Mutagenic Effect of Three Ion Beams on Rice and Identification of Heritable Mutations by Whole Genome Sequencing

 , and
, and
Abstract
:1. Introduction
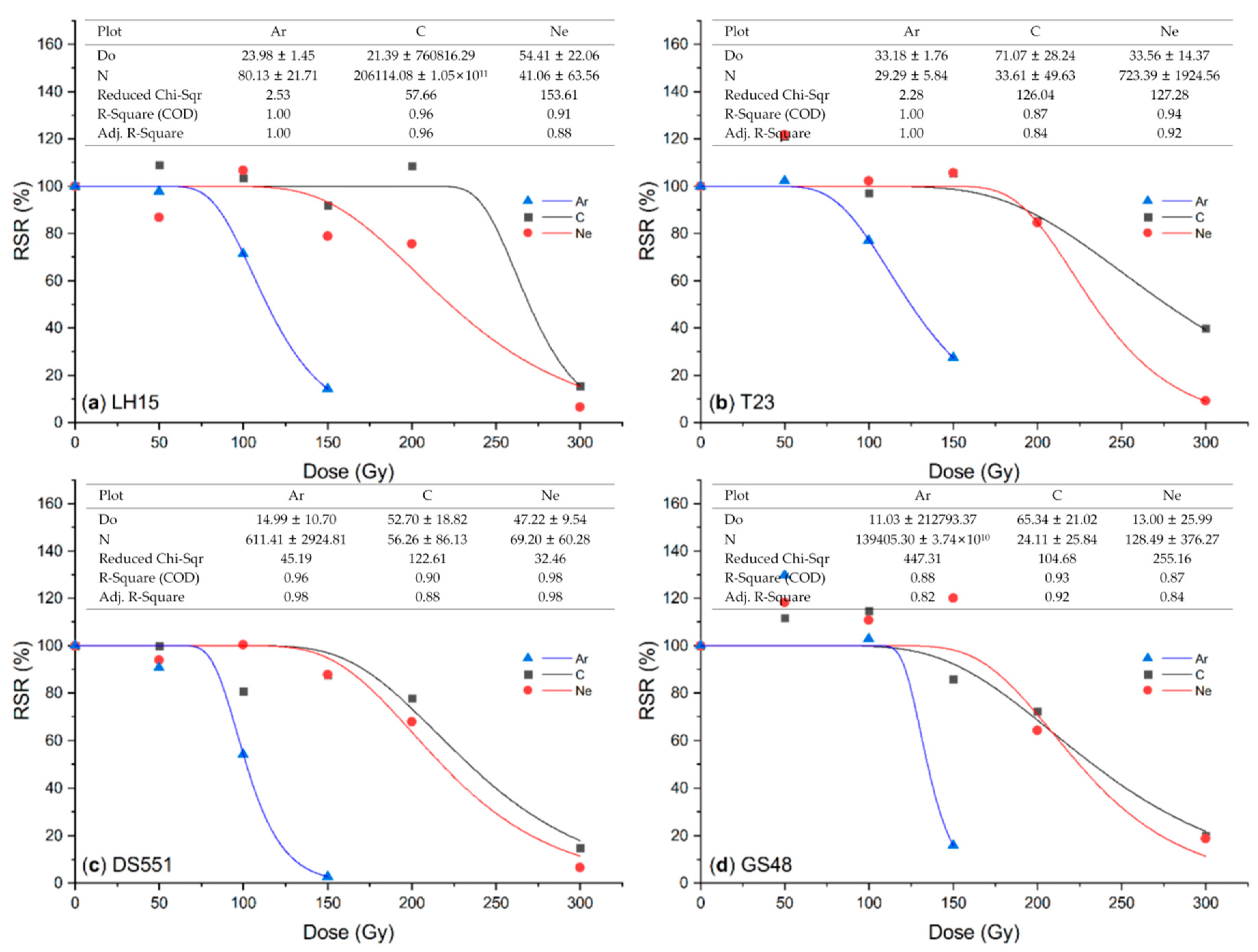
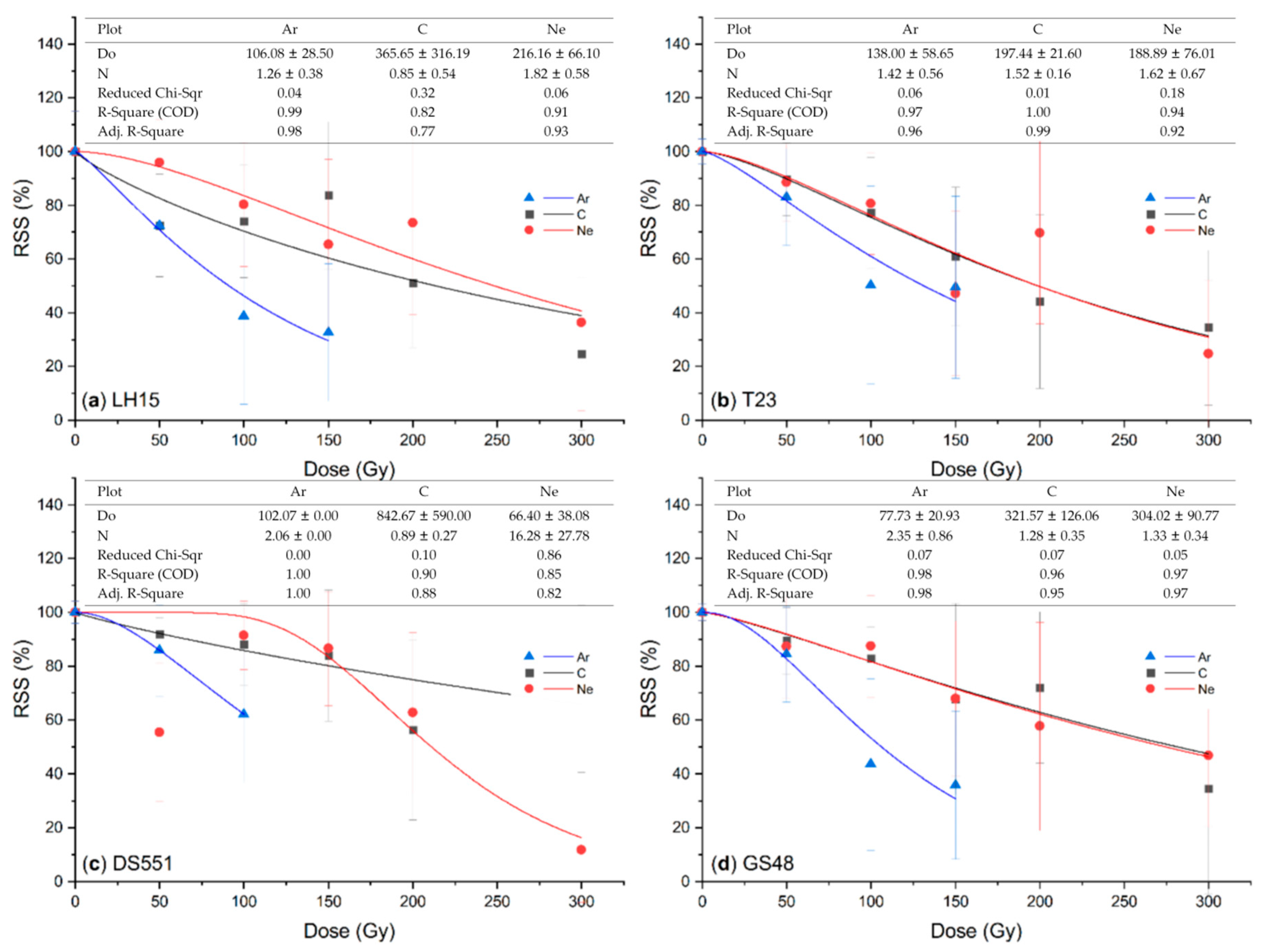
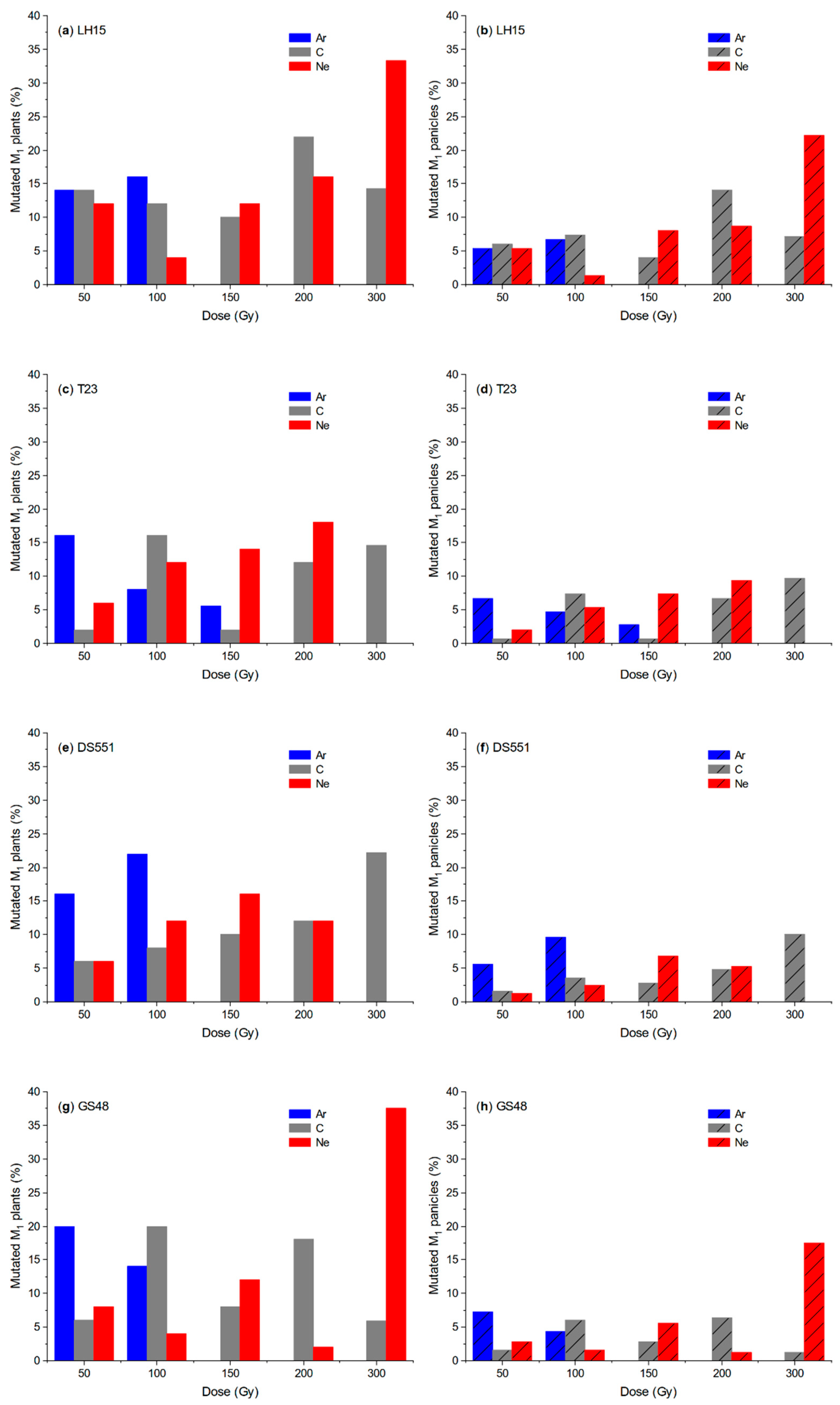
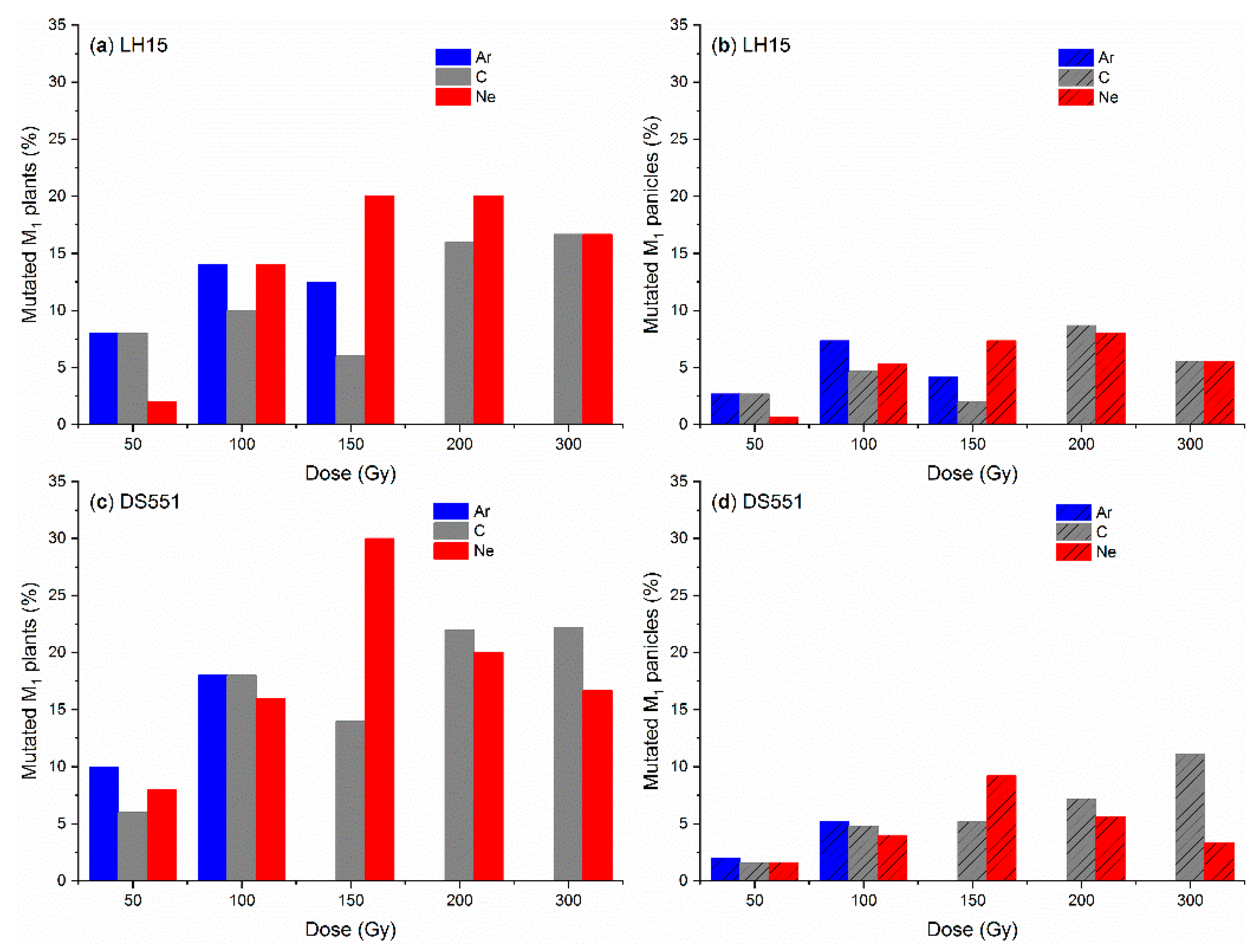
2. Results
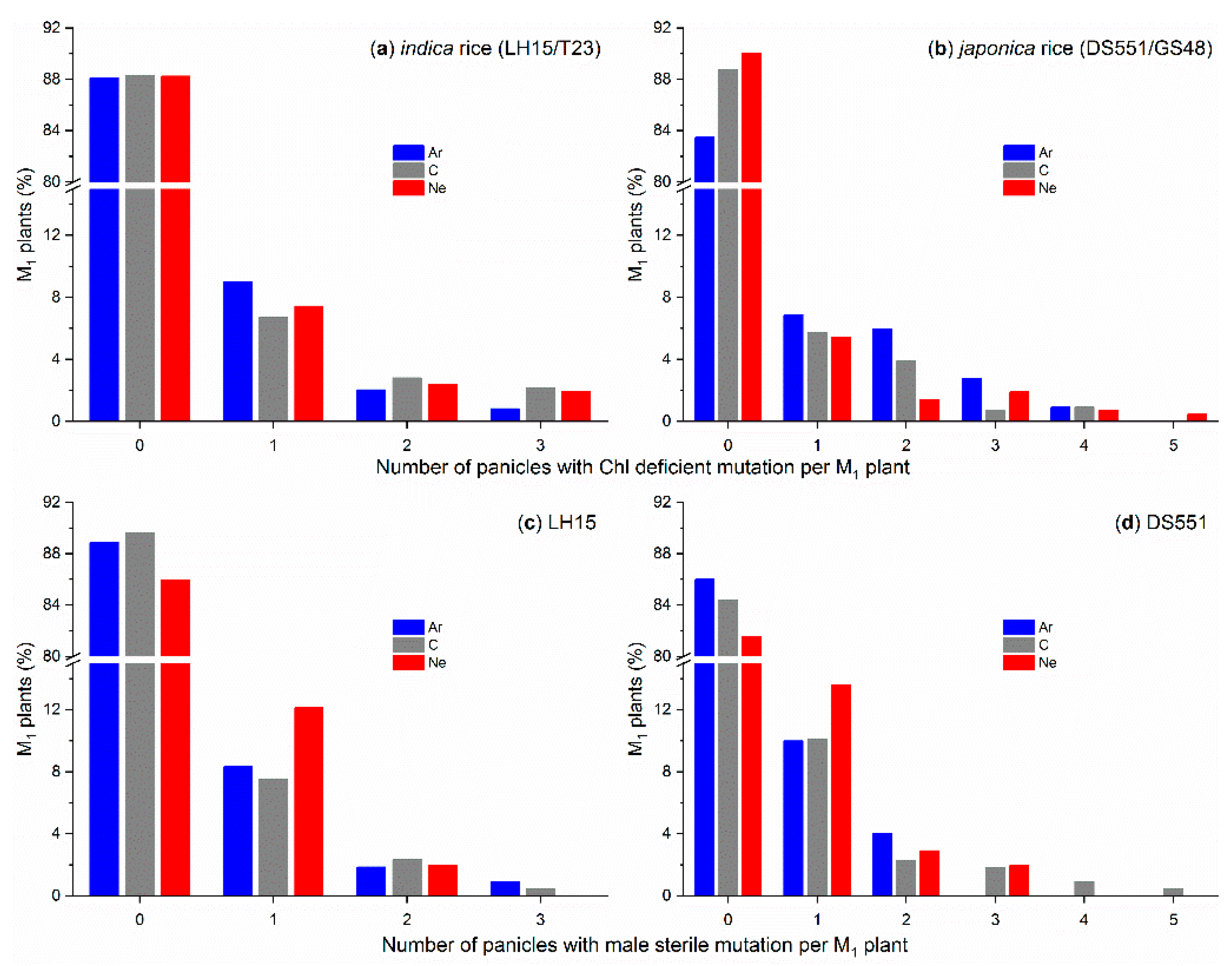
2.1. Effect of Ion Beam Radiation on M1 Plants
2.2. Morphological Mutants in M2 Populations
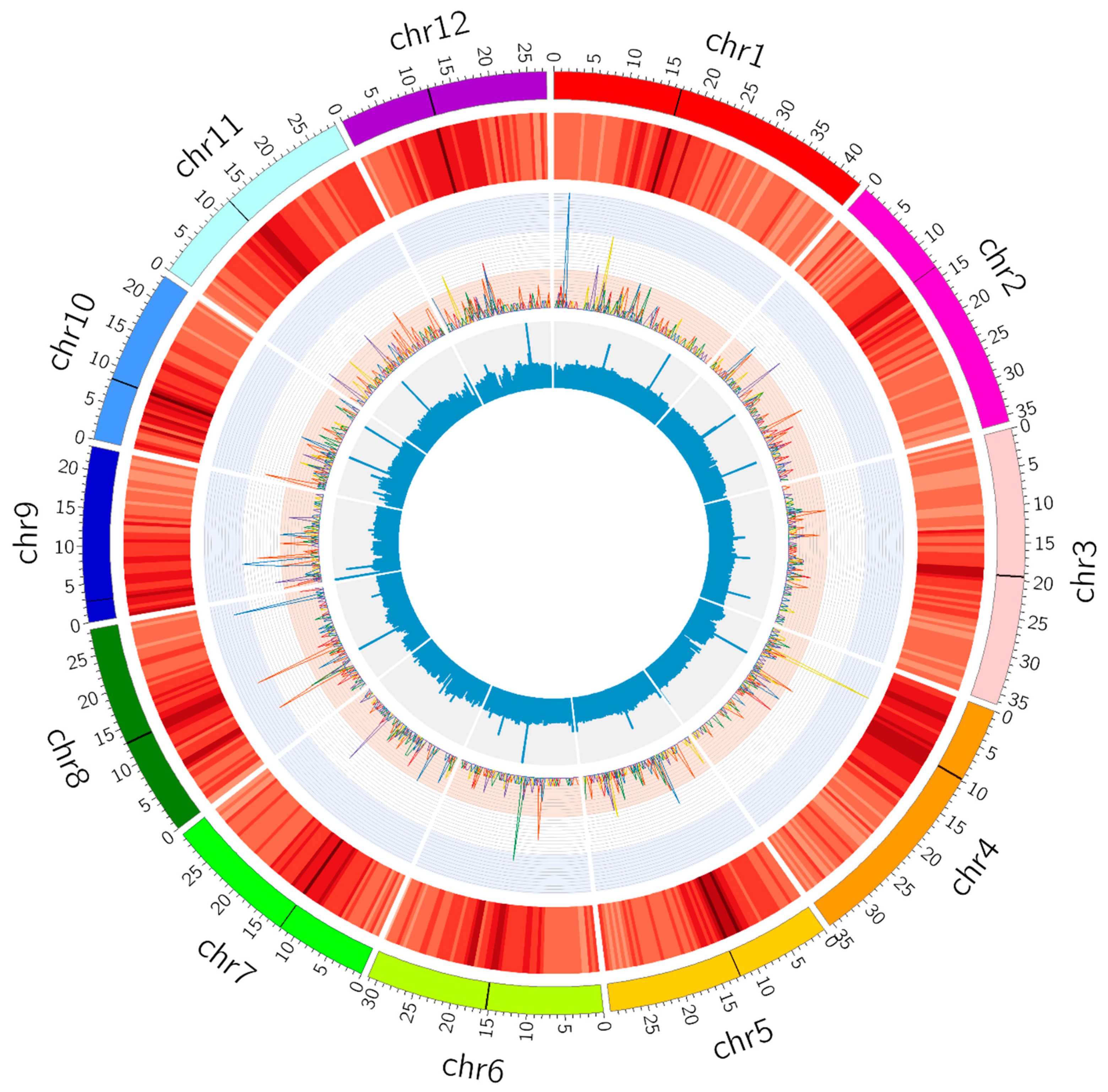
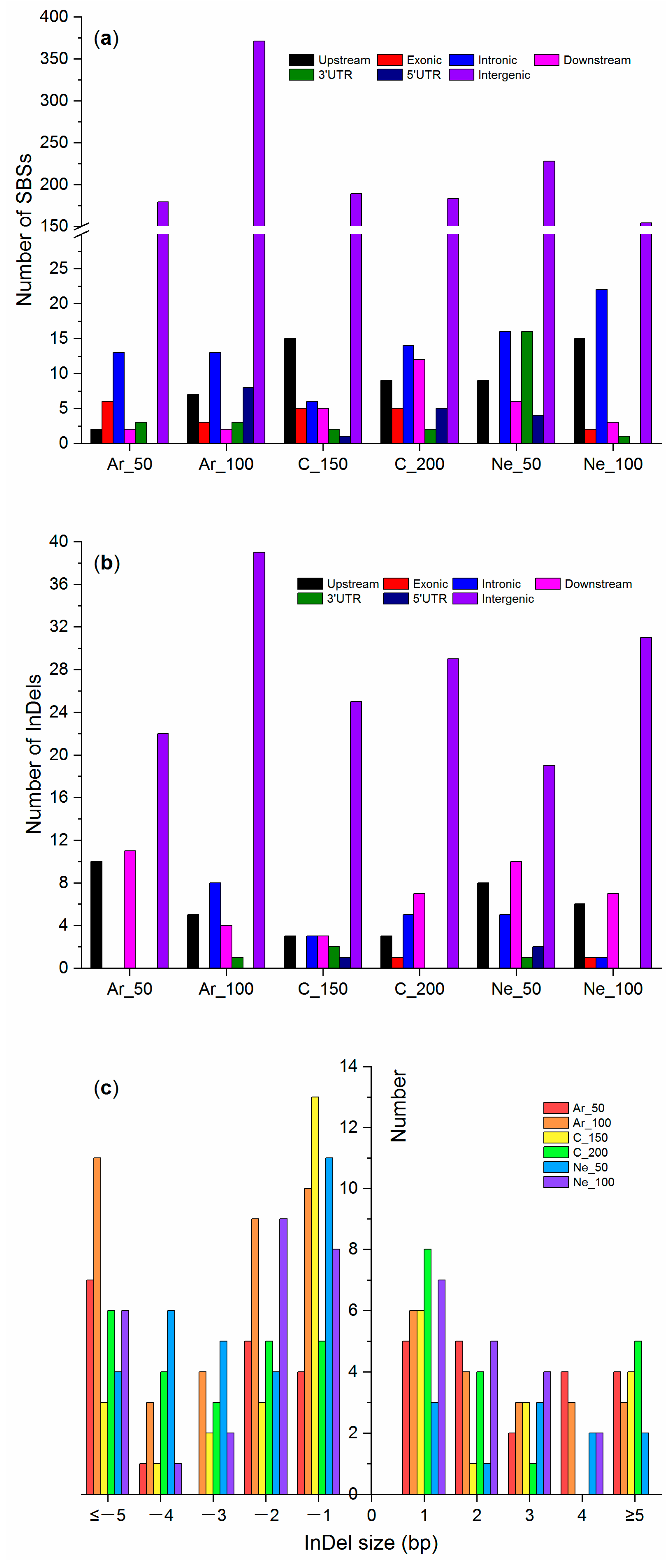
2.3. Genome-wide Mutations in M4 Plants
3. Discussion
3.1. Ar, C and Ne Ion Beams Could All Be Used for Mutation Induction and Breeding in Rice
3.2. How to Make Full Use of Irradiated Seeds?
3.3. Mutations Identified via Genome/Exome Sequencing
4. Materials and Methods
4.1. Irradiation of Ion Beams and Mutant Development
4.2. Genome Sequencing
4.3. Mutation Detection and Annotation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Abe, T.; Ryuto, H.; Fukunishi, N. Ion beam radiation mutagenesis. In Plant Mutation Breeding and Biotechnology, 1st ed.; Shu, Q.Y., Forster, B.P., Nakagawa, H., Eds.; Plant Breeding and Genetics Section, Joint FAO/IAEA Division of Nuclear Techniques in Food and Agriculture, International Atomic Energy Agency: Vienna, Austria, 2012; Chapter 9; pp. 99–106. [Google Scholar]
- Tanaka, A.; Shikazono, N.; Hase, Y. Studies on biological effects of ion beams on lethality, molecular nature of mutation, mutation rate, and spectrum of mutation phenotype for mutation breeding in higher plants. J. Radiat. Res. 2010, 51, 223–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaguchi, H.; Hase, Y.; Tanaka, A.; Shikazono, N.; Degi, K.; Shimizu, A.; Morishita, T. Mutagenic effects of ion beam irradiation on rice. Breed. Sci. 2009, 59, 169–177. [Google Scholar] [CrossRef] [Green Version]
