
The Persistence of Bacterial Pathogens in Surface Water and Its Impact on Global Food Safety

 ,
,
Abstract
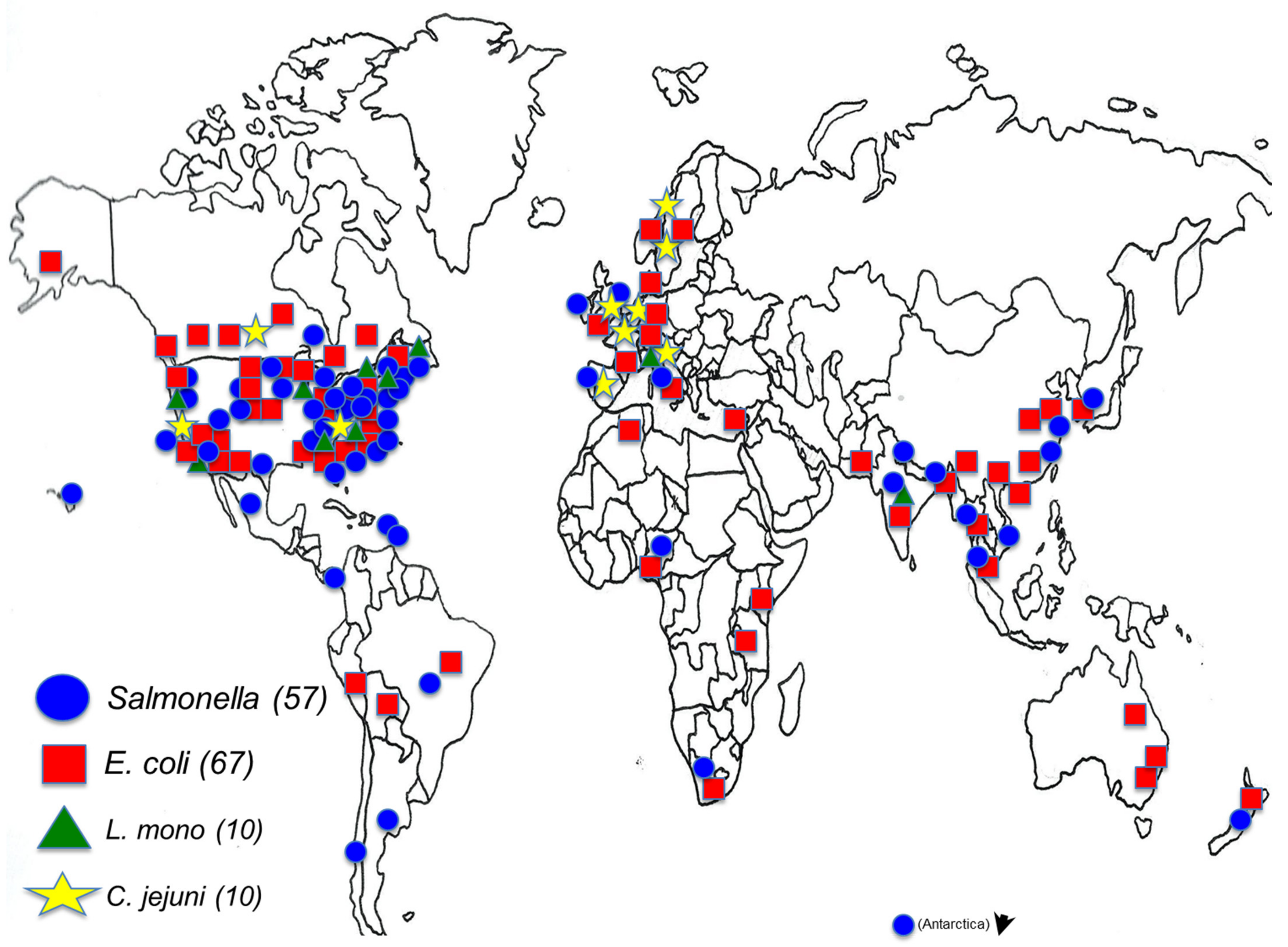
:1. Introduction
2. The Pathogens
2.1. Pathogenic E. coli: O157 and Other Shiga Toxin-Producing E. coli (STEC)
2.2. Salmonella
2.3. Campylobacter and Emerging Arcobacter
2.4. Listeria Monocytogenes
3. Antimicrobial Resistance (AMR) in Environmental Waters
4. The Role of Genomics in Detection, Traceability, and Characterization of Enteric Bacterial Pathogens Associated with Water
4.1. Whole Genome Sequencing (WGS)
4.2. Metagenomics
4.3. Long-Read Sequencing
5. Global Initiatives
5.1. Latin American Water Study
5.2. Global Water and Food Safety Summit
6. Advances in Methods and Preventive Measures
6.1. Water Collection Methods
6.2. Development of Preventive Measures
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- European Food Safety Authority; European Centre for Disease Prevention and Control. The European Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2014. EFSA J. 2015, 13, 4329. [Google Scholar] [CrossRef]
- Tack, D.M.; Marder, E.P.; Griffin, P.M.; Cieslak, P.R.; Dunn, J.; Hurd, S.; Scallan, E.; Lathrop, S.; Muse, A.; Ryan, P.; et al. Preliminary Incidence and Trends of Infections with Pathogens Transmitted Commonly Through Food—Foodborne Diseases Active Surveillance Network, 10 U.S. Sites, 2015–2018. MMWR Morb. Mortal. Wkly. Rep. 2019, 68, 369–373. [Google Scholar] [CrossRef] [Green Version]
- FDA. FSMA Final Rule on Produce Safety (Federal Register Notice). Available online: https://www.govinfo.gov/content/pkg/FR-2015-11-27/pdf/2015-28159.pdf (accessed on 3 March 2021).
- Bell, R.L.; Zheng, J.; Burrows, E.; Allard, S.; Wang, C.Y.; Keys, C.E.; Melka, D.C.; Strain, E.; Luo, Y.; Allard, M.W.; et al. Ecological prevalence, genetic diversity, and epidemiological aspects of Salmonella isolated from tomato agricultural regions of the Virginia Eastern Shore. Front. Microbiol. 2015, 6, 415. [Google Scholar] [CrossRef] [PubMed]
- Micallef, S.A.; Goldstein, R.E.R.; George, A.; Kleinfelter, L.; Boyer, M.S.; McLaughlin, C.R.; Estrin, A.; Ewing, L.; Beaubrun, J.J.-G.; Hanes, D.E.; et al. Occurrence and antibiotic resistance of multiple Salmonella serotypes recovered from water, sediment and soil on mid-Atlantic tomato farms. Environ. Res. 2012, 114, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Cooley, M.B.; Quinones, B.; Oryang, D.; Mandrell, R.E.; Gorski, L. Prevalence of shiga toxin producing Escherichia coli, Salmonella enterica, and Listeria monocytogenes at public access watershed sites in a California Central Coast agricultural region. Front. Cell. Infect. Microbiol. 2014, 4, 30. [Google Scholar] [CrossRef] [Green Version]
- Solaiman, S.; Allard, S.M.; Callahan, M.T.; Jiang, C.; Handy, E.; East, C.; Haymaker, J.; Bui, A.; Craddock, H.; Murray, R.; et al. Longitudinal Assessment of the Dynamics of Escherichia coli, Total Coliforms, Enterococcus spp., and Aeromonas spp. in Alternative Irrigation Water Sources: A CONSERVE Study. Appl. Environ. Microbiol. 2020, 86. [Google Scholar] [CrossRef]
- Sharma, M.; Handy, E.T.; East, C.L.; Kim, S.; Jiang, C.; Callahan, M.T.; Allard, S.M.; Micallef, S.; Craighead, S.; Anderson-Coughlin, B.; et al. Prevalence of Salmonella and Listeria monocytogenes in non-traditional irrigation waters in the Mid-Atlantic United States is affected by water type, season, and recovery method. PLoS ONE 2020, 15, e0229365. [Google Scholar] [CrossRef] [PubMed]
- Allard, S.M.; Callahan, M.T.; Bui, A.; Ferelli, A.M.C.; Chopyk, J.; Chattopadhyay, S.; Mongodin, E.F.; Micallef, S.A.; Sapkota, A.R. Creek to Table: Tracking fecal indicator bacteria, bacterial pathogens, and total bacterial communities from irrigation water to kale and radish crops. Sci. Total Environ. 2019, 666, 461–471. [Google Scholar] [CrossRef] [PubMed]
- Haymaker, J.; Sharma, M.; Parveen, S.; Hashem, F.; May, E.B.; Handy, E.T.; White, C.; East, C.; Bradshaw, R.; Micallef, S.A.; et al. Prevalence of Shiga-toxigenic and atypical enteropathogenic Escherichia coli in untreated surface water and reclaimed water in the Mid-Atlantic U.S. Environ. Res. 2019, 172, 630–636. [Google Scholar] [CrossRef] [PubMed]
- Center for Food Safety and Applied Nutrition. Environmental Studies. Available online: https://www.fda.gov/food/science-research-food/environmental-studies (accessed on 8 March 2021).
- Decol, L.T.; Casarin, L.S.; Hessel, C.T.; Batista, A.C.F.; Allende, A.; Tondo, E.C. Microbial quality of irrigation water used in leafy green production in Southern Brazil and its relationship with produce safety. Food Microbiol. 2017, 65, 105–113. [Google Scholar] [CrossRef]
- Havelaar, A.H.; Kirk, M.D.; Torgerson, P.R.; Gibb, H.J.; Hald, T.; Lake, R.J.; Praet, N.; Bellinger, D.C.; de Silva, N.R.; Gargouri, N.; et al. World Health Organization Global Estimates and Regional Comparisons of the Burden of Foodborne Disease in 2010. PLoS Med. 2015, 12, e1001923. [Google Scholar] [CrossRef] [Green Version]
- Levine, M.M. Escherichia coli that Cause Diarrhea: Enterotoxigenic, Enteropathogenic, Enteroinvasive, Enterohemorrhagic, and Enteroadherent. J. Infect. Dis. 1987, 155, 377–389. [Google Scholar] [CrossRef] [PubMed]
- Brooks, J.T.; Sowers, E.G.; Wells, J.G.; Greene, K.D.; Griffin, P.M.; Hoekstra, R.M.; Strockbine, N.A. Non-O157 Shiga Toxin–Producing Escherichia coli Infections in the United States, 1983–2002. J. Infect. Dis. 2005, 192, 1422–1429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- CDC. FoodNet Fast. Available online: https://wwwn.cdc.gov/foodnetfast/ (accessed on 29 March 2021).
- USDA, F. Shiga Toxin-Producing Escherichia coli in Certain Raw Beef Products. Available online: https://www.federalregister.gov/documents/2012/05/31/2012-13283/shiga-toxin-producing-escherichia-coli-in-certain-raw-beef-products (accessed on 29 March 2021).
