
Linking Taxonomic, Phylogenetic and Functional Plant Diversity with Ecosystem Services of Cliffs and Screes in Greece

 , , ,
, , ,  ,
,
Abstract
:1. Introduction
2. Results
2.1. Plant Taxa of Cliffs and Screes
2.2. Biodiversity Patterns
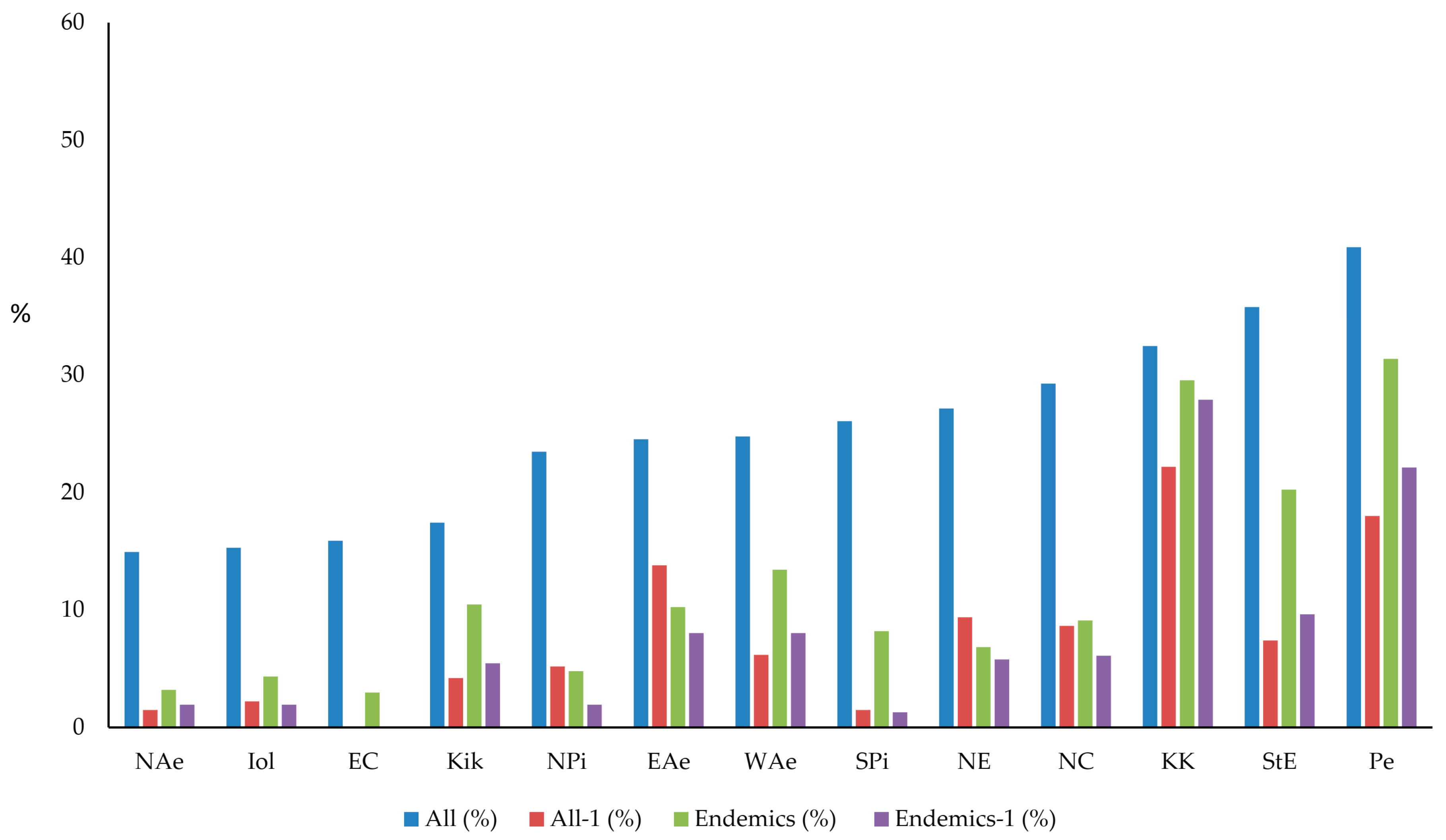
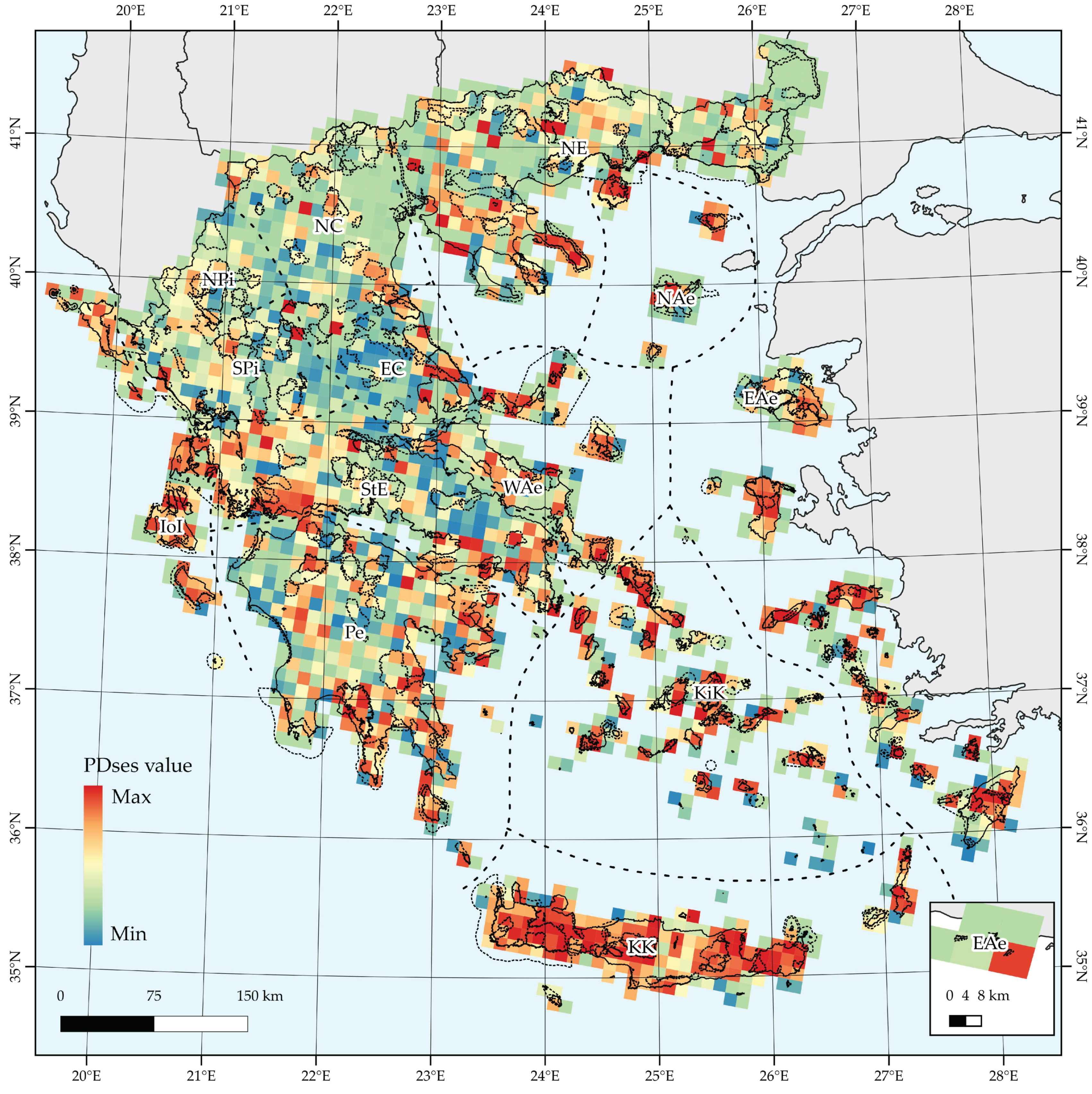
2.2.1. Taxonomic and Phylogenetic Diversity
2.2.2. Functional Diversity
- Life form: 22.43% of the 2033 cells are highlighted as the most diverse with five different life forms (Figure S5a). Almost one-fifth of these cells (17.54%) are in the floristic region of Kriti-Karpathos (KK), followed by Pe (15.79%) and StE (14.25%).
- Growth form: A small fraction of the total cells (3.64%) are highlighted as the most diverse with 8–10 different growth forms (Figure S5b). One-fourth of these cells (24.32%) are in the floristic region of StE, followed by Pe (16.22%), NPi and NC (13.51% each).
- Longevity: More than one-third of the total cells (37.58%) are highlighted as the most diverse with three different longevity statuses, i.e., annual, biennial, and perennial (Figure S5c). Nearly one fifth of these cells (18.59%) are located in the floristic region of Pe, followed by KK (16.88%) and StE (14.53%).