- Ishikawa, S.; Ishimaru, Y.; Igura, M.; Kuramata, M.; Abe, T.; Senoura, T.; Hase, Y.; Arao, T.; Nishizawa, N.K.; Nakanishi, H. Ion-beam irradiation, gene identification, and marker-assisted breeding in the development of low-cadmium rice. Proc. Natl. Acad. Sci. USA 2012, 109, 19166–19171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takano, N.; Takahashi, Y.; Yamamoto, M.; Teranishi, M.; Yamaguchi, H.; Sakamoto, A.N.; Hase, Y.; Fujisawa, H.; Wu, J.; Matsumoto, T.; et al. Isolation of a novel UVB-tolerant rice mutant obtained by exposure to carbon-ion beams. J. Radiat. Res. 2013, 54, 637–648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ichitani, K.; Yamaguchi, D.; Taura, S.; Fukutoku, Y.; Onoue, M.; Shimizu, K.; Hashimoto, F.; Sakata, Y.; Sato, M. Genetic analysis of ion-beam induced extremely late heading mutants in rice. Breed. Sci. 2014, 64, 222–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sevanthi, A.M.V.; Kandwal, P.; Kale, P.B.; Prakash, C.; Ramkumar, M.K.; Yadav, N.; Mahato, A.K.; Sureshkumar, V.; Behera, M.; Deshmukh, R.K.; et al. Whole genome characterization of a few EMS-induced mutants of upland rice variety Nagina 22 reveals a staggeringly high frequency of SNPs which show high phenotypic plasticity towards the wild-type. Front. Plant Sci. 2018, 9, 1179. [Google Scholar] [CrossRef]
