol. Biol. Rep. 2014, 41, 201–207. [Google Scholar] [CrossRef]
- Parasramka, M.A.; Maji, S.; Matsuda, A.; Yan, I.K.; Patel, T. Long non-coding RNAs as novel targets for therapy in hepatocellular carcinoma. Pharmacol. Therap. 2016, 161, 67–78. [Google Scholar] [CrossRef] [Green Version]
- Roberts, T.C.; Wood, M.J.A. Therapeutic targeting of non-coding RNAs. Essays Biochem. 2013, 54, 127–145. [Google Scholar] [CrossRef] [Green Version]
- Cha, D.J.; Franklin, J.L.; Dou, Y.; Liu, Q.; Higginbotham, J.N.; Demory Beckler, M.; Weaver, A.M.; Vickers, K.; Prasad, N.; Levy, S.; et al. KRAS-dependent sorting of miRNA to exosomes. eLife 2015, 4, e07197. [Google Scholar] [CrossRef]
- Hinger, S.A.; Cha, D.J.; Franklin, J.L.; Higginbotham, J.N.; Dou, Y.; Ping, J.; Shu, L.; Prasad, N.; Levy, S.; Zhang, B.; et al. Diverse long RNAs are differentially sorted into extracellular vesicles secreted by colorectal cancer cells. Cell Rep. 2018, 25, 715–725.e4. [Google Scholar] [CrossRef]
- Dou, Y.; Cha, D.J.; Franklin, J.L.; Higginbotham, J.N.; Jeppesen, D.K.; Weaver, A.M.; Prasad, N.; Levy, S.; Coffey, R.J.; Patton, J.G.; et al. Circular RNAs are down-regulated in KRAS mutant colon cancer cells and can be transferred to exosomes. Sci. Rep. 2016, 6, 37982. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, K.; Yan, I.K.; Haga, H.; Patel, T. Modulation of hypoxia-signaling pathways by extracellular linc-Ror. J. Cell Sci. 2014, 127, 1585–1594. [Google Scholar] [CrossRef]
- Mohankumar, S.; Patel, T. Extracellular vesicle long noncoding RNA as potential biomarkers of liver cancer. Brief. Funct. Genom. 2016, 15, 249–256. [Google Scholar] [CrossRef]
- Olivas, W.M.; Muhlrad, D.; Parker, R. Analysis of the yeast genome: Identification of new non-coding and small Orf-containing RNAs. Nucleic Acids Res. 1997, 25, 4619–4625. [Google Scholar] [CrossRef]
- Bazzini, A.A.; Johnstone, T.G.; Christiano, R.; Mackowiak, S.D.; Obermayer, B.; Fleming, E.S.; Vejnar, C.E.; Lee, M.T.; Rajewsky, N.; Walther, T.C.; et al. Identification of small Orfs in vertebrates using ribosome footprinting and evolutionary conservation. EMBO J. 2014, 33, 981–993. [Google Scholar] [CrossRef]
- Andrews, S.J.; Rothnagel, J.A. Emerging evidence for functional peptides encoded by short open reading frames. Nat. Rev. Genet. 2014, 15, 193–204. [Google Scholar] [CrossRef]
- Anderson, D.M.; Anderson, K.M.; Chang, C.-L.; Makarewich, C.A.; Nelson, B.R.; McAnally, J.R.; Kasaragod, P.; Shelton, J.M.; Liou, J.; Bassel-Duby, R.; et al. A micropeptide encoded by a putative long noncoding RNA regulates muscle performance. Cell 2015, 160, 595–606. [Google Scholar] [CrossRef]
- Nelson, B.R.; Makarewich, C.A.; Anderson, D.M.; Winders, B.R.; Troupes, C.D.; Wu, F.; Reese, A.L.; McAnally, J.R.; Chen, X.; Kavalali, E.T.; et al. A peptide encoded by a transcript annotated as long noncoding RNA enhances serca activity in muscle. Science 2016, 351, 271–275. [Google Scholar] [CrossRef]
- Matsumoto, A.; Pasut, A.; Matsumoto, M.; Yamashita, R.; Fung, J.; Monteleone, E.; Saghatelian, A.; Nakayama, K.I.; Clohessy, J.G.; Pandolfi, P.P. mTORC1 and muscle regeneration are regulated by the LINC00961-encoded spar polypeptide. Nature 2017, 541, 228–232. [Google Scholar] [CrossRef]
- Rion, N.; Rüegg, M.A. LncRNA-encoded peptides: More than translational noise? Cell Res. 2017, 27, 604–605. [Google Scholar] [CrossRef] [Green Version]
- Sun, Z.; Nair, A.; Chen, X.; Prodduturi, N.; Wang, J.; Kocher, J.-P. Uclncr: Ultrafast and comprehensive long non-coding rna detection from rna-seq. Sci. Rep-Uk 2017, 7, 14196. [Google Scholar] [CrossRef]
Figure 1.