- Dispersal mode: More than one-tenth of the total cells (11.02%) are highlighted as the most diverse with 6–7 different dispersal modes, i.e., autochory, barochory, anemochory, and zoochory with epizoochory, endozoochory, and myrmecochory (Figure S5d). Nearly one fifth of these cells (18.30%) are located in the floristic region of Pe, followed by StE (14.29%) and KK (14.29%).
- Pollination type: Almost one-third of the total cells (31.14%) are highlighted as the most diverse with 3-4 different pollination types (Figure S5e). Nearly one fifth of these cells (15.17%) are located in the floristic region of Pe, followed by KK (14.54%) and StE (12.16%).
- Petal color: 27 out of 2033 cells (1.32%) are highlighted as the most diverse with 26–39 different petal colors (Figure S5f). Most of these cells (8 out of 27: 29.63%) are in the floristic region of StE, followed by Pe, SPi, and NPi (5 cells at each: 18.52%).
2.2.3. Distribution and Area of Cliffs and Screes in SACs
2.3. Ecosystem Services Indicators
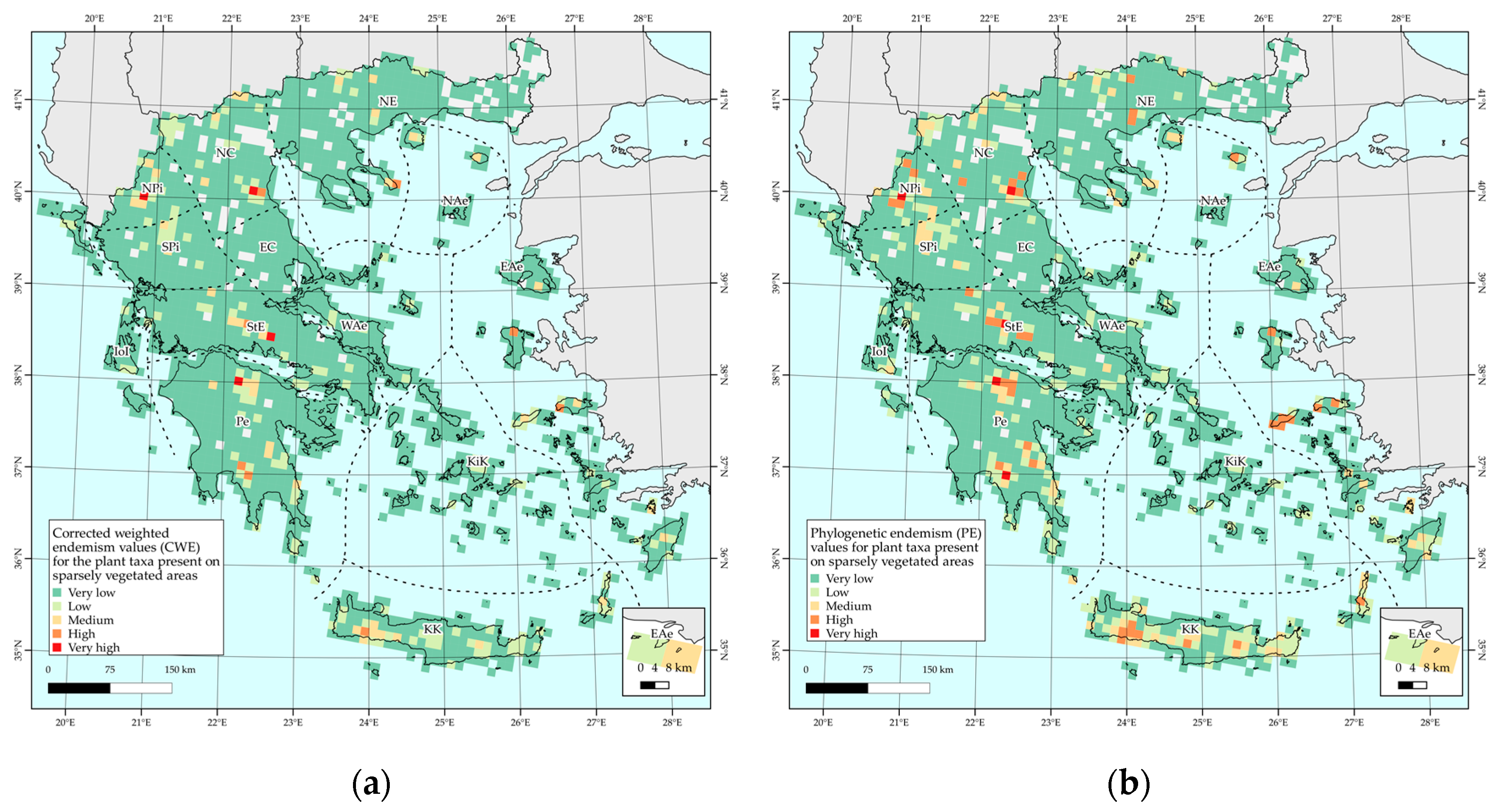
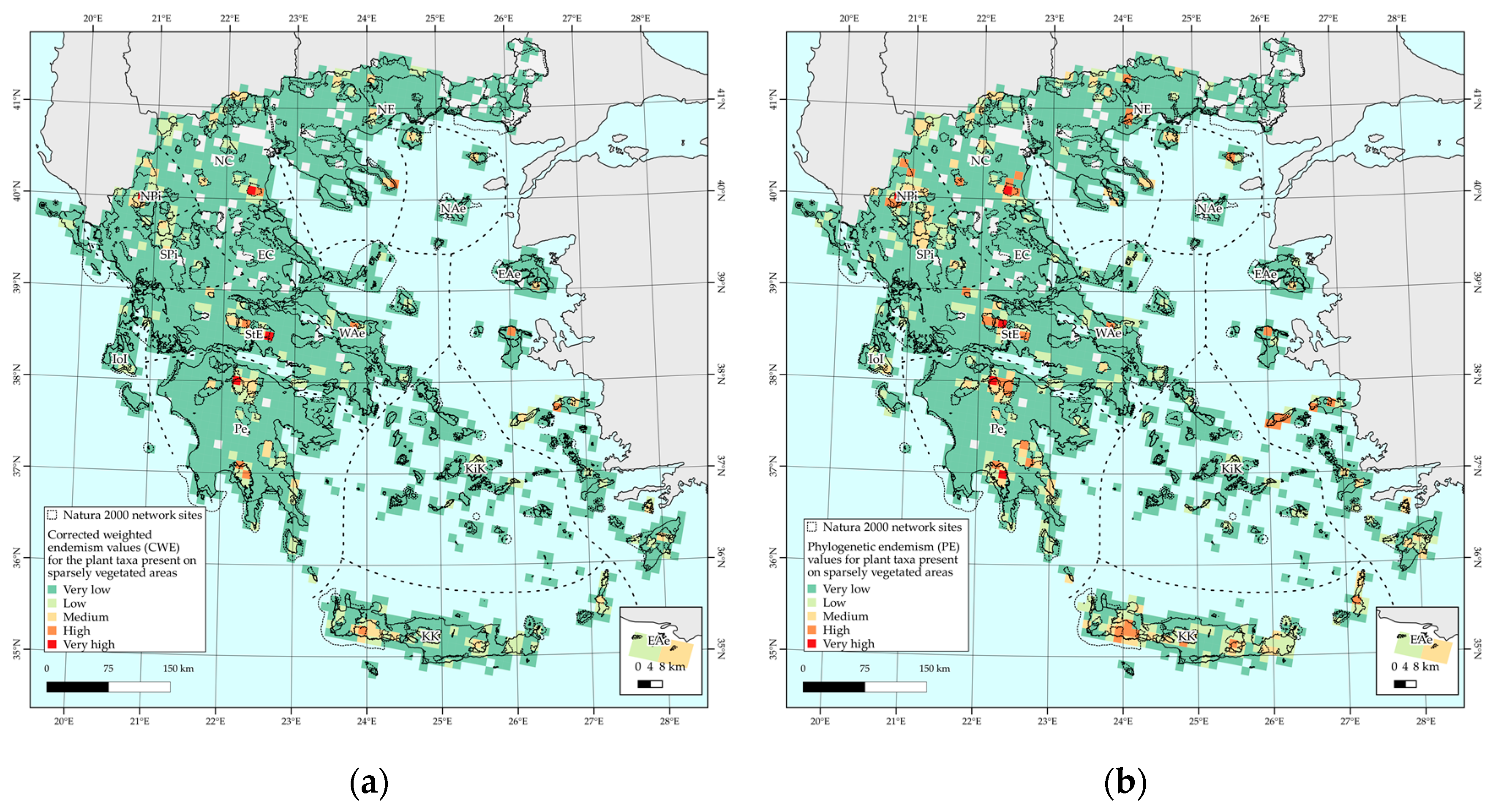
2.4. Biodiversity Hotspots and Protected Area Network Overlap
3. Discussion
3.1. Taxonomic and Phylogenetic Patterns
3.2. Functional Diversity and Ecosystem Services
3.3. Ecosystem Services and MAES Implementation
4. Materials and Methods
4.1. Plant Taxa of Cliffs and Screes
4.2. Biodiversity Patterns
4.3. Ecosystem Services
4.4. Priority Hotspots and Protected Area Overlap
4.5. Thematic Mapping
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Maes, J.; Teller, A.; Erhard, M.; Liquete, C.; Braat, L.; Berry, P.; Egoh, B.; Puydarrieux, P.; Fiorina, C.; Santos-Martín, F.; et al. Mapping and Assessment of Ecosystems and Their Services-An Analytical Framework for Ecosystem Assessments under Action 5 of the EU Biodiversity Strategy to 2020. 2013. Available online: https://ec.europa.eu/environment/nature/knowledge/ecosystem_assessment/pdf/MAESWorkingPaper2013.pdf (accessed on 25 February 2021).
