
Impact of Three Different Light Spectra on the Yield, Morphology and Growth Trajectory of Three Different Cannabis sativa L. Strains

 ,
,  , , ,
, , ,
Abstract
:1. Introduction
2. Results
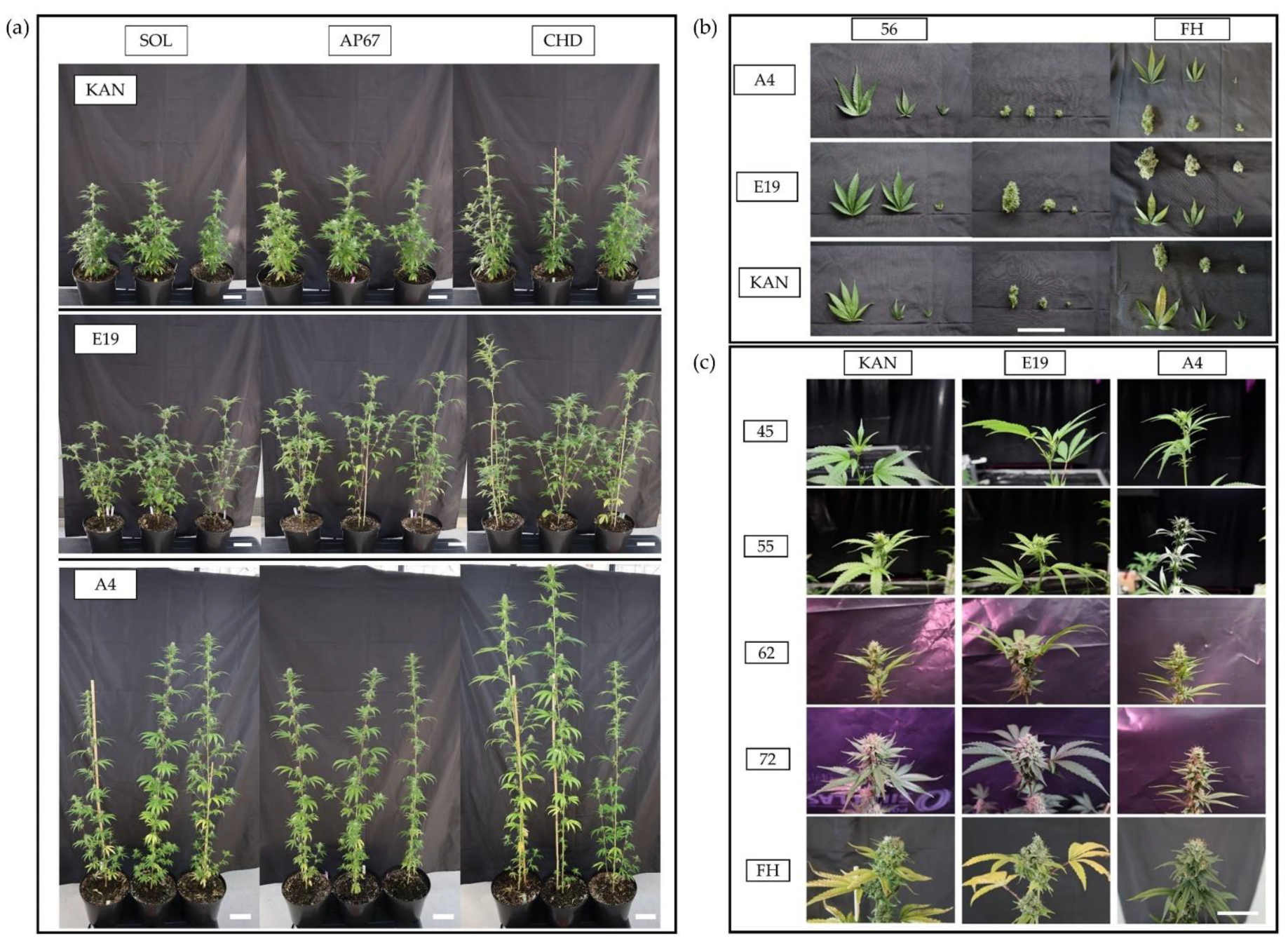
2.1. Plant Architecture
2.1.1. Node Number and Length
2.1.2. Canopy Height
2.1.3. Branch Length and Number
2.1.4. Mean Stem Diameter
2.2. Growth
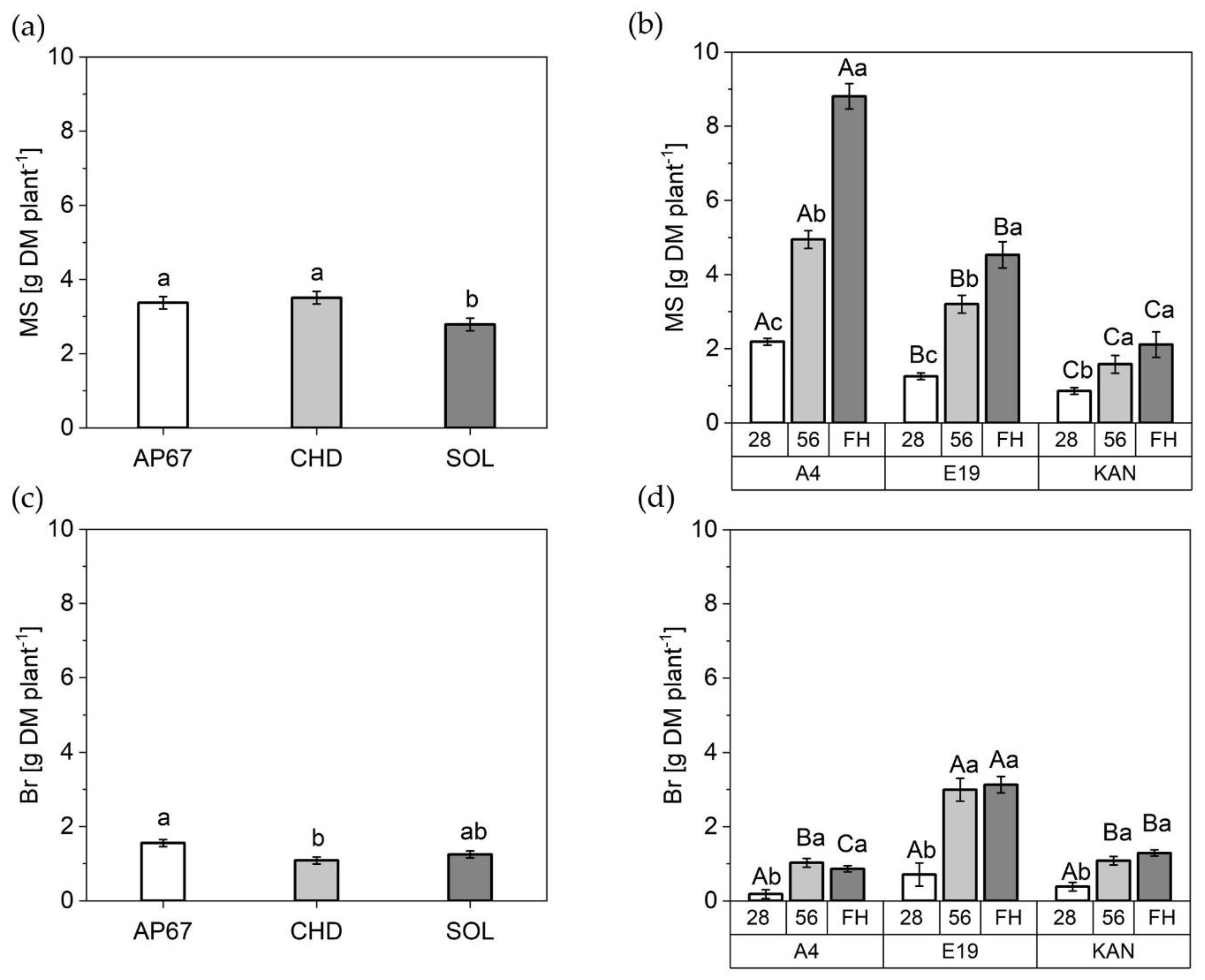
2.2.1. Dry Matter of Leaves
2.2.2. Specific Leaf Area
2.2.3. Stem Dry Matter
2.2.4. Photosynthesis Rate
2.2.5. Nitrogen Concentration of the Leaf Fractions
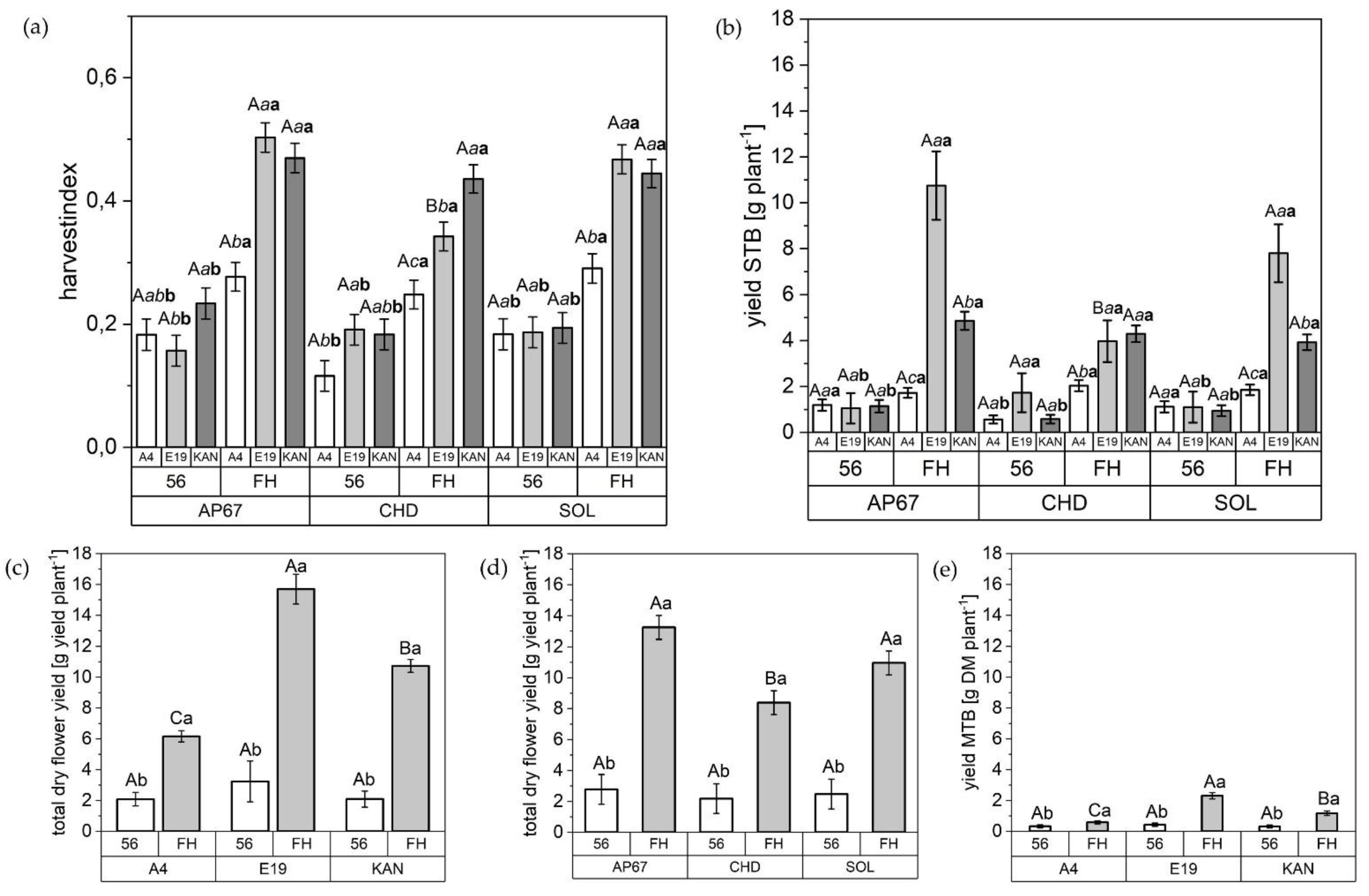
2.3. Harvest
3. Discussion
3.1. Growth Behavior of the Strains
3.2. Assessing the Influence of Light
3.3. Limitations and Practical Application
4. Materials and Methods
4.1. Experimental Setup
4.2. Plant Material and Growing Conditions
4.3. Experimental Light Setting
4.4. Data Collection
4.4.1. Destructive Sampling
4.4.2. Non-Destructive Measurements
4.4.3. Photosynthetic Rate
4.5. Chemical Analysis
4.6. Data Analysis
4.6.1. Calculating of Growing Degree Days
4.6.2. Assessing the Growth Trajectory
4.6.3. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

References
- Russo, E.B. History of Cannabis and Its Preparations in Saga, Science, and Sobriquet. Chem. Biodivers. 2007, 4, 1614–1648. [Google Scholar] [CrossRef]
- Monthony, A.S.; Page, S.R.; Hesami, M.; Jones, A.M.P. The Past, Present and Future of Cannabis sativa Tissue Culture. Plants 2021, 10, 185. [Google Scholar] [CrossRef] [PubMed]
- Zivovinovic, S.; Alder, R.; Allenspach, M.D.; Steuer, C. Determination of cannabinoids in Cannabis sativa L. samples for recreational, medical, and forensic purposes by reversed-phase liquid chromatography-ultraviolet detection. J. Anal. Sci. Technol. 2018, 9, 27. [Google Scholar] [CrossRef] [Green Version]
- Backer, R.; Schwinghamer, T.; Rosenbaum, P.; McCarty, V.; Eichhorn Bilodeau, S.; Lyu, D.; Ahmed, M.B.; Robinson, G.; Lefsrud, M.; Wilkins, O.; et al. Closing the Yield Gap for Cannabis: A Meta-Analysis of Factors Determining Cannabis Yield. Front. Plant Sci. 2019, 10, 495. [Google Scholar] [CrossRef] [Green Version]
- Fischedick, J.T. Identification of Terpenoid Chemotypes Among High (-)-trans-Δ9- Tetrahydrocannabinol-Producing Cannabis sativa L. Cultivars. Cannabis Cannabinoid Res. 2017, 2, 34–47. [Google Scholar] [CrossRef] [Green Version]
- Eichhorn Bilodeau, S.; Wu, B.-S.; Rufyikiri, A.-S.; MacPherson, S.; Lefsrud, M. An Update on Plant Photobiology and Implications for Cannabis Production. Front. Plant Sci. 2019, 10, 296. [Google Scholar] [CrossRef]