Schematic representation of the genomic loci of different long non-coding RNA (lncRNA) and circular RNA (circRNA) biogenesis. (A) lncRNA classification depends on the genomic position: (i) intergenic RNAs (lincRNAs) are located between two protein coding-genes, (ii) intronic lncRNAs are positioned within an intronic region of a protein coding-gene, (iii) antisense lncRNAs (aslncRNAs) are transcribed from complementary strands, (iv) bidirectional lncRNAs originate from bidirectional transcription of protein-coding genes, and (v) enhancer RNAs (eRNAs) originate from enhancer regions and mediate transcription factor positioning into protein coding-genes promoters. (B) circRNAs are lncRNAs that undergo back splicing and can originate from transcripts containing only intronic, one or more exonic, or both intronic and exonic fragments (elciRNAs).
Figure 1.
Schematic representation of the genomic loci of different long non-coding RNA (lncRNA) and circular RNA (circRNA) biogenesis. (A) lncRNA classification depends on the genomic position: (i) intergenic RNAs (lincRNAs) are located between two protein coding-genes, (ii) intronic lncRNAs are positioned within an intronic region of a protein coding-gene, (iii) antisense lncRNAs (aslncRNAs) are transcribed from complementary strands, (iv) bidirectional lncRNAs originate from bidirectional transcription of protein-coding genes, and (v) enhancer RNAs (eRNAs) originate from enhancer regions and mediate transcription factor positioning into protein coding-genes promoters. (B) circRNAs are lncRNAs that undergo back splicing and can originate from transcripts containing only intronic, one or more exonic, or both intronic and exonic fragments (elciRNAs).

Figure 2.
LncRNA participation in gene expression regulation. Many types of lncRNA controls can occur to regulate gene expression, such as: (1) chromatin and chromosome condensation through histone modifications, (2) transcription factors (TF) direct recruitment, (3) binding to RNA polymerase (pol) II, (4) alternative splicing, (5) mRNA stability, (6) miRNA availability, (7) polysomes recruitment, and (8) modulation of gene expression in neighbor cells through packaging into extracellular vesicles.
Figure 2.
LncRNA participation in gene expression regulation. Many types of lncRNA controls can occur to regulate gene expression, such as: (1) chromatin and chromosome condensation through histone modifications, (2) transcription factors (TF) direct recruitment, (3) binding to RNA polymerase (pol) II, (4) alternative splicing, (5) mRNA stability, (6) miRNA availability, (7) polysomes recruitment, and (8) modulation of gene expression in neighbor cells through packaging into extracellular vesicles.

Figure 3.
Interaction between microRNAs (miRNAs) and lncRNAs. (A) lncRNAs can act as miRNA sponges in both linear or circular forms, thus reducing interaction between miRNA-target mRNA; (B) lncRNA can harbor miRNA precursors, producing mature miRNAs after Dicer and/or Drosha cleavage; (C) miRNA can target lncRNA for degradation, similar to 3’ untranslated region binding to mRNAs; and (D) lncRNA–miRNA competition for the mRNA binding site. circRNA—circular lncRNA; AGO—argonaute proteins; miRISC—miRNA-induced silencing complex; Dicer—endoribonuclease; Drosha—endoribonuclease.
