A Study on the Effect of Macro- and Micro- Nutrients on Nannochloropsis oceanica Growth, Fatty Acid Composition and Magnetic Harvesting Efficiency

 , ,
, ,  , , ,
, , ,
Abstract
:1. Introduction
2. Results and Discussion
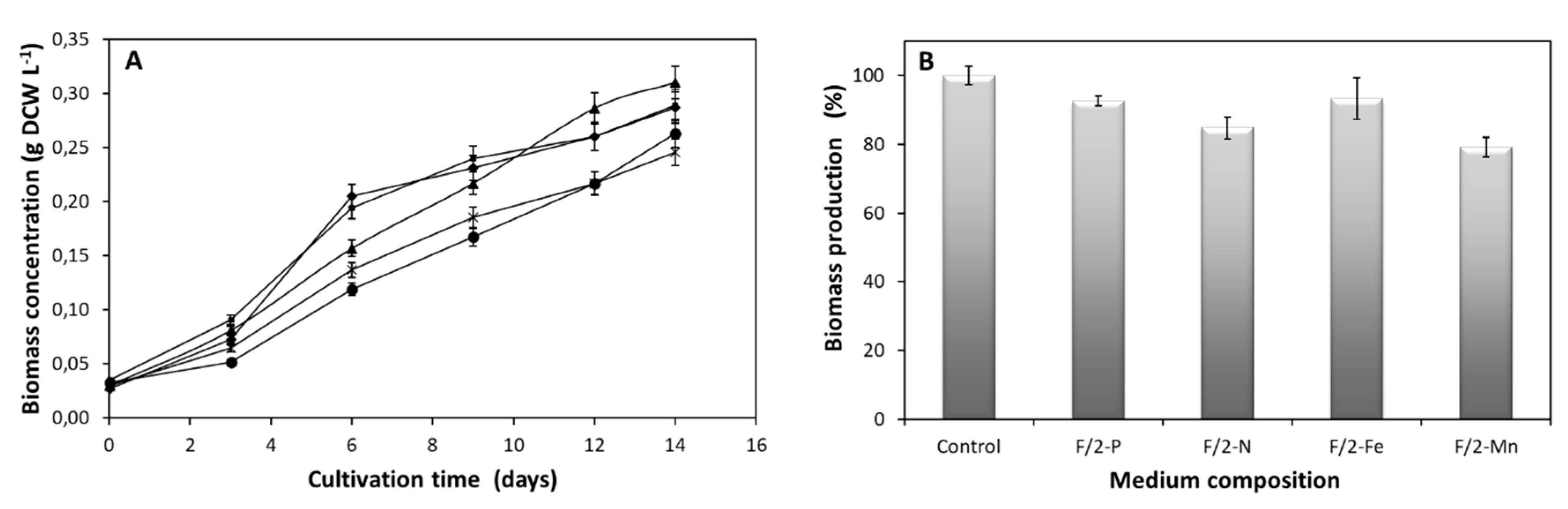
2.1. Effect of Macro- and Micro-Elements on N. oceanica Growth, Lipid Biosynthesis and Chlorophyll Content
2.2. Effect of Nutrients on Fatty Acid Profile
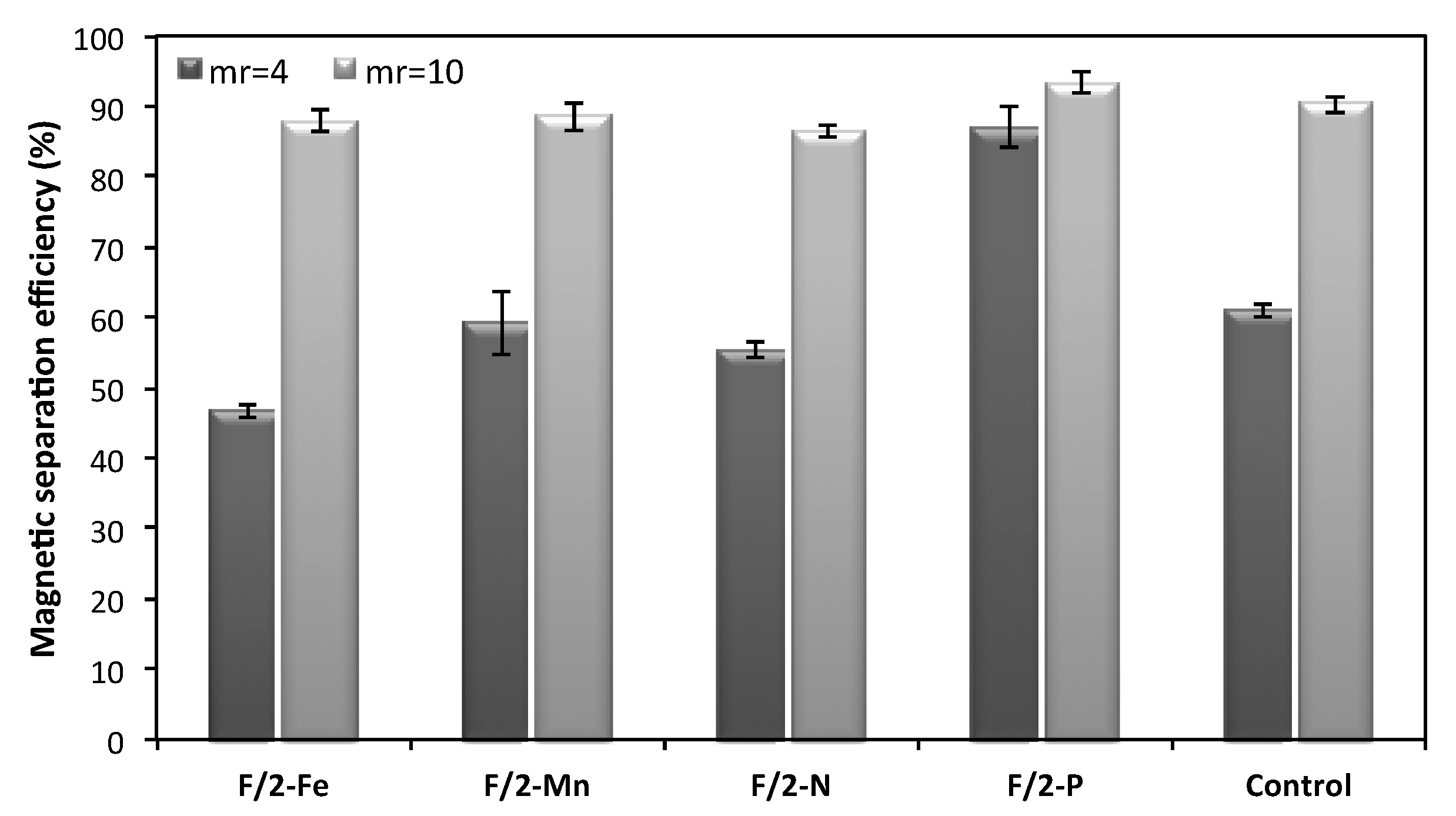
2.3. Effect of Culture Media Composition on Magnetic Harvesting Efficiency
3. Materials and Methods
3.1. Organism and Growth Conditions
3.2. Microalgae Dry Weight
3.3. Extraction and Quantification of Chlorophyll
3.4. Determination of Lipid Content and Lipid Composition
3.5. Kinetic and Yield Parameters
3.6. Magnetic Separation of Nannochloropsis Oceanica Microalgal Biomass
3.7. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- da Silva Ferreira, V.; Sant'Anna, C. The effect of physicochemical conditions and nutrient sources on maximizing the growth and lipid productivity of green microalgae. Phycol. Res. 2017, 65, 3–13. [Google Scholar] [CrossRef] [Green Version]
- Dou, X.; Lu, X.-H.; Lu, M.-Z.; Yu, L.-S.; Xue, R.; Ji, J.-B. The effects of trace elements on the lipid productivity and fatty acid composition of Nannochloropis oculata. J. Renew. Energy 2013, 2013, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, S.B.; Soren, H. The biochemical properties of manganese in plants. Plants 2019, 8, 381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peloquin, J.M.; Britt, R.D. EPR/ENDOR characterization of the physical and electronic structure of the OEC Mn cluster. Biochim. Biophys. Acta 2001, 1503, 96–111. [Google Scholar] [CrossRef] [Green Version]
- Rizwan, M.; Mujtaba, G.; Lee, K. Effects of iron sources on the growth and lipid/carbohydrate production of marine microalga Dunaliella tertiolecta. Biotechnol. Bioprocess. Eng. 2017, 22, 68–75. [Google Scholar] [CrossRef]
- Lourenço, S.O.; Barbarino, E.; Lavín, P.L.; Lanfer Marquez, U.M.; Aidar, E. Distribution of intracellular nitrogen in marine microalgae: Calculation of new nitrogen-to-protein conversion factors. Eur. J. Phycol. 2004, 39, 17–32. [Google Scholar] [CrossRef]