- NACMCF. Response to Questions Posed by the Food and Drug Administration Regarding Virulence Factors and Attributes that Define Foodborne Shiga Toxin-Producing Escherichia coli (STEC) as Severe Human Pathogens (†). J. Food Prot. 2019, 82, 724–767. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Biological Hazards. Scientific Opinion on VTEC-seropathotype and scientific criteria regarding pathogenicity assessment. EFSA J. 2013, 11, 3138. [Google Scholar] [CrossRef]
- The Interagency Food Safety Analytics Collaboration (IFSAC). Foodborne Illness Source Attribution Estimates for 2018 for Salmonella, Escherichia coli O157, Listeria monocytogenes, and Campylobacter Using Multi-Year Outbreak Surveillance Data, United States; The Interagency Food Safety Analytics Collaboration: Atlanta, GA, USA; Washington, DC, USA, 2020. [Google Scholar]
- Pires, S.M.; Majowicz, S.; Gill, A.; Devleesschauwer, B. Global and regional source attribution of Shiga toxin-producing Escherichia coli infections using analysis of outbreak surveillance data. Epidemiol. Infect. 2019, 147, e236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Painter, J.A.; Hoekstra, R.M.; Ayers, T.; Tauxe, R.V.; Braden, C.R.; Angulo, F.J.; Griffin, P.M. Attribution of foodborne illnesses, hospitalizations, and deaths to food commodities by using outbreak data, United States, 1998–2008. Emerg. Infect. Dis. 2013, 19, 407–415. [Google Scholar] [CrossRef] [PubMed]
- Ferens, W.A.; Hovde, C.J. Escherichia coli O157:H7: Animal Reservoir and Sources of Human Infection. Foodborne Pathog. Dis. 2010, 8, 465–487. [Google Scholar] [CrossRef] [Green Version]
- Carstens, C.K.; Salazar, J.K.; Darkoh, C. Multistate Outbreaks of Foodborne Illness in the United States Associated with Fresh Produce From 2010 to 2017. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rangel, J.M.; Sparling, P.H.; Crowe, C.; Griffin, P.M.; Swerdlow, D.L. Epidemiology of Escherichia coli O157:H7 outbreaks, United States, 1982–2002. Emerg. Infect. Dis. 2005, 11, 603–609. [Google Scholar] [CrossRef]
- Heiman, K.E.; Mody, R.K.; Johnson, S.D.; Griffin, P.M.; Gould, L.H. Escherichia coli O157 Outbreaks in the United States, 2003–2012. Emerg. Infect. Dis. 2015, 21, 1293–1301. [Google Scholar] [CrossRef] [PubMed]
- FDA. Available online: https://www.fda.gov/food/outbreaks-foodborne-illness/investigation-summary-factors-potentially-contributing-contamination-romaine-lettuce-implicated-fall (accessed on 29 March 2021).
- FDA. Available online: https://www.fda.gov/food/outbreaks-foodborne-illness/factors-potentially-contributing-contamination-romaine-lettuce-implicated-three-outbreaks-e-coli (accessed on 29 March 2021).
- Sharapov, U.M.; Wendel, A.M.; Davis, J.P.; Keene, W.E.; Farrar, J.; Sodha, S.; Hyytia-Trees, E.; Leeper, M.; Gerner-Smidt, P.; Griffin, P.M.; et al. Multistate Outbreak of Escherichia coli O157:H7 Infections Associated with Consumption of Fresh Spinach: United States, 2006. J. Food Prot. 2016, 79, 2024–2030. [Google Scholar] [CrossRef] [PubMed]
- FDA. Available online: https://www.fda.gov/Food/RecallsOutbreaksEmergencies/Outbreaks/ucm235477.htm (accessed on 29 March 2021).
- Ding, H.; Fu, T.-J.; Smith, M.A. Microbial Contamination in Sprouts: How Effective Is Seed Disinfection Treatment? J. Food Sci. 2013, 78, R495–R501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- CDC. Outbreak of E. coli Infections Linked to Clover Sprouts. Available online: https://www.cdc.gov/ecoli/2020/o103h2-02-20/index.html (accessed on 29 March 2021).
- Foley, C.; Harvey, E.; Bidol, S.A.; Henderson, T.; Njord, R.; DeSalvo, T.; Haupt, T.; Mba-Jonas, A.; Bailey, C.; Bopp, C. Outbreak of Escherichia coli O104:H4 infections associated with sprout consumption—Europe and North America, May–July 2011. MMWR Morb. Mortal. Wkly. Rep. 2013, 62, 1029–1031. [Google Scholar]
- Center for Food Safety and Applied Nutrition. Compliance with and Recommendations for Implementation of the Standards for the Growing, Harvesting, Packing, and Holding of Produce for Human Consumption for Sprout Operations: Guidance for Industry (Draft Guidance). Fed. Regist. 2017, 82, 7751–7753. [Google Scholar]
- European Union. Commission Regulation (EU) No 209/2013 of 11 March 2013 Amending Regulation (EC) No 2073/2005 as Regards Microbiological Criteria for Sprouts and the Sampling Rules for Poultry Carcases and Fresh Poultry Meat Text with EEA Relevance. Available online: https://eur-lex.europa.eu/eli/reg/2013/209/oj (accessed on 8 October 2021).
- Lauber, C.L.; Glatzer, L.; Sinsabaugh, R.L. Prevalence of Pathogenic Escherichia coli in Recreational Waters. J. Great Lakes Res. 2003, 29, 301–306. [Google Scholar] [CrossRef]
- Ishii, S.; Hansen, D.L.; Hicks, R.E.; Sadowsky, M.J. Beach Sand and Sediments are Temporal Sinks and Sources of Escherichia coli in Lake Superior. Environ. Sci. Technol. 2007, 41, 2203–2209. [Google Scholar] [CrossRef]
- Cho, S.; Hiott, L.M.; Barrett, J.B.; McMillan, E.A.; House, S.L.; Humayoun, S.B.; Adams, E.S.; Jackson, C.R.; Frye, J.G. Prevalence and characterization of Escherichia coli isolated from the Upper Oconee Watershed in Northeast Georgia. PLoS ONE 2018, 13, e0197005. [Google Scholar] [CrossRef] [Green Version]
- Hamelin, K.; Bruant, G.; El-Shaarawi, A.; Hill, S.; Edge, T.A.; Fairbrother, J.; Harel, J.; Maynard, C.; Masson, L.; Brousseau, R. Occurrence of Virulence and Antimicrobial Resistance Genes in Escherichia coli Isolates from Different Aquatic Ecosystems within the St. Clair River and Detroit River Areas. Appl. Environ. Microbiol. 2007, 73, 477–484. [Google Scholar] [CrossRef] [Green Version]
- Topalcengiz, Z.; Strawn, L.K.; Danyluk, M.D. Microbial quality of agricultural water in Central Florida. PLoS ONE 2017, 12, e0174889. [Google Scholar] [CrossRef] [PubMed]
- Sidhu, J.P.; Hodgers, L.; Ahmed, W.; Chong, M.N.; Toze, S. Prevalence of human pathogens and indicators in stormwater runoff in Brisbane, Australia. Water Res. 2012, 46, 6652–6660. [Google Scholar] [CrossRef] [PubMed]
- Ram, S.; Vajpayee, P.; Tripathi, U.; Singh, R.L.; Seth, P.K.; Shanker, R. Determination of antimicrobial resistance and virulence gene signatures in surface water isolates of Escherichia coli. J. Appl. Microbiol. 2008, 105, 1899–1908. [Google Scholar] [CrossRef] [PubMed]
- Cho, K.; Wolny, J.; Kase, J.A.; Unno, T.; Pachepsky, Y. Interactions of algae, aquatic vegetation, and E. coli in freshwater environments. Water Res. 2021. submitted. [Google Scholar]
- Lee, C.S.; Kim, M.; Lee, C.; Yu, Z.; Lee, J. The Microbiota of Recreational Freshwaters and the Implications for Environmental and Public Health. Front. Microbiol. 2016, 7, 1826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halac, S.R.; Bazán, R.V.; Larrosa, N.B.; Nadal, A.F.; Ruibal-Conti, A.L.; Rodríguez, M.I.; Ruiz, M.A.; López, A.G. First report on negative association between cyanobacteria and fecal indicator bacteria at San Roque reservoir (Argentina): Impact of environmental factors. J. Freshw. Ecol. 2019, 34, 273–291. [Google Scholar] [CrossRef] [Green Version]
- Canada.ca. Waterborne outbreak of gastroenteritis associated with a contaminated municipal water supply, Walkerton, Ontario, May–June 2000. Can. Commun. Dis. Rep. 2000, 26, 170–173. [Google Scholar]
- Probert, W.S.; Miller, G.M.; Ledin, K.E. Contaminated Stream Water as Source for Escherichia coli O157 Illness in Children. Emerg. Infect. Dis. 2017, 23, 1216–1218. [Google Scholar] [CrossRef] [Green Version]
- Gu, G.; Luo, Z.; Cevallos-Cevallos, J.M.; Adams, P.; Vellidis, G.; Wright, A.; van Bruggen, A.H. Factors affecting the occurrence of Escherichia coli O157 contamination in irrigation ponds on produce farms in the Suwannee River Watershed. Can. J. Microbiol. 2013, 59, 175–182. [Google Scholar] [CrossRef]
- Cooley, M.; Carychao, D.; Crawford-Miksza, L.; Jay, M.T.; Myers, C.; Rose, C.; Keys, C.; Farrar, J.; Mandrell, R.E. Incidence and Tracking of Escherichia coli O157:H7 in a Major Produce Production Region in California. PLoS ONE 2007, 2, e1159. [Google Scholar] [CrossRef]
- Maal-Bared, R.; Bartlett, K.H.; Bowie, W.R.; Hall, E.R. Phenotypic antibiotic resistance of Escherichia coli and E. coli O157 isolated from water, sediment and biofilms in an agricultural watershed in British Columbia. Sci. Total. Environ. 2013, 443, 315–323. [Google Scholar] [CrossRef]
- Majowicz, S.E.; Musto, J.; Scallan, E.; Angulo, F.J.; Kirk, M.; O’Brien, S.J.; Jones, T.F.; Fazil, A.; Hoekstra, R.M. The global burden of nontyphoidal Salmonella gastroenteritis. Clin. Infect. Dis. 2010, 50, 882–889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minor, T.; Lasher, A.; Klontz, K.; Brown, B.; Nardinelli, C.; Zorn, D. The Per Case and Total Annual Costs of Foodborne Illness in the United States. Risk Anal. 2015, 35, 1125–1139. [Google Scholar] [CrossRef] [PubMed]
- Callejón, R.M.; Rodríguez-Naranjo, M.I.; Ubeda, C.; Hornedo-Ortega, R.; Garcia-Parrilla, M.C.; Troncoso, A.M. Reported foodborne outbreaks due to fresh produce in the United States and European Union: Trends and causes. Foodborne Pathog. Dis. 2015, 12, 32–38. [Google Scholar] [CrossRef]
- CDC. Outbreak of Salmonella Newport Infections Linked to Onions. Available online: https://www.cdc.gov/Salmonella/newport-07-20/index.html (accessed on 12 June 2021).