- Hussain, M.; Iqbal, M.A.; Till, B.J.; Rahman, M.U. Identification of induced mutations in hexaploid wheat genome using exome capture assay. PLoS ONE 2018, 13, e0201918. [Google Scholar] [CrossRef] [Green Version]
- Li, W.J.; Guo, H.J.; Wang, Y.B.; Xie, Y.D.; Zhao, L.S.; Gu, J.Y.; Zhao, S.R.; Zhao, B.C.; Wang, G.J.; Liu, L.X. Identification of novel alleles induced by EMS-mutagenesis in key genes of kernel hardness and starch biosynthesis in wheat by TILLING. Genes Genom. 2017, 39, 387–395. [Google Scholar] [CrossRef]
- Sidhu, G.; Mohan, A.; Zheng, P.; Dhaliwal, A.K.; Main, D.; Gill, K.S. Sequencing-based high throughput mutation detection in bread wheat. BMC Genomics 2015, 16, 962. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Luu, N.S.; Chen, Z.; Chen, B.; Cui, X.; Wu, J.; Zhang, Z.; Lu, T. Generation and characterization of a Foxtail Millet (Setaria italica) mutant library. Front. Plant Sci. 2019, 10, 369. [Google Scholar] [CrossRef] [Green Version]
- Mestanza, C.; Riegel, R.; Vásquez, S.C.; Veliz, D.; Cruz-Rosero, N.; Canchignia, H.; Silva, H. Discovery of mutations in Chenopodium quinoa Willd through EMS mutagenesis and mutation screening using pre-selection phenotypic data and next-generation sequencing. J. Agric. Sci. 2018, 156, 1196–1204. [Google Scholar] [CrossRef]
- Jiao, Y.; Burke, J.; Chopra, R.; Burow, G.; Chen, J.; Wang, B.; Hayes, C.; Emendack, Y.; Ware, D.; Xin, Z. A sorghum mutant resource as an efficient platform for gene discovery in grasses. Plant Cell 2016, 28, 1551–1562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shirasawa, K.; Hirakawa, H.; Nunome, T.; Tabata, S.; Isobe, S. Genome-wide survey of artificial mutations induced by ethyl methanesulfonate and gamma rays in tomato. Plant Biotechnol. J. 2016, 14, 51–60. [Google Scholar] [CrossRef] [PubMed]