Figure 3.
Interaction between microRNAs (miRNAs) and lncRNAs. (A) lncRNAs can act as miRNA sponges in both linear or circular forms, thus reducing interaction between miRNA-target mRNA; (B) lncRNA can harbor miRNA precursors, producing mature miRNAs after Dicer and/or Drosha cleavage; (C) miRNA can target lncRNA for degradation, similar to 3’ untranslated region binding to mRNAs; and (D) lncRNA–miRNA competition for the mRNA binding site. circRNA—circular lncRNA; AGO—argonaute proteins; miRISC—miRNA-induced silencing complex; Dicer—endoribonuclease; Drosha—endoribonuclease.

Figure 4.
Patho-physiological implications of lncRNAs expression in a time- and infection-dependent manner. (A) Expression of HOTAIR complexed with PRC2 and LSD1 leads to trimethylation of the histone 3 at the lysine 27 and demethylation of the lysine 4, controlling gene expression during embryonic development, but leading to breast cancer metastasis when expressed in adult breast tissue; (B) disruption of the NRON lncRNA expression in T CD4+ lymphocyte by the HIV Nef protein leads to NFAT translocation to the nucleus and LTR transcription. Abbreviations: HOTAIR—Hox transcript antisense intergenic RNA; PRC2—polycomb repressive complex; LSD1—lysine-specific histone demethylase 1; Me—methyl groups; H—histone; K—lysine; NEF—negative regulatory factor, NRON—non-coding repressor of NFAT; NFAT—nuclear factor of activated T-cells; HIV—human immunodeficiency virus; LTR—long terminal repeat; T CD4—CD4 T lymphocytes.
Figure 4.
Patho-physiological implications of lncRNAs expression in a time- and infection-dependent manner. (A) Expression of HOTAIR complexed with PRC2 and LSD1 leads to trimethylation of the histone 3 at the lysine 27 and demethylation of the lysine 4, controlling gene expression during embryonic development, but leading to breast cancer metastasis when expressed in adult breast tissue; (B) disruption of the NRON lncRNA expression in T CD4+ lymphocyte by the HIV Nef protein leads to NFAT translocation to the nucleus and LTR transcription. Abbreviations: HOTAIR—Hox transcript antisense intergenic RNA; PRC2—polycomb repressive complex; LSD1—lysine-specific histone demethylase 1; Me—methyl groups; H—histone; K—lysine; NEF—negative regulatory factor, NRON—non-coding repressor of NFAT; NFAT—nuclear factor of activated T-cells; HIV—human immunodeficiency virus; LTR—long terminal repeat; T CD4—CD4 T lymphocytes.

© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Fernandes, J.C.R.; Acuña, S.M.; Aoki, J.I.; Floeter-Winter, L.M.; Muxel, S.M. Long Non-Coding RNAs in the Regulation of Gene Expression: Physiology and Disease. Non-Coding RNA 2019, 5, 17. https://doi.org/10.3390/ncrna5010017
AMA Style
Fernandes JCR, Acuña SM, Aoki JI, Floeter-Winter LM, Muxel SM. Long Non-Coding RNAs in the Regulation of Gene Expression: Physiology and Disease. Non-Coding RNA. 2019; 5(1):17. https://doi.org/10.3390/ncrna5010017
Chicago/Turabian StyleFernandes, Juliane C. R., Stephanie M. Acuña, Juliana I. Aoki, Lucile M. Floeter-Winter, and Sandra M. Muxel. 2019. "Long Non-Coding RNAs in the Regulation of Gene Expression: Physiology and Disease" Non-Coding RNA 5, no. 1: 17. https://doi.org/10.3390/ncrna5010017
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.