- Kakabouki, I.; Mavroeidis, A.; Tataridas, A.; Kousta, A.; Efthimiadou, A.; Karydogianni, S.; Katsenios, N.; ROUSSIS, I.; Papastylianou, P. Effect of Rhizophagus irregularis on Growth and Quality of Cannabis sativa Seedlings. Plants 2021, 10, 1333. [Google Scholar] [CrossRef]
- Burgel, L.; Hartung, J.; Graeff-Hönninger, S. Impact of Different Growing Substrates on Growth, Yield and Cannabinoid Content of Two Cannabis sativa L. Genotypes in a Pot Culture. Horticulturae 2020, 6, 62. [Google Scholar] [CrossRef]
- Smith, J.T.; Jackson, B.E.; Whipker, B.E.; Fonteno, W.C. Industrial hemp vegetative growth affected by substrate composition. Acta Hortic. 2021, 1305, 83–90. [Google Scholar] [CrossRef]
- André, A.; Leupin, M.; Kneubühl, M.; Pedan, V.; Chetschik, I. Evolution of the Polyphenol and Terpene Content, Antioxidant Activity and Plant Morphology of Eight Different Fiber-Type Cultivars of Cannabis Sativa L. Cultivated at Three Sowing Densities. Plants 2020, 9, 1740. [Google Scholar] [CrossRef] [PubMed]
- Burgel, L.; Hartung, J.; Schibano, D.; Graeff-Hönninger, S. Impact of Different Phytohormones on Morphology, Yield and Cannabinoid Content of Cannabis sativa L. Plants 2020, 9, 725. [Google Scholar] [CrossRef]
- Jerushalmi, S.; Maymon, M.; Dombrovsky, A.; Freeman, S. Fungal Pathogens Affecting the Production and Quality of Medical Cannabis in Israel. Plants 2020, 9, 882. [Google Scholar] [CrossRef]
- Booth, J.K.; Bohlmann, J. Terpenes in Cannabis sativa—From plant genome to humans. Plant Sci. 2019, 284, 67–72. [Google Scholar] [CrossRef]
- McPartland, J.M.; Small, E. A classification of endangered high-THC cannabis (Cannabis sativa subsp. indica) domesticates and their wild relatives. PhytoKeys 2020, 144, 81–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mudge, E.M.; Brown, P.N.; Murch, S.J. The Terroir of Cannabis: Terpene Metabolomics as a Tool to Understand Cannabis sativa Selections. Planta Med. 2019, 85, 781–796. [Google Scholar] [CrossRef] [Green Version]
- Sawler, J.; Stout, J.M.; Gardner, K.M.; Hudson, D.; Vidmar, J.; Butler, L.; Page, J.E.; Myles, S. The Genetic Structure of Marijuana and Hemp. PLoS ONE 2015, 10, e0133292. [Google Scholar] [CrossRef] [Green Version]