- Ryckebosch, E.; Bruneel, C.; Muylaert, K.; Foubert, I. Microlagae as an alternative source of omega-3-long chain polyunsaturated fatty acids. Lipid Technol. 2012, 24, 128–130. [Google Scholar] [CrossRef]
- Mata, T.M.; Almeidab, R.; Caetanoa, N.S. Effect of the culture nutrients on the biomass and lipid productivities of microalgae Dunaliella tertiolecta. Chem. Eng. 2013, 32, 973. [Google Scholar]
- Chen, M.; Tang, H.; Ma, H.; Holland, T.C.; Ng, K.S.; Salley, S.O. Effect of nutrients on growth and lipid accumulation in the green algae Dunaliella tertiolecta. Bioresour. Technol. 2011, 102, 1649–1655. [Google Scholar] [CrossRef]
- Battah, M.; El-Ayoty, Y.; Abomohra, A.E.F.; El-Ghany, S.A.; Esmael, A. Effect of Mn2+, Co2+ and H2O2 on biomass and lipids of the green microalga Chlorella vulgaris as a potential candidate for biodiesel production. Ann. Microbiol. 2014, 65, 155–162. [Google Scholar] [CrossRef]
- Zienkiewicz, A.; Zienkiewicz, K.; Poliner, E.; Pulman, J.A.; Du, Z.Y.; Stefano, G.; Tsai, C.H.; Horn, P.; Feussner, I.; Farre, E.M.; et al. The Microalga Nannochloropsis during transition from quiescence to autotrophy in response to nitrogen availability. Plant. Physiol. 2020, 182, 819–839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merzlyak, M.N.; Chivkunova, O.B.; Gorelova, O.A.; Reshetnikova, I.V.; Solovchenko, A.E.; Khozin Goldberg, I.; Cohen, Z. Effect of nitrogen starvation on optical properties, pigments, and arachidonic acid content of the unicellular green alga Parietochlorisincisa (Trebouxiophyceae, Chlorophyta). J. Phycol. 2007, 43, 833–843. [Google Scholar] [CrossRef]
- Satpati, G.G.; Gorain, P.C.; PaL, R. Efficacy of EDTA and phosphorous on biomass yield and total lipid accumulation in two green microalgae with special emphasis on neutral lipid detection by flow cytometry. Adv. Biol. 2016. [Google Scholar] [CrossRef]
- Prochazkova, G.; Safarik, I.; Branyik, T. Harvesting microalgae with microwave synthesized magnetic microparticles. Bioresour. Technol. 2013, 130, 472–477. [Google Scholar] [CrossRef]
