Reducing Cadmium Accumulation in Plants: Structure–Function Relations and Tissue-Specific Operation of Transporters in the Spotlight

1
International Research Center for Environmental Membrane Biology, Foshan University, Foshan 528000, China
2
Tasmanian Institute of Agriculture, University of Tasmania, Hobart TAS 7001, Australia
*
Author to whom correspondence should be addressed.
Plants 2020, 9(2), 223; https://doi.org/10.3390/plants9020223
Submission received: 31 January 2020
/
Revised: 5 February 2020
/
Accepted: 6 February 2020
/
Published: 9 February 2020
(This article belongs to the Special Issue Root System Structure and Function: A Themed Issue in Honor of Professor Yoav Waisel)
Abstract
:Cadmium (Cd) is present in many soils and, when entering the food chain, represents a major health threat to humans. Reducing Cd accumulation in plants is complicated by the fact that most known Cd transporters also operate in the transport of essential nutrients such as Zn, Fe, Mn, or Cu. This work summarizes the current knowledge of mechanisms mediating Cd uptake, radial transport, and translocation within the plant. It is concluded that real progress in the field may be only achieved if the transport of Cd and the above beneficial micronutrients is uncoupled, and we discuss the possible ways of achieving this goal. Accordingly, we suggest that the major focus of research in the field should be on the structure–function relations of various transporter isoforms and the functional assessment of their tissue-specific operation. Of specific importance are two tissues. The first one is a xylem parenchyma in plant roots; a major “controller” of Cd loading into the xylem and its transport to the shoot. The second one is a phloem tissue that operates in the last step of a metal transport. Another promising and currently underexplored avenue is to understand the role of non-selective cation channels in Cd uptake and reveal mechanisms of their regulation.
1. Cd Toxicity as an Issue
1.1. Cadmium in Soils
Cadmium (Cd), a non-essential metal element for people and plants, is widely present in soils in Europe, China, Japan, and America [1]. The origin of cadmium can be divided into two types; one is from natural sources, the other one is of anthropogenic origin [2]. While cadmium is also present in the air and water, most of it eventually goes to the soil. Mineral oil and heavy metals are the main pollutants causing soil contamination, accounting for about 60%. In nature, Cd is released into the environment through rivers and the atmosphere, mainly from weathering of soil parent material and volcanic activities, at an estimated rate of about 820 metric tons per year [3,4]. In contrast, anthropogenic Cd emissions are more serious, accounting for 8000 to 10,000 mt per year [5]. The major sources of anthropogenic Cd are from cadmium-related manufacturing, application of chemical fertilizers, contaminated sewage sludge and waste water, sewage effluents, and agricultural run-offs [6]. Cadmium migrates into the surrounding soil and atmosphere through solid waste combustion. Compared with urban soils, the content of cadmium in agricultural soil is relatively low [7]. However, due to the intensive use of phosphate fertilizer or leakage of factory sewage, Cd accumulation in agricultural soils is becoming a major issue. Cadmium is of a particular concern for plants, because it accumulates in leaves in high quantities, which can be 10–500 times higher than in plants grown in non-polluted environments, and may then enter the food chain by being eaten by animals or humans [8]. The ability of plants to absorb cadmium mainly depends on the concentration of soil Cd and plant ability of accumulation and translocation, and is also affected by pH, temperature, and redox potential, as well as by concentrations of other elements and soil organic matter [9]. For example, excessive use of nitrogen and phosphate fertilizer in many areas of China led to increased soil acidification, while acidic soil accelerates the absorption of cadmium by plants [10]. Drought stress may also increase the rate of cadmium uptake 2–3-fold, compared with non-stressed conditions [11,12]. As described in Section 3.1, Cd uptake by plant roots shares the same pathway with zinc (Zn). Zn is an essential micronutrient and usually accompanies Cd with a ratio of 100:1 in most natural systems and anthropogenic environments [13,14,15]. Due to their similar geochemistry, Cd and Zn interactions are often of a synergistic nature. However, Cd toxicity is almost 10 times higher than that of Zn [16,17], and Cd is not required for plant growth and development, while Zn is considered essential for many metabolic functions.
1.2. Cadmium Toxicity: Health Implications
Most studies have shown that excessive cadmium accumulation leads to kidney dysfunction and lung damage. Cadmium is also a carcinogen that can cause kidney cancer and breast canceranemia. Other negative health effects include heart failure, hypertension, cerebral infarction, proteinuria, eye cataract formation, osteoporosis, emphysema, and renal insufficiency [18,19]. For example, the average weekly cadmium accumulation in Japan during the 1980s to 2000s was up to 4 mg Cd per kilogram of body weight [20]; this has resulted in an outbreak of the Itai-Itai disease caused by Cd contamination in rice in the river basin of Toyama [6,21]. In a study on rats, a high level of lipid peroxidation was observed under the continued oral application of 3.5 mg∙kg−1 of Cd, resulting in a high MDA level in liver, kidney, and serum [22]. Acute Cd treatments at higher doses resulted in a massive and rapid hepatic necrosis [23]. Cadmium accumulates in the human body mainly through the food chain, and most food comes from edible parts or seeds of crops [24]. An estimated 98% of cadmium intake comes from terrestrial foods, only 1% from aquatic foods, and 1% from cadmium in a drinking water [25]. Plant foods are generally considered to be the main source of Cd exposure in the population, and grains account for a large proportion of the total dietary intake. Wheat and rice are the major food crops for the global population [26,27]. Both species can easily absorb cadmium from the soil and accumulate it in grains [28,29]. In China, the average intake of cadmium for the general population more than doubled between 1990 and 2015 [30,31]. Of a specific concern is also Cd accumulation in leafy vegetables. Lettuce (Lactuca sativa L.) and endive (Cichorium endivia L.) are two important salad leafy vegetables; both may accumulate significant amounts of Cd in edible parts that can then enter the food chain. In these species, some studies have reported Cd contents of up to 9 mg∙kg−1 in their leaves, fourfold higher than that in roots and 20–30-fold higher than the actual Cd content in the soil (Table 1) [32,33]. Thus, preventing excessive accumulation of Cd in grains and leafy vegetables is critical for population health [34].
1.3. Cadmium Impact on Plant Growth and Performance
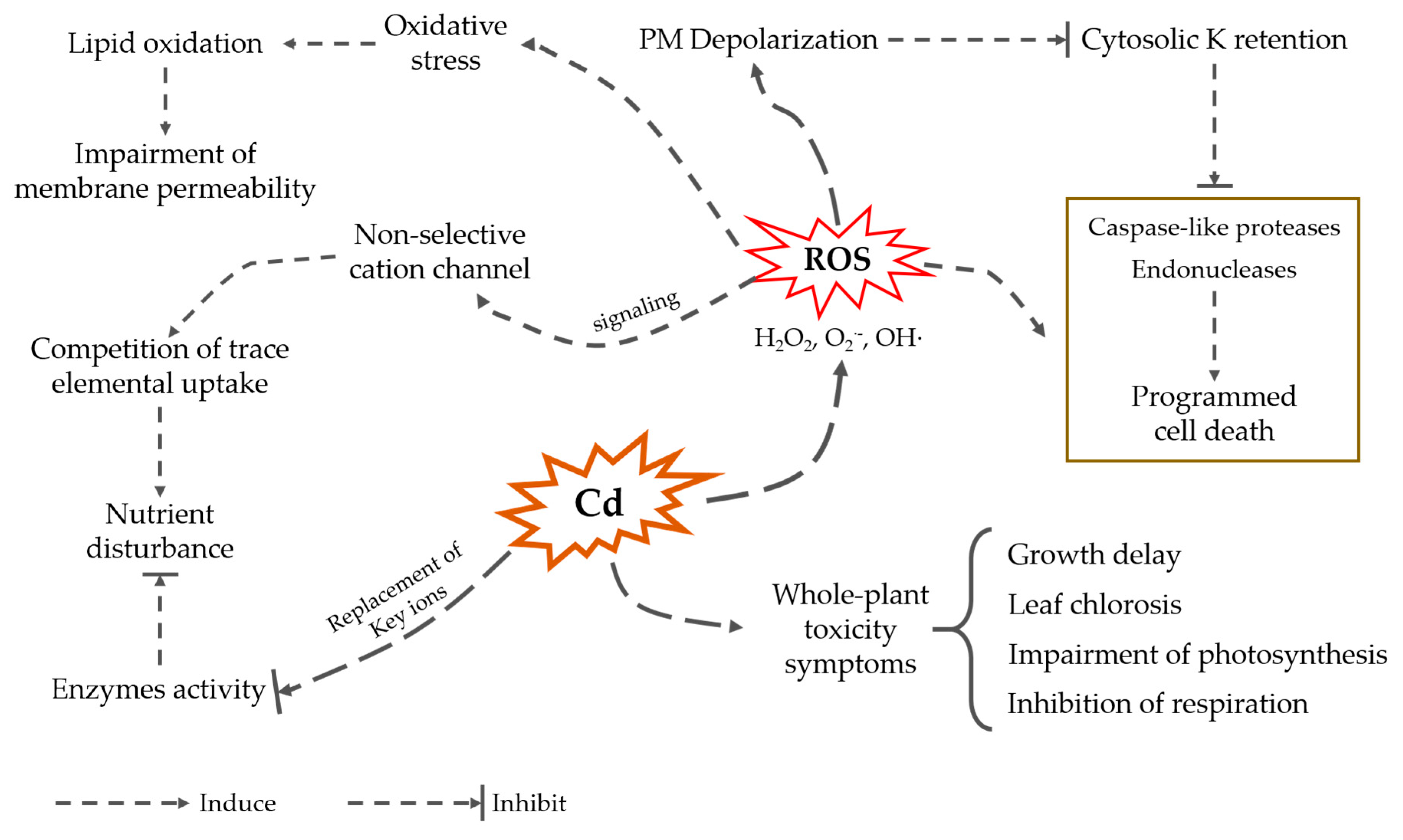
Cadmium causes a wide range of deleterious effects on plants, affecting plant metabolism and causing oxidative stress, nutrient uptake disturbance, and even plant death (Figure 1; [35]). At the whole-plant level, symptoms of Cd toxicity include growth delay, leaf chlorosis, and inhibition of photosynthesis and respiration [36]. Cd can impair plant development and growth by interfering with biochemical and physiologically related signaling pathways, such as affecting photosynthesis by interfering with photosynthetic electron transport, resulting in decreased chlorophyll content and stomatal conductance [37]. Moreover, the inhibition of root Fe(III) reductase induced by Cd led to Fe(II) deficiency, and it seriously affected photosynthesis [38]. Cadmium also changes membrane permeability, leading to a decrease in water content and affecting water balance [39].
Cadmium is a non-essential element; thus, plants have not developed a specialized uptake system for its absorption. Instead, Cd enters plants and is transported across various membranes by other metal transporters [40,41,42,43] or non-selective cation channels [44,45]. This results in a competition with acquisition of some essential metals. In addition, cadmium can also replace ions on the active sites of some enzymes, thus disrupting their activity and absorption of essential nutrients [46].
Cadmium also induces depolarization of the root cell plasma membrane, thus reducing the driving force for cation uptake (e.g., potassium). Potassium plays a key role in determining the fate of cells. In both mammalian [47] and plant [48,49] systems, high cytosolic K+ concentrations are required to suppress the activity of caspase-like proteases and endonucleases and, hence, prevent programmed cell death (PCD). Therefore, it could be envisaged that when cadmium ions affect the absorption and efflux of potassium, the imbalance of potassium ions in the cytoplasm will be caused, thus activating the cell PCD process.