- García, A.; Aguado, E.; Parra, G.; Manzano, S.; Martínez, C.; Megías, Z.; Cebrián, G.; Romero, J.; Beltrán, S.; Garrido, D.; et al. Phenomic and genomic characterization of a mutant platform in Cucurbita pepo. Front. Plant Sci. 2018, 9, 1049. [Google Scholar] [CrossRef] [PubMed]
- Elhiti, M.; Wang, H.Y.; Austin, R.S.; Chen, B.; Brown, D.; Wang, A.M. Generation of chemically induced mutations using in vitro propagated shoot tip tissues for genetic improvement of fruit trees. Plant Cell Tissue Organ Cult. 2016, 124, 447–452. [Google Scholar] [CrossRef]
- Mohd-Yusoff, N.F.; Ruperao, P.; Tomoyoshi, N.E.; Edwards, D.; Gresshoff, P.M.; Biswas, B.; Batley, J. Scanning the effects of ethyl methanesulfonate on the whole genome of Lotus japonicus using second-generation sequencing analysis. G3 (Bethesda) 2015, 5, 559–567. [Google Scholar] [CrossRef] [Green Version]
- Maghuly, F.; Pabinger, S.; Krainer, J.; Laimer, M. The pattern and distribution of induced mutations in J. curcas using reduced representation sequencing. Front. Plant Sci. 2018, 9, 524. [Google Scholar] [CrossRef]
- Li, S.; Zheng, Y.C.; Cui, H.R.; Fu, H.W.; Shu, Q.Y.; Huang, J.Z. Frequency and type of inheritable mutations induced by γ rays in rice as revealed by whole genome sequencing. J. Zhejiang Univ. Sci. B 2016, 17, 905–915. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Li, S.; Huang, J.; Fan, L.; Shu, Q. Identification and characterization of γ-ray-induced mutations in rice cytoplasmic genomes by whole-genome sequencing. Cytogenet. Genome Res. 2020. [Google Scholar] [CrossRef]