- Wang, X.; Zhao, Y.; Jiang, X.; Liu, L.; Li, X.; Li, H.; Liang, W. In-situ self-assembly of plant polyphenol-coated Fe3O4 particles for oleaginous microalgae harvesting. J. Environ. Manag. 2018, 214, 335–345. [Google Scholar] [CrossRef]
- Ge, S.; Agbakpe, M.; Zhang, W.; Kuang, L.; Wu, Z.; Wang, X. Recovering magnetic Fe3O4–ZnO nanocomposites from algal biomass based on hydrophobicity shift under UV irradiation. ACS Appl. Mater. Interfaces 2015, 7, 11677–11682. [Google Scholar] [CrossRef]
- Fraga-García, P.; Kubbutat, P.; Brammen, M.; Schwaminger, S.; Berensmeier, S. Bare Iron Oxide Nanoparticles for Magnetic Harvesting of Microalgae: From Interaction Behavior to Process Realization. Nanomaterials 2018, 8, 292. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.R.; Wang, F.; Wang, S.K.; Liu, C.Z.; Guo, C. Efficient harvesting of marine microalgae Nannochloropsis maritima using magnetic nanoparticles. Bioresour. Technol. 2013, 138, 387–390. [Google Scholar] [CrossRef]
- Boli, E.; Savvidou, M.; Logothetis, D.; Louli, V.; Pappa, G.; Voutsas, E.; Magoulas, K. Magnetic harvesting of marine algae Nannochloropsis oceanica. Sep. Sci. Technol. 2017, 1–8. [Google Scholar] [CrossRef]
- Savvidou, M.G.; Lymperopoulou, T.V.; Mamma, D.; Balta-Brouma, K.P.; Sotiroudis, T.G.; Kekos, D.; Kolisis, F.N. A study on the combined effects of carbon and nitrogen source on high added value products synthesis by Nannochloropsis oceanica CCMP1779 using response surface methodology. Biocatal. Agric. Biotechnol. 2017, 10, 298–307. [Google Scholar] [CrossRef]
- Savvidou, M.G.; Katsabea, A.; Kotidis, P.; Mamma, D.; Lymperopoulou, T.V.; Kekos, D.; Kolisis, F.N. Studies on the catalytic behavior of a membrane-bound lipolytic enzyme from the microalgae Nannochloropsis oceanica CCMP1779. Enzym. Microb. Technol. 2018, 116, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Álvarez-Díaz, P.D.; Ruiz, J.; Arbib, Z.; Barragán, J.; Garrido-Pérez, C.; Perales, J.A. Lipid production of microalga Ankistrodes musfalcatus increased by nutrient and light starvation in a two-stage cultivation process. Appl. Biochem. Biotechnol. 2014, 174, 1471–1483. [Google Scholar] [CrossRef] [PubMed]