The toxicity threshold for Cd in plants varies between plant species. Cd concentration in grains has been found to increase proportionally to the total Cd content in soils, and 0.3 and 0.1 mg∙kg−1 levels are considered as a likely threshold for barley and wheat, respectively [14]. Necrosis symptoms can be observed in tomato when grown in half-strength Hoagland solution containing 10 μM Cd for two weeks [50]. At the same time, in some hyperaccumulator species concentrations of Cd in some tissues may exceed these values by several orders of magnitude (e.g., 15,000 mg∙kg−1 in the pith tissue in Sedum alfredii; [51]).
2. Current Trends in Breeding Programs
Given the importance of Cd toxicity issue for human health, the ideal solution for this problem would be via remediation of contaminated soils and preventing further anthropogenic-driven Cd contamination. However, achieving this goal may require a significant amount of time and orchestrated efforts and therefore can be considered as strategic only. An immediate solution may come through biological/genetic means, by creating cultivars with minimal, or no, Cd accumulation. Numerous attempts have been made to understand the genetic basis of Cd transport and accumulation in plants, in order to reduce Cd load. Given that over 50% of all calories consumed in the human diet comes from three major cereal crops (rice, wheat, and maize), reduction of Cd allocation to cereal grain is considered as the most essential objective in the breeding programs. This implies understanding the modes controlling Cd transfer from the roots to the shoots, followed by their modification by genetic means. Rice (Oryza sativa L.) is the main food feeding more than half of the global population; thus, the majority of research was done on this cereal. During the last decade, a series of QTL (quantitative trait loci) associated with Cd transportation in rice has been reported. Here, six different QTLs were found to be associated with Cd concentration in shoots during the seedling stage [52,53,54,55]. Genetic and physiological investigations suggest that the major QTL detected on the long arm of chromosome 11 is responsible for the specific translocation of Cd from the roots to the shoots [52]. Chromosome 7 harbors several putative metal transporters encoding genes such as OsZIP8, OsHMA3, and OsNramp1 [42,56]. Three major QTL (qCCBR1-1, qCCBR4-2, and qCCBR9-1) associated with Cd concentration in brown rice and another one named qCCMR11-2 in the milled rice explained more than 20% of the phenotypic variance respectively. These findings may potentially facilitate marker-assisted selection of rice varieties with low Cd accumulation in grain. It was shown that OsCDT1, OsCDT3, and OsCDT4 located on chromosome 6, 1, and 2, respectively, encode a Cys-rich peptide in rice. This peptide chelates Cd at the cellular surface and prevents its further trans-membrane transportation, resulting in less cytosolic accumulation of Cd [57]. Further investigation demonstrated a differential binding affinity responding to diverse metal among CDTs [58], suggesting a possible genetic solution for modification of selectivity for these toxic metals. Recently, a new QTL, qGCd7.1, associated with high Cd accumulation was detected as a new OsHMA3 allele [59]. However, this new allele was weak at both transcriptional and protein levels in the test genotypes compared with the fully functional OsHMA3 lines. Liu et al. [60] also reported that a QTL for grain Cd concentration located on chromosome 7 (GCC7), which is responsible for differential shoot and grain Cd accumulation, had a different promoter activity of OsHMA3 regulating the expression level in shoot and grain. Together, it is plausible that GCC7 has a capacity to interact with qGCd7.1 regulating the Cd uptake. Using genome-wide association studies, Liu et al. [61] reported 17 QTLs associated with grain Cd concentration. Among them, a novel candidate gene encoding OsARM1, a MYB transcription factor, previously known as responsible for freezing tolerance [62,63,64], was predicted to respond to Cd stress. Overexpression of the Cd-induced MYB49 gene in Arabidopsis resulted in a significant increase in Cd accumulation [65]. MYB49 was also interacting with ABI5 which was up-regulated by Cd-induced ABA accumulation, implicating ABA signaling in control of Cd uptake and accumulation in plants [65]. In addition, one significant QTL referring to seed cadmium concentration was identified and validated in the F6:7 NILs in soybean [66]. This major QTL located on chromosome 9 was revealed to account for more than 50% of the genetic variation in RILs populations and contributed to a phenotype with low seed Cd concentration. Further, an effect of the Cda1 locus on seed Cd concentration was confirmed by utilization of marker-selected soybean genotypes with a significant separation between high and low Cd accumulation [67]. However, despite these findings, no major progress in developing cultivars with low Cd-accumulating ability was achieved. The reasons for this are twofold. First, with so many QTLs reported, it is not practical to implement Cd tolerance traits without transferring some other undesirable genes. Second, at the physiological level, Cd transporters also operate as transporters of essential micronutrients such as Fe, Zn, or Mn. Thus, attempts to reduce Cd accumulations in plants may compromise their capacity for taking up these beneficial nutrients.
3. Molecular Mechanisms of Cd Uptake and Transport
3.1. Cd Uptake by Roots
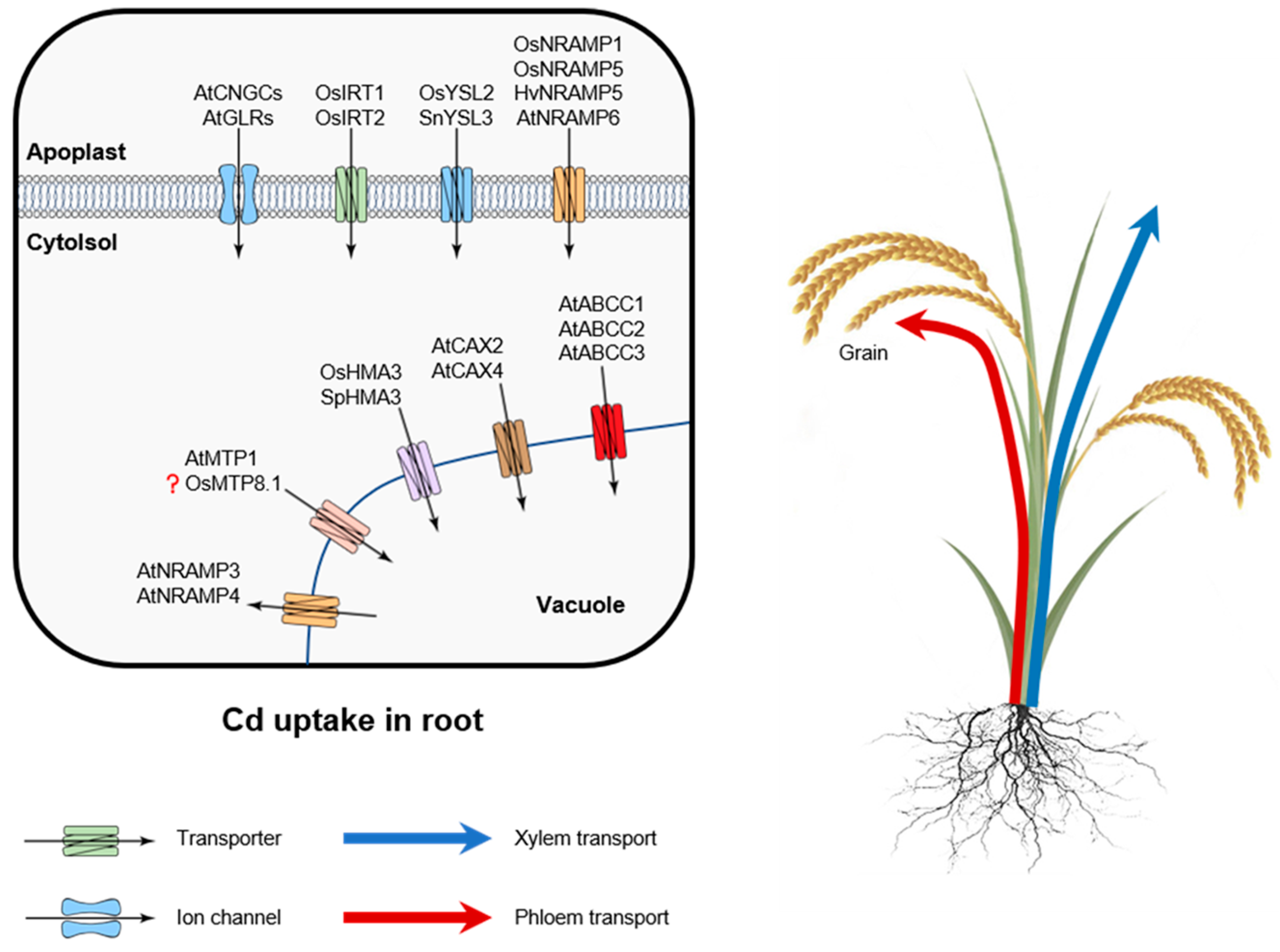
Plant cells have no Cd-selective transporter; hence, Cd uptake occurs through plasma membrane transporters involved in the uptake of other divalent cations, including Ca, Mg, Fe, Zn, Mn, and Cu [68]. Most of studies in the field were conducted on rice [69,70,71]. Several families of transporters have been implied to be responsible for Cd accumulation (Figure 2).
When present in ionic form, Cd uptake by plant roots could be mediated by three major transport systems [40,72,73,74]. One of them is NRAMP (natural resistance-associated macrophage protein). The best-known family members are OsNRAMP1, OsNRAMP5, and AtNRAMP6. Cd can be also taken via ZIP (zinc/iron-regulated transporter-like protein) transporters, such as AtIRT1 and TcZNT1/TcZIP4, and finally, by low-affinity calcium transporters (such as TaLCT1 in wheat). Cadmium can be also transported by transport systems mediating plant Fe uptake. Two distinct strategies are used by plants to accumulate Fe from soils. The first one relies on the uptake of a reduced form of Fe(II) (so-called Strategy I). Strategy II is only observed in grasses and implies uptake of chelated Fe(III). Here, Fe(III)-phytosiderophore complexes (Fe(III)-PS) are taken into the root cells by yellow stripe 1 or yellow stripe-like 1 (YS1/YSL1) transporters identified in maize [75] and barley [76]. In chelated form, Cd may also be transported by YSL (yellow stripe-like 1) proteins such as OsYSL2 in rice and SnYSL3 in Solanum nigrum [41,77]. Plants belonging to Strategy I have the ability to transport Cd across plasma membrane by the same mechanisms employed for Fe2+ transport. OsIRT1 was shown to be implicated in Cd influx, for example, upon reaeration of soil after flooding [42,78]. In Arabidopsis, the rate of root Cd uptake was much lower in atirt1 knockout mutant [79]. Ogawa et al. [78] showed that OsIRT1 and OsIRT2, two rice Fe2+ transporters, were also able to transport Cd when expressed in yeast. However, the role of these transporters for Cd uptake in planta seems to be rather minor in rice [73].