- Henry, I.M.; Zinkgraf, M.S.; Groover, A.T.; Comai, L. A system for dosage-based functional genomics in poplar. Plant Cell 2015, 27, 2370–2383. [Google Scholar] [CrossRef] [Green Version]
- Belfield, E.J.; Gan, X.; Mithani, A.; Brown, C.; Jiang, C.; Franklin, K.; Alvey, E.; Wibowo, A.; Jung, M.; Bailey, K.; et al. Genome-wide analysis of mutations in mutant lineages selected following fast-neutron irradiation mutagenesis of Arabidopsis thaliana. Genome Res. 2012, 22, 1306–1315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, G.T.; Chern, M.S.; Jain, R.; Martin, J.A.; Schackwitz, W.S.; Jiang, L.; Vega-Sánchez, M.E.; Lipzen, A.M.; Barry, K.W.; Schmutz, J.; et al. Genome-wide sequencing of 41 rice (Oryza sativa L.) mutated lines reveals diverse mutations induced by fast-neutron irradiation. Mol. Plant 2016, 9, 1078–1081. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Luo, S.; Li, X.; Yang, J.; Cui, T.; Li, W.; Yu, L.; Feng, H.; Chen, Y.; Mu, J.; et al. Identification of substitutions and small insertion-deletions induced by carbon-ion beam irradiation in Arabidopsis thaliana. Front. Plant Sci. 2017, 8, 1851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, Y.; Luo, S.W.; Yu, L.X.; Cui, T.; Chen, X.; Yang, J.Y.; Li, X.; Li, W.J.; Wang, J.F.; Zhou, L.B. Strategies for identification of mutations induced by carbon-ion beam irradiation in Arabidopsis thaliana by whole genome re-sequencing. Mutat. Res. 2018, 807, 21–30. [Google Scholar] [CrossRef]
- Hase, Y.; Satoh, K.; Kitamura, S.; Oono, Y. Physiological status of plant tissue affects the frequency and types of mutations induced by carbon-ion irradiation in Arabidopsis. Sci. Rep. 2018, 8, 1394. [Google Scholar] [CrossRef] [Green Version]