- Olson, R.J.; Chisholm, S.W. Effects of light and nitrogen limitation on the cell cycle of the dinoflagellate Amphidinium carteri. J. Plankton Res. 1986, 8, 785–793. [Google Scholar] [CrossRef]
- Loukin, S.; Kung, C. Manganese effectively supports yeast cell-cycle progression in place of calcium. J. Cell Biol. 1995, 131, 1025–1037. [Google Scholar] [CrossRef] [Green Version]
- Böttger, L.H.; Miller, E.P.; Andresen, C.; Matzanke, B.F.; Küpper, F.C.; Carrano, C.J. A typical iron storage in marine brown algae: A multidisciplinary study of iron transport and storage in Ectocarpus siliculosus. J. Exp. Bot. 2012, 63, 5763–5772. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Cao, Y.; Xu, H.; Liu, Y.; Sun, J.; Qiao, D.; Cao, Y. Effect of nitrogen-starvation, light intensity and iron on triacylglyceride/carbohydrate production and fatty acid profile of Neochloris oleoabundans HK-129 by a two-stage process. Bioresour. Technol. 2014, 155, 204–212. [Google Scholar] [CrossRef]
- Ahmad, A.; Osman, S.M.; San Cha, T. Phosphate-induced changes in fatty acid biosynthesis in Chlorella sp. KS-MA2 strain. J. Biotechnol. Comput. Biol. Bionanotechnol. 2016, 97, 295–304. [Google Scholar] [CrossRef]
- Liang, K.; Zhang, Q.; Gu, M.; Cong, W. Effect of phosphorus on lipid accumulation in freshwater microalga Chlorella sp. J. Appl. Phycol. 2013, 25, 311–318. [Google Scholar] [CrossRef]
- Wang, Z.T.; Ullrich, N.; Joo, S.; Waffenschmidt, S.; Goodenough, U. Algal lipid bodies: Stress induction, purification, and biochemical characterization in wild-type and starchless Chlamydomonas reinhardtii. Eukaryot. Cell 2009, 8, 1856–1868. [Google Scholar] [CrossRef] [Green Version]
- Converti, A.; Casazza, A.A.; Ortiz, E.Y.; Perego, P.; Del Borghi, M. Effect of temperature and nitrogen concentration on the growth and lipid content of Nannochloropsis oculata and Chlorella vulgaris for biodiesel production. Chem. Eng. Process. Process. Intensif. 2009, 48, 1146–1151. [Google Scholar] [CrossRef]