Of all the transporters above, NRAMP members have been demonstrated to be involved in numerous functions including uptake, translocation, intracellular transport, and detoxification of transition metals in many species [41,71,79,80,81,82,83]. In rice, OsNRAMP5, which plays a major role in Mn2+ uptake, may also represent a major route of Cd uptake by roots [73,82]. However, this view was challenged by Takahashi et al. [84] who showed rice mutants lacking functional OsNRAMP5 only showed a ~20% reduction in root Cd content, while accumulating significant amounts of Cd in the shoot. In barley, HvNRAMP5, which shares 84% identity with OsNRAMP5, can also mediate the uptake of Cd and Mn [83]. Interestingly, other orthologues of NRAMPs perform different specificity to NRAMP5. Among of them, OsNRAMP3 only has a capacity to transport Mn rather than Cd or Fe [85], while OsNRAMP1 can mediate uptake of trivalent Al ion [86]. In Arabidopsis, AtNRAMP3 and AtNRAMP4 can be functional to transport Cd2+ [87,88]. AtNRAMP6 is also involved in intracellular Cd2+ transport [40]. The above results indicate that plants can transport cadmium through these low affinity metal transporters. However, the genetic variation affecting the selectivity of the transport for specific metals needs to be further investigated.
The summary of reported Cd transporters, their tissue-specific localization, and substrate specificity are given in the table below (Table 2).
In the roots of a halophytic plant Suaeda salsa, Cd2+ influx was inhibited by Ca2+ uptake blockers. Higher plants lack Ca2+-selective channels [90], and Ca2+ uptake in plants is believed to occur mainly via non-selective cation (NSCC) channels [89]. Thus, these results suggest that some Cd2+ can be transported into cells through NSCC. The molecular nature and mechanisms of regulation of Cd uptake via NSCC remain elusive. Altogether, Arabidopsis genome harbors 40 NSCC channels from two major groups; 20 of these channels are classified as “cyclic nucleotide-gated channels” (CNGC), and the other 20 as “glutamate receptors” [90]. The specific roles each of these members in Cd uptake require investigation.
3.2. Long-Distant Cd Transport
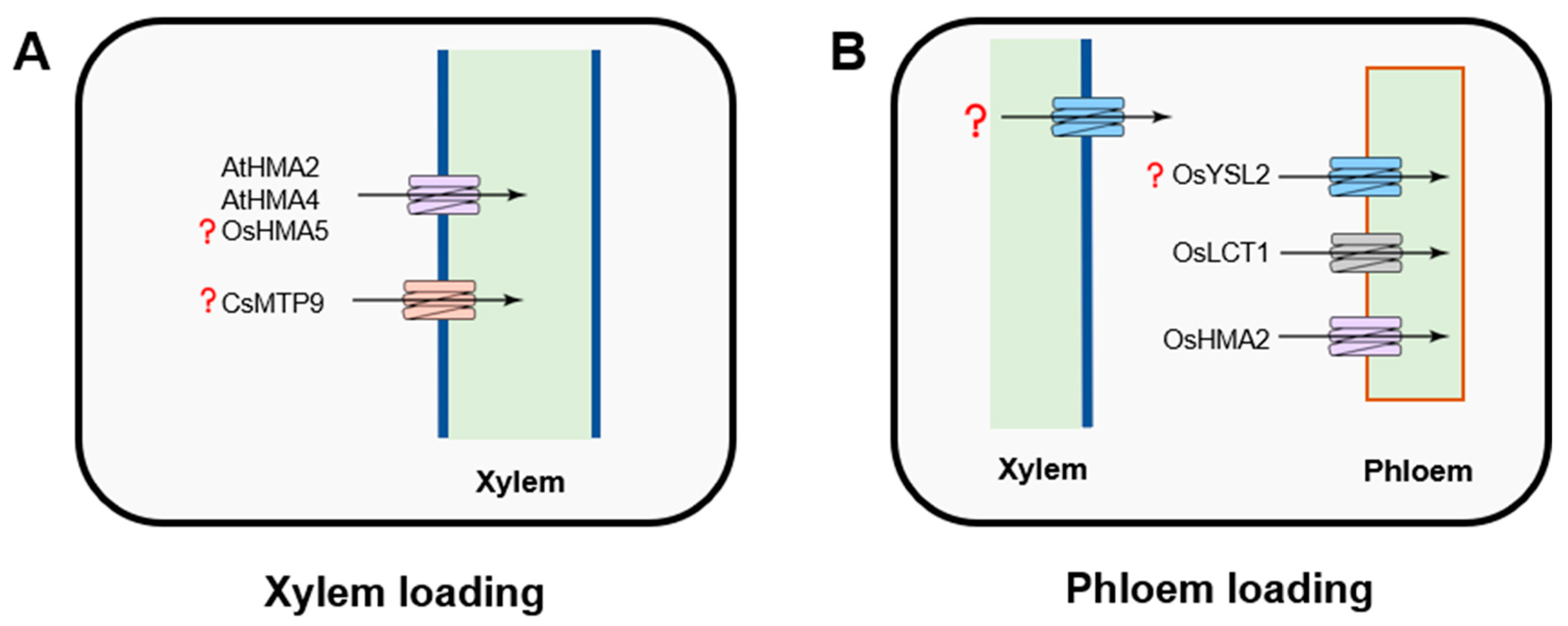
Once taken by the root, Cd is then loaded into the xylem to be transported to the shoot. Cd translocation via the xylem is a key determinant of variation in grain Cd accumulation. The differences in the root-to-shoot Cd translocation rates of cultivars of O. sativa ssp. indica (high Cd-accumulating) and O. sativa ssp. japonica (low Cd-accumulating) rice explain the genotypic variation in Cd-accumulation observed in these two subspecies [26]. This translocation is mostly a function of retention in roots and xylem loading activity. The cation diffusion facilitator (CDF) family, otherwise known as the metal tolerance protein (MTP) family in plants, is believed to play an essential role in this process (Figure 3A). The MTPs/CDFs possess an ability to transport multiple ions and, depending on their selectivity, the members of MTP family are classified into three major clusters, including Zn-CDF, Mn-CDF, and Zn/Fe-CDF [97,98,99,100,117,118].
Functionally, MTP members operate as efflux transporters. It was shown that CsMTP9, known as an efflux transporter for Mn, rescued the Cd-hypersensitive phenotype when expressed in yeast, resulting from strengthened Cd2+ efflux activity [101]. However, OsMTP9 does not have a capacity to transport Cd, suggesting a different unidentified transporter involved in this pathway [102]. In rice, OsMTP8.1 has been described as a vacuolar Mn-specific transporter, but its role in Cd uptake has not been evaluated [103]. In Arabidopsis, MTP1 has been described as a Zn2+/H+ vacuolar transporter, which also showed a capacity to mediate Cd flux in Thlaspi goesingense [99,104]. Podar et al. [119] also reported that MTP1 is a Zn2+/Co2+-specific transporter, with a high affinity to Zn2+ when expressed in barley. Importantly, the structure–function analysis of MTP1 suggests that it is possible to increase the selectivity of AtMTP1 towards Zn by modifying a five-residue sequence within the MTP1 N-segment of the His-rich intracytoplasmic loop. This His-rich loop was suggested to be a determinant of the identity of the metal ion [100].
P1B-type heavy metal-transporting ATPases (HMAs) transport heavy metals (Cu+, Cu2+, Zn2+, Co2+, Cd2+, Pb2+) across membranes are present in most organisms and crucial for the cellular metal homeostasis. Most of them are not fully characterized. OsHMA5 has also been localized to the root pericycle cells and xylem but seems to have more affinity to copper rather than other metal ions [105]. In Arabidopsis, HMA2 and HMA4 are considered to load Cd and Zn into the xylem [106,107]. The latest study on the identification of mutations from amino acids of HMA4 transmembrane helices (TMs) indicated a potential solution to alter the function of this transporter in Arabidopsis hma2/hma4 double mutant. The transporter with substitution of E169 and K667 by Ala residues can still transport Cd2+ as the native protein, while Zn2+ transport is completely silenced [107].
Although the long-distant transport via phloem is crucial for Cd accumulation in seeds and grains, the mechanistic basis of this process is poorly understood (Figure 3B). So far, OsHMA2 has been described to function in intervascular Cd delivery to developing tissues in the uppermost node of rice [85]. The loss of OsHMA2 function in insertion mutants results in decreased leaf and grain Cd concentrations [120]. An increase in Cd tolerance in rice and reduction of Cd accumulation in grains can be observed in the overexpression lines of OsHMA3 [54,121]. Another transporter that mediates phloem-based Cd distribution is LCT1 (low-affinity cation transporter 1) [74,108]. OsLCT1 encodes a plasma membrane-localized protein, which is mainly expressed in phloem parenchyma cells in leaf blades and nodes during the reproductive stage [108]. OsLCT1 transcript level is strongly upregulated during the reproductive stage in rice, and a decrease in Cd concentration in grains can be observed in its oslct1 knockdown line [74]. OsYSL2, an orthologue of ZmYSL1, is mainly expressed in parenchyma, suggesting a potential capacity to transport Cd in phloem [77]. At this stage, given the limited knowledge on molecular mechanisms of phloem loading, LCT1 and HMA2 are the likely candidate genes for further modification to restrain the translocation of Cd, particularly into grain in cereal crops.
4. Tissue Tolerance Mechanisms
To minimize the toxicity of Cd exposure and its accumulation, plants have evolved various detoxification mechanisms to surmount the adverse effects. Sequestration in root vacuoles has been demonstrated as the major process limiting the translocation of Cd to shoots and seeds [54,122]. As the largest organelle inside the mature plant cell, vacuole is an important reservoir of ions and metabolites and is crucial for the detoxification process as well as normal cell development [123,124].
Cd has a strong affinity for thiol-containing molecules such as cysteine, glutathione, and phytochelatins (PCs). Phytochelatins react with heavy metal ions by glutathione S-transferase catalyzation in the cytosol, and afterwards, they are sequestered into the vacuole for degradation. In sequestration into vacuoles, ATP-binding cassette transporters mediate influx of PC-metal (loid) complexes. In Schizosaccharomyces pombe, the first ABC transporter HMT1 located at the tonoplast was reported to aid transporting PC-Cd complexes formed in the cytosol [125,126,127]. A functional homolog of HMT1 has been then reported in Caenorhabditis elegans and Drosophila [128,129]. In Arabidopsis, ABCC1 and ABCC2 have been identified as the main transporters mediating PC uptake into vacuoles [109,130]. Cd staining experiments showed that Cd was mainly present in the cytosol in abcc1abcc2 double mutant, whereas almost all Cd has been compartmentalized in the vacuoles of the wild type [131]. Orthologs of AtABCC1 have been identified in numerous grasses including rice, maize, and barley. In barley, a vacuole kinetic analysis also suggests that this transport mechanism is conserved across species [132]. AtABCC3 was also shown to play an important role on Cd tolerance. Plants overexpressing AtABCC3 showed Cd-tolerant phenotypes, while a mutant defective in atabcc3 was more sensitive to Cd stress. The function of AtABCC3 is highly dependent on PCs. When AtABCC3 was expressed in the cad1-3 mutant (defective in PC synthesis) or AtABCC3-overexpressing plants were subjected to BSO (an PC biosynthesis inhibitor), the Cd-tolerant phenotype disappeared [110].