- Centers for Disease Control and Prevention (CDC). Surveillance for Foodborne Disease Outbreaks, United States, 2017; Annual Report; CDC: Atlanta, GA, USA, 2019. [Google Scholar]
- FDA. Factors Potentially Contributing to the Contamination of Peaches Implicated in the Summer 2020 Outbreak of Salmonella Enteritidis; FDA: White Oak, MD, USA, 2021. [Google Scholar]
- Hanning, I.B.; Nutt, J.D.; Ricke, S.C. Salmonellosis outbreaks in the United States due to fresh produce: Sources and potential intervention measures. Foodborne Pathog. Dis. 2009, 6, 635–648. [Google Scholar] [CrossRef] [PubMed]
- Burris, K.P.; Simmons, O.D.; Webb, H.M.; Deese, L.M.; Moore, R.G.; Jaykus, L.A.; Zheng, J.; Reed, E.; Ferreira, C.M.; Brown, E.W.; et al. Colonization and Internalization of Salmonella enterica and Its Prevalence in Cucumber Plants. Front. Microbiol. 2020, 11, 1135. [Google Scholar] [CrossRef]
- Burris, K.P.; Simmons, O.D., 3rd; Webb, H.M.; Moore, R.G.; Jaykus, L.A.; Zheng, J.; Reed, E.; Ferreira, C.M.; Brown, E.; Bell, R.L. Salmonella enterica colonization and fitness in pre-harvest cantaloupe production. Food Microbiol. 2021, 93, 103612. [Google Scholar] [CrossRef]
- Zheng, J.; Allard, S.; Reynolds, S.; Millner, P.; Arce, G.; Blodgett, R.J.; Brown, E.W. Colonization and Internalization of Salmonella enterica in tomato plants. Appl. Environ. Microbiol. 2013, 79, 2494–2502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golberg, D.; Kroupitski, Y.; Belausov, E.; Pinto, R.; Sela, S. Salmonella Typhimurium internalization is variable in leafy vegetables and fresh herbs. Int. J. Food Microbiol. 2011, 145, 250–257. [Google Scholar] [CrossRef]
- Johnson, N.; Litt, P.K.; Kniel, K.E.; Bais, H. Evasion of Plant Innate Defense Response by Salmonella on Lettuce. Front. Microbiol. 2020, 11, 500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, C.H.; Shollenberger, L.M. Survivability and long-term preservation of bacteria in water and in phosphate-buffered saline. Lett. Appl. Microbiol. 2003, 37, 45–50. [Google Scholar] [CrossRef]
- Liu, H.; Whitehouse, C.A.; Li, B. Presence and Persistence of Salmonella in Water: The Impact on Microbial Quality of Water and Food Safety. Front. Public Health 2018, 6, 159. [Google Scholar] [CrossRef] [PubMed]
- Burton, G.A., Jr.; Gunnison, D.; Lanza, G.R. Survival of pathogenic bacteria in various freshwater sediments. Appl. Environ. Microbiol. 1987, 53, 633–638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Byappanahalli, M.N.; Sawdey, R.; Ishii, S.; Shively, D.A.; Ferguson, J.A.; Whitman, R.L.; Sadowsky, M.J. Seasonal stability of Cladophora-associated Salmonella in Lake Michigan watersheds. Water Res. 2009, 43, 806–814. [Google Scholar] [CrossRef] [PubMed]
- Gaertner, J.P.; Mendoza, J.A.; Forstner, M.R.; Hahn, D. Recovery of Salmonella from biofilms in a headwater spring ecosystem. J. Water Health 2011, 9, 458–466. [Google Scholar] [CrossRef] [PubMed]
- Hendricks, C.W. Increased recovery rate of Salmonellae from stream bottom sediments versus surface waters. Appl. Microbiol. 1971, 21, 379–380. [Google Scholar] [CrossRef] [PubMed]
- Van Donsel, D.J.; Geldreich, E.E. Relationships of Salmonellae to fecal coliforms in bottom sediments. Water Res. 1971, 5, 1079–1087. [Google Scholar] [CrossRef]
- Andie, D.A.; Weiser, H.H.; Maloney, G.W. Survival of Bacterial Enteric Pathogens in Farm Pond Water. J. AWWA 1967, 59, 503–508. [Google Scholar] [CrossRef]
- Hendricks, C.W.; Morrison, S.M. Multiplication and growth of selected enteric bacteria in clear mountain stream water. Water Res. 1967, 1, 567–576. [Google Scholar] [CrossRef]
- Cevallos-Cevallos, J.M.; Gu, G.; Richardson, S.M.; Hu, J.; van Bruggen, A.H. Survival of Salmonella enterica Typhimurium in water amended with manure. J. Food Prot. 2014, 77, 2035–2042. [Google Scholar] [CrossRef]
- Santo Domingo, J.W.; Harmon, S.; Bennett, J. Survival of Salmonella species in river water. Curr. Microbiol. 2000, 40, 409–417. [Google Scholar] [CrossRef]
- Topalcengiz, Z.; McEgan, R.; Danyluk, M.D. Fate of Salmonella in Central Florida Surface Waters and Evaluation of EPA Worst Case Water as a Standard Medium. J. Food Prot. 2019, 82, 916–925. [Google Scholar] [CrossRef] [PubMed]
- Roszak, D.B.; Grimes, D.J.; Colwell, R.R. Viable but nonrecoverable stage of Salmonella enteritidis in aquatic systems. Can. J. Microbiol. 1984, 30, 334–338. [Google Scholar] [CrossRef]
- Aguirre-Sanchez, J.R.; Ibarra-Rodriguez, J.R.; Vega-Lopez, I.F.; Martínez-Urtaza, J.; Chaidez-Quiroz, C. Genomic signatures of adaptation to natural settings in non-typhoidal Salmonella enterica Serovars Saintpaul, Thompson and Weltevreden. Infect. Genet. Evol. 2021, 90, 104771. [Google Scholar] [CrossRef]
- Deaven, A.M.; Ferreira, C.M.; Reed, E.A.; Chen See, J.R.; Lee, N.A.; Almaraz, E.; Rios, P.C.; Marogi, J.G.; Lamendella, R.; Zheng, J.; et al. Salmonella Genomics and Population Analyses Reveal High Inter- and Intraserovar Diversity in Freshwater. Appl. Environ. Microbiol. 2021, 87. [Google Scholar] [CrossRef] [PubMed]
- Draper, A.D.; Doores, S.; Gourama, H.; LaBorde, L.F. Microbial Survey of Pennsylvania Surface Water Used for Irrigating Produce Crops. J. Food Prot. 2016, 79, 902–912. [Google Scholar] [CrossRef] [PubMed]
- Gu, G.; Strawn, L.K.; Zheng, J.; Reed, E.A.; Rideout, S.L. Diversity and Dynamics of Salmonella enterica in Water Sources, Poultry Litters, and Field Soils Amended with Poultry Litter in a Major Agricultural Area of Virginia. Front. Microbiol. 2019, 10, 2868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haley, B.J.; Cole, D.J.; Lipp, E.K. Distribution, diversity, and seasonality of waterborne Salmonellae in a rural watershed. Appl. Environ. Microbiol. 2009, 75, 1248–1255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, D.; Tertuliano, M.; Vellidis, G.; Harris, C.; Grossman, M.K.; Rajeev, S.; Levy, K. Evaluation of Grower-Friendly, Science-Based Sampling Approaches for the Detection of Salmonella in Ponds Used for Irrigation of Fresh Produce. Foodborne Pathog. Dis. 2018, 15, 627–636. [Google Scholar] [CrossRef]
- Luo, Z.; Gu, G.; Ginn, A.; Giurcanu, M.C.; Adams, P.; Vellidis, G.; van Bruggen, A.H.; Danyluk, M.D.; Wright, A.C. Distribution and Characterization of Salmonella enterica Isolates from Irrigation Ponds in the Southeastern United States. Appl. Environ. Microbiol. 2015, 81, 4376–4387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McEgan, R.; Mootian, G.; Goodridge, L.D.; Schaffner, D.W.; Danyluk, M.D. Predicting Salmonella Populations from Biological, Chemical, and Physical Indicators in Florida Surface Waters. Appl. Environ. Microbiol. 2013, 79, 4094–4105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patchanee, P.; Molla, B.; White, N.; Line, D.E.; Gebreyes, W.A. Tracking Salmonella contamination in various watersheds and phenotypic and genotypic diversity. Foodborne Pathog. Dis. 2010, 7, 1113–1120. [Google Scholar] [CrossRef] [PubMed]
- Rajabi, M.; Jones, M.; Hubbard, M.; Rodrick, G.; Wright, A.C. Distribution and Genetic Diversity of Salmonella enterica in the Upper Suwannee River. Int. J. Microbiol. 2011, 2011, 461321. [Google Scholar] [CrossRef] [Green Version]
- Strawn, L.K.; Gröhn, Y.T.; Warchocki, S.; Worobo, R.W.; Bihn, E.A.; Wiedmann, M. Risk factors associated with Salmonella and Listeria monocytogenes contamination of produce fields. Appl. Environ. Microbiol. 2013, 79, 7618–7627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Truitt, L.N.; Vazquez, K.M.; Pfuntner, R.C.; Rideout, S.L.; Havelaar, A.H.; Strawn, L.K. Microbial Quality of Agricultural Water Used in Produce Preharvest Production on the Eastern Shore of Virginia. J. Food Prot. 2018, 81, 1661–1672. [Google Scholar] [CrossRef] [PubMed]
- Benjamin, L.; Atwill, E.R.; Jay-Russell, M.; Cooley, M.; Carychao, D.; Gorski, L.; Mandrell, R.E. Occurrence of generic Escherichia coli, E. coli O157 and Salmonella spp. in water and sediment from leafy green produce farms and streams on the Central California coast. Int. J. Food Microbiol. 2013, 165, 65–76. [Google Scholar] [CrossRef]
- Gorski, L.; Parker, C.T.; Liang, A.; Cooley, M.B.; Jay-Russell, M.T.; Gordus, A.G.; Atwill, E.R.; Mandrell, R.E. Prevalence, distribution, and diversity of Salmonella enterica in a major produce region of California. Appl. Environ. Microbiol. 2011, 77, 2734–2748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Partyka, M.L.; Bond, R.F.; Chase, J.A.; Atwill, E.R. Spatial and temporal variability of bacterial indicators and pathogens in six California reservoirs during extreme drought. Water Res. 2018, 129, 436–446. [Google Scholar] [CrossRef] [PubMed]
- Walters, S.P.; Thebo, A.L.; Boehm, A.B. Impact of urbanization and agriculture on the occurrence of bacterial pathogens and stx genes in coastal waterbodies of central California. Water Res. 2011, 45, 1752–1762. [Google Scholar] [CrossRef]
- Arvanitidou, M.; Kanellou, K.; Vagiona, D.G. Diversity of Salmonella spp. and fungi in northern Greek rivers and their correlation to fecal pollution indicators. Environ. Res. 2005, 99, 278–284. [Google Scholar] [CrossRef]
- Díaz-Torres, O.; Lugo-Melchor, O.Y.; de Anda, J.; Gradilla-Hernández, M.S.; Amézquita-López, B.A.; Meza-Rodríguez, D. Prevalence, Distribution, and Diversity of Salmonella Strains Isolated from a Subtropical Lake. Front. Microbiol. 2020, 11, 521146. [Google Scholar] [CrossRef] [PubMed]
- Flockhart, L.; Pintar, K.; Cook, A.; McEwen, S.; Friendship, R.; Kelton, D.; Pollari, F. Distribution of Salmonella in Humans, Production Animal Operations and a Watershed in a FoodNet Canada Sentinel Site. Zoonoses Public Health 2017, 64, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, B.G.-V.; Espinar, A.C.; Carmona, M.J.B. A comparative study of strains of Salmonella isolated from irrigation waters, vegetables and human infections. Epidemiol. Infect. 1987, 98, 271–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, Y.N.; Tsai, H.C.; Hsu, B.M.; Chiou, C.S. The association of Salmonella enterica from aquatic environmental and clinical samples in Taiwan. Sci. Total Environ. 2018, 624, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.H.; Hsu, B.M.; Chou, M.Y.; Tsai, H.L.; Kao, P.M.; Wang, H.J.; Hsiao, H.Y.; Su, M.J.; Huang, Y.L. Application of molecular biological techniques to analyze Salmonella seasonal distribution in stream water. FEMS Microbiol. Lett. 2014, 352, 87–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polo, F.; Figueras, M.J.; Inza, I.; Sala, J.; Fleisher, J.M.; Guarro, J. Relationship between presence of Salmonella and indicators of faecal pollution in aquatic habitats. FEMS Microbiol. Lett. 1998, 160, 253–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, J.L.; Slawson, R.M.; Taylor, W.D. Salmonella serotype diversity and seasonality in urban and rural streams. J. Appl. Microbiol. 2013, 114, 907–922. [Google Scholar] [CrossRef] [PubMed]