- Schwabe, A.L.; McGlaughlin, M.E. Genetic tools weed out misconceptions of strain reliability in Cannabis sativa: Implications for a budding industry. J. Cannabis Res. 2019, 1, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Small, E.; Jui, P.Y.; Lefkovitch, L.P. A Numerical Taxonomic Analysis of Cannabis with Special Reference to Species Delimitation. Syst. Bot. 1976, 1, 67. [Google Scholar] [CrossRef]
- Small, E. Evolution and Classification of Cannabis sativa (Marijuana, Hemp) in Relation to Human Utilization. Bot. Rev. 2015, 81, 189–294. [Google Scholar] [CrossRef]
- McPartland, J.M. Cannabis Systematics at the Levels of Family, Genus, and Species. Cannabis Cannabinoid Res. 2018, 3, 203–212. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Chen, X.; Guo, H.; Trindade, L.M.; Salentijn, E.M.J.; Guo, R.; Guo, M.; Xu, Y.; Yang, M. Latitudinal Adaptation and Genetic Insights Into the Origins of Cannabis sativa L. Front. Plant Sci. 2018, 9, 1876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caplan, D.; Stemeroff, J.; Dixon, M.; Zheng, Y. Vegetative propagation of cannabis by stem cuttings: Effects of leaf number, cutting position, rooting hormone, and leaf tip removal. Can. J. Plant Sci. 2018, 98, 1126–1132. [Google Scholar] [CrossRef]
- Caplan, D.; Dixon, M.; Zheng, Y. Optimal Rate of Organic Fertilizer during the Flowering Stage for Cannabis Grown in Two Coir-based Substrates. HortScience 2017, 52, 1796–1803. [Google Scholar] [CrossRef]
- Bernstein, N.; Gorelick, J.; Zerahia, R.; Koch, S. Impact of N, P, K, and Humic Acid Supplementation on the Chemical Profile of Medical Cannabis (Cannabis sativa L). Front. Plant Sci. 2019, 10, 736. [Google Scholar] [CrossRef] [Green Version]
- Chandra, S.; Lata, H.; Khan, I.A.; Elsohly, M.A. Photosynthetic response of Cannabis sativa L. to variations in photosynthetic photon flux densities, temperature and CO2 conditions. Physiol. Mol. Biol. Plants 2008, 14, 299–306. [Google Scholar] [CrossRef] [Green Version]
- Chandra, S.; Lata, H.; Mehmedic, Z.; Khan, I.A.; Elsohly, M.A. Light dependence of photosynthesis and water vapor exchange characteristics in different high Δ9-THC yielding strains of Cannabis sativa L. J. Appl. Res. Med. Aromat. Plants 2015, 2, 39–47. [Google Scholar] [CrossRef]
- Danziger, N.; Bernstein, N. Light matters: Effect of light spectra on cannabinoid profile and plant development of medical cannabis (Cannabis sativa L.). Ind. Crop. Prod. 2021, 164, 113351. [Google Scholar] [CrossRef]
- Cho, L.-H.; Yoon, J.; An, G. The control of flowering time by environmental factors. Plant J. 2017, 90, 708–719. [Google Scholar] [CrossRef]