In contrast to ABCCs, for which Cd transport is coupled to phytochelatins, the heavy metal-transporting ATPases (HMAs), cation diffusion facilitator (CDF), and Ca2+ exchangers transport the ionic form of Cd (Cd2+). P1B-type ATPases have also been shown to play a key role on sequestering Cd in root vacuoles [54]. Of interest is the fact that OsHMA3 seems to be highly specific for Cd, while the Arabidopsis halleri HMA3 has a preference for Zn. However, the A. thaliana HMA3 shows broad substrate specificity being able to transport Co, Pb, Cd, and Zn [111,112]. OsHMA3, a vacuolar P1B-type ATPase was found to contribute 85.6% of the variance in Cd content between low-and high-cadmium accumulation varieties of rice [54]. The important role of OsHMA3 as controlling shoot Cd accumulation was demonstrated by QTL analysis of low and high Cd-accumulating rice cultivars [133]. In Arabidopsis, the function of HMA3 was attributed to limiting long-distance transport of Cd from the root to the shoot as this transporter is predominantly expressed in the root and thus operates in its sequestration here [113]. The crucial role of HMA3 for plants’ ability to handle high concentrations of Cd in shoots is also broadly reported for hyperaccumulating species. Working with hyperaccumulating Sedum alfredii species, Zhang et al. [134] reported high level of SaHMA3h expression leading to efficient detoxifying ability in this species. S. alfredii plants also had a larger number of gene copies as compared with its non-hyperaccumulator ecotype. In S. plumbizincicola shoots, particularly in young leaf cells, SpHMA3 is critical for Cd detoxification and acts by sequestering Cd into vacuoles [114]. Some transporters like CAXs and NRAMPs, which are tonoplast-localized, also play a role in Cd tolerance [135]. Most of the CAXs are Ca2+ specific, but in Arabidopsis, AtCAX2 and AtCAX4 were shown to have the ability to transport other metals like Cd, Zn, and Mn [115,116]. A heterologous overexpression of SaCAX2a from hyperaccumulating S. alfredii plants in tobacco resulted in improved Cd tolerance [134]. Likewise, the atnramp3atnramp4 double knockout mutant exhibits increased sensitivity to Cd [136] but also displayed Fe deficiency symptoms. NRAMP3 and NRAMP4 are also believed to be involved in the remobilization of essential metals by exporting them across the tonoplast membrane from the vacuole [137].
Besides those divalent cation transporters functioning in cellular sequestration, organic acids are considered to have an involvement in detoxification mechanisms. Of interest is that organic acids such as citric, malic, and carboxylic acids may stabilize Cd2+ in a complex form. The root hair of Cd hyperaccumulator Thlaspi caerulescens contains high levels of citric and malonic acids that distribute high Cd levels in cell walls, ensuring that plants can tolerate Cd-induced stress [138]. In leaves of the Cd hyperaccumulator Solanum nigrum, both acetic and citric acid bind with Cd, enabling the formation of Cd-organic acid complex for chelation [139]. Ehsan et al. [140] found that citric acid in culture solution forms a citric complex of Cd that can be easily translocated from the roots to shoots of Brassica napus without any toxicity occurrence, thus peaking its phytoextraction potential. The secretion of malic and oxalic acids in tissues of Agrogyron elongatum grown in the nutrient medium was positively correlated with peak Cd levels and tolerance mechanisms for coping with Cd toxicity [141]. In addition, the binding affinity of organic acids to Cd forms a complex with Cd2+ thus reducing the availability of free toxic Cd2+ in the growth medium and alleviates phytotoxicity induced by excess Cd.
5. Conclusions and the Way Forward
Cd has long been recognized as a major health threat to humans. It represents one of the most toxic substances released into the environment, and practically all human populations are environmentally exposed to Cd, mostly through plant-derived food. Thus, the ability to regulate Cd loading into the xylem and its delivery to the shoot with a transpiration stream in leafy vegetables is critical to avoid Cd entering the food chain. Equally important is the prevention of Cd being accumulated in cereal grains (by phloem-based retranslocation mechanism). While a significant progress has been made in revealing the molecular nature of transporters mediating Cd uptake and transport across cellular membranes, reducing Cd accumulation in plants is complicated by the fact that most known Cd transporters also operate in the transport of essential nutrients such as Zn, Fe, Mn, or Cu. Thus, the real progress in the field may be only achieved if transport of these nutrients and Cd is uncoupled. This calls for more studies dealing with structure–function relations of various transporters isoforms. In this context, Kawachi et al. [98,100] conducted a structure–function analysis of Arabidopsis MTP1 transporter from the cation diffusion facilitator family. Based on the crystal structure of the Escherichia coli and using site-directed mutagenesis, they showed that Ala substitution of either Asn258 in TM5 or Ser101 in TM2 reduced AtMTP1 selectivity for Zn2+ in yeast. They also showed that deletions in N-terminal and His-rich intra-molecular cytosolic domains and mutations of single residues flanking the transmembrane pore or participating in intra- or inter-molecular domain interactions selectively affected yeast’s ability to accumulate Co and Cd [100]. Thus, uncoupling of Cd transport from transport of essential micronutrients is technically possible and just required some more orchestrated efforts. The major hurdle in this process is a high tissue specificity of transporter operation. Two tissues are of a specific importance in this context. The first one is a xylem parenchyma in plant roots, a major “controller” of Cd loading into the xylem and its transport to the shoot. The second one is a phloem tissue that operates in the last step of the metal transport from mature leaves (where Cd is delivered) to the developing grains. Ion remobilization from leaves and transport to developing grains is currently terra incognita, hinting at many unexploited ways to improve crops genetically. Another promising and currently underexplored avenue is to understand the role of non-selective cation channels in Cd uptake and reveal mechanisms of their regulation. Arabidopsis genome harbors 40 non-selective cation channels (NSCC) [90]; 20 of them belong to ionotropic “glutamate receptor-like” (GLR) family. Other plant families also have over 10 GLR members in their genomes [93,94,95,96]. GLRs are believed to be tetramers consisting of different subunits, with preferential expression in root tissues [142]. The other 20 channels are classified as “cyclic nucleotide-gated channels” (CNGC). CNGCs are structurally similar to members of the superfamily of six transmembrane “Shaker-like” pore-loop ion channels [91,92]. All these channels need to be functionally characterized for their ability to uptake Cd2+ in functional (MIFE or patch-clamp electrophysiology) assays and then validated in genetic studies.
Author Contributions
Conceptualization, S.S. and X.H.; data collection, X.H., Q.W., S.D. and S.S.; writing—X.H., S.D., Q.W., and S.S.; critical revision, S.S., X.H. and M.Y.; funding acquisition, M.Y. and S.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by National Natural Science Foundation of China (31672228, 31870249), the Science and Technology Department of Guangdong Province (2018A050506085, 163-2018-XMZC-0001-05-0049), and the Science and Technology Bureau of Foshan (20161201010027).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Panos, P.; Marc, V.L.; Yusuf, Y.; Luca, M. Contaminated sites in Europe: Review of the current situation based on data collected through a European network. J. Environ. Public Health 2013, 2013, 158764. [Google Scholar]
- Alloway, B.J. Heavy Metals in Soils: Trace Metals and Metalloids in Soils and Their Bioavailability; Springer: Dordrecht, The Netherlands, 2013. [Google Scholar]
- Cook, M.E.; Morrow, H. Anthropogenic sources of cadmium in Canada. In National Workshop on Cadmium Transport into Plants; Canadian Network of Toxicology Centres: Ottawa, ON, Canada, 1995. [Google Scholar]
- Hayat, M.T.; Nauman, M.; Nazir, N.; Ali, S.; Bangash, N. Environmental hazards of cadmium: Past, present, and future. In Cadmium Toxicity and Tolerance in Plants. From Physiology to Remediation; Hasanuzzaman, M., Prasad, M.N.V., Fujita, M., Eds.; Academic Press: London, UK, 2019; pp. 163–183. [Google Scholar]
- World Health Organisation (WHO). Environmental Health Criteria 134—Cadmium International Programme on Chemical Safety (IPCS) Monograph; World Health Organization: Geneva, Switzerland, 1992. [Google Scholar]
- Xue, D.W.; Jiang, H.; Deng, X.X.; Zhang, X.Q.; Wang, H.; Xu, X.B.; Hu, J.; Zeng, D.; Guo, L.; Qian, Q. Comparative proteomic analysis provides new insights into cadmium accumulation in rice grain under cadmium stress. J. Hazard. Mater. 2014, 280, 269–278. [Google Scholar] [CrossRef]
- Clemens, S.; Ma, J.F. Toxic heavy metal and metalloid accumulation in crop plants and foods. Annu. Rev. Plant Biol. 2016, 67, 489–512. [Google Scholar] [CrossRef] [Green Version]
- Zu, Y.; Li, Y.; Chen, J.; Chen, H.Y.; Li, Q.; Schratz, C. Hyper accumulation of Pb, Zn and Cd in herbaceous grown on lead-zinc mining area in Yunnan, China. Environ. Int. 2005, 1, 755–762. [Google Scholar]
- Sanita, D.T.L.; Gabbrielli, R. Response to cadmium in higher plants. Environ. Exp. Bot. 1999, 41, 105–130. [Google Scholar] [CrossRef]