- Hirano, T.; Kazama, Y.; Ishii, K.; Ohbu, S.; Shirakawa, Y.; Abe, T. Comprehensive identification of mutations induced by heavy-ion beam irradiation in Arabidopsis thaliana. Plant J. 2015, 82, 93–104. [Google Scholar] [CrossRef]
- Kazama, Y.; Ishii, K.; Hirano, T.; Wakana, T.; Yamada, M.; Ohbu, S.; Abe, T. Different mutational function of low- and high-linear energy transfer heavy-ion irradiation demonstrated by whole-genome resequencing of Arabidopsis mutants. Plant J. 2017, 92, 1020–1030. [Google Scholar] [CrossRef] [Green Version]
- Ichida, H.; Morita, R.; Shirakawa, Y.; Hayashi, Y.; Abe, T. Targeted exome sequencing of unselected heavy-ion beam-irradiated populations reveals less-biased mutation characteristics in the rice genome. Plant J. 2019, 98, 301–314. [Google Scholar] [CrossRef]
- Li, F.; Shimizu, A.; Nishio, T.; Tsutsumi, N.; Kato, H. Comparison and characterization of mutations induced by gamma-ray and carbon-ion irradiation in rice (Oryza sativa L.) using whole-genome resequencing. G3 (Bethesda) 2019, 9, 3743–3751. [Google Scholar] [CrossRef] [Green Version]
- Yang, G.; Luo, W.; Zhang, J.; Yan, X.; Du, Y.; Zhou, L.; Li, W.; Wang, H.; Chen, Z.; Guo, T. Genome-wide comparisons of mutations induced by carbon-ion beam and gamma-rays irradiation in rice via resequencing multiple mutants. Front. Plant Sci. 2019, 10, 1514. [Google Scholar] [CrossRef] [Green Version]