- Kokkoris, I.P.; Drakou, E.G.; Maes, J.; Dimopoulos, P. Ecosystem services supply in protected mountains of Greece: Setting the baseline for conservation management. Int. J. Biodivers. Sci. Ecosyst. Serv. Manag. 2018, 14. [Google Scholar] [CrossRef] [Green Version]
- Kokkoris, I.P.; Mallinis, G.; Bekri, E.S.; Vlami, V.; Zogaris, S.; Chrysafis, I.; Mitsopoulos, I.; Dimopoulos, P. National set of MAES indicators in Greece: Ecosystem services and management implications. Forests 2020, 11, 595. [Google Scholar] [CrossRef]
- Kokkoris, I.P.; Dimopoulos, P.; Xystrakis, F.; Tsiripidis, I. National scale ecosystem condition assessment with emphasis on forest types in Greece. One Ecosyst. 2018, 3. [Google Scholar] [CrossRef] [Green Version]
- Kokkoris, I.P.; Bekri, E.S.; Skuras, D.; Vlami, V.; Zogaris, S.; Maroulis, G.; Dimopoulos, D.; Dimopoulos, P. Integrating MAES implementation into protected area management under climate change: A fine-scale application in Greece. Sci. Total Environ. 2019, 695. [Google Scholar] [CrossRef] [PubMed]
- Hatziiordanou, L.; Fitoka, E.; Hadjicharalampous, E.; Eleftheria Votsi, N.; Palaskas, D.; Abdul Malak, D. Indicators for mapping and assessment of ecosystem condition and of the ecosystem service habitat maintenance in support of the EU biodiversity strategy to 2020. One Ecosyst. 2019, 4. [Google Scholar] [CrossRef]
- Vlami, V.; Kokkoris, I.P.; Zogaris, S.; Cartalis, C.; Kehayias, G.; Dimopoulos, P. Cultural landscapes and attributes of “culturalness” in protected areas: An exploratory assessment in Greece. Sci. Total Environ. 2017, 595. [Google Scholar] [CrossRef]
- Nikolaidou, C.; Votsi, N.E.P.; Sgardelis, S.P.; Maxwell Halley, J.; Pantis, J.; Tsiafouli, M.A. Ecosystem Service capacity is higher in areas of multiple designation types. One Ecosyst. 2017, 2. [Google Scholar] [CrossRef] [Green Version]
- Kotsiras, K.; Kokkoris, I.P.; Strid, A.; Dimopoulos, P. Integrating plant diversity data into mapping and assessment of ecosystem and their services (MAES) implementation in Greece: Woodland and forest pilot. Forests 2020, 11, 956. [Google Scholar] [CrossRef]
- LIFE-IP 4 NATURA. Available online: https://edozoume.gr/en/ (accessed on 25 February 2021).
- Kougioumoutzis, K.; Kokkoris, I.P.; Panitsa, M.; Kallimanis, A.; Strid, A.; Dimopoulos, P. Plant endemism centres and biodiversity hotspots in Greece. Biology 2021, 10, 72. [Google Scholar] [CrossRef] [PubMed]
- Haight, J.; Hammill, E. Protected areas as potential refugia for biodiversity under climatic change. Biol. Conserv. 2020, 241. [Google Scholar] [CrossRef]
- Hoffmann, S.; Irl, S.D.H.; Beierkuhnlein, C. Predicted climate shifts within terrestrial protected areas worldwide. Nat. Commun. 2019, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monsarrat, S.; Jarvie, S.; Svenning, J.C. Anthropocene refugia: Integrating history and predictive modelling to assess the space available for biodiversity in a human-dominated world. Philos. Trans. R. Soc. B Biol. Sci. 2019, 374. [Google Scholar] [CrossRef] [Green Version]
- Stein, B.A.; Staudt, A.; Cross, M.S.; Dubois, N.S.; Enquist, C.; Griffis, R.; Hansen, L.J.; Hellmann, J.J.; Lawler, J.J.; Nelson, E.J.; et al. Preparing for and managing change: Climate adaptation for biodiversity and ecosystems. Front. Ecol. Environ. 2013, 11. [Google Scholar] [CrossRef]
- Veron, S.; Faith, D.P.; Pellens, R.; Pavoine, S. Priority areas for phylogenetic diversity: Maximising gains in the mediterranean basin. In Phylogenetic Diversity: Applications and Challenges in Biodiversity Science; Springer International Publishing: Cham, Switzerland, 2018. [Google Scholar]
- Kougioumoutzis, K.; Kokkoris, I.P.; Panitsa, M.; Trigas, P.; Strid, A.; Dimopoulos, P. Plant diversity patterns and conservation implications under climate-change scenarios in the mediterranean: The case of crete (Aegean, Greece). Diversity 2020, 12, 270. [Google Scholar] [CrossRef]
- Kougioumoutzis, K.; Kokkoris, I.P.; Panitsa, M.; Trigas, P.; Strid, A.; Dimopoulos, P. Spatial phylogenetics, biogeographical patterns and conservation implications of the endemic flora of Crete (Aegean, Greece) under climate change scenarios. Biology 2020, 9, 199. [Google Scholar] [CrossRef] [PubMed]
- Kougioumoutzis, K.; Kokkoris, I.P.; Panitsa, M.; Strid, A.; Dimopoulos, P. Extinction Risk Assessment of the Greek Endemic Flora. Biology 2021, 10, 195. [Google Scholar] [CrossRef] [PubMed]
- Weinstein, B.G.; Tinoco, B.; Parra, J.L.; Brown, L.M.; McGuire, J.A.; Gary Stiles, F.; Graham, C.H. Taxonomic, phylogenetic, and trait beta diversity in south American hummingbirds. Am. Nat. 2014, 184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Webb, C.O.; Ackerly, D.D.; McPeek, M.A.; Donoghue, M.J. Phylogenies and community ecology. Annu. Rev. Ecol. Syst. 2002, 33, 475–505. [Google Scholar] [CrossRef] [Green Version]
- Cavender-Bares, J.; Kozak, K.H.; Fine, P.V.A.; Kembel, S.W. The merging of community ecology and phylogenetic biology. Ecol. Lett. 2009, 12. [Google Scholar] [CrossRef] [PubMed]
- Siefert, A.; Ravenscroft, C.; Weiser, M.D.; Swenson, N.G. Functional beta-diversity patterns reveal deterministic community assembly processes in eastern North American trees. Glob. Ecol. Biogeogr. 2013, 22. [Google Scholar] [CrossRef]
- Swenson, N.G. The assembly of tropical tree communities–the advances and shortcomings of phylogenetic and functional trait analyses. Ecography 2013, 36. [Google Scholar] [CrossRef]