- Polat, H.; Topalcengiz, Z.; Danyluk, M.D. Prediction of Salmonella presence and absence in agricultural surface waters by artificial intelligence approaches. J. Food Saf. 2020, 40, e12733. [Google Scholar] [CrossRef]
- Li, B.; Vellidis, G.; Liu, H.; Jay-Russell, M.; Zhao, S.; Hu, Z.; Wright, A.; Elkins, C.A. Diversity and antimicrobial resistance of Salmonella enterica isolates from surface water in Southeastern United States. Appl. Environ. Microbiol. 2014, 80, 6355–6365. [Google Scholar] [CrossRef] [Green Version]
- Andrews, W.H.; Jacobson, A.; Hammack, T.S. Salmonella. In Bacteriological Analytical Manual; FDA: White Oak, MD, USA, 2021. [Google Scholar]
- ISO. ISO 19250:2010 Water Quality—Detection of Salmonella spp.; ISO: Geneva, Switzerland, 2010. [Google Scholar]
- Gorski, L. Selective enrichment media bias the types of Salmonella enterica strains isolated from mixed strain cultures and complex enrichment broths. PLoS ONE 2012, 7, e34722. [Google Scholar] [CrossRef]
- Siceloff, A.T.; Ohta, N.; Norman, K.N.; Loneragan, G.H.; Norby, B.; Scott, H.M.; Shariat, N.W. Antimicrobial Resistance Hidden within Multiserovar Salmonella Populations. Antimicrob. Agents Chemother. 2021, 65. [Google Scholar] [CrossRef]
- Bell, R.L.; Jarvis, K.G.; Ottesen, A.R.; McFarland, M.A.; Brown, E.W. Recent and emerging innovations in Salmonella detection: A food and environmental perspective. Microb. Biotechnol. 2016, 9, 279–292. [Google Scholar] [CrossRef] [PubMed]
- Kaakoush, N.O.; Castano-Rodriguez, N.; Mitchell, H.M.; Man, S.M. Global Epidemiology of Campylobacter Infection. Clin. Microbiol. Rev. 2015, 28, 687–720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramees, T.P.; Dhama, K.; Karthik, K.; Rathore, R.S.; Kumar, A.; Saminathan, M.; Tiwari, R.; Malik, Y.S.; Singh, R.K. Arcobacter: An emerging food-borne zoonotic pathogen, its public health concerns and advances in diagnosis and control—A comprehensive review. Vet. Q. 2017, 37, 136–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Igwaran, A.; Okoh, A.I. Human campylobacteriosis: A public health concern of global importance. Heliyon 2019, 5, e02814. [Google Scholar] [CrossRef] [PubMed]
- Black, R.E.; Levine, M.M.; Clements, M.L.; Hughes, T.P.; Blaser, M.J. Experimental Campylobacter jejuni infection in humans. J. Infect. Dis. 1988, 157, 472–479. [Google Scholar] [CrossRef]
- Fitzgerald, C. Campylobacter. Clin. Lab. Med. 2015, 35, 289–298. [Google Scholar] [CrossRef]
- Kapperud, G.; Espeland, G.; Wahl, E.; Walde, A.; Herikstad, H.; Gustavsen, S.; Tveit, I.; Natas, O.; Bevanger, L.; Digranes, A. Factors associated with increased and decreased risk of Campylobacter infection: A prospective case-control study in Norway. Am. J. Epidemiol. 2003, 158, 234–242. [Google Scholar] [CrossRef] [PubMed]
- Moreira, N.A.; Bondelind, M. Safe drinking water and waterborne outbreaks. J. Water Health 2017, 15, 83–96. [Google Scholar] [CrossRef] [PubMed]
- Taylor, D.N.; Porter, B.W.; Williams, C.A.; Miller, H.G.; Bopp, C.A.; Blake, P.A. Campylobacter enteritis: A large outbreak traced to commercial raw milk. West. J. Med. 1982, 137, 365–369. [Google Scholar]
- Vogt, R.L.; Sours, H.E.; Barrett, T.; Feldman, R.A.; Dickinson, R.J.; Witherell, L. Campylobacter enteritis associated with contaminated water. Ann. Intern. Med. 1982, 96, 292–296. [Google Scholar] [CrossRef]
- Palmer, S.R.; Gully, P.R.; White, J.M.; Pearson, A.D.; Suckling, W.G.; Jones, D.M.; Rawes, J.C.; Penner, J.L. Water-borne outbreak of Campylobacter gastroenteritis. Lancet 1983, 1, 287–290. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention. Surveillance for waterborne disease outbreaks associated with drinking water and other nonrecreational water—United States, 2009–2010. MMWR Morb. Mortal. Wkly. Rep. 2013, 62, 714–720. [Google Scholar]
- Schuster, C.J.; Ellis, A.G.; Robertson, W.J.; Charron, D.F.; Aramini, J.J.; Marshall, B.J.; Medeiros, D.T. Infectious disease outbreaks related to drinking water in Canada, 1974–2001. Can. J. Public Health 2005, 96, 254–258. [Google Scholar] [CrossRef]
- Merritt, A.; Miles, R.; Bates, J. An outbreak of Campylobacter enteritis on an island resort, north Queensland. Commun. Dis. Intell. 1999, 23, 215–219, discussion 220. [Google Scholar] [PubMed]
- Hanninen, M.L.; Haajanen, H.; Pummi, T.; Wermundsen, K.; Katila, M.L.; Sarkkinen, H.; Miettinen, I.; Rautelin, H. Detection and typing of Campylobacter jejuni and Campylobacter coli and analysis of indicator organisms in three waterborne outbreaks in Finland. Appl. Environ. Microbiol. 2003, 69, 1391–1396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pitkanen, T.; Miettinen, I.T.; Nakari, U.M.; Takkinen, J.; Nieminen, K.; Siitonen, A.; Kuusi, M.; Holopainen, A.; Hanninen, M.L. Faecal contamination of a municipal drinking water distribution system in association with Campylobacter jejuni infections. J. Water Health 2008, 6, 365–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pitkanen, T. Review of Campylobacter spp. in drinking and environmental waters. J. Microbiol. Methods 2013, 95, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Jalava, K.; Rintala, H.; Ollgren, J.; Maunula, L.; Gomez-Alvarez, V.; Revez, J.; Palander, M.; Antikainen, J.; Kauppinen, A.; Rasanen, P.; et al. Novel microbiological and spatial statistical methods to improve strength of epidemiological evidence in a community-wide waterborne outbreak. PLoS ONE 2014, 9, e104713. [Google Scholar] [CrossRef] [Green Version]
- Guzman-Herrador, B.; Carlander, A.; Ethelberg, S.; de Blasio, B.F.; Kuusi, M.; Lund, V.; Lofdahl, M.; MacDonald, E.; Nichols, G.; Schonning, C.; et al. Waterborne outbreaks in the Nordic countries, 1998 to 2012. Euro Surveill. 2015, 20. [Google Scholar] [CrossRef] [Green Version]
- Gibney, K.B.; O’Toole, J.; Sinclair, M.; Leder, K. Burden of Disease Attributed to Waterborne Transmission of Selected Enteric Pathogens, Australia, 2010. Am. J. Trop. Med. Hyg. 2017, 96, 1400–1403. [Google Scholar] [CrossRef]
- Shrestha, R.D.; Midwinter, A.C.; Marshall, J.C.; Collins-Emerson, J.M.; Pleydell, E.J.; French, N.P. Campylobacter jejuni Strains Associated with Wild Birds and Those Causing Human Disease in Six High-Use Recreational Waterways in New Zealand. Appl. Environ. Microbiol. 2019, 85. [Google Scholar] [CrossRef] [PubMed]
- Kubota, K.; Kasuga, F.; Iwasaki, E.; Inagaki, S.; Sakurai, Y.; Komatsu, M.; Toyofuku, H.; Angulo, F.J.; Scallan, E.; Morikawa, K. Estimating the burden of acute gastroenteritis and foodborne illness caused by Campylobacter, Salmonella, and Vibrio parahaemolyticus by using population-based telephone survey data, Miyagi Prefecture, Japan, 2005 to 2006. J. Food Prot. 2011, 74, 1592–1598. [Google Scholar] [CrossRef] [PubMed]
- Abe, T.; Haga, S.; Yokoyama, K.; Watanabe, N. An outbreak of Campylobacter jejuni subsp. jejuni infection via tap water. JPN J. Infect. Dis. 2008, 61, 327. [Google Scholar] [PubMed]
- VanDerslice, J.; Briscoe, J. Environmental interventions in developing countries: Interactions and their implications. Am. J. Epidemiol. 1995, 141, 135–144. [Google Scholar] [CrossRef] [PubMed]
- Ashbolt, N.J. Microbial contamination of drinking water and disease outcomes in developing regions. Toxicology 2004, 198, 229–238. [Google Scholar] [CrossRef]
- Gupta, S.S.; Mohammed, M.H.; Ghosh, T.S.; Kanungo, S.; Nair, G.B.; Mande, S.S. Metagenome of the gut of a malnourished child. Gut Pathog. 2011, 3, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saidi, S.M.; Iijima, Y.; Sang, W.K.; Mwangudza, A.K.; Oundo, J.O.; Taga, K.; Aihara, M.; Nagayama, K.; Yamamoto, H.; Waiyaki, P.G.; et al. Epidemiological study on infectious diarrheal diseases in children in a coastal rural area of Kenya. Microbiol. Immunol. 1997, 41, 773–778. [Google Scholar] [CrossRef]
- Lengerh, A.; Moges, F.; Unakal, C.; Anagaw, B. Prevalence, associated risk factors and antimicrobial susceptibility pattern of Campylobacter species among under five diarrheic children at Gondar University Hospital, Northwest Ethiopia. BMC Pediatr. 2013, 13, 82. [Google Scholar] [CrossRef] [Green Version]
- Chukwu, M.O.; Abia, A.L.K.; Ubomba-Jaswa, E.; Obi, L.; Dewar, J.B. Characterization and Phylogenetic Analysis of Campylobacter Species Isolated from Paediatric Stool and Water Samples in the Northwest Province, South Africa. Int. J. Environ. Res. Public Health 2019, 16, 2205. [Google Scholar] [CrossRef] [Green Version]
- Collado, L.; Figueras, M.J. Taxonomy, epidemiology, and clinical relevance of the genus Arcobacter. Clin. Microbiol. Rev. 2011, 24, 174–192. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Cataluña, A.; Salas-Massó, N.; Diéguez, A.L.; Balboa, S.; Lema, A.; Romalde, J.L.; Figueras, M.J. Revisiting the Taxonomy of the Genus Arcobacter: Getting Order from the Chaos. Front. Microbiol. 2018, 9, 2077. [Google Scholar] [CrossRef]
- Ho, H.T.; Lipman, L.J.; Gaastra, W. Arcobacter, what is known and unknown about a potential foodborne zoonotic agent! Vet. Microbiol. 2006, 115, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Fong, T.T.; Mansfield, L.S.; Wilson, D.L.; Schwab, D.J.; Molloy, S.L.; Rose, J.B. Massive microbiological groundwater contamination associated with a waterborne outbreak in Lake Erie, South Bass Island, Ohio. Environ. Health Perspect. 2007, 115, 856–864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, T.T.; Lee, J. Global Distribution and Prevalence of Arcobacter in Food and Water. Zoonoses Public Health 2015, 62, 579–589. [Google Scholar] [CrossRef]