- Feng, H.Y.; Yu, Z.L. Ion implantation mutagenesis. In Plant Mutation Breeding and Biotechnology, 1st ed.; Shu, Q.Y., Forster, B.P., Nakagawa, H., Eds.; Plant Breeding and Genetics Section, Joint FAO/IAEA Division of Nuclear Techniques in Food and Agriculture, International Atomic Energy Agency: Vienna, Austria, 2012; Chapter 10; pp. 107–122. [Google Scholar]
- Matsumoto, Y.; Furusawa, Y.; Aoki-Nakano, M.; Matsufuji, N.; Hirayama, R.; Kanai, T.; Ando, K.; Sakurai, H. Estimation of RBE values for carbon-ion beams in the wide dose range using multicellular spheroids. Radiat. Prot. Dosim. 2019, 183, 45–49. [Google Scholar] [CrossRef] [PubMed]
- Robinson, J.T.; Thorvaldsdóttir, H.; Winckler, W.; Guttman, M.; Lander, E.S.; Getz, G.; Mesirov, J.P. Integrative genomics viewer. Nat. Biotechnol. 2011, 29, 24–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, R.K.; Jain, M. NGS QC Toolkit: A toolkit for quality control of next generation sequencing data. PLoS ONE 2012, 7, e30619. [Google Scholar] [CrossRef] [PubMed]
- The Reference Nipponbare Genome. Available online: ftp://ftp.ensemblgenomes.org/pub/plants/release-41/fasta/oryza_sativa/dna/Oryza_sativa.IRGSP-1.0.dna.toplevel.fa.gz (accessed on 24 April 2020).
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. 1000 Genome Project Data Processing Subgroup. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Picard Toolkit, Broad Institute, GitHub Repository. Available online: http://broadinstitute.github.io/picard/ (accessed on 23 March 2020).
- Calculation of the Coverage and Average Depth. Available online: https://github.com/zhengyunchao/Inheritable_Mutations_Induced_by_Ion_Beams_in_Rice/blob/master/Calculation_of_the_coverage_and_average_depth.pl (accessed on 24 April 2020).