- Mazel, F.; Wüest, R.O.; Lessard, J.P.; Renaud, J.; Ficetola, G.F.; Lavergne, S.; Thuiller, W. Global patterns of β-diversity along the phylogenetic time-scale: The role of climate and plate tectonics. Glob. Ecol. Biogeogr. 2017, 26. [Google Scholar] [CrossRef]
- Graham, C.H.; Parra, J.L.; Rahbek, C.; McGuire, J.A. Phylogenetic structure in tropical hummingbird communities. Proc. Natl. Acad. Sci. USA 2009, 106. [Google Scholar] [CrossRef] [Green Version]
- Raus, T.; Karadimou, E.; Dimopoulos, P. Taxonomic and functional plant diversity of the Santorini-Christiana island group (Aegean Sea, Greece). Willdenowia 2019, 49. [Google Scholar] [CrossRef] [Green Version]
- Rosenfeld, J.S. Functional redundancy in ecology and conservation. Oikos 2002, 98, 156–162. [Google Scholar] [CrossRef] [Green Version]
- Díaz, S.; Cabido, M. Vive la différence: Plant functional diversity matters to ecosystem processes. Trends Ecol. Evol. 2001, 16, 646–655. [Google Scholar] [CrossRef]
- Dwyer, J.M.; Hobbs, R.J.; Wainwright, C.E.; Mayfield, M.M. Climate moderates release from nutrient limitation in natural annual plant communities. Glob. Ecol. Biogeogr. 2015, 24. [Google Scholar] [CrossRef]
- Blonder, B.; Kapas, R.E.; Dalton, R.M.; Graae, B.J.; Heiling, J.M.; Opedal, Ø.H. Microenvironment and functional-trait context dependence predict alpine plant community dynamics. J. Ecol. 2018, 106. [Google Scholar] [CrossRef]
- Verde, N.; Kokkoris, I.P.; Georgiadis, C.; Kaimaris, D.; Dimopoulos, P.; Mitsopoulos, I.; Mallinis, G. National scale land cover classification for ecosystem services mapping and assessment, using multitemporal copernicus EO data and google earth engine. Remote Sens. 2020, 12, 3303. [Google Scholar] [CrossRef]
- Screes. Available online: https://eunis.eea.europa.eu/habitats/291 (accessed on 25 February 2021).
- Inland Cliffs, Rock Pavements and Outcrops. Available online: https://eunis.eea.europa.eu/habitats/301 (accessed on 25 February 2021).
- Harrison, S.; Noss, R. Endemism hotspots are linked to stable climatic refugia. Ann. Bot. 2017, 119, 207–214. [Google Scholar] [CrossRef]
- Löffler, J.; Anschlag, K.; Baker, B.; Finch, O.D.; Diekkrüger, B.; Wundram, D.; Schröder, B.; Pape, R.; Lundberg, A. Mountain ecosystem response to global change. Erdkunde 2011, 65. [Google Scholar] [CrossRef] [Green Version]
- Cuesta, F.; Muriel, P.; Llambí, L.D.; Halloy, S.; Aguirre, N.; Beck, S.; Carilla, J.; Meneses, R.I.; Cuello, S.; Grau, A.; et al. Latitudinal and altitudinal patterns of plant community diversity on mountain summits across the tropical Andes. Ecography 2017, 40. [Google Scholar] [CrossRef]
- Körner, C. Alpine ecosystems. In Encyclopedia of Biodiversity, 2nd ed.; Levin, S.A., Ed.; Academic Press: Amsterdam, The Netherlands, 2013; Volume 1, pp. 148–157. [Google Scholar]
- Médail, F.; Verlaque, R. Ecological characteristics and rarity of endemic plants from Southeast France and Corsica: Implications for biodiversity conservation. Biol. Conserv. 1997, 80. [Google Scholar] [CrossRef]
- Strid, A.; Tan, K. Flora Hellenica; Koeltz: Königstein, Germany, 1997; Volume 1, p. 187. [Google Scholar]
- Larson, D.W.; Matthes, U.; Kelly, P. Cliff Ecology: Pattern and Process in Cliff Ecosystems; Cambridge University Press: Cambridge, UK, 2000. [Google Scholar]
- Dimopoulos, P.; Sýkora, K.V.; Mucina, L.; Georgiadis, T. The high-rank syntaxa of the rock-cliff and scree vegetation of the mainland Greece and Crete. Folia Geobot. 1997, 32. [Google Scholar] [CrossRef]
- Mucina, L.; Bültmann, H.; Dierßen, K.; Theurillat, J.P.; Raus, T.; Čarni, A.; Šumberová, K.; Willner, W.; Dengler, J.; García, R.G.; et al. Vegetation of Europe: Hierarchical floristic classification system of vascular plant, bryophyte, lichen, and algal communities. Appl. Veg. Sci. 2016, 19. [Google Scholar] [CrossRef]
- Valachovič, M.; Dierssen, K.; Dimopoulos, P.; Hadač, E.; Loidi, J.; Mucina, L.; Rossi, G.; Tendero, F.V.; Tomaselli, M. The vegetation on screes—A synopsis of higher syntaxa in Europe. Folia Geobot. 1997, 32. [Google Scholar] [CrossRef]
- Davis, P.H. Cliff Vegetation in the Eastern Mediterranean. J. Ecol. 1951, 39. [Google Scholar] [CrossRef]
- Escudero, A. Community patterns on exposed cliffs in a Mediterranean calcareous mountain. Vegetatio 1996, 125. [Google Scholar] [CrossRef]
- Bergmeier, E.; Dimopoulos, P.; Mucina, L. Validation of some alliances of the Aegean chasmophytic vegetation of the Asplenietea trichomanis. Lazaroa 2011, 32. [Google Scholar] [CrossRef]
- Aronne, G.; Arena, C.; De Micco, V.; Giovanetti, M.; Buonanno, M. Full light and soil drought constrain plant growth in Mediterranean cliffs: The case of Primula palinuri Petagna. Plant Biosyst. 2018, 152. [Google Scholar] [CrossRef]
- Fois, M.; Cuena-Lombraña, A.; Fenu, G.; Bacchetta, G. Using species distribution models at local scale to guide the search of poorly known species: Review, methodological issues and future directions. Ecol. Modell. 2018, 385. [Google Scholar] [CrossRef] [Green Version]
- Panitsa, M.; Kontopanou, A. Diversity of chasmophytes in the vascular flora of Greece: Floristic analysis and phytogeographical patterns. Bot. Serbica 2017, 41. [Google Scholar] [CrossRef]
- Trigas, P.; Tsiftsis, S.; Tsiripidis, I.; Iatrou, G. Distribution Patterns and Conservation Perspectives of the Endemic Flora of Peloponnese (Greece). Folia Geobot. 2012, 47. [Google Scholar] [CrossRef]
- EEA Reference Grid. Available online: https://www.eea.europa.eu/data-and-maps/data/eea-reference-grids-2 (accessed on 25 February 2021).