- Amaducci, S.; Scordia, D.; Liu, F.H.; Zhang, Q.; Guo, H.; Testa, G.; Cosentino, S.L. Key cultivation techniques for hemp in Europe and China. Ind. Crop. Prod. 2015, 68, 2–16. [Google Scholar] [CrossRef]
- Lalge, A.; Cerny, P.; Trojan, V.; and Vyhnanek, T. The effects of red, blue and white light on the growth and development of Cannabis sativa L. Mendel. Net. 2017, 8, 646–651. [Google Scholar]
- Hawley, D.; Graham, T.; Stasiak, M.; Dixon, M. Improving Cannabis Bud Quality and Yield with Subcanopy Lighting. HortScience 2018, 53, 1593–1599. [Google Scholar] [CrossRef]
- Folta, K.M.; Carvalho, S.D. Photoreceptors and Control of Horticultural Plant Traits. HortScience 2015, 50, 1274–1280. [Google Scholar] [CrossRef] [Green Version]
- Bantis, F.; Smirnakou, S.; Ouzounis, T.; Koukounaras, A.; Ntagkas, N.; Radoglou, K. Current status and recent achievements in the field of horticulture with the use of light-emitting diodes (LEDs). Sci. Hortic. 2018, 235, 437–451. [Google Scholar] [CrossRef]
- Mills, E. The carbon footprint of indoor Cannabis production. Energy Policy 2012, 46, 58–67. [Google Scholar] [CrossRef]
- Akvilė, V.; Margit, O.; Pavelas, D. LED Lighting in Horticulture. In Light Emitting Diodes for Agriculture; Springer: Singapore, 2017; pp. 113–147. [Google Scholar]
- Singh, D.; Basu, C.; Meinhardt-Wollweber, M.; Roth, B. LEDs for energy efficient greenhouse lighting. Renew. Sustain. Energy Rev. 2015, 49, 139–147. [Google Scholar] [CrossRef] [Green Version]
- Morrow, R.C. LED Lighting in Horticulture. HortScience 2008, 43, 1947–1950. [Google Scholar] [CrossRef] [Green Version]
- Magagnini, G.; Grassi, G.; Kotiranta, S. The Effect of Light Spectrum on the Morphology and Cannabinoid Content of Cannabis sativa L. MCA 2018, 1, 19–27. [Google Scholar] [CrossRef]
- Namdar, D.; Charuvi, D.; Ajjampura, V.; Mazuz, M.; Ion, A.; Kamara, I.; Koltai, H. LED lighting affects the composition and biological activity of Cannabis sativa secondary metabolites. Ind. Crop. Prod. 2019, 132, 177–185. [Google Scholar] [CrossRef]
- Eaves, J.; Eaves, S.; Morphy, C.; Murray, C. The relationship between light intensity, cannabis yields, and profitability. Agron. J. 2020, 112, 1466–1470. [Google Scholar] [CrossRef]
- Rodriguez-Morrison, V.; Llewellyn, D.; Zheng, Y. Cannabis Yield, Potency, and Leaf Photosynthesis Respond Differently to Increasing Light Levels in an Indoor Environment. Front. Plant Sci. 2021, 12, 646020. [Google Scholar] [CrossRef] [PubMed]