- Talay, F.; Molva, C.; Atabay, H.I. Isolation and identification of Arcobacter species from environmental and drinking water samples. Folia Microbiol. 2016, 61, 479–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Reilly, C.E.; Bowen, A.B.; Perez, N.E.; Sarisky, J.P.; Shepherd, C.A.; Miller, M.D.; Hubbard, B.C.; Herring, M.; Buchanan, S.D.; Fitzgerald, C.C.; et al. A waterborne outbreak of gastroenteritis with multiple etiologies among resort island visitors and residents: Ohio, 2004. Clin. Infect. Dis. 2007, 44, 506–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rice, E.W.; Rodgers, M.R.; Wesley, I.V.; Johnson, C.H.; Tanner, S.A. Isolation of Arcobacter butzleri from ground water. Lett. Appl. Microbiol. 1999, 28, 31–35. [Google Scholar] [CrossRef]
- Jacob, J.; Woodward, D.; Feuerpfeil, I.; Johnson, W.M. Isolation of Arcobacter butzleri in raw water and drinking water treatment plants in Germany. Zent. Hyg. Umw. 1998, 201, 189–198. [Google Scholar]
- Pedati, C.; Koirala, S.; Safranek, T.; Buss, B.F.; Carlson, A.V. Campylobacteriosis Outbreak Associated with Contaminated Municipal Water Supply—Nebraska, 2017. MMWR Morb. Mortal. Wkly. Rep. 2019, 68, 169–173. [Google Scholar] [CrossRef] [Green Version]
- Murray, R.T.; Cruz-Cano, R.; Nasko, D.; Blythe, D.; Ryan, P.; Boyle, M.M.; Wilson, S.M.; Sapkota, A.R. Association between private drinking water wells and the incidence of Campylobacteriosis in Maryland: An ecological analysis using Foodborne Diseases Active Surveillance Network (FoodNet) data (2007–2016). Environ. Res. 2020, 188, 109773. [Google Scholar] [CrossRef]
- Whiley, H.; van den Akker, B.; Giglio, S.; Bentham, R. The role of environmental reservoirs in human campylobacteriosis. Int. J. Environ. Res. Public Health 2013, 10, 5886–5907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphy, H.M.; Thomas, M.K.; Schmidt, P.J.; Medeiros, D.T.; McFadyen, S.; Pintar, K.D. Estimating the burden of acute gastrointestinal illness due to Giardia, Cryptosporidium, Campylobacter, E. coli O157 and norovirus associated with private wells and small water systems in Canada. Epidemiol. Infect. 2016, 144, 1355–1370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyllestad, S.; Iversen, A.; MacDonald, E.; Amato, E.; Borge, B.A.S.; Boe, A.; Sandvin, A.; Brandal, L.T.; Lyngstad, T.M.; Naseer, U.; et al. Large waterborne Campylobacter outbreak: Use of multiple approaches to investigate contamination of the drinking water supply system, Norway, June 2019. Eurosurveilliance 2020, 25. [Google Scholar] [CrossRef]
- Mulder, A.C.; Franz, E.; de Rijk, S.; Versluis, M.A.J.; Coipan, C.; Buij, R.; Muskens, G.; Koene, M.; Pijnacker, R.; Duim, B.; et al. Tracing the animal sources of surface water contamination with Campylobacter jejuni and Campylobacter coli. Water Res. 2020, 187, 116421. [Google Scholar] [CrossRef]
- Phiri, B.J.; French, N.P.; Biggs, P.J.; Stevenson, M.A.; Reynolds, A.D.; Garcia, R.J.; Hayman, D.T.S. Microbial contamination in drinking water at public outdoor recreation facilities in New Zealand. J. Appl. Microbiol. 2021, 130, 302–312. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.; Agidi, S.; Marion, J.W.; Lee, J. Arcobacter in Lake Erie beach waters: An emerging gastrointestinal pathogen linked with human-associated fecal contamination. Appl. Environ. Microbiol. 2012, 78, 5511–5519. [Google Scholar] [CrossRef]
- Webb, A.L.; Taboada, E.N.; Selinger, L.B.; Boras, V.F.; Inglis, G.D. Efficacy of wastewater treatment on Arcobacter butzleri density and strain diversity. Water Res. 2016, 105, 291–296. [Google Scholar] [CrossRef]
- Niedermeyer, J.A.; Miller, W.G.; Yee, E.; Harris, A.; Emanuel, R.E.; Jass, T.; Nelson, N.; Kathariou, S. Search for Campylobacter spp. Reveals High Prevalence and Pronounced Genetic Diversity of Arcobacter butzleri in Floodwater Samples Associated with Hurricane Florence in North Carolina, USA. Appl. Environ. Microbiol. 2020, 86. [Google Scholar] [CrossRef]
- Diergaardt, S.M.; Venter, S.N.; Spreeth, A.; Theron, J.; Brozel, V.S. The occurrence of campylobacters in water sources in South Africa. Water Res. 2004, 38, 2589–2595. [Google Scholar] [CrossRef]
- St-Pierre, K.; Levesque, S.; Frost, E.; Carrier, N.; Arbeit, R.D.; Michaud, S. Thermotolerant coliforms are not a good surrogate for Campylobacter spp. in environmental water. Appl. Environ. Microbiol. 2009, 75, 6736–6744. [Google Scholar] [CrossRef] [Green Version]
- Ghaju Shrestha, R.; Tanaka, Y.; Sherchand, J.B.; Haramoto, E. Identification of 16S rRNA and Virulence-Associated Genes of Arcobacter in Water Samples in the Kathmandu Valley, Nepal. Pathogens 2019, 8, 110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sorensen, J.P.; Lapworth, D.J.; Read, D.S.; Nkhuwa, D.C.; Bell, R.A.; Chibesa, M.; Chirwa, M.; Kabika, J.; Liemisa, M.; Pedley, S. Tracing enteric pathogen contamination in sub-Saharan African groundwater. Sci. Total Environ. 2015, 538, 888–895. [Google Scholar] [CrossRef] [Green Version]
- Cousins, M.; Sargeant, J.M.; Fisman, D.N.; Greer, A.L. Identifying the environmental drivers of Campylobacter infection risk in southern Ontario, Canada using a One Health approachs. Zoonoses Public Health 2020, 67, 516–524. [Google Scholar] [CrossRef]
- Van Driessche, E.; Houf, K. Survival capacity in water of Arcobacter species under different temperature conditions. J. Appl. Microbiol. 2008, 105, 443–451. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, S.; Araujo, R. Occurrence of thermotolerant Campylobacter species in surface waters of a Mediterranean area and in its prevailing pollution sources. J. Appl. Microbiol. 2010, 109, 1027–1034. [Google Scholar] [CrossRef] [PubMed]
- Vereen, E., Jr.; Lowrance, R.R.; Jenkins, M.B.; Adams, P.; Rajeev, S.; Lipp, E.K. Landscape and seasonal factors influence Salmonella and Campylobacter prevalence in a rural mixed use watershed. Water Res. 2013, 47, 6075–6085. [Google Scholar] [CrossRef] [Green Version]
- Mughini-Gras, L.; Penny, C.; Ragimbeau, C.; Schets, F.M.; Blaak, H.; Duim, B.; Wagenaar, J.A.; de Boer, A.; Cauchie, H.M.; Mossong, J.; et al. Quantifying potential sources of surface water contamination with Campylobacter jejuni and Campylobacter coli. Water Res. 2016, 101, 36–45. [Google Scholar] [CrossRef]
- David, J.M.; Pollari, F.; Pintar, K.D.M.; Nesbitt, A.; Butler, A.J.; Ravel, A. Do contamination of and exposure to chicken meat and water drive the temporal dynamics of Campylobacter cases? Epidemiol. Infect. 2017, 145, 3191–3203. [Google Scholar] [CrossRef] [Green Version]
- Cools, I.; Uyttendaele, M.; Caro, C.; D’Haese, E.; Nelis, H.J.; Debevere, J. Survival of Campylobacter jejuni strains of different origin in drinking water. J. Appl. Microbiol. 2003, 94, 886–892. [Google Scholar] [CrossRef]
- Trigui, H.; Thibodeau, A.; Fravalo, P.; Letellier, A.; Faucher, S.P. Survival in water of Campylobacter jejuni strains isolated from the slaughterhouse. Springerplus 2015, 4, 799. [Google Scholar] [CrossRef]
- Patrone, V.; Campana, R.; Vallorani, L.; Dominici, S.; Federici, S.; Casadei, L.; Gioacchini, A.M.; Stocchi, V.; Baffone, W. CadF expression in Campylobacter jejuni strains incubated under low-temperature water microcosm conditions which induce the viable but non-culturable (VBNC) state. Antonie Van Leeuwenhoek 2013, 103, 979–988. [Google Scholar] [CrossRef]
- Humphrey, T.J.; Muscat, I. Incubation temperature and the isolation of Campylobacter jejuni from food, milk or water. Lett. Appl. Microbiol. 1989, 9, 137–139. [Google Scholar] [CrossRef]
- Khan, I.U.; Hill, S.; Nowak, E.; Edge, T.A. Effect of incubation temperature on the detection of thermophilic Campylobacter species from freshwater beaches, nearby wastewater effluents, and bird fecal droppings. Appl. Environ. Microbiol. 2013, 79, 7639–7645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mason, M.J.; Humphrey, T.J.; Martin, K.W. Isolation of sublethally injured campylobacters from poultry and water sources. Br. J. Biomed. Sci. 1998, 56, 2–5. [Google Scholar]
- Humphrey, T.J.; Martin, K.W.; Slader, J.; Durham, K. Campylobacter spp. in the kitchen: Spread and persistence. Symp. Ser. Soc. Appl. Microbiol. 2001, 115S–120S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moran, L.; Kelly, C.; Cormican, M.; McGettrick, S.; Madden, R.H. Restoring the selectivity of Bolton broth during enrichment for Campylobacter spp. from raw chicken. Lett. Appl. Microbiol. 2011, 52, 614–618. [Google Scholar] [CrossRef]
- Carrillo, C.D.; Kenwell, R.; Iugovaz, I.; Oyarzabal, O.A. Recovery of Campylobacter spp. from Food and Environmental Sources. Methods Mol. Biol. 2017, 1512, 9–18. [Google Scholar] [CrossRef]
- Hiett, K.L. Campylobacter jejuni Isolation/Enumeration from Environmental Samples. Methods Mol. Biol. 2017, 1512, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Chon, J.W.; Kim, D.H.; Bae, D.; Song, K.Y.; Kim, H.; Sung, K.; Seo, K.H. Comparison of Direct Syringe Filtration and Membrane Filtration for the Selective Isolation of Campylobacter jejuni from Ready-to-Eat Sprouts. Foodborne Pathog. Dis. 2019, 16, 371–375. [Google Scholar] [CrossRef]
- Kim, J.; Oh, E.; Banting, G.S.; Braithwaite, S.; Chui, L.; Ashbolt, N.J.; Neumann, N.F.; Jeon, B. An Improved Culture Method for Selective Isolation of Campylobacter jejuni from Wastewater. Front. Microbiol. 2016, 7, 1345. [Google Scholar] [CrossRef] [PubMed]
- Reddington, K.; Eccles, D.; O’Grady, J.; Drown, D.M.; Hansen, L.H.; Nielsen, T.K.; Ducluzeau, A.L.; Leggett, R.M.; Heavens, D.; Peel, N.; et al. Metagenomic analysis of planktonic riverine microbial consortia using nanopore sequencing reveals insight into river microbe taxonomy and function. Gigascience 2020, 9. [Google Scholar] [CrossRef]
- Lv, R.; Wang, K.; Feng, J.; Heeney, D.D.; Liu, D.; Lu, X. Detection and Quantification of Viable but Non-culturable Campylobacter jejuni. Front. Microbiol. 2019, 10, 2920. [Google Scholar] [CrossRef]
- Levican, A.; Collado, L.; Figueras, M.J. The Use of Two Culturing Methods in Parallel Reveals a High Prevalence and Diversity of Arcobacter spp. in a Wastewater Treatment Plant. Biomed. Res. Int. 2016, 2016, 8132058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramees, T.P.; Rathore, R.S.; Bagalkot, P.S.; Sailo, B.; Mohan, H.V.; Kumar, A.; Dhama, K.; Singh, R.K. Genotyping and genetic diversity of Arcobacter butzleri and Arcobacter cryaerophilus isolated from different sources by using ERIC-PCR from India. Vet. Q. 2014, 34, 211–217. [Google Scholar] [CrossRef] [PubMed]