- Kokkoris, I.P.; Kokkinos, V.; Michos, E.; Kalogeropoulos, R.; Charalambides, M.; Kounelis, A.; Iliadou, E.; Damianidis, C.K.; Mallinis, G.; Bouras, C.; et al. MAES_GR: A Web-Based, Spatially Enabled Field Survey Platform for the MAES Implementation in Greece. Land 2021, 10, 381. [Google Scholar] [CrossRef]
- Cooper, A. Plant species coexistence in cliff habitats. J. Biogeogr. 1997, 24. [Google Scholar] [CrossRef]
- Snogerup, S. Evolutionary and plant geographical aspects of chasmophytic communities. In Plant Life of South-West Asia; Davis, P.H., Harper, P.C., Hedge, I.C., Eds.; Botanical Society of Edinburgh: Edinburgh, UK, 1971; pp. 157–170. [Google Scholar]
- Surina, B.; Martinčič, A. Chasmophytes on screes? A rule and not an exception in the vegetation of the Karst (southwest Slovenia). Plant Biosyst. 2012, 146. [Google Scholar] [CrossRef]
- Cadotte, M.W.; Carscadden, K.; Mirotchnick, N. Beyond species: Functional diversity and the maintenance of ecological processes and services. J. Appl. Ecol. 2011, 48, 1079–1087. [Google Scholar] [CrossRef]
- Naeem, S. Disentangling the impacts of diversity on ecosystem functioning in combinatorial experiments. Ecology 2002, 83. [Google Scholar] [CrossRef]
- Biswas, S.R.; Mallik, A.U. Species diversity and functional diversity relationship varies with disturbance intensity. Ecosphere 2011, 2. [Google Scholar] [CrossRef]
- Tomaselli, M.; Foggi, B.; Carbognani, M.; Gennai, M.; Petraglia, A. The rock-face vegetation in the northern apennines and neighbouring mountain areas, from the coast line to the highest summits. Phytocoenologia 2019, 49. [Google Scholar] [CrossRef]
- Lenoir, J.; Svenning, J.C. Climate-related range shifts–a global multidimensional synthesis and new research directions. Ecography 2015, 38. [Google Scholar] [CrossRef]
- Steinbauer, M.J.; Grytnes, J.A.; Jurasinski, G.; Kulonen, A.; Lenoir, J.; Pauli, H.; Rixen, C.; Winkler, M.; Bardy-Durchhalter, M.; Barni, E.; et al. Accelerated increase in plant species richness on mountain summits is linked to warming. Nature 2018, 556. [Google Scholar] [CrossRef]
- Alexander, J.M.; Diez, J.M.; Levine, J.M. Novel competitors shape species’ responses to climate change. Nature 2015, 525. [Google Scholar] [CrossRef]
- Thompson, J.D. Plant Evolution in the Mediterranean; Oxford University Press: Oxford, UK, 2020. [Google Scholar]
- Garnier, E.; Navas, M.-L.; Grigulis, K. Plant Functional Diversity: Organism Traits, Community Structure, and Ecosystem Properties; Oxford University Press: Oxford, UK, 2016. [Google Scholar]
- Cardinale, B.J.; Duffy, J.E.; Gonzalez, A.; Hooper, D.U.; Perrings, C.; Venail, P.; Narwani, A.; MacE, G.M.; Tilman, D.; Wardle, D.A.; et al. Biodiversity loss and its impact on humanity. Nature 2012, 486, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Naeem, S.; Duffy, J.E.; Zavaleta, E. The functions of biological diversity in an age of extinction. Science 2012, 336, 1401–1406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, A.; Keppel, G.; Weigelt, P.; Zotz, G.; Kreft, H. Functional traits are key to understanding orchid diversity on islands. Ecography 2021. [Google Scholar] [CrossRef]
- De Bello, F.; Lavorel, S.; Díaz, S.; Harrington, R.; Cornelissen, J.H.C.; Bardgett, R.D.; Berg, M.P.; Cipriotti, P.; Feld, C.K.; Hering, D.; et al. Towards an assessment of multiple ecosystem processes and services via functional traits. Biodivers. Conserv. 2010, 19. [Google Scholar] [CrossRef]
- Carpenter, S.R.; Mooney, H.A.; Agard, J.; Capistrano, D.; Defries, R.S.; Diaz, S.; Dietz, T.; Duraiappah, A.K.; Oteng-Yeboah, A.; Pereira, H.M.; et al. Science for managing ecosystem services: Beyond the Millennium Ecosystem Assessment. Proc. Natl. Acad. Sci. USA 2009, 106, 1305–1312. [Google Scholar] [CrossRef] [Green Version]
- Lev-Yadun, S. Defensive functions of white coloration in coastal and dune plants. Isr. J. Plant Sci. 2006, 54, 317–325. [Google Scholar] [CrossRef]