- Wilson, P.J.; Thompson, K.E.N.; Hodgson, J.G. Specific leaf area and leaf dry matter content as alternative predictors of plant strategies. New Phytol. 1999, 143, 155–162. [Google Scholar] [CrossRef]
- Spitzer-Rimon, B.; Duchin, S.; Bernstein, N.; Kamenetsky, R. Architecture and Florogenesis in Female Cannabis sativa Plants. Front. Plant Sci. 2019, 10, 350. [Google Scholar] [CrossRef] [Green Version]
- Bernstein, N.; Gorelick, J.; Koch, S. Interplay between chemistry and morphology in medical cannabis (Cannabis sativa L.). Ind. Crop. Prod. 2019, 129, 185–194. [Google Scholar] [CrossRef]
- Knops, J.M.H.; Reinhart, K. Specific Leaf Area Along a Nitrogen Fertilization Gradient. Am. Midl. Nat. 2000, 144, 265–272. [Google Scholar] [CrossRef]
- Gaudreau, S.; Missihoun, T.; Germain, H. Early topping: An alternative to standard topping increases yield in cannabis production. Plant Sci. Today 2020, 7, 627–630. [Google Scholar] [CrossRef]
- Folina, A.; Kakabouki, I.; Tourkochoriti, E.; Roussis, I.; Pateroulakis, H.; Bilalis, D. Evaluation of the Effect of Topping on Cannabidiol (CBD) Content in Two Industrial Hemp (Cannabis sativa L.) Cultivars. Bull. UASVM Hortic. 2020, 77, 46. [Google Scholar] [CrossRef]
- Naim-Feil, E.; Pembleton, L.W.; Spooner, L.E.; Malthouse, A.L.; Miner, A.; Quinn, M.; Polotnianka, R.M.; Baillie, R.C.; Spangenberg, G.C.; Cogan, N.O.I. The characterization of key physiological traits of medicinal cannabis (Cannabis sativa L.) as a tool for precision breeding. BMC Plant Biol. 2021, 21, 294. [Google Scholar] [CrossRef] [PubMed]
- Chandra, S.; Lata, H.; Elsohly, M.A.; Walker, L.A.; Potter, D. Cannabis cultivation: Methodological issues for obtaining medical-grade product. Epilepsy Behav. 2017, 70, 302–312. [Google Scholar] [CrossRef] [PubMed]
- Meier, U.; Bleiholder, H.; Buhr, L.; Feller, C.; Hack, H.; Heß, M.; Lancashire, P.D.; Schnock, U.; Stauß, R.; van den Boom, T.; et al. The BBCH system to coding the phenological growth stages of plants—History and publications. J. Fur Kult. 2009, 61, 41–52. [Google Scholar] [CrossRef]
- Mediavilla, V.; Jonquera, M.; Schmid-Slembrouck, I.; Soldati, A. Decimal code for growth stages of hemp (Cannabis sativa L.). J. Int. Hemp Assoc. 1998, 5, 68–74. [Google Scholar]
- Glivar, T.; Eržen, J.; Kreft, S.; Zagožen, M.; Čerenak, A.; Čeh, B.; Tavčar Benković, E. Cannabinoid content in industrial hemp (Cannabis sativa L.) strains grown in Slovenia. Ind. Crop. Prod. 2020, 145, 112082. [Google Scholar] [CrossRef]