- Levican, A.; Figueras, M.J. Performance of five molecular methods for monitoring Arcobacter spp. BMC Microbiol. 2013, 13, 220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, P.T.; JuArez, O.; Restaino, L. A New Method for Detection of Arcobacter butzleri, Arcobacter cryaerophilus, and Arcobacter skirrowii Using a Novel Chromogenic Agar. J. Food Prot 2021, 84, 160–168. [Google Scholar] [CrossRef]
- Shrestha, R.G.; Tanaka, Y.; Malla, B.; Tandukar, S.; Bhandari, D.; Inoue, D.; Sei, K.; Sherchand, J.B.; Haramoto, E. Development of a Quantitative PCR Assay for Arcobacter spp. and its Application to Environmental Water Samples. Microbes Environ. 2018, 33, 309–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, I.U.H.; Becker, A.; Cloutier, M.; Plotz, M.; Lapen, D.R.; Wilkes, G.; Topp, E.; Abdulmawjood, A. Loop-mediated isothermal amplification: Development, validation and application of simple and rapid assays for quantitative detection of species of Arcobacteraceae family- and species-specific Aliarcobacter faecis and Aliarcobacter lanthieri. J. Appl. Microbiol. 2020. [Google Scholar] [CrossRef]
- Laishram, M.; Rathlavath, S.; Lekshmi, M.; Kumar, S.; Nayak, B.B. Isolation and characterization of Arcobacter spp. from fresh seafood and the aquatic environment. Int. J. Food Microbiol. 2016, 232, 87–89. [Google Scholar] [CrossRef] [PubMed]
- Acharya, K.; Blackburn, A.; Mohammed, J.; Haile, A.T.; Hiruy, A.M.; Werner, D. Metagenomic water quality monitoring with a portable laboratory. Water Res. 2020, 184, 116112. [Google Scholar] [CrossRef]
- Bloomfield, S.; Wilkinson, D.; Rogers, L.; Biggs, P.; French, N.; Mohan, V.; Savoian, M.; Venter, P.; Midwinter, A. Campylobacter novaezeelandiae sp. nov., isolated from birds and water in New Zealand. Int. J. Syst. Evol. Microbiol. 2020, 70, 3775–3784. [Google Scholar] [CrossRef] [PubMed]
- Farber, J.M.; Peterkin, P.I. Listeria monocytogenes, a food-borne pathogen. Microbiol. Rev. 1991, 55, 476–511. [Google Scholar] [CrossRef] [PubMed]
- Barbuddhe, S.B.; Chakraborty, T. Listeria as an enteroinvasive gastrointestinal pathogen. Curr. Top. Microbiol. Immunol. 2009, 337, 173–195. [Google Scholar] [CrossRef] [PubMed]
- Pohl, A.M.; Pouillot, R.; Bazaco, M.C.; Wolpert, B.J.; Healy, J.M.; Bruce, B.B.; Laughlin, M.E.; Hunter, J.C.; Dunn, J.R.; Hurd, S.; et al. Differences Among Incidence Rates of Invasive Listeriosis in the U.S. FoodNet Population by Age, Sex, Race/Ethnicity, and Pregnancy Status, 2008–2016. Foodborne Pathog. Dis. 2019, 16, 290–297. [Google Scholar] [CrossRef]
- Kwon, H.J.; Chen, Z.; Evans, P.; Meng, J.; Chen, Y. Characterization of Mobile Genetic Elements Using Long-Read Sequencing for Tracking Listeria monocytogenes from Food Processing Environments. Pathogens 2020, 9, 822. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Gonzalez-Escalona, N.; Hammack, T.S.; Allard, M.W.; Strain, E.A.; Brown, E.W. Core genome multilocus sequence typing for identification of globally distributed clonal groups and differentiation of outbreak strains of Listeria monocytogenes. Appl. Environ. Microbiol. 2016, 82, 6258–6272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qing, J.; Barlow, A.; Coppock, C.; Stocker, M.; Harriger, D.; Wells, E.; Pachepsky, Y.; Macarisin, D. Prevalence of generic Listeria spp. and Listeria monocytogenes in surface waters in the Mid-Atlantic region of the United States. J. Food Prot. 2018, 81, 147. [Google Scholar]
- Falardeau, J.; Johnson, R.P.; Pagotto, F.; Wang, S. Occurrence, characterization, and potential predictors of verotoxigenic Escherichia coli, Listeria monocytogenes, and Salmonella in surface water used for produce irrigation in the Lower Mainland of British Columbia, Canada. PLoS ONE 2017, 12, e0185437. [Google Scholar] [CrossRef] [Green Version]
- Stea, E.C.; Purdue, L.M.; Jamieson, R.C.; Yost, C.K.; Truelstrup Hansen, L. Comparison of the Prevalences and Diversities of Listeria Species and Listeria monocytogenes in an Urban and a Rural Agricultural Watershed. Appl. Environ. Microbiol. 2015, 81, 3812–3822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weller, D.; Wiedmann, M.; Strawn, L.K. Irrigation Is Significantly Associated with an Increased Prevalence of Listeria monocytogenes in Produce Production Environments in New York State. J. Food Prot. 2015, 78, 1132–1141. [Google Scholar] [CrossRef] [PubMed]
- Chapin, T.K.; Nightingale, K.K.; Worobo, R.W.; Wiedmann, M.; Strawn, L.K. Geographical and meteorological factors associated with isolation of Listeria species in New York State produce production and natural environments. J. Food Prot. 2014, 77, 1919–1928. [Google Scholar] [CrossRef]
- Linke, K.; Ruckerl, I.; Brugger, K.; Karpiskova, R.; Walland, J.; Muri-Klinger, S.; Tichy, A.; Wagner, M.; Stessl, B. Reservoirs of listeria species in three environmental ecosystems. Appl. Environ. Microbiol. 2014, 80, 5583–5592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, G.; Strawn, L.K.; Ottesen, A.R.; Ramachandran, P.; Reed, E.A.; Zheng, J.; Boyer, R.R.; Rideout, S.L. Correlation of Salmonella enterica and Listeria monocytogenes in Irrigation Water to Environmental Factors, Fecal Indicators, and Bacterial Communities. Front. Microbiol. 2020, 11, 557289. [Google Scholar] [CrossRef] [PubMed]
- Sauders, B.D.; Overdevest, J.; Fortes, E.; Windham, K.; Schukken, Y.; Lembo, A.; Wiedmann, M. Diversity of Listeria species in urban and natural environments. Appl. Environ. Microbiol. 2012, 78, 4420–4433. [Google Scholar] [CrossRef] [Green Version]
- Strawn, L.K.; Fortes, E.D.; Bihn, E.A.; Nightingale, K.K.; Grohn, Y.T.; Worobo, R.W.; Wiedmann, M.; Bergholz, P.W. Landscape and meteorological factors affecting prevalence of three food-borne pathogens in fruit and vegetable farms. Appl. Environ. Microbiol. 2013, 79, 588–600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colburn, K.G.; Kaysner, C.A.; Abeyta, C., Jr.; Wekell, M.M. Listeria species in a California coast estuarine environment. Appl. Environ. Microbiol. 1990, 56, 2007–2011. [Google Scholar] [CrossRef] [Green Version]
- Wilkes, G.; Edge, T.; Gannon, V.; Jokinen, C.; Lyautey, E.; Medeiros, D.; Neumann, N.; Ruecker, N.; Topp, E.; Lapen, D.R. Seasonal relationships among indicator bacteria, pathogenic bacteria, Cryptosporidium oocysts, Giardia cysts, and hydrological indices for surface waters within an agricultural landscape. Water Res. 2009, 43, 2209–2223. [Google Scholar] [CrossRef] [PubMed]
- Lyautey, E.; Lapen, D.R.; Wilkes, G.; McCleary, K.; Pagotto, F.; Tyler, K.; Hartmann, A.; Piveteau, P.; Rieu, A.; Robertson, W.J.; et al. Distribution and characteristics of Listeria monocytogenes isolates from surface waters of the South Nation River watershed, Ontario, Canada. Appl. Environ. Microbiol. 2007, 73, 5401–5410. [Google Scholar] [CrossRef] [Green Version]
- Ijabadeniyi, O.; Debusho, L.K.; Vanderlinde, M.; Buys, E.M. Irrigation water as a potential preharvest source of bacterial contamination of vegetables. J. Food Saf. 2011, 31, 452–461. [Google Scholar] [CrossRef] [Green Version]
- Frances, N.; Hornby, H.; Hunter, P.R. The isolation of Listeria species from fresh-water sites in Cheshire and North Wales. Epidemiol. Infect. 1991, 107, 235–238. [Google Scholar] [CrossRef] [Green Version]
- Rodas-Suarez, O.R.; Flores-Pedroche, J.F.; Betancourt-Rule, J.M.; Quinones-Ramirez, E.I.; Vazquez-Salinas, C. Occurrence and antibiotic sensitivity of Listeria monocytogenes strains isolated from oysters, fish, and estuarine water. Appl. Environ. Microbiol. 2006, 72, 7410–7412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arvanitidou, M.; Papa, A.; Constantinidis, T.C.; Danielides, V.; Katsouyannopoulos, V. The occurrence of Listeria spp. and Salmonella spp. in surface waters. Microbiol. Res. 1997, 152, 395–397. [Google Scholar] [CrossRef]
- Schaffter, N.; Zumstein, J.; Parriaux, A.l. Factors influencing the bacteriological water quality in mountainous surface and groundwaters. Acta Hydrochim. Hydrobiol. 2004, 32, 225–234. [Google Scholar] [CrossRef]
- Macarisin, D. Listeria monocytogenes Prevalence and Population Diversity in Surface Waters. In Proceedings of the Global Water and Food Safety Summit, College Park, MD, USA, 19–21 November 2019. [Google Scholar]
- Qing, J.; Barlow, A.; Stocker, M.; Pachepsky, Y.; Allard, M.; Brown, E.; Chen, Y.; Macarisin, D. Occurrence and population diversity of Listeria monocytogenes in two irrigation ponds in Maryland. J. Food Prot. 2019, 82, 299. [Google Scholar]
- Macarisin, D.; Qing, J.; Harriger, D.; Picard, R.; Wells, E.; Pachepsky, Y.; Allard, M.; Brown, E.; Chen, Y. Listeria monocytogenes levels and population diversity in surface waters in the United States Mid-Atlantic region. J. Food Prot. 2019, 82, 299. [Google Scholar]
- Maury, M.M.; Tsai, Y.H.; Charlier, C.; Touchon, M.; Chenal-Francisque, V.; Leclercq, A.; Criscuolo, A.; Gaultier, C.; Roussel, S.; Brisabois, A.; et al. Uncovering Listeria monocytogenes hypervirulence by harnessing its biodiversity. Nat. Genet. 2016, 48, 308–313. [Google Scholar] [CrossRef] [Green Version]
- Orsi, R.H.; den Bakker, H.C.; Wiedmann, M. Listeria monocytogenes lineages: Genomics, evolution, ecology, and phenotypic characteristics. Int. J. Med. Microbiol. 2011, 301, 79–96. [Google Scholar] [CrossRef]
- Macarisin, D.; Qing, J.; Barlow, A.; Coppock, C.; Harriger, D.; Wells, E.; Pachepsky, Y. Levels of Listeria monocytogenes and bacterial fecal indicators in surface waters in the Mid-Atlantic region of the United States. J. Food Prot. 2018, 81, 147. [Google Scholar]
- Sheth, I.; Li, F.; Hur, M.; Laasri, A.; Jesus, A.J.D.; Kwon, H.J.; Macarisin, D.; Hammack, T.S.; Jinneman, K.; Chen, Y. Comparison of three enrichment schemes for the detection of low levels of desiccation-stressed Listeria spp. from select environmental surfaces. Food Control. 2018, 84, 493–498. [Google Scholar] [CrossRef]
- Hitchins, A.D.; Jinneman, K.; Chen, Y. Food and Drug Administration Bacteriological Analytical Manual Chapter 10, Detection and Enumeration of Listeria monocytogenes in Foods. Available online: http://www.fda.gov/Food/FoodScienceResearch/LaboratoryMethods/ucm071400.htm (accessed on 12 June 2021).