- Lv, J.M.; Cheng, L.H.; Xu, X.H.; Zhang, L.; Chen, H.L. Enhanced lipid production of Chlorella vulgaris by adjustment of cultivation conditions. Bioresour. Technol. 2010, 101, 6797–6804. [Google Scholar] [CrossRef] [PubMed]
- Juneja, A.; Ceballos, R.; Murthy, G. Effects of environmental factors and nutrient availability on the biochemical composition of algae for biofuels production: A review. Energies 2013, 6, 4607–4638. [Google Scholar] [CrossRef] [Green Version]
- Millaleo, R.; Reyes-Díaz, M.; Alberdi, M.; Ivanov, A.G.; Krol, M.; Hüner, N.P.A. Excess manganese differentially inhibits photosystem I versus II in Arabidopsis thaliana. J. Exp. Bot. 2012, 64, 343–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xing, W.; Huang, W.M.; Li, D.H.; Liu, Y.D. Effects of iron on growth, pigment content, photosystem II efficiency, and siderophores production of Microcystis aeruginosa and Microcystis wesenbergii. Curr. Microbiol. 2007, 55, 94–98. [Google Scholar] [CrossRef] [PubMed]
- Varsano, T.; Wolf, S.G.; Pick, U. A chlorophyll a/b- binding protein homolog that is induced by iron deficiency is associated with enlarged photosystem I units in the eukaryotic alga Dunaliella Salina. J. Biol. Chem. 2006, 281, 10305–10315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Kassas, H.Y. Growth and fatty acid profile of the marine microalga Picochlorum sp. grown under nutrient stress conditions. Egypt. J. Aquat. Res. 2013, 39, 233–239. [Google Scholar] [CrossRef] [Green Version]
- Yodsuwan, N.; Sawayama, S.; Sirisansaneeyakul, S. Effect of nitrogen concentration on growth, lipid production and fatty acid profiles of the marine diatom Phaeodactylum tricornutum. Agric. Nat. Resour. 2017, 51, 190–197. [Google Scholar] [CrossRef]
- Cañavate, J.P.; Armada, I.; Hachero-Cruzado, I. Common and species-specific effects of phosphate on marine microalgae fatty acids shape their function in phytoplankton trophic ecology. Microb. Ecol. 2017, 74, 623–639. [Google Scholar] [CrossRef]
- Urzica, E.I.; Vieler, A.; Hong-Hermesdorf