- Hitz, T.; Hartung, J.; Graeff-Hönninger, S.; Munz, S. Morphological Response of Soybean (Glycine max (L.) Merr.) Cultivars to Light Intensity and Red to Far-Red Ratio. Agronomy 2019, 9, 428. [Google Scholar] [CrossRef] [Green Version]
- Folta, K.M.; Maruhnich, S.A. Green light: A signal to slow down or stop. J. Exp. Bot. 2007, 58, 3099–3111. [Google Scholar] [CrossRef]
- Zhou, X.; GE, Z.M.; Kellomäki, S.; Wang, K.Y.; Peltola, H.; Martikainen, P. Effects of elevated CO2 and temperature on leaf characteristics, photosynthesis and carbon storage in aboveground biomass of a boreal bioenergy crop (Phalaris arundinacea L.) under varying water regimes. GCB Bioenergy 2011, 3, 223–234. [Google Scholar] [CrossRef]
- Reich, P.B.; Ellsworth, D.S.; Walters, M.B. Leaf structure (specific leaf area) modulates photosynthesis-nitrogen relations: Evidence from within and across species and functional groups. Funct. Ecol. 1998, 12, 948–958. [Google Scholar] [CrossRef]
- Casal, J.J. Shade avoidance. Arab. Book 2012, 10, e0157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruberti, I.; Sessa, G.; Ciolfi, A.; Possenti, M.; Carabelli, M.; Morelli, G. Plant adaptation to dynamically changing environment: The shade avoidance response. Biotechnol. Adv. 2012, 30, 1047–1058. [Google Scholar] [CrossRef]
- Tibbitts, T.W.; Morgan, D.C.; Warrington, I.J. Growth of lettuce, spinach, mustard, and wheat plants under four combinations of high-pressure sodium, metal halide, and tungsten halogen lamps at equal PPFD. J. Am. Soc. Hort. Sci. 1983, 108, 622–630. [Google Scholar]
- Wheeler, R.M.; Mackowiak, C.L.; Sager, J.C. Soybean stem growth under high-pressure sodium with supplemental blue lighting. Agron. J. 1991, 83, 903–906. [Google Scholar] [CrossRef]
- Nelson, J.A.; Bugbee, B. Economic analysis of greenhouse lighting: Light emitting diodes vs. high intensity discharge fixtures. PLoS ONE 2014, 9, e99010. [Google Scholar] [CrossRef] [Green Version]
- Ouzounis, T.; Rosenqvist, E.; Ottosen, C.-O. Spectral Effects of Artificial Light on Plant Physiology and Secondary Metabolism: A Review. HortScience 2015, 50, 1128–1135. [Google Scholar] [CrossRef] [Green Version]
- Sellaro, R.; Crepy, M.; Trupkin, S.A.; Karayekov, E.; Buchovsky, A.S.; Rossi, C.; Casal, J.J. Cryptochrome as a sensor of the blue/green ratio of natural radiation in Arabidopsis. Plant Physiol. 2010, 154, 401–409. [Google Scholar] [CrossRef] [Green Version]
- VDLUFA. VDLUFA-Methodenbuch Band II.1 Die Untersuchung von Düngemitteln; VDLUFA-Verl.: Darmstadt, Germany, 2000. [Google Scholar]