- Zhao, K.L.; Fu, W.J.; Ye, Z.Q.; Zhang, C.S. Contamination and spatial variation of heavy metals in the soil-rice system in Nanxun County, Southeastern China. J. Environ. Res. Public Health 2015, 12, 1577–1594. [Google Scholar] [CrossRef] [PubMed]
- Bauddh, K.; Singh, R.P. Growth, tolerance efficiency and phytoremediation potential of Ricinus communis (L.) and Brassica juncea (L.) in salinity and drought affected cadmium contaminated soil. Ecotoxicol. Environ. Saf. 2012, 85, 13–22. [Google Scholar] [CrossRef]
- Shi, G.; Xia, S.; Ye, J.; Huang, Y.; Liu, C.; Zhang, Z. PEG-simulated drought stress decreases cadmium accumulation in castor bean by altering root morphology. Environ. Exp. Bot. 2015, 111, 127–134. [Google Scholar] [CrossRef]
- Demeyer, A.; Nkana, J.V.; Verloo, M.G. Characteristics of wood ash and influence on soil properties and nutrient uptake: An overview. Bioresour. Technol. 2001, 77, 287–295. [Google Scholar] [CrossRef]
- Kabata-Pendias, A. Trace Elements in Soils and Plants, 4th ed.; CRC Press: Boca Raton, FL, USA, 2011. [Google Scholar]
- Smith, S.R. A critical review of the bioavailability and impacts of heavy metals in municipal solid waste composts compared to sewage sludge. Environ. Int. 2009, 35, 142–156. [Google Scholar] [CrossRef]
- De Oliveira, V.H.; Tibbett, M. Cd and Zn interactions and toxicity in ectomycorrhizal basidiomycetes in axenic culture. PeerJ 2018, 6, e4478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johansen, J.L.; Rønn, R.; Ekelund, F. Toxicity of cadmium and zinc to small soil protists. Environ. Pollut. 2018, 242, 1510–1517. [Google Scholar] [CrossRef] [PubMed]
- Godt, J.; Scheidig, F.; Grosse-Siestrup, C.; Esche, V.; Brandenburg, P.; Reich, A.; Groneberg, D.A. The toxicity of cadmium and resulting hazards for human health. J. Occup. Med. Toxicol. 2016, 1, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nawrot, T.; Plusquin, M.; Hogervorst, J.; Roels, H.A.; Celis, H.; Thijs, L.; Vangronsveld, J.; Van Hecke, E.; Staessen, J.A. Environmental exposure to cadmium and risk of cancer: A prospective populationbased study. Lancet Oncol. 2006, 7, 119–126. [Google Scholar] [CrossRef] [Green Version]
- Tsukahara, T.; Ezaki, T.; Moriguchi, J.; Furuki, K.; Shimbo, S.; Matsuda-Inoguchi, N.; Ikeda, M. Rice as the most influential source of cadmium intake among general Japanese population. Sci. Total Environ. 2003, 305, 41–51. [Google Scholar] [CrossRef]
- Inaba, T.; Kobayashi, E.; Suwazono, Y.; Uetani, M.; Oishi, M.; Nakagawa, H.; Nogawa, K. Estimation of cumulative cadmium intake causing Itai-itai disease. Toxicol. Lett. 2005, 159, 192–201. [Google Scholar] [CrossRef]
- Kara, H.; Karatas, F.; Canatan, H.; Servi, K. Effects of exogenous metallothionein on acute cadmium toxicity in rats. Biol. Trace Elem. Res. 2005, 104, 223. [Google Scholar] [CrossRef]
- Dudley, R.E.; Svoboda, D.J.; Klaassen, C.D. Acute exposure to cadmium causes severe liver injury in rats. Toxicol. Appl. Pharm. 1982, 65, 302–313. [Google Scholar] [CrossRef]
- Touceda-González, M.; Brader, G.; Antonielli, L.; Ravindran, V.B.; Waldner, G.; Friesl-Hanl, W.; Corretto, E.; Campisano, A.; Pancher, M.; Angela, S. Combined amendment of immobilizers and the plant growth-promoting strain Burkholderia phytofirmans PsJN favours plant growth and reduces heavy metal uptake. Soil Biol. Biochem. 2015, 91, 140–150. [Google Scholar] [CrossRef] [Green Version]
- Van Assche, F.J. A Stepwise Model to Quantify the Relative Contribution of Different Environmental Sources to Human Cadmium Exposure. In Proceedings of the 8th International Nickel-Cadmium Battery Conference, Prague, Czech Republic, 21–22 September 1998. [Google Scholar]
- Clemens, S.; Aarts, M.G.M.; Thomine, S.; Verbruggen, N. Plant science: The key to preventing slow cadmium poisoning. Trends Plant Sci. 2013, 18, 92–99. [Google Scholar] [CrossRef]
- Uraguchi, S.; Fujiwara, T. Rice breaks ground for cadmium free cereals. Curr. Opin. Plant Biol. 2013, 16, 328–334. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Liu, X.N.; Ding, C.; Wu, L. The dynamic simulation of rice growth parameters under cadmium stress with the assimilation of multi-period spectral indices and crop model. Field Crops Res. 2015, 183, 225–234. [Google Scholar] [CrossRef]
- Song, W.E.; Chen, S.B.; Liu, J.F.; Chen, L.; Song, N.N.; Li, N.; Liu, B.V. Variation of Cd concentration in various rice cultivars and derivation of cadmium toxicity thresholds for paddy soil by species-sensitivity distribution. J. Integr. Agric. 2015, 14, 1845–1854. [Google Scholar] [CrossRef]
- Chen, H.; Yang, X.; Wang, P.; Wang, Z.; Li, M.; Zhao, F.J. Dietary cadmium intake from rice and vegetables and potential health risk: A case study in Xiangtan, southern China. Sci. Total Environ. 2018, 639, 271–277. [Google Scholar] [CrossRef]
- Song, Y.; Wang, Y.; Mao, W.; Sui, H.; Yong, L.; Yang, D.; Jiang, D.; Zhang, L.; Gong, Y. Dietary cadmium exposure assessment among the Chinese population. PLoS ONE 2017, 12, e0177978. [Google Scholar] [CrossRef] [Green Version]
- Baldantoni, D.; Morra, L.; Zaccardelli, M.; Alfani, A. Cadmium accumulation in leaves of leafy vegetables. Ecotoxicol. Environ. Saf. 2016, 123, 89–94. [Google Scholar] [CrossRef]
- Wang, J.; Yu, N.; Mu, G.; Shinwari, K.I.; Shen, Z.; Zheng, L. Screening for Cd-safe cultivars of Chinese cabbage and a preliminary study on the mechanisms of Cd accumulation. Int. J. Environ. Res. Public Health 2017, 14, 395. [Google Scholar] [CrossRef] [Green Version]
- FAO Food and Agriculture Organization. ProdStat. Core Production Data Base, Core Production Data Base, Electronic Resource. 2012. Available online: http://faostat.fao.org/ (accessed on 11 October 2019).
- Mohamed, A.A.; Castagna, A.; Ranieri, A.; Sanita di Toppi, L. Cadmium tolerance in Brassica juncea roots and shoots is affected by antioxidant status and phytochelatin biosynthesis. Plant Physiol. Biochem. 2012, 57, 15–22. [Google Scholar] [CrossRef]
- Navarro-Leon, E.; Oviedo-Silva, J.; Ruiz, J.M.; Blasco, B. Possible role of HMA4a TILLING mutants of Brassica rapa in cadmium. Ecotoxicol. Environ. Saf. 2019, 180, 88–94. [Google Scholar] [CrossRef]
- Hassan, M.J.; Zhang, G.; Zhu, Z. Influence of cadmium toxicity on plant growth and nitrogen uptake in rice as affected by nitrogen form. J. Plant Nutr. 2008, 31, 251–262. [Google Scholar] [CrossRef]
- Alcantara, E.; Romera, F.J.; Canete, M.; De La Guardia, M.D. Effects of heavy metals on both induction and function of root Fe(III) reductase in Fe-deficient cucumber (Cucumis sativus L.) plants. J. Exp. Bot. 1994, 45, 1893–1898. [Google Scholar] [CrossRef]
- Sharma, V.; Pant, D. Structural basis for expanding the application of bioligand in metal bioremediation: A review. Bioresour. Technol. 2018, 252, 188–197. [Google Scholar] [CrossRef] [PubMed]
- Cailliatte, R.; Lapeyre, B.; Briat, J.F.; Mari, S.; Curie, C. The NRAMP6 metal transporter contributes to cadmium toxicity. Biochem. J. 2009, 422, 217–228. [Google Scholar] [CrossRef] [PubMed]
- Feng, S.; Tan, J.; Zhang, Y.; Liang, S.; Xiang, S.; Wang, H.; Chai, T. Isolation and characterization of a novel cadmium-regulated Yellow Stripe-Like transporter (SnYSL3) in Solanum nigrum. Plant Cell Rep. 2017, 36, 281–296. [Google Scholar] [CrossRef]
- Takahashi, R.; Ishimaru, Y.; Senoura, T.; Shimo, H.; Ishikawa, S.; Arao, T.; Nakanishi, H.; Nishizawa, N.K. The OsNRAMP1 iron transporter is involved in Cd accumulation in rice. J. Exp. Bot. 2011, 62, 4843–4850. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.L.; Yin, X.B.; Banuelos, G.S.; Lin, Z.Q.; Liu, Y.; Li, M.; Yuan, L.X. Indications of selenium protection against cadmium and lead toxicity in oilseed rape (Brassica napus L.). Front. Plant Sci. 2016, 7, 1875. [Google Scholar] [CrossRef]
- Lindberg, S.; Landberg, T.; Greger, M. A new method to detect cadmium uptake in protoplasts. Planta 2004, 219, 526–532. [Google Scholar] [CrossRef]
- Li, D.D.; Zhou, D.M. Toxicity and subcellular distribution of cadmium in wheat as affected by dissolved organic acids. J. Environ. Sci. 2012, 24, 903–911. [Google Scholar] [CrossRef]
- Jasinski, M.; Sudre, D.; Schansker, G.; Schellenberg, M.; Constant, S.; Martinoia, E. Atosa1, a member of the abc1-like family, as a new factor in cadmium and oxidative stress response. Plant Physiol. 2008, 147, 719–731. [Google Scholar] [CrossRef] [Green Version]
- Bortner, C.D.; Hughes, F.M.; Cidlowski, J.A. A primary role for K+ and Na+ efflux in the activation of apoptosis. J. Biol. Chem. 1977, 272, 32436–32442. [Google Scholar] [CrossRef] [Green Version]
- Demidchik, V.; Cuin, T.A.; Svistunenko, D.; Smith, S.J.; Miller, A.J.; Shabala, S.; Sokolik, A.; Yurin, V. Arabidopsis root K+ efflux conductance by hydroxyl radicals: Single-channel properties, genetic basis and involvement in stress-induced cell death. J. Cell Sci. 2010, 123, 1468–1479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shabala, S. Physiological and cellular aspects of phytotoxicity tolerance in plants: The role of membrane transporters and implications for crop breeding for waterlogging tolerance. New Phytol. 2007, 190, 289–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Millán, A.F.; Sagardoy, R.; Solanas, M.; Abadía, A.; Abadía, J. Cadmium toxicity in tomato (Lycopersicon esculentum) plants grown in hydroponics. Environ. Exp. Bot. 2009, 65, 376–385. [Google Scholar] [CrossRef]
- Tian, S.; Lu, L.; Labavitch, J.; Yang, X.; He, Z.; Hu, H.; Sarangi, R.; Newville, M.; Commisso, J.; Brown, P. Cellular sequestration of cadmium in the hyperaccumulator plant species Sedum alfredii. Plant Physiol. 2011, 157, 1914–1925. [Google Scholar] [CrossRef] [Green Version]