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A.; Garimella, K.; Altshuler, D.; Gabriel, S.; Daly, M.; et al. The Genome Analysis Toolkit: A MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koboldt, D.C.; Zhang, Q.; Larson, D.E.; Shen, D.; McLellan, M.D.; Lin, L.; Miller, C.A.; Mardis, E.R.; Ding, L.; Wilson, R.K. VarScan 2: Somatic mutation and copy number alteration discovery in cancer by exome sequencing. Genome Res. 2012, 22, 568–576. [Google Scholar] [CrossRef] [Green Version]
- Removal of Common Variants in Any Two Mutants. Available online: https://github.com/zhengyunchao/Heritable_Mutations_Induced_by_Ion_Beams_in_Rice/blob/master/Removal_of_common_variants_in_any_two_mutants.pl (accessed on 24 April 2020).
- Filtration of Variants Called by GATK. Available online: https://github.com/zhengyunchao/Heritable_Mutations_Induced_by_Ion_Beams_in_Rice/blob/master/Filtration_of_Variants_Called_by_GATK.pl (accessed on 24 April 2020).
- Filtration of Variants Called by SAMtools. Available online: https://github.com/zhengyunchao/Heritable_Mutations_Induced_by_Ion_Beams_in_Rice/blob/master/Filtration_of_Variants_Called_by_SAMtools.pl (accessed on 24 April 2020).
- Filtration of Variants Called by VarScan2. Available online: https://github.com/zhengyunchao/Heritable_Mutations_Induced_by_Ion_Beams_in_Rice/blob/master/Filtration_of_Variants_Called_by_VarScan2.pl (accessed on 24 April 2020).
- Wang, K.; Li, M.; Hakonarson, H. ANNOVAR: Functional annotation of genetic variants from next-generation sequencing data. Nucleic Acids Res. 2010, 38, e164. [Google Scholar] [CrossRef]
- Krzywinski, M.; Schein, J.; Birol, I.; Connors, J.; Gascoyne, R.; Horsman, D.; Jones, S.J.; Marra, M.A. Circos: An information aesthetic for comparative genomics. Genome Res. 2009, 19, 1639–1645. [Google Scholar] [CrossRef] [Green Version]
- Viewing Alignments in the Integrative Genomics Viewer (IGV). Available online: http://software.broadinstitute.org/software/igv/AlignmentData (accessed on 24 April 2020).









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Material 1 | No. of Mapped Reads | Mapping Rate (%) | Genome Coverage (%) | Sequencing Depth (×) |
|---|---|---|---|---|
| DS551 | 135,748,093 | 98.79 | 97.30 | 53.46 |
| Ar_50 | 153,313,517 | 95.63 | 97.37 | 59.82 |
| Ar_100 | 169,292,288 | 98.47 | 97.41 | 66.52 |
| C_150 | 143,348,549 | 98.29 | 97.29 | 55.29 |
| C_200 | 154,943,939 | 98.18 | 97.30 | 61.15 |
| Ne_50 | 161,256,389 | 98.67 | 97.34 | 63.27 |
| Ne_100 | 164,791,769 | 98.16 | 97.34 | 64.84 |
| Mutant 1 | Total | Single Base Substitution (SBS) | Insertion/Deletion (InDel) | ||
|---|---|---|---|---|---|
| Subtotal (Ti/Tv) 2 | Ex. (Syn/Nonsyn /Stop-Gain/Loss) 3 | Subtotal | Ex. (Frame-/Nonframe -Shift/Stop-Gain/Loss) 3 | ||
| Ar_50 | 236 | 205 (141/64) | 6 (2/4/0/0) | 31 | 0 (0/0/0/0) |
| Ar_100 | 453 | 404 (297/107) | 2 (1/1/0/0) | 49 | 0 (0/0/0/0) |
| C_150 | 256 | 223 (139/84) | 5 (1/4/0/0) | 33 | 0 (0/0/0/0) |
| C_200 | 269 | 228 (145/83) | 5 (1/4/0/0) | 41 | 1 (1/0/0/0) |
| Ne_50 | 307 | 275 (156/119) | 0 (0/0/0/0) | 32 | 0 (0/0/0/0) |
| Ne_100 | 238 | 197 (128/69) | 2 (2/0/0/0) | 41 | 1 (1/0/0/0) |
| Ion Beam | Energy (MeV/u) | LET 1 (keV/μm) | Doses (Gy) | Dose Rate (Gy/Min.) |
|---|---|---|---|---|
| 40Ar18+ | 500 | 92 | 50, 100, 150, 200, 300 | 7.1 |
| 12C6+ | 290 | 13 | 50, 100, 150, 200, 300 | 7.7 |
| 20Ne10+ | 400 | 31 | 50, 100, 150, 200, 300 | 5.5 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, Y.; Li, S.; Huang, J.; Fu, H.; Zhou, L.; Furusawa, Y.; Shu, Q. Mutagenic Effect of Three Ion Beams on Rice and Identification of Heritable Mutations by Whole Genome Sequencing. Plants 2020, 9, 551. https://doi.org/10.3390/plants9050551
Zheng Y, Li S, Huang J, Fu H, Zhou L, Furusawa Y, Shu Q. Mutagenic Effect of Three Ion Beams on Rice and Identification of Heritable Mutations by Whole Genome Sequencing. Plants. 2020; 9(5):551. https://doi.org/10.3390/plants9050551
Chicago/Turabian StyleZheng, Yunchao, Shan Li, Jianzhong Huang, Haowei Fu, Libin Zhou, Yoshiya Furusawa, and Qingyao Shu. 2020. "Mutagenic Effect of Three Ion Beams on Rice and Identification of Heritable Mutations by Whole Genome Sequencing" Plants 9, no. 5: 551. https://doi.org/10.3390/plants9050551