- Birch, C.J.; Hammer, G.L.; Rickert, K.G. Temperature and photoperiod sensitivity of development in five cultivars of maize (Zea mays L.) from emergence to tassel initiation. Field Crop. Res. 1998, 55, 93–107. [Google Scholar] [CrossRef] [Green Version]
- Verhulst, P.F. A note on population growth. Corresp. Math. Phys. 1838, 10, 113–121. [Google Scholar]
- Mao, L.; Zhang, L.; Sun, X.; van der Werf, W.; Evers, J.B.; Zhao, X.; Zhang, S.; Song, X.; Li, Z. Use of the beta growth function to quantitatively characterize the effects of plant density and a growth regulator on growth and biomass partitioning in cotton. Field Crop. Res. 2018, 224, 28–36. [Google Scholar] [CrossRef]
- Piepho, H.P.; Buchse, A.; Emrich, K. A Hitchhiker’s Guide to Mixed Models for Randomized Experiments. J. Agron. Crop. Sci. 2003, 189, 310–322. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factor | Height | Number of Nodes | Mean Stem Diameter | |||||
|---|---|---|---|---|---|---|---|---|
| hmax | k | tm | a | b | a | b | ||
| [cm] | [GDD] | |||||||
| Strain | A4 | 136.10 a | 0.0058 a | 410.72 a | 8.54 a | 0.035 a | 3.37 a | 0.0034 a |
| E19 | 95.25 b | 0.0057 a | 347.06 b | 7.75 b | 0.017 b | 3.06 b | 0.0027 b | |
| KAN | 66.11 c | 0.0045 b | 364.65 b | 7.96 b | 0.019 b | 3.06 b | 0.0018 c | |
| Light | AP67 | 94.65 b | 0.0053 | 378.83 | ||||
| CHD | 114.03 a | 0.0053 | 378.83 | |||||
| SOL | 87.78 b | 0.0053 | 378.83 | |||||
| p | Light | 0.001 | 0.367 | 0.737 | 0.993 | 0.264 | 0.117 | 0.164 |
| Strain | 0.001 | 0.001 | 0.001 | 0.017 | 0.001 | 0.009 | 0.001 | |
| Light Strain | 0.203 | 0.580 | 0.408 | 0.113 | 0.645 | 0.421 | 0.409 | |
| Factor | Internode Length | Length Side Branches | No. Branches | |||||
|---|---|---|---|---|---|---|---|---|
| a | b | a | b | a | b | |||
| Light strain | AP67 | A4 | 1.66 Aa | 0.0021 b | 0.37 | 0.017 Ba | −0.88 Ba | 0.044 Aa |
| E19 | 1.74 Aa | 0.0028 a | 0.37 | 0.048 Aa | 1.79 Aa | 0.018 Bb | ||
| KAN | 1.46 Aa | 0.0011 c | 0.37 | 0.020 Ba | 2.93 Aa | 0.022 Ba | ||
| CHD | A4 | 1.27 Ba | 0.0021 b | 0.37 | 0.007 Ba | −2.90 Cb | 0.046 Aa | |
| E19 | 1.85 Aa | 0.0028 a | 0.37 | 0.028 Ab | −1.00 Bb | 0.030 Ba | ||
| KAN | 1.46 ABa | 0.0011 c | 0.37 | 0.024 ABa | 1.72 Aa | 0.025 Ba | ||
| SOL | A4 | 1.65 Aa | 0.0021 b | 0.37 | 0.008 Ba | 0.10 Ba | 0.043 Aa | |
| E19 | 1.59 ABa | 0.0028 a | 0.37 | 0.043 Aa | 2.38 Aa | 0.017 Bb | ||
| KAN | 1.39 Ba | 0.0011 c | 0.37 | 0.020 Ba | 2.28 Aa | 0.023 Ba | ||