- Ueno, D.; Kono, I.; Yokosho, K.; Ando, T.; Yano, M.; Ma, J.F. A major quantitative trait locus controlling cadmium translocation in rice (Oryza sativa). New Phytol. 2009, 182, 644–653. [Google Scholar] [CrossRef]
- Ueno, D.; Koyama, E.; Kono, I.; Ando, T.; Yano, M.; Ma, J.F. Identification of a novel major quantitative trait locus controlling distribution of Cd between roots and shoots in Rice. Plant Cell Physiol. 2009, 50, 2223–2233. [Google Scholar] [CrossRef] [Green Version]
- Ueno, D.; Yamaji, N.; Kono, I.; Huang, C.F.; Ando, T.; Yano, M.; Ma, J.F. Gene limiting cadmium accumulation in rice. Proc. Natl. Acad. Sci. USA 2010, 107, 16500–16505. [Google Scholar] [CrossRef] [Green Version]
- Yan, J.; Wang, P.; Wang, P.; Yang, M.; Lian, X.; Tang, Z.; Huang, C.; Salt, D.; Zhao, F.J. A loss-of-function allele of associated with high cadmium accumulation in shoots and grain of rice cultivars. Plant Cell Environ. 2016, 39, 1941–1954. [Google Scholar] [CrossRef] [Green Version]
- Hu, D.; Sheng, Z.; Li, Q.; Chen, W.; Wei, X.; Xie, L.; Jiao, G.; Shao, G.; Wang, J.; Tang, S.; et al. Identification of QTLs associated with cadmium concentration in rice grains. J. Integr. Agric. 2018, 17, 1563–1573. [Google Scholar] [CrossRef] [Green Version]
- Kuramata, M.; Masuya, S.; Takahashi, Y.; Kitagawa, E.; Inoue, C.; Ishikawa, S.; Youssefian, S.; Kusano, T. Novel cysteine-rich peptides from Digitaria ciliaris and Oryza sativa enhance tolerance to cadmium by limiting its cellular accumulation. Plant Cell Physiol. 2009, 50, 106–117. [Google Scholar] [CrossRef] [Green Version]
- Xia, J.X.; Yamaji, N.; Ma, J.F. A plasma membrane-localized small peptide is involved in rice aluminum tolerance. Plant J. 2013, 76, 345–355. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Yang, M.; Li, Y.; Tian, J.; Zhang, Y.; Liang, L.; Liu, Z.; Chen, K.; Li, Y.; Lv, K.; et al. Comprehensive analysis of variation of cadmium accumulation in rice and detection of a new weak allele of OsHMA3. J. Exp. Bot. 2019, 70, 6389–6400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.; Gao, Z.; Shang, L.; Yang, C.; Ruan, B.; Zeng, D.; Guo, L.; Zhao, F.; Huang, C.; Qian, Q. Natural variation in the promoter of OsHMA3 contributes to differential grain cadmium accumulation between indica and japonica rice. J. Integr. Plant Biol. 2009, 1672–9072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Chen, S.; Chen, M.; Zheng, G.; Peng, Y.; Shi, X.; Qin, P.; Xu, X.; Teng, S. Association study reveals genetic loci responsible for arsenic, cadmium and lead accumulation in rice grain in contaminated farmlands. Front. Plant Sci. 2009, 10, 61. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Xu, Y.; Ma, Q.; Xu, W.; Wang, T.; Xue, Y.; Chong, K. Overexpression of an R1R2R3 MYB gene, OsMYB3R-2, increases tolerance to freezing, drought, and salt stress in transgenic Arabidopsis. Plant Physiol. 2007, 143, 1739–1751. [Google Scholar] [CrossRef] [Green Version]
- Ma, Q.; Dai, X.; Xu, Y.; Guo, J.; Liu, Y.; Chen, N.; Xiao, J.; Zhang, D.; Xu, Z.; Zhang, X.; et al. Enhanced tolerance to chilling stress in OsMYB3R2 transgenic rice is mediated by alteration in cell cycle and ectopic expression of stress genes. Plant Physiol. 2009, 150, 244–256. [Google Scholar] [CrossRef]
- Lv, Y.; Yang, M.; Hu, D.; Yang, Z.; Ma, S.; Li, X.; Xiong, L. The OsMYB30 transcription factor suppresses cold tolerance by interacting with a JAZ protein and suppressing b-amylase expression. Plant Physiol. 2017, 173, 1475–1491. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Wang, R.; Ju, Q.; Li, W.; Tran, L.P.; Xu, J. The R2R3-MYB transcription factor MYB49 regulates cadmium accumulation. Plant Physiol. 2019, 180, 529–542. [Google Scholar] [CrossRef]
- Benitez, E.R.; Hajika, M.; Yamada, T.; Takahashi, K.; Oki, N.; Yamada, N.; Nakamura, T.; Kanamaru, K. A Major QTL controlling seed cadmium accumulation in soybean. Crop Sci. 2010, 50, 1728–1734. [Google Scholar] [CrossRef]
- Vollmann, J.; Lošák, T.; Pachner, M.; Watanabe, D.; Musilová, L.; Hlušek, J. Soybean cadmium concentration: Validation of a QTL affecting seed cadmium accumulation for improved food safety. Euphytica 2014, 203, 177–184. [Google Scholar] [CrossRef]
- Clemens, S. Toxic metal accumulation, responses to exposure and mechanisms of tolerance in plants. Biochimie 2006, 88, 1707–1719. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.H.; Hou, H.; Zhao, L.; Sun, Z.J.; Lu, Y.F.; Li, H. Mitigation of Cd accumulation in rice from Cd-contaminated paddy soil by foliar dressing of S and P. Sci. Total Environ. 2019, 690, 321–328. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Wang, F.; Liu, S.; Du, Y.; Li, F.R.; Du, R.Y.; Wen, D.; Zhao, J. Comparative responses to silicon and selenium in relation to cadmium uptake, compartmentation in roots, and xylem transport in flowering Chinese cabbage (Brassica campestris L. ssp. chinensis var. utilis) under cadmium stress. J. Exp. Bot. 2016, 131, 173–180. [Google Scholar] [CrossRef]
- Li, Y.; Liang, X.F.; Huang, Q.Q.; Xu, Y.M.; Yang, F. Inhibition of Cd accumulation in grains of wheat and rice under rotation mode using composite silicate amendment. RSC Adv. 2019, 61, 35539–35548. [Google Scholar] [CrossRef] [Green Version]
- Lux, A.; Vaculik, M.; Martinka, M.; Liskova, D.; Kulkarni, M.G.; Stirk, W.A.; Van, S. Cadmium induces hypodermal periderm formation in the roots of the monocotyledonous medicinal plant Merwilla plumbea. Ann. Bot. 2011, 107, 285–292. [Google Scholar] [CrossRef]
- Sasaki, A.; Yamaji, N.; Yokosho, K.; Ma, J.F. Nramp5 is a major transporter responsible for manganese and cadmium uptake in rice. Plant Cell 2012, 24, 2155–2167. [Google Scholar] [CrossRef] [Green Version]
- Uraguchi, S.; Kamiya, T.; Sakamoto, T.; Kasai, K.; Sato, Y.; Nagamura, Y.; Yoshida, A.; Kyozuka, J.; Ishikawa, S.; Fujiwara, T. Low-affinity cation transporter (OsLCT1) regulates cadmium transport into rice grains. Proc. Natl. Acad. Sci. USA 2011, 108, 20959–20964. [Google Scholar] [CrossRef] [Green Version]
- Curie, C.; Panaviene, Z.; Loulergue, C.; Dellaporta, S.L.; Briat, J.F.; Walker, E.L. Maize yellow stripe1 encodes a membrane protein directly involved in Fe (III) uptake. Nature 2001, 409, 346–349. [Google Scholar] [CrossRef]
- Murata, Y.; Ma, J.F.; Yamaji, N.; Ueno, D.; Nomoto, K.; Iwashita, T. A specific transporter for iron (III)–phytosiderophore in barley roots. Plant J. 2006, 46, 563–572. [Google Scholar] [CrossRef]
- Koike, S.; Inoue, H.; Mizuno, D.; Takahashi, M.; Nakanishi, H.; Mori, S.; Nishizawa, N.K. OsYSL2 is a rice metal-nicotianamine transporter that is regulated by iron and expressed in the phloem. Plant J. 2004, 39, 415–424. [Google Scholar] [CrossRef]
- Ogawa, I.; Nakanishi, H.; Ishimaru, Y.; Takahashi, M.; Mori, S.; Nishizawa, N.K. Iron deficiency enhanced Cd uptake and translocation by Fe2+ transporters, OsIRT1 and OsIRT2, in rice. Plant Cell Physiol. 2006, 47, S231. [Google Scholar]
- Yoshihara, T.; Hodoshima, H.; Miyano, Y.; Shoji, K.; Shimada, H.; Goto, F. Cadmium inducible Fe deficiency responses observed from macro and molecular views in tobacco plants. Plant Cell Rep. 2006, 25, 365–373. [Google Scholar] [CrossRef] [PubMed]
- Lanquar, V.; Lelièvre, F.; Barbier-Brygoo, H.; Thomine, S. Regulation and function of AtNRAMP4 metal transporter protein. Soil Sci. 2004, 50, 1141–1150. [Google Scholar] [CrossRef]
- Nevo, Y.; Nelson, N. The NRAMP family of metal-ion transporters. BBA-Mol. Cell Res. 2006, 1763, 609–620. [Google Scholar] [CrossRef] [Green Version]
- Ishimaru, Y.; Takahashi, R.; Bashir, K.; Shimo, H.; Senoura, T.; Sugimoto, K.; Ono, K.; Yano, M.; Ishikawa, S.; Arao, T.; et al. Characterizing the role of rice NRAMP5 in manganese, iron and cadmium transport. Sci. Rep. 2012, 2, 286. [Google Scholar] [CrossRef]
- Wu, D.; Yamaji, N.; Yamane, M.; Kashino-Fujii, M.; Sato, K.; Ma, J. The HvNramp5 transporter mediates uptake of cadmium and manganese, but not iron. Plant Physiol. 2016, 172, 1899–1910. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, R.; Ishimaru, Y.; Shimo, H.; Bashir, K.; Senoura, T.; Sugimoto, K.; Ono, K.; Suzui, N.; Kawachi, N.; Ishii, S.; et al. From laboratory to field: OsNRAMP5-knockdown rice is a promising candidate for Cd phytoremediation in paddy fields. PLoS ONE 2014, 9, e98816. [Google Scholar] [CrossRef]
- Yamaji, N.; Sasaki, A.; Xia, J.X.; Yokosho, K.; Ma, J.F. A node-based switch for preferential distribution of manganese in rice. Nat. Commun. 2013, 4, 1–11. [Google Scholar] [CrossRef]
- Xia, J.; Yamaji, N.; Kasai, T.; Ma, J.F. Plasma membrane-localized transporter for aluminum in rice. Proc. Natl. Acad. Sci. USA 2010, 107, 18381–18385. [Google Scholar] [CrossRef] [Green Version]
- Thomine, S.; Lelievre, F.; Debarbieux, E.; Schroeder, J.I.; Barbier-Brygoo, H. AtNRAMP3, a multispecific vacuolar metal transporter involved in plant responses to iron deficiency. Plant J. 2003, 34, 685–695. [Google Scholar] [CrossRef]
- Thomine, S.; Wang, R.; Ward, J.M.; Crawford, N.M.; Schroeder, J.I. Cadmium and iron transport by members of a plant transporter gene family in Arabidopsis with homology to NRAMP genes. Proc. Natl. Acad. Sci. USA 2000, 97, 4991–4996. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Liu, X.; Peijnenburg, W.J.G.M.; Zhao, J.; Chen, X.; Yu, J.; Wu, H. Pathways of cadmium fluxes in the root of the halophyte Suaeda salsa. Ecotoxicol. Environ. Saf. 2012, 75, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Demidchik, V.; Shabala, S.; Isayenkov, S.; Cuin, T.A.; Pottosin, I. Calcium transport across plant membranes: Mechanisms and functions. New Phytol. 2018, 220, 49–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeFalco, T.A.; Marshall, C.B.; Munro, K.; Kang, H.G.; Moeder, W.; Ikura, M.; Snedden, W.A.; Yoshioka, K. Multiple calmodulin-binding sites positively and negatively regulate Arabidopsis CYCLIC NUCLEOTIDE-GATED CHANNEL12. Plant Cell 2016, 28, 1738–1751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jha, S.K.; Sharma, M.; Pandey, G.K. Role of cyclic nucleotide gated channels in stress management in plants. Curr. Genom. 2016, 17, 315–329. [Google Scholar] [CrossRef]