- Ottesen, A.; Ramachandran, P.; Reed, E.; White, J.R.; Hasan, N.; Subramanian, P.; Ryan, G.; Jarvis, K.; Grim, C.; Daquiqan, N.; et al. Enrichment dynamics of Listeria monocytogenes and the associated microbiome from naturally contaminated ice cream linked to a listeriosis outbreak. BMC Microbiol. 2016, 16, 275. [Google Scholar] [CrossRef] [PubMed]
- Besse, N.G.; Barre, L.; Buhariwalla, C.; Vignaud, M.L.; Khamissi, E.; Decourseulles, E.; Nirsimloo, M.; Chelly, M.; Kalmokoff, M. The overgrowth of Listeria monocytogenes by other Listeria spp. in food samples undergoing enrichment cultivation has a nutritional basis. Int. J. Food Microbiol. 2010, 136, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Dailey, R.C.; Welch, L.J.; Hitchins, A.D.; Smiley, R.D. Effect of Listeria seeligeri or Listeria welshimeri on Listeria monocytogenes detection in and recovery from buffered Listeria enrichment broth. Food Microbiol. 2015, 46, 528–534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oravcova, K.; Trncikova, T.; Kuchta, T.; Kaclikova, E. Limitation in the detection of Listeria monocytogenes in food in the presence of competing Listeria innocua. J. Appl. Microbiol. 2008, 104, 429–437. [Google Scholar] [CrossRef] [PubMed]
- Bruhn, J.B.; Vogel, B.F.; Gram, L. Bias in the Listeria monocytogenes enrichment procedure: Lineage 2 strains outcompete lineage 1 strains in University of Vermont selective enrichments. Appl. Environ. Microbiol. 2005, 71, 961–967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorski, L.; Walker, S.; Liang, A.S.; Nguyen, K.M.; Govoni, J.; Carychao, D.; Cooley, M.B.; Mandrell, R.E. Comparison of subtypes of Listeria monocytogenes isolates from naturally contaminated watershed samples with and without a selective secondary enrichment. PLoS ONE 2014, 9, e92467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reissbrodt, R. New chromogenic plating media for detection and enumeration of pathogenic Listeria spp.—An overview. Int. J. Food Microbiol. 2004, 95, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Pouillot, R.; Burall, L.S.; Strain, E.A.; Van Doren, J.M.; De Jesus, A.J.; Laasri, A.; Wang, H.; Ali, L.; Tatavarthy, A.; et al. Comparative evaluation of direct plating and most probable number for enumeration of low levels of Listeria monocytogenes in naturally contaminated ice cream products. Int. J. Food Microbiol. 2017, 241, 15–22. [Google Scholar] [CrossRef] [Green Version]
- CDC. Antibiotic Resistance Threats in the United States, 2019; U.S. Department of Health and Human Services, CDC: Atlanta, GA, USA, 2019. [Google Scholar]
- WHO. Antimicrobial Resistance: Global Report on Surveillance; World Health Organization: Geneva, Switzerland, 2014. [Google Scholar]
- Sommer, M.O.; Church, G.M.; Dantas, G. The human microbiome harbors a diverse reservoir of antibiotic resistance genes. Virulence 2010, 1, 299–303. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Yuan, Q.; Mathieu, J.; Stadler, L.; Senehi, N.; Sun, R.; Alvarez, P.J.J. Antibiotic resistance genes from livestock waste: Occurrence, dissemination, and treatment. NPJ Clean Water 2020, 3, 4. [Google Scholar] [CrossRef] [Green Version]
- Popowska, M.; Rzeczycka, M.; Miernik, A.; Krawczyk-Balska, A.; Walsh, F.; Duffy, B. Influence of soil use on prevalence of tetracycline, streptomycin, and erythromycin resistance and associated resistance genes. Antimicrob. Agents Chemother. 2012, 56, 1434–1443. [Google Scholar] [CrossRef] [Green Version]
- Heuer, H.; Schmitt, H.; Smalla, K. Antibiotic resistance gene spread due to manure application on agricultural fields. Curr. Opin. Microbiol. 2011, 14, 236–243. [Google Scholar] [CrossRef]
- Xu, Y.; Li, H.; Shi, R.; Lv, J.; Li, B.; Yang, F.; Zheng, X.; Xu, J. Antibiotic resistance genes in different animal manures and their derived organic fertilizer. Environ. Sci. Eur. 2020, 32, 102. [Google Scholar] [CrossRef]
- Karkman, A.; Johnson, T.A.; Lyra, C.; Stedtfeld, R.D.; Tamminen, M.; Tiedje, J.M.; Virta, M. High-throughput quantification of antibiotic resistance genes from an urban wastewater treatment plant. FEMS Microbiol. Ecol. 2016, 92. [Google Scholar] [CrossRef] [Green Version]
- Berendonk, T.U.; Manaia, C.M.; Merlin, C.; Fatta-Kassinos, D.; Cytryn, E.; Walsh, F.; Bürgmann, H.; Sørum, H.; Norström, M.; Pons, M.N.; et al. Tackling antibiotic resistance: The environmental framework. Nat. Rev. Microbiol. 2015, 13, 310–317. [Google Scholar] [CrossRef]
- Meinersmann, R.J.; Berrang, M.E.; Jackson, C.R.; Fedorka-Cray, P.; Ladely, S.; Little, E.; Frye, J.G.; Mattsson, B. Salmonella, Campylobacter and Enterococcus spp.: Their antimicrobial resistance profiles and their spatial relationships in a synoptic study of the Upper Oconee River basin. Microb. Ecol. 2008, 55, 444–452. [Google Scholar] [CrossRef]
- Dolejská, M.; Bierosová, B.; Kohoutová, L.; Literák, I.; Cízek, A. Antibiotic-resistant Salmonella and Escherichia coli isolates with integrons and extended-spectrum beta-lactamases in surface water and sympatric black-headed gulls. J. Appl. Microbiol. 2009, 106, 1941–1950. [Google Scholar] [CrossRef]
- Casanova, L.M.; Hill, V.R.; Sobsey, M.D. Antibiotic-resistant Salmonella in swine wastes and farm surface waters. Lett. Appl. Microbiol. 2020, 71, 117–123. [Google Scholar] [CrossRef] [PubMed]
- McEgan, R.; Chandler, J.C.; Goodridge, L.D.; Danyluk, M.D. Diversity of Salmonella isolates from central Florida surface waters. Appl. Environ. Microbiol. 2014, 80, 6819–6827. [Google Scholar] [CrossRef] [Green Version]
- Jokinen, C.C.; Koot, J.; Cole, L.; Desruisseau, A.; Edge, T.A.; Khan, I.U.; Koning, W.; Lapen, D.R.; Pintar, K.D.; Reid-Smith, R.; et al. The distribution of Salmonella enterica serovars and subtypes in surface water from five agricultural regions across Canada. Water Res. 2015, 76, 120–131. [Google Scholar] [CrossRef] [PubMed]
- Servais, P.; Passerat, J. Antimicrobial resistance of fecal bacteria in waters of the Seine river watershed (France). Sci. Total Environ. 2009, 408, 365–372. [Google Scholar] [CrossRef] [PubMed]
- Agga, G.E.; Arthur, T.M.; Durso, L.M.; Harhay, D.M.; Schmidt, J.W. Antimicrobial-Resistant Bacterial Populations and Antimicrobial Resistance Genes Obtained from Environments Impacted by Livestock and Municipal Waste. PLoS ONE 2015, 10, e0132586. [Google Scholar] [CrossRef] [PubMed]
- Titilawo, Y.; Obi, L.; Okoh, A. Antimicrobial resistance determinants of Escherichia coli isolates recovered from some rivers in Osun State, South-Western Nigeria: Implications for public health. Sci. Total. Environ. 2015, 523, 82–94. [Google Scholar] [CrossRef]
- Lyimo, B.; Buza, J.; Subbiah, M.; Smith, W.; Call, D.R. Comparison of antibiotic resistant Escherichia coli obtained from drinking water sources in northern Tanzania: A cross-sectional study. BMC Microbiol. 2016, 16, 254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Yu, D.; He, S.; Ye, H.; Zhang, L.; Wen, Y.; Zhang, W.; Shu, L.; Chen, S. Prevalence of Antibiotic-Resistant Escherichia coli in Drinking Water Sources in Hangzhou City. Front. Microbiol. 2017, 8, 1133. [Google Scholar] [CrossRef]
- Watkinson, A.J.; Micalizzi, G.B.; Graham, G.M.; Bates, J.B.; Costanzo, S.D. Antibiotic-resistant Escherichia coli in wastewaters, surface waters, and oysters from an urban riverine system. Appl. Environ. Microbiol. 2007, 73, 5667–5670. [Google Scholar] [CrossRef] [Green Version]
- Blaak, H.; de Kruijf, P.; Hamidjaja, R.A.; van Hoek, A.H.; de Roda Husman, A.M.; Schets, F.M. Prevalence and characteristics of ESBL-producing E. coli in Dutch recreational waters influenced by wastewater treatment plants. Vet. Microbiol. 2014, 171, 448–459. [Google Scholar] [CrossRef]
- Bajaj, P.; Singh, N.S.; Kanaujia, P.K.; Virdi, J.S. Distribution and molecular characterization of genes encoding CTX-M and AmpC β-lactamases in Escherichia coli isolated from an Indian urban aquatic environment. Sci. Total Environ. 2015, 505, 350–356. [Google Scholar] [CrossRef] [PubMed]
- Jørgensen, S.B.; Søraas, A.V.; Arnesen, L.S.; Leegaard, T.M.; Sundsfjord, A.; Jenum, P.A. A comparison of extended spectrum β-lactamase producing Escherichia coli from clinical, recreational water and wastewater samples associated in time and location. PLoS ONE 2017, 12, e0186576. [Google Scholar] [CrossRef] [Green Version]
- Rice, E.W.; Messer, J.W.; Johnson, C.H.; Reasoner, D.J. Occurrence of high-level aminoglycoside resistance in environmental isolates of enterococci. Appl. Environ. Microbiol. 1995, 61, 374–376. [Google Scholar] [CrossRef] [Green Version]
- Moore, D.F.; Guzman, J.A.; McGee, C. Species distribution and antimicrobial resistance of enterococci isolated from surface and ocean water. J. Appl. Microbiol. 2008, 105, 1017–1025. [Google Scholar] [CrossRef]
- Lata, P.; Ram, S.; Agrawal, M.; Shanker, R. Enterococci in river Ganga surface waters: Propensity of species distribution, dissemination of antimicrobial-resistance and virulence-markers among species along landscape. BMC Microbiol. 2009, 9, 140. [Google Scholar] [CrossRef] [Green Version]
- Cho, S.; Hiott, L.M.; McDonald, J.M.; Barrett, J.B.; McMillan, E.A.; House, S.L.; Adams, E.S.; Frye, J.G.; Jackson, C.R. Diversity and antimicrobial resistance of Enterococcus from the Upper Oconee Watershed, Georgia. J. Appl. Microbiol. 2020, 128, 1221–1233. [Google Scholar] [CrossRef] [PubMed]
- Iversen, A.; Kühn, I.; Franklin, A.; Möllby, R. High prevalence of vancomycin-resistant enterococci in Swedish sewage. Appl. Environ. Microbiol. 2002, 68, 2838–2842. [Google Scholar] [CrossRef] [Green Version]
- Novais, C.; Coque, T.M.; Ferreira, H.; Sousa, J.C.; Peixe, L. Environmental contamination with vancomycin-resistant enterococci from hospital sewage in Portugal. Appl. Environ. Microbiol. 2005, 71, 3364–3368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenberg Goldstein, R.E.; Micallef, S.A.; Gibbs, S.G.; George, A.; Claye, E.; Sapkota, A.; Joseph, S.W.; Sapkota, A.R. Detection of vancomycin-resistant enterococci (VRE) at four U.S. wastewater treatment plants that provide effluent for reuse. Sci. Total Environ. 2014, 466–467, 404–411. [Google Scholar] [CrossRef]
- Young, S.; Nayak, B.; Sun, S.; Badgley, B.D.; Rohr, J.R.; Harwood, V.J. Vancomycin-Resistant Enterococci and Bacterial Community Structure following a Sewage Spill into an Aquatic Environment. Appl. Environ. Microbiol. 2016, 82, 5653–5660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haberecht, H.B.; Nealon, N.J.; Gilliland, J.R.; Holder, A.V.; Runyan, C.; Oppel, R.C.; Ibrahim, H.M.; Mueller, L.; Schrupp, F.; Vilchez, S.; et al. Antimicrobial-Resistant Escherichia coli from Environmental Waters in Northern Colorado. J. Environ. Public Health 2019, 2019, 3862949. [Google Scholar] [CrossRef] [Green Version]
- Tokajian, S.; Moghnieh, R.; Salloum, T.; Arabaghian, H.; Alousi, S.; Moussa, J.; Abboud, E.; Youssef, S.; Husni, R. Extended-spectrum β-lactamase-producing Escherichia coli in wastewaters and refugee camp in Lebanon. Future Microbiol. 2018, 13, 81–95. [Google Scholar] [CrossRef] [Green Version]
- Hendriksen, R.S.; Munk, P.; Njage, P.; van Bunnik, B.; McNally, L.; Lukjancenko, O.; Röder, T.; Nieuwenhuijse, D.; Pedersen, S.K.; Kjeldgaard, J.; et al. Global monitoring of antimicrobial resistance based on metagenomics analyses of urban sewage. Nat. Commun. 2019, 10, 1124. [Google Scholar] [CrossRef]
- FDA. FDA Releases NARMS Strategic Plan, Announces October 2020 NARMS Virtual Public Meeting. Available online: https://www.fda.gov/animal-veterinary/cvm-updates/fda-releases-narms-strategic-plan-announces-october-2020-narms-virtual-public-meeting (accessed on 17 February 2021).
- EPA. Development and Implementation of a Surface Water Pilot Study Within the National Antimicrobial Resistance Monitoring System (NARMS). Available online: https://cfpub.epa.gov/si/si_public_record_Report.cfm?dirEntryId=350147&Lab=CESER (accessed on 17 February 2021).