| p | Light | 0.799 | 0.277 | 0.436 | 0.169 | 0.010 | 0.028 | |
| Strain | 0.001 | 0.001 | 0.433 | 0.001 | 0.001 | 0.001 | ||
| Light Strain | 0.014 | 0.101 | 0.629 | 0.019 | 0.016 | 0.041 | ||
| Trait | Light Spectra/Strain | Yield in g DM/Plant |
|---|---|---|
| Side bud (SB) light | AP67 | 3.36 ± 0.23 a |
| CHD | 2.33 ± 0.24 b | |
| SOL | 2.91 ± 0.23 ab | |
| Side bud (SB) strain | A4 | 2.33 ± 0.15 b |
| E19 | 3.37 ± 0.30 a | |
| KAN | 2.90 ± 0.12 a | |
| Side bud (SB) date | 56 | 0.92 ± 0.20 b |
| FH | 4.82 ± 0.15 a | |
| p-values side Bud | ||
| Rep | <0.0001 | |
| Light | 0.0166 | |
| Strain | 0.0013 | |
| Date | <0.0001 | |
| Long Day Period (18/6) | Short Day Period (12/12) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Nutrients in mg | Week 1 (1–7) | Week 2 (8–14) | Week 3 (15–21) | Week 4 (22–28) | Week 1 (29–35) | Week 2 (36–42) | Week 3 (43–49) | Week 4 (50–56) | Week 5 (57–63) | Week 6 (64–70) | Week 7 (71–77) | Week 8 (78–85) |
| N | 8.1 | 16.2 | 32.4 | 32.4 | / | 310 | 60 | 80 | 60 | 40 | 20 | / |
| P205 | 5.4 | 10.8 | 21.6 | 21.6 | / | 80 | 120 | 160 | 120 | 80 | 40 | / |
| K2O | 8.1 | 16.2 | 32.4 | 32.4 | / | 120 | 180 | 240 | 180 | 120 | 60 | / |
| Spectral Range (nm) | AP67 | CHD | SOL |
|---|---|---|---|
| PAR 400–700 | 680 | 680 | 680 |
| 300–400 | 2.31 | 11.00 | 8.04 |
| 400–500 | 104.50 | 111.00 | 121.60 |
| 500–600 | 137.50 | 264.00 | 251.49 |
| 600–700 | 438.00 | 305.00 | 308.04 |
| 700–800 | 122.76 | 140.00 | 34.31 |
| R:FR | 4.04 | 2.83 | 13.49 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reichel, P.; Munz, S.; Hartung, J.; Präger, A.; Kotiranta, S.; Burgel, L.; Schober, T.; Graeff-Hönninger, S. Impact of Three Different Light Spectra on the Yield, Morphology and Growth Trajectory of Three Different Cannabis sativa L. Strains. Plants 2021, 10, 1866. https://doi.org/10.3390/plants10091866
Reichel P, Munz S, Hartung J, Präger A, Kotiranta S, Burgel L, Schober T, Graeff-Hönninger S. Impact of Three Different Light Spectra on the Yield, Morphology and Growth Trajectory of Three Different Cannabis sativa L. Strains. Plants. 2021; 10(9):1866. https://doi.org/10.3390/plants10091866
Chicago/Turabian StyleReichel, Philipp, Sebastian Munz, Jens Hartung, Achim Präger, Stiina Kotiranta, Lisa Burgel, Torsten Schober, and Simone Graeff-Hönninger. 2021. "Impact of Three Different Light Spectra on the Yield, Morphology and Growth Trajectory of Three Different Cannabis sativa L. Strains" Plants 10, no. 9: 1866. https://doi.org/10.3390/plants10091866