- Lam, H.M.; Chiu, J.; Hsieh, M.H.; Meisel, L.; Oliveira, I.C.; Shin, M.; Coruzzi, G. Glutamate-receptor genes in plants. Nature 1998, 396, 125–126. [Google Scholar] [CrossRef]
- Davenport, R. Glutamate receptors in plants. Ann. Bot. 2002, 90, 549–557. [Google Scholar] [CrossRef]
- Aouini, A.; Matsukura, C.; Ezura, H.; Asamizu, E. Characterisation of 13 glutamate receptor-like genes encoded in the tomato genome by structure, phylogeny and expression profiles. Gene 2012, 493, 36–43. [Google Scholar] [CrossRef]
- Ni, J.; Yu, Z.; Du, G.; Zhang, Y.; Taylor, J.L.; Shen, C.; Xu, J.; Liu, X.; Wang, Y.; Wu, Y. Heterologous expression and functional analysis of rice GLUTAMATE RECEPTOR-LIKE family indicates its role in glutamate triggered calcium flux in rice roots. Rice 2016, 9, 9. [Google Scholar] [CrossRef] [Green Version]
- Montanini, B.; Blaudez, D.; Jeandroz, S.; Sanders, D.; Chalot, M. Phylogenetic and functional analysis of the Cation Diffusion Facilitator (CDF) family: Improved signature and prediction of substrate specificity. BMC Genom. 2007, 8, 107. [Google Scholar] [CrossRef] [Green Version]
- Kawachi, M.; Kobae, Y.; Mimura, T.; Maeshima, M. Deletion of ahistidine-rich loop of AtMTP1, a vacuolar Zn2+/H+ antiporter of Arabidopsis thaliana, stimulates the transport activity. J. Biol. Chem. 2008, 283, 8374–8383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desbrosses-Fonrouge, A.G.; Voigt, K.; Schroder, A.; Arrivault, S.; Thomine, S.; Kramer, U. Arabidopsis thaliana MTP1 is a Zn transporter in the vacuolar membrane which mediates Zn detoxification and drives leaf Zn accumulation. FEBS Lett. 2005, 579, 4165–4174. [Google Scholar] [CrossRef] [PubMed]
- Kawachi, M.; Kobae, Y.; Kogawa, S.; Mimura, T.; Kramer, U.; Maeshima, M. Amino acid screening based on structural modeling identifies critical residues for the function, ion selectivity and structure of Arabidopsis MTP1. FEBS J. 2012, 279, 2339–2356. [Google Scholar] [CrossRef] [PubMed]
- Migocka, M.; Papierniak, A.; Kosieradzka, A.; Posyniak, E.; Maciaszczyk-Dziubinska, E.; Biskup, R.; Garbiec, A.; Marchewka, T. Cucumber metal tolerance protein CsMTP9 is a plasma membrane H+-coupled antiporter involved in the Mn2+ and Cd2+ efflux from root cells. Plant J. 2015, 84, 1045–1058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ueno, D.; Sasaki, A.; Yamaji, N.; Miyaji, T.; Fujii, Y.; Takemoto, Y.; Moriyama, S.; Che, J.; Moriyama, Y.; Iwasaki, K.; et al. A polarly localized transporter for efficient manganese uptake in rice. Nat. Plants 2015, 1, 1–8. [Google Scholar] [CrossRef]
- Chen, Z.; Fujii, Y.; Yamaji, N.; Masuda, S.; Takemoto, Y.; Kamiya, T. Mn tolerance in rice is mediated by MTP8.1 a member of the cation diffusion facilitator family. J. Exp. Bot. 2013, 64, 4375–4387. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Gustin, J.L.; Lahner, B.; Persans, M.W.; Baek, D.; Yun, D.J.; Salt, D.E. The plant CDF family member TgMTP1 from the Ni/Zn hyperaccumulator Thlaspi goesingense acts to enhance efflux of Zn at the plasma membrane when expressed in Saccharomyce cerevisiae. Plant J. 2004, 39, 237–251. [Google Scholar] [CrossRef]
- Deng, F.; Yamaji, N.; Xia, J.; Ma, J.F. A member of the heavy metal P-type ATPase OsHMA5 is involved in xylem loading of copper in rice. Plant Physiol. 2013, 163, 1353–1362. [Google Scholar] [CrossRef] [Green Version]
- Hussain, D.; Haydon, M.J.; Wang, Y.; Wong, E.; Sherson, S.M.; Young, J.; Camakaris, J.; Harper, J.F.; Cobbett, C.S. P-type ATPase heavy metal transporters with roles in essential zinc homeostasis in Arabidopsis. Plant Cell 2004, 16, 1327–1339. [Google Scholar] [CrossRef] [Green Version]
- Lekeux, G.; Crowet, J.; Nouet, C.; Joris, M.; Jadoul, A.; Bosman, B.; Carnol, M.; Motte, P.; Lins, L.; Galleni, M.; et al. Homology modeling and in vivo functional characterization of the zinc permeation pathway in a heavy metal P-type ATPase. J. Exp. Bot. 2019, 70, 329–341. [Google Scholar] [CrossRef]
- Uraguchi, S.; Kamiya, T.; Clemens, S.; Fujiwara, T. Charaterization of OsLCT1, a cadmium transporter from Indica rice (Oryza sativa). Physiol. Plant. 2014, 151, 339–347. [Google Scholar] [CrossRef] [PubMed]
- Song, W.Y.; Park, J.; Mendoza-Cozatl, D.G.; Suter-Grotemeyer, M.; Shim, D.; Hortensteiner, S.; Geisler, M.; Weder, B.; Rea, P.A.; Rentsch, D. Arsenic tolerance in Arabidopsis is mediated by two ABCC-type phytochelatin transporters. Proc. Natl. Acad. Sci. USA 2010, 107, 21187–21192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brunetti, P.; Zanella, L.; De, P.A.; Di, L.D.; Cecchetti, V.; Falasca, G. Cadmium-inducible expression of the ABC-type transporter AtABCC3 increases phytochelatin-mediated cadmium tolerance in Arabidopsis. J. Exp. Bot. 2015, 66, 3815–3829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becher, M.; Talke, I.N.; Krall, L.; Kramer, U. Cross-species microarray transcript profiling reveals high constitutive expression of metal homeostasis genes in shoots of the zinc hyperaccumulator Arabidopsis halleri. Plant J. 2004, 37, 251–268. [Google Scholar] [CrossRef]
- Morel, M.; Crouzet, J.; Gravot, A.; Auroy, P.; Leonhardt, N.; Vavasseur, A. AtHMA3, a P1B-ATPase allowing Cd/Zn/Co/Pb vacuolar storage in Arabidopsis. Plant Physiol. 2009, 149, 894–904. [Google Scholar] [CrossRef] [Green Version]
- Chao, D.Y.; Silva, A.; Baxter, I.; Huang, Y.S.; Nordborg, M.; Danku, J. Genome-wide association studies identify heavy metal ATPase3 as the primary determinant of natural variation in leaf cadmium in Arabidopsis thaliana. PLoS Genet. 2012, 8, e1002923. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Zhao, H.; Wu, L.; Liu, A.; Zhao, F.J.; Xu, W. Heavy metal ATPase3 (HMA3) confers cadmium hypertolerance on the cadmium/zinc hyperaccumulator Sedum plumbizincicola. New Phytol. 2017, 215, 687–698. [Google Scholar] [CrossRef] [Green Version]
- Korenkov, V.; Hirschi, K.; Crutchfield, J.D.; Wagner, G.J. Enhancing tonoplast Cd/H antiport activity increases Cd, Zn, and Mn tolerance, and impacts root/shoot Cd partitioning in Nicotiana tabacum L. Planta 2007, 226, 1379–1387. [Google Scholar] [CrossRef]
- Korenkov, V.; King, B.; Hirschi, K.; Wagner, G.J. Root-selective expression of AtCAX4 and AtCAX2 results in reduced lamina cadmium in field-grown Nicotiana tabacum L. Plant Biotechnol. J. 2009, 7, 219–226. [Google Scholar] [CrossRef]
- Migocka, M.; Kosieradzka, A.; Papierniak, A.; Maciaszczyk-Dziubinska, E.; Posyniak, E.; Garbiec, A.; Filleur, S. Two metal-tolerance proteins, MTP1 and MTP4, are involved in Zn homeostasis and Cd sequestration in cucumber cells. J. Exp. Bot. 2014, 66, 1001–1015. [Google Scholar] [CrossRef] [Green Version]
- Migocka, M.; Papierniak, A.; Maciaszczyk-Dziubinska, E.; Pozdzik, P.; Posyniak, E.; Garbiec, A.; Filleur, S. Cucumber metal transport protein MTP8 confers increased tolerance to manganese when expressed in yeast and Arabidopsis thaliana. J. Exp. Bot. 2014, 65, 5367–5384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Podar, D.; Scherer, J.; Noordally, Z.; Herzyk, P.; Nies, D.; Sanders, D. Metal selectivity determinants in a family of transition metal transporters. J. Biol. Chem. 2012, 287, 3185–3196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, R.; Ishimaru, Y.; Shimo, H.; Ogo, Y.; Senoura, T.; Nishizawa, N.K.; Nakanishi, H. The OsHMA2 transporter is involved in root to shoot translocation of Zn and Cd in rice. Plant Cell Environ. 2012, 35, 1948–1957. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, A.; Yamaji, N.; Ma, J. Overexpression of OsHMA3 enhances Cd tolerance and expression of Zn transporter genes in rice. J. Exp. Bot. 2014, 65, 6013–6021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kopittke, P.M.; DeJonge, M.D.; Wang, P.; Mckenna, B.A.; Lombi, E.; Paterson, D.J. Laterally resolved speciation of arsenic in roots of wheat and rice using fluorescence-XANES imaging. New Phytol. 2013, 201, 1251–1262. [Google Scholar] [CrossRef] [PubMed]
- Martinoia, E.; Meyer, S.; De Angeli, A.; Nagy, R. Vacuolar transporters in their physiological context. Annu. Rev. Plant. Biol. 2012, 63, 183–213. [Google Scholar] [CrossRef]
- Mathys, W. The role of malate, oxalate, and mustard oil glucosides in the evolution of zinc resistance in herbage plants. Physiol. Plant. 1997, 40, 130–136. [Google Scholar] [CrossRef]
- Ortiz, D.F.; Kreppel, L.; Speiser, D.M.; Scheel, G.; McDonald, G.; Ow, D.W. Heavy metal tolerance in the fission yeast requires an ATP-binding cassette type vacuolar membrane transporter. EMBO J. 1992, 11, 3491–3499. [Google Scholar] [CrossRef]
- Ortiz, D.F.; Ruscitti, T.; McCue, K.F.; Ow, D.W. Transport of metal- binding peptides by HMT1, a fission Yeast ABC-type vacuolar membrane protein. J. Biol. Chem. 1995, 270, 4721–4728. [Google Scholar] [CrossRef] [Green Version]
- Kuriakose, S.V.; Prasad, M.N.V. Cadmium stress affects seed germination and seedling growth in Sorghumbicolor (L.) Moench by changing the activities of hydrolyzing enzymes. Plant Growth Reg. 2008, 54, 143–156. [Google Scholar] [CrossRef]