- Allard, M.W.; Bell, R.; Ferreira, C.M.; Gonzalez-Escalona, N.; Hoffmann, M.; Muruvanda, T.; Ottesen, A.; Ramachandran, P.; Reed, E.; Sharma, S.; et al. Genomics of foodborne pathogens for microbial food safety. Curr. Opin. Biotechnol. 2018, 49, 224–229. [Google Scholar] [CrossRef]
- Lienau, E.K.; Strain, E.; Wang, C.; Zheng, J.; Ottesen, A.R.; Keys, C.E.; Hammack, T.S.; Musser, S.M.; Brown, E.W.; Allard, M.W.; et al. Identification of a salmonellosis outbreak by means of molecular sequencing. N. Engl. J. Med. 2011, 364, 981–982. [Google Scholar] [CrossRef] [PubMed]
- Feldgarden, M.; Brover, V.; Haft, D.H.; Prasad, A.B.; Slotta, D.J.; Tolstoy, I.; Tyson, G.H.; Zhao, S.; Hsu, C.-H.; McDermott, P.F.; et al. Validating the AMRFinder Tool and Resistance Gene Database by Using Antimicrobial Resistance Genotype-Phenotype Correlations in a Collection of Isolates. Antimicrob. Agents Chemother. 2019, 63, e00483-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffmann, M.; Miller, J.; Melka, D.; Allard, M.W.; Brown, E.W.; Pettengill, J.B. Temporal Dynamics of Salmonella enterica subsp. enterica Serovar Agona Isolates from a Recurrent Multistate Outbreak. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- FDA. FDA Resitome Tracker. Available online: https://www.fda.gov/animal-veterinary/national-antimicrobial-resistance-monitoring-system/global-resistome-data (accessed on 3 March 2021).
- Hoffmann, M.; Luo, Y.; Monday, S.R.; Gonzalez-Escalona, N.; Ottesen, A.R.; Muruvanda, T.; Wang, C.; Kastanis, G.; Keys, C.; Janies, D.; et al. Tracing Origins of the Salmonella Bareilly Strain Causing a Food-borne Outbreak in the United States. J. Infect. Dis. 2015. [Google Scholar] [CrossRef] [Green Version]
- Allard, M.W.; Strain, E.; Melka, D.; Bunning, K.; Musser, S.M.; Brown, E.W.; Timme, R. Practical Value of Food Pathogen Traceability through Building a Whole-Genome Sequencing Network and Database. J. Clin. Microbiol. 2016, 54, 1975–1983. [Google Scholar] [CrossRef] [Green Version]
- Allard, M.W.; Strain, E.; Rand, H.; Melka, D.; Correll, W.A.; Hintz, L.; Stevens, E.; Timme, R.; Lomonaco, S.; Chen, Y.; et al. Whole genome sequencing uses for foodborne contamination and compliance: Discovery of an emerging contamination event in an ice cream facility using whole genome sequencing. Infect. Genet. Evol. 2019, 73, 214–220. [Google Scholar] [CrossRef] [PubMed]
- Allard, M.W.; Zheng, J.; Cao, G.; Timme, R.; Stevens, E.; Brown, E.W. Food Safety Genomics and Connections to One Health and the Clinical Microbiology Laboratory. Clin. Lab. Med. 2020, 40, 553–563. [Google Scholar] [CrossRef]
- Allard, M.W.; Luo, Y.; Strain, E.; Pettengill, J.; Timme, R.; Wang, C.; Li, C.; Keys, C.E.; Zheng, J.; Stones, R.; et al. On the evolutionary history, population genetics and diversity among isolates of Salmonella Enteritidis PFGE pattern JEGX01.0004. PLoS ONE 2013, 8, e55254. [Google Scholar] [CrossRef] [PubMed]
- Pightling, A.W.; Pettengill, J.B.; Luo, Y.; Baugher, J.D.; Rand, H.; Strain, E. Interpreting Whole-Genome Sequence Analyses of Foodborne Bacteria for Regulatory Applications and Outbreak Investigations. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Wilson, D.; Dolan, G.; Aird, H.; Sorrell, S.; Dallman, T.J.; Jenkins, C.; Robertson, L.; Gorton, R. Farm-to-fork investigation of an outbreak of Shiga toxin-producing Escherichia coli O157. Microb. Genom. 2018, 4, e000160. [Google Scholar] [CrossRef]
- Zheng, J.; Luo, Y.; Reed, E.; Bell, R.; Brown, E.W.; Hoffmann, M. Whole-Genome Comparative Analysis of Salmonella enterica Serovar Newport Strains Reveals Lineage-Specific Divergence. Genome Biol. Evol. 2017, 9, 1047–1050. [Google Scholar] [CrossRef] [Green Version]
- JIFSAN. Global Water and Food Safety Summit. Available online: https://jifsan.umd.edu/events/water-safety (accessed on 2 February 2021).
- Schulz, J.; Ruddat, I.; Hartung, J.; Hamscher, G.; Kemper, N.; Ewers, C. Antimicrobial-Resistant Escherichia coli Survived in Dust Samples for More than 20 Years. Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maguire, M.; Kase, J.A.; Roberson, D.; Muruvanda, T.; Brown, E.W.; Allard, M.; Musser, S.M.; González-Escalona, N. Precision long-read metagenomics sequencing for food safety by detection and assembly of Shiga toxin-producing Escherichia coli in irrigation water. PLoS ONE 2021, 16, e0245172. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, S.; Frosth, S.; Svensson, L.; Fernstrom, L.L.; Skarin, H.; Hansson, I. Detection of Campylobacter spp. in water by dead-end ultrafiltration and application at farm level. J. Appl. Microbiol. 2019, 127, 1270–1279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sikorski, M.J.; Levine, M.M. Reviving the "Moore Swab": A Classic Environmental Surveillance Tool Involving Filtration of Flowing Surface Water and Sewage Water to Recover Typhoidal Salmonella Bacteria. Appl. Environ. Microbiol. 2020, 86, e00060-00020. [Google Scholar] [CrossRef]
- Fernandez, H.; Otth, L.; Wilson, M. Isolation of thermotolerant species of Campylobacter from river water using two collection methods. Arch. Med. Vet. 2003, 35. [Google Scholar] [CrossRef]
- Bisha, B.; Perez-Mendez, A.; Danyluk, M.D.; Goodridge, L.D. Evaluation of modified moore swabs and continuous flow centrifugation for concentration of Salmonella and Escherichia coli O157:H7 from large volumes of water. J. Food Prot. 2011, 74, 1934–1937. [Google Scholar] [CrossRef]
- McEgan, R.; Rodrigues, C.A.; Sbodio, A.; Suslow, T.V.; Goodridge, L.D.; Danyluk, M.D. Detection of Salmonella spp. from large volumes of water by modified Moore swabs and tangential flow filtration. Lett. Appl. Microbiol. 2013, 56, 88–94. [Google Scholar] [CrossRef]
- Sbodio, A.; Maeda, S.; Lopez-Velasco, G.; Suslow, T.V. Modified Moore swab optimization and validation in capturing E. coli O157:H7 and Salmonella enterica in large volume field samples of irrigation water. Food Res. Int. 2013, 51, 654–662. [Google Scholar] [CrossRef]
- Hill, V.R.; Polaczyk, A.L.; Hahn, D.; Narayanan, J.; Cromeans, T.L.; Roberts, J.M.; Amburgey, J.E. Development of a rapid method for simultaneous recovery of diverse microbes in drinking water by ultrafiltration with sodium polyphosphate and surfactants. Appl. Environ. Microbiol. 2005, 71, 6878–6884. [Google Scholar] [CrossRef] [Green Version]
- Smith, C.M.; Hill, V.R. Dead-end hollow-fiber ultrafiltration for recovery of diverse microbes from water. Appl. Environ. Microbiol. 2009, 75, 5284–5289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mull, B.; Hill, V.R. Recovery of diverse microbes in high turbidity surface water samples using dead-end ultrafiltration. J. Microbiol. Methods 2012, 91, 429–433. [Google Scholar] [CrossRef] [Green Version]
- Mull, B.; Hill, V.R. Recovery and detection of Escherichia coli O157:H7 in surface water, using ultrafiltration and real-time PCR. Appl. Environ. Microbiol. 2009, 75, 3593–3597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pachepsky, Y.; Shelton, D.R.; McLain, J.E.T.; Patel, J.; Mandrell, R.E. Irrigation Waters as a Source of Pathogenic Microorganisms in Produce: A Review. In Advances in Agronomy; Sparks, D.L., Ed.; Academic Press: Cambridge, MA, USA, 2011; Volume 113, pp. 75–141. [Google Scholar]
- Pandey, P.K.; Kass, P.H.; Soupir, M.L.; Biswas, S.; Singh, V.P. Contamination of water resources by pathogenic bacteria. AMB Express 2014, 4, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FDA. Efficacy Protocol for Reduction of Foodborne Bacteria in Preharvest Agricultural Water. Available online: https://www.fda.gov/food/food-safety-modernization-act-fsma/efficacy-protocol-reduction-foodborne-bacteria-preharvest-agricultural-water (accessed on 17 February 2021).
- Dery, J.; Gerrity, D.; Rock, C. Minimizing Risks: Use of Surface Water in Pre-Harvest Agricultural Irrigation; Part II: Sodium and Calcium Hypochlorite (Chlorine) Treatment Methods. Available online: https://extension.arizona.edu/sites/extension.arizona.edu/files/pubs/az1831-2020.pdf (accessed on 17 February 2021).
- LeChevallier, M.W.; Evans, T.M.; Seidler, R.J. Effect of turbidity on chlorination efficiency and bacterial persistence in drinking water. Appl. Environ. Microbiol. 1981, 42, 159–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Obi, C.; Igumbor, J.; Momba, M.; Samie, A. Interplay of factors involving chlorine dose, turbidity flow capacity and pH on microbial quality of drinking water in small water treatment plants. Water SA 2008, 34, 565–572. [Google Scholar] [CrossRef] [Green Version]
- Silverman, G.S.; Nagy, L.A.; Olson, B.H. Variations in particulate matter, algae, and bacteria in an uncovered, finished-drinking-water reservoir. J. AWWA 1983, 75, 191–196. [Google Scholar] [CrossRef]
- Amrutha, B.; Sundar, K.; Shetty, P.H. Study on E. coli and Salmonella biofilms from fresh fruits and vegetables. J. Food Sci. Technol. 2017, 54, 1091–1097. [Google Scholar] [CrossRef] [Green Version]
- Møretrø, T.; Heir, E.; Nesse, L.L.; Vestby, L.K.; Langsrud, S. Control of Salmonella in food related environments by chemical disinfection. Food Res. Int. 2012, 45, 532–544. [Google Scholar] [CrossRef]


{kind=link}
| Method | Ease of Use a | Field Deployable | Easy to Transport | Volume Sampled | Cost b |
|---|---|---|---|---|---|
| Grab | easy | Yes | no | 100 mL to 1 L c | $ |
| Moore swab | easy | Yes | yes | NA | $ |
| modified Moore Swab | intermediate | Yes | yes | 10 L | $$ |
| Tangential Filtration | complicated | not readily | no | 10 L-100 L | $$$ |
| Dead End Ultrafiltration | intermediate | Yes | yes | up to 100 L | $$$ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bell, R.L.; Kase, J.A.; Harrison, L.M.; Balan, K.V.; Babu, U.; Chen, Y.; Macarisin, D.; Kwon, H.J.; Zheng, J.; Stevens, E.L.; et al. The Persistence of Bacterial Pathogens in Surface Water and Its Impact on Global Food Safety. Pathogens 2021, 10, 1391. https://doi.org/10.3390/pathogens10111391
Bell RL, Kase JA, Harrison LM, Balan KV, Babu U, Chen Y, Macarisin D, Kwon HJ, Zheng J, Stevens EL, et al. The Persistence of Bacterial Pathogens in Surface Water and Its Impact on Global Food Safety. Pathogens. 2021; 10(11):1391. https://doi.org/10.3390/pathogens10111391
Chicago/Turabian StyleBell, Rebecca L., Julie A. Kase, Lisa M. Harrison, Kannan V. Balan, Uma Babu, Yi Chen, Dumitru Macarisin, Hee Jin Kwon, Jie Zheng, Eric L. Stevens, and et al. 2021. "The Persistence of Bacterial Pathogens in Surface Water and Its Impact on Global Food Safety" Pathogens 10, no. 11: 1391. https://doi.org/10.3390/pathogens10111391