- Cheminal, A.; Kokkoris, I.P.; Strid, A.; Dimopoulos, P. Medicinal and aromatic lamiaceae plants in greece: Linking diversity and distribution patterns with ecosystem services. Forests 2020, 11, 661. [Google Scholar] [CrossRef]
- Solomou, A.D.; Martinos, K.; Skoufogianni, E.; Danalatos, N.G. Medicinal and Aromatic Plants Diversity in Greece and Their Future Prospects: A Review. Agric. Sci. 2016, 4, 9–20. [Google Scholar] [CrossRef] [Green Version]
- Petrakou, K.; Iatrou, G.; Lamari, F.N. Ethnopharmacological survey of medicinal plants traded in herbal markets in the Peloponnisos, Greece. J. Herb. Med. 2020, 19. [Google Scholar] [CrossRef]
- Dee, L.E.; Cowles, J.; Isbell, F.; Pau, S.; Gaines, S.D.; Reich, P.B. When Do Ecosystem Services Depend on Rare Species? Trends Ecol. Evol. 2019, 34. [Google Scholar] [CrossRef]
- Gascon, C.; Brooks, T.M.; Contreras-Macbeath, T.; Heard, N.; Konstant, W.; Lamoreux, J.; Launay, F.; Maunder, M.; Mittermeier, R.A.; Molur, S.; et al. The importance and benefits of species. Curr. Biol. 2015, 25, R431–R438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dimopoulos, P.; Drakou, E.G.; Kokkoris, I.P.; Katsanevakis, S.; Kallimanis, A.; Tsiafouli, M.; Bormpoudakis, D.; Kormas, K.; Arends, J. The need for the implementation of an Ecosystem Services assessment in Greece: Drafting the national agenda. One Ecosyst. 2017, 2. [Google Scholar] [CrossRef]
- Maes, J.; Teller, A.; Erhard, M.; Grizzetti, B.; Barredo, J.I.; Paracchini, M.L.; Condé, S.; Somma, F.; Orgiazzi, A.; Jones, A.; et al. Mapping and Assessment of Ecosystems and their Services: An Analytical Framework for Ecosystem Condition; Publications office of the European Union: Luxemburg, 2018; ISBN 978-92-79-74288-0. [Google Scholar]
- Strid, A.; Tan, K. Flora Hellenica. 1; Koeltz Scientific Books: Königstein, Germany, 1997. [Google Scholar]
- Strid, A.; Tan, K. Flora Hellenica. 2.; A. R. G. Gantner: Ruggell, Liechtenstein, 2002. [Google Scholar]
- Kontopanou, A.; Panitsa, M. Habitat islands on the aegean islands (Greece): Elevational gradient of chasmophytic diversity, endemism, phytogeographical patterns and need for monitoring and conservation. Diversity 2020, 12, 33. [Google Scholar] [CrossRef] [Green Version]
- Dimopoulos, P.; Raus, T.; Bergmeier, E.; Constantinidis, T.; Iatrou, G.; Kokkini, S.; Strid, A.; Tzanoudakis, D. Vascular plants of Greece: An annotated checklist. Englera 2013, 1, 372. [Google Scholar]
- Dimopoulos, P.; Raus, T.; Bergmeier, E.; Constantinidis, T.; Iatrou, G.; Kokkini, S.; Strid, A.; Tzanoudakis, D. Vascular plants of Greece: An annotated checklist. Supplement. Willdenowia 2016, 46. [Google Scholar] [CrossRef] [Green Version]
- Dimopoulos, P.; Raus, T.; Strid, A. Flora of Greece Web. Vascular Plants of Greece, an Annotated Checklist. Available online: http://portal.cybertaxonomy.org/flora-greece/ (accessed on 25 February 2021).
- QGIS Development Team QGIS Geographic Information System. Open Source Geospatial Found. Proj. 2020. Available online: http://qgis.osgeo.org (accessed on 25 February 2021).
- Crisp, M.D.; Laffan, S.; Linder, H.P.; Monro, A. Endemism in the Australian flora. J. Biogeogr. 2001, 28, 183–198. [Google Scholar] [CrossRef]
- Linder, H.P. Plant diversity and endemism in sub-Saharan tropical Africa. J. Biogeogr. 2001, 28, 169–182. [Google Scholar] [CrossRef]
- Linder, H.P. On areas of endemism, with an example from the African restionaceae. Syst. Biol. 2001, 50, 892–912. [Google Scholar] [CrossRef]
- Guerin, G.R.; Ruokolainen, L.; Lowe, A.J. A georeferenced implementation of weighted endemism. Methods Ecol. Evol. 2015, 6. [Google Scholar] [CrossRef]
- Guerin, G.R. biomapME: Biodiversity Mapping and Macroecology. R Package v2.0. 2020. Available online: https://github.com/GregGuerin/biomapME (accessed on 25 February 2021).