- Sooksa-Nguan, T.; Yakubov, B.; Kozlovskyy, V.; Barkume, C.M.; Howe, K.J.; Thannhauser, T.W. Drosophila ABCtransporter, DmHMT-1, confers tolerance to cadmium. DmHMT-1 and its yeast homolog, SpHMT-1, are not essential for vacuolar phytochelatin sequestration. J. Biol. Chem. 2009, 284, 354–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vatamaniuk, O.K.; Bucher, E.A.; Sundaram, M.V.; Rea, P.A. CeHMT-1, a putative phytochelatin transporter, is required for cadmium tolerance in Caenorhab ditiselegans. J. Biol. Chem. 2005, 280, 23684–23690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Y.P.; Li, Z.S.; Drozdowicz, Y.M.; Hortensteiner, S.; Martinoia, E.; Rea, P.A. AtMRP2, an Arabidopsis ATP binding cassette transporter able to transport glutathione S-conjugates and chlorophyll catabolites: Functional comparisons with AtMRP1. Plant Cell 1998, 10, 267–282. [Google Scholar] [PubMed] [Green Version]
- Park, J.; Song, W.Y.; Ko, D.; Eom, Y.; Hansen, T.H.; Schiller, M.; Lee, T.G.; Martinoia, E.; Lee, Y. The phytochelatin transporters AtABCC1 and AtABCC2 mediate tolerance to cadmium and mercury. Plant J. 2012, 69, 278–288. [Google Scholar] [CrossRef]
- Song, W.Y.; Mendoza-Cozatl, D.; Lee, Y.; Schroeder, J.; Ahn, S.N.; Lee, H.S. Phytochelatin-metal(loid) transport into vacuoles shows different substrate preferences in barley and Arabidopsis. Plant Cell Environ. 2014, 37, 1192–1201. [Google Scholar] [CrossRef] [Green Version]
- Miyadate, H.; Adachi, S.; Hiraizumi, A.; Tezuka, K.; Nakazawa, N.; Kawamoto, T. OsHMA3, a P-1B-type of ATPase affects root-to-shoot cadmium translocation in rice by mediating efflux into vacuoles. New Phytol. 2011, 189, 190–199. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, M.; Shohag, M.J.; Tian, S.; Song, H.; Feng, Y. Enhanced expression of SaHMA3 plays critical roles in Cd hyperaccumulation and hypertolerance in Cd hyperaccumulator Sedum alfredii Hance. Planta 2016, 243, 577–589. [Google Scholar] [CrossRef]
- Oomen, R.J.; Wu, J.; Lelievre, F.; Blanchet, S.; Richaud, P.; Barbier-Brygoo, H. Functional characterization of NRAMP3 and NRAMP4 from the metal hyperaccumulator Thlaspi caerulescens. New Phytol. 2009, 181, 637–650. [Google Scholar] [CrossRef]
- Baliardini, C.; Meyer, C.L.; Salis, P.; Saumitou-Laprade, P.; Verbruggen, N. CATION EXCHANGER1 cosegregates with cadmium tolerance in the metal hyperaccumulator Arabidopsis halleri and plays a role in limiting oxidative stress in Arabidopsis spp. Plant Physiol. 2015, 169, 549–559. [Google Scholar] [CrossRef] [Green Version]
- Lanquar, V.; Lelièvre, F.; Bolte, S.; Hamès, C.; Alcon, C.; Neumann, D. Mobilization of vacuolar iron by AtNRAMP3 and AtNRAMP4 is essential for seed germination on low iron. EMBO J. 2005, 24, 4041–4051. [Google Scholar] [CrossRef]
- Boominathan, R.; Doran, P.M. Organic acid complexation, heavy metal distribution and the effect of ATPase inhibition in hairy roots of hyperaccumulator plant species. J. Biotechnol. 2003, 101, 131–146. [Google Scholar] [CrossRef]
- Sun, R.L.; Zhou, Q.X.; Jin, C.X. Cadmium accumulation in relation to organic acids in leaves of Solanum nigrum L. as a newly found cadmium hyperaccumulator. Plant Soil 2006, 285, 125–134. [Google Scholar] [CrossRef]
- Ehsan, S.; Ali, S.; Noureen, S.; Mahmood, K.; Farid, M.; Ishaque, W.; Shakoor, M.B.; Rizwan, M. Citric acid assisted phytoremediation of cadmium by Brassica napus L. Ecotoxicol. Environ. Saf. 2014, 106, 164–172. [Google Scholar] [CrossRef]
- Yang, H.; Yang, Z.M.; Zhou, L.X.; Wong, J.W. Ability of Agrogyron elongatum to accumulate the single metal of cadmium, copper, nickel and lead and root exudation of organic acids. J. Environ. Sci. 2001, 13, 368–375. [Google Scholar]
- Price, M.B.; Kong, D.; Okumoto, S. Inter-subunit interactions between Glutamate-Like Receptors in Arabidopsis. Plant Signal. Behav. 2013, 12, e27034. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Cadmium effect on plant metabolism and growth.

Figure 2.
Uptake and intracellular compartmentation of cadmium in plant roots. Cd taken up by IRT1/2 (zinc/iron-regulated transporter-like protein), YSL2/3 (yellow stripe-like protein), and NRAMP1/5/6 (natural resistance-associated macrophage protein) transporters that are located at the plasma membrane of the root epidermis. Cd can be also transported into cells through non-selective cation (NSCC) channels such as CNGCs (cyclic nucleotide-gated channels) and GLRs (glutamate receptors). ABCC (ATP-binding cassette transporters), CAX (cation exchanger), HMA3 (metal-transporting ATPases), MTP1 (metal tolerance protein), and NRAMP3/4 mediate Cd transport and sequestration in the vacuole.
Figure 2.
Uptake and intracellular compartmentation of cadmium in plant roots. Cd taken up by IRT1/2 (zinc/iron-regulated transporter-like protein), YSL2/3 (yellow stripe-like protein), and NRAMP1/5/6 (natural resistance-associated macrophage protein) transporters that are located at the plasma membrane of the root epidermis. Cd can be also transported into cells through non-selective cation (NSCC) channels such as CNGCs (cyclic nucleotide-gated channels) and GLRs (glutamate receptors). ABCC (ATP-binding cassette transporters), CAX (cation exchanger), HMA3 (metal-transporting ATPases), MTP1 (metal tolerance protein), and NRAMP3/4 mediate Cd transport and sequestration in the vacuole.

Figure 3.
Transporters mediating xylem (A) and phloem (B) cadmium loading. HMA2 and LCT1 (low-affinity calcium transporter) are functioning in the translocation of Cd into the phloem. The major transporter that regulates xylem Cd loading is HMA2/4. The role of MTP9 in the long-distance Cd transport remains unclear.
Figure 3.
Transporters mediating xylem (A) and phloem (B) cadmium loading. HMA2 and LCT1 (low-affinity calcium transporter) are functioning in the translocation of Cd into the phloem. The major transporter that regulates xylem Cd loading is HMA2/4. The role of MTP9 in the long-distance Cd transport remains unclear.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Cd content in leafy vegetable plants (selected examples).
| Leafy Vegetable Species | Range of Cd in Shoot (mg∙kg−1) | Mean or Range of Cd in Soil (mg∙kg−1) | Reference |
|---|---|---|---|
| Lactuca sativa L. | 5.8 to 9.1 | 0.12 to 0.31 | Baldantoni et al. [32] |
| Cichorium endivia L. | 0.61 to 3.80 | 0.13 to 0.51 | Baldantoni et al. [32] |
| Brassica pekinensis L. | 1.05 to 3.51 | 2.42 | Wang et al. [33] |
Table 2.
Transporters mediating cadmium uptake and translocation in plants.
| Transporter | Localization | Function/Substrate | Reference |
|---|---|---|---|
| OsNRAMP1 | Roots and shoots (PM) | Influx of Cd, Al | [42] |
| OsZIP8 | Root (PM) | Influx of Zn, Fe | [52] |
| OsHMA3 | Roots (tonoplast) | Cd sequestration in root vacuoles | [52] |
| SnYSL3 | Vascular tissues and epidermal cells of the roots and stems (PM) | Transport of nicotianamine complexes containing Fe(II), Cu, Zn, and Cd | [41] |
| OsYSL2 | Vascular bundles, Roots (PM) | Influx of nicotianamine complexes containing Fe(II), Mn, Ni, and Cd | [77] |
| OsIRT1 | Roots (PM) | Uptake of Fe, Zn, Mn, and Cd | [78] |
| OsIRT2 | Roots (PM) | Uptake of Fe, Zn, Mn, and Cd | [78] |
| OsNRAMP5 | Roots (PM) | Uptake of Mn and Cd | [82] |
| HvNRAMP5 | Roots (PM) | Uptake of Mn and Cd | [83] |
| OsNRAMP3 | Vascular bundles, roots, leaves (tonoplast) | Uptake of Mn | [85] |
| AtNRAMP3 | Vascular bundles, roots, leaves (tonoplast) | Efflux of Fe and Cd | [87,88] |
| AtNRAMP4 | Vascular bundles, roots, leaves (tonoplast) | Efflux of Fe and Cd | [87,88] |
| AtNRAMP6 | Roots, young leaves (PM) | Influx of Mn, | [40] |
| AtCNGCs | Roots (PM) | Transporter for multiple cations | [89,90,91,92] |
| AtGLRs | Roots (PM) | Transporter for multiple cations | [89,90,93,94,95,96] |
| AtMTP1 | Roots and leaves (tonoplast) | Transporter for Zn and Cd | [97,98,99,100] |
| CsMTP9 | Roots endodermal cells (PM) | Efflux of Mn and Cd | [101] |
| OsMTP9 | Roots (PM) | Efflux of Mn | [102] |
| OsMTP8.1 | Roots (tonoplast) | Sequestration of Mn into vacuoles | [103] |
| TgMTP1 | Roots and leaves (tonoplast) | Transporter for Zn and Cd | [104] |
| OsHMA5 | Roots, vascular bundles (tonoplast) | Loading of Cu in xylem | [105] |
| AtHMA2 | Roots, vascular tissue (PM) | Delivery of Zn and Cd to xylem | [106,107] |
| AtHMA4 | Roots, vascular tissue (PM) | Delivery of Zn and Cd to xylem | [106,107] |
| OsLCT1 | Leaves, nodes, phloem parenchyma (PM) | Efflux of Cd, Ca, Mg, and Mn | [74,108] |
| AtABCC1 | Roots and shoots (tonoplast) | Uptake of PCs | [109] |
| AtABCC2 | Roots and shoots (tonoplast) | Uptake of PCs | [109] |
| AtABCC3 | Roots and shoots (tonoplast) | Uptake of PCs | [110] |
| AhHMA3 | Roots, shoots (tonoplast) | Sequestration of Zn into vacuoles | [111] |
| AtHMA3 | Vascular tissues (tonoplast) | Transport of Zn, Co, Pb, and Cd | [112,113] |
| SpHMA3 | Roots, shoots (tonoplast) | Sequestration of Cd into vacuoles | [114] |
| AtCAX2 | Roots (tonoplast) | Vacuolar Cd, Zn, and Mn transport | [115,116] |
| AtCAX4 | Roots (tonoplast) | Vacuolar Cd, Zn, and Mn transport | [115,116] |
Abbreviations: PM, plasma membrane.
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Huang, X.; Duan, S.; Wu, Q.; Yu, M.; Shabala, S. Reducing Cadmium Accumulation in Plants: Structure–Function Relations and Tissue-Specific Operation of Transporters in the Spotlight. Plants 2020, 9, 223. https://doi.org/10.3390/plants9020223
AMA Style
Huang X, Duan S, Wu Q, Yu M, Shabala S. Reducing Cadmium Accumulation in Plants: Structure–Function Relations and Tissue-Specific Operation of Transporters in the Spotlight. Plants. 2020; 9(2):223. https://doi.org/10.3390/plants9020223
Chicago/Turabian StyleHuang, Xin, Songpo Duan, Qi Wu, Min Yu, and Sergey Shabala. 2020. "Reducing Cadmium Accumulation in Plants: Structure–Function Relations and Tissue-Specific Operation of Transporters in the Spotlight" Plants 9, no. 2: 223. https://doi.org/10.3390/plants9020223
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.