- Rosauer, D.; Laffan, S.W.; Crisp, M.D.; Donnellan, S.C.; Cook, L.G. Phylogenetic endemism: A new approach for identifying geographical concentrations of evolutionary history. Mol. Ecol. 2009, 18. [Google Scholar] [CrossRef] [PubMed]
- Faith, D.P. Conservation evaluation and phylogenetic diversity. Biol. Conserv. 1992, 61. [Google Scholar] [CrossRef]
- Mazel, F.; Renaud, J.; Guilhaumon, F.; Mouillot, D.; Gravel, D.; Thuiller, W. Mammalian phylogenetic diversity-area. Ecology 2015, 96, 2814–2822. [Google Scholar] [CrossRef] [Green Version]
- Daru, B.H.; Karunarathne, P.; Schliep, K. phyloregion: R package for biogeographical regionalization and macroecology. Methods Ecol. Evol. 2020, 11. [Google Scholar] [CrossRef]
- Daru, B.H.; Elliott, T.L.; Park, D.S.; Davies, T.J. Understanding the Processes Underpinning Patterns of Phylogenetic Regionalization. Trends Ecol. Evol. 2017, 32, 845–860. [Google Scholar] [CrossRef] [PubMed]
- Daru, B.H.; Farooq, H.; Antonelli, A.; Faurby, S. Endemism patterns are scale dependent. Nat. Commun. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Tsirogiannis, C.; Sandel, B. PhyloMeasures: A package for computing phylogenetic biodiversity measures and their statistical moments. Ecography 2016, 39. [Google Scholar] [CrossRef]
- González-Orozco, C.E.; Pollock, L.J.; Thornhill, A.H.; Mishler, B.D.; Knerr, N.; Laffan, S.W.; Miller, J.T.; Rosauer, D.F.; Faith, D.P.; Nipperess, D.A.; et al. Phylogenetic approaches reveal biodiversity threats under climate change. Nat. Clim. Chang. 2016, 6. [Google Scholar] [CrossRef] [Green Version]
- Cañadas, E.M.; Fenu, G.; Peñas, J.; Lorite, J.; Mattana, E.; Bacchetta, G. Hotspots within hotspots: Endemic plant richness, environmental drivers, and implications for conservation. Biol. Conserv. 2014, 170. [Google Scholar] [CrossRef]
- Strid, A. Mountain Flora of Greece. 1; Cambridge University Press: Cambridge, UK, 1986. [Google Scholar]
- Strid, A.; Tan, K. Mountain Flora of Greece; University Press: Edinburgh, UK, 1991; Volume 2. [Google Scholar]
- Tutin, T.G.; Burges, N.A.; Chater, A.O.; Edmondson, J.R.; Heywood, V.H.; Moore, D.M.; Valentine, D.H.; Walters, S.M.; Webb, D.A. Flora Europaea 1, 2nd ed.; Cambridge University Press: Cambridge, UK, 1993. [Google Scholar]
- Tutin, T.G.; Heywood, V.H.; Burges, N.A.; Moore, D.M.; Valentine, D.H.; Walters, S.M.; Webb, D.A. Flora Europaea 2–5; Cambridge University Press: Cambridge, UK, 1976–1981. [Google Scholar]
- Harrison, P.A.; Berry, P.M.; Simpson, G.; Haslett, J.R.; Blicharska, M.; Bucur, M.; Dunford, R.; Egoh, B.; Garcia-Llorente, M.; Geamănă, N.; et al. Linkages between biodiversity attributes and ecosystem services: A systematic review. Ecosyst. Serv. 2014, 9. [Google Scholar] [CrossRef] [Green Version]
- Cresswell, C.J.; Cunningham, H.M.; Wilcox, A.; Randall, N.P. What specific plant traits support ecosystem services such as pollination, bio-control and water quality protection in temperate climates? A systematic map. Environ. Evid. 2018, 7. [Google Scholar] [CrossRef] [Green Version]
- Hanisch, M.; Schweiger, O.; Cord, A.F.; Volk, M.; Knapp, S. Plant functional traits shape multiple ecosystem services, their trade-offs and synergies in grasslands. J. Appl. Ecol. 2020, 57, 1535–1550. [Google Scholar] [CrossRef]
- Hanson, J. Wdpar: Interface to the World Database on Protected Areas; R Package Version; R Core Team: Vienna, Austria, 2019; Volume 1. [Google Scholar]
- Pebesma, E. Simple features for R: Standardized support for spatial vector data. R J. 2018, 10. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Section | Class | Code | Relevant ES Indicators |
|---|---|---|---|
| Provisioning (biotic) | Fibers and other materials from wild plants for direct use or processing (excluding genetic materials) | 1.1.5.2 | G, L |
| Seeds, spores and other plant materials collected for maintaining or establishing a population | 1.2.1.1 | G, L | |
| Higher and lower plants (whole organisms) used to breed new strains or varieties | 1.2.1.2 | T, P | |
| Individual genes extracted from higher and lower plants for the design and construction of new biological entities | 1.2.1.3 | T, P | |
| Regulating and maintenance (biotic) | Pollination (or ‘gamete’ dispersal in a marine context) | 2.2.2.1 | D, C, PL |
| Maintaining nursery populations and habitats (including gene pool protection) | 2.2.2.3 | T, P, G, D | |
| Control of erosion rates | 2.2.1.1 | G, L, LV | |
| Cultural (biotic) | Characteristics of living systems that enable activities promoting health, recuperation, or enjoyment through passive or observational interactions | 3.1.1.2 | T, P, G, L, C, PL |
| Characteristics of living systems that enable scientific investigation or the creation of traditional ecological knowledge | 3.1.2.1 | T, P, PL | |
| Characteristics of living systems that enable education and training | 3.1.2.2 | T, P, G, L, D, C | |
| Characteristics of living systems that enable aesthetic experiences | 3.1.2.4 | G, L, D, C |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Panitsa, M.; Kokkoris, I.P.; Kougioumoutzis, K.; Kontopanou, A.; Bazos, I.; Strid, A.; Dimopoulos, P. Linking Taxonomic, Phylogenetic and Functional Plant Diversity with Ecosystem Services of Cliffs and Screes in Greece. Plants 2021, 10, 992. https://doi.org/10.3390/plants10050992
Panitsa M, Kokkoris IP, Kougioumoutzis K, Kontopanou A, Bazos I, Strid A, Dimopoulos P. Linking Taxonomic, Phylogenetic and Functional Plant Diversity with Ecosystem Services of Cliffs and Screes in Greece. Plants. 2021; 10(5):992. https://doi.org/10.3390/plants10050992
Chicago/Turabian StylePanitsa, Maria, Ioannis P. Kokkoris, Konstantinos Kougioumoutzis, Anna Kontopanou, Ioannis Bazos, Arne Strid, and Panayotis Dimopoulos. 2021. "Linking Taxonomic, Phylogenetic and Functional Plant Diversity with Ecosystem Services of Cliffs and Screes in Greece" Plants 10, no. 5: 992. https://doi.org/10.3390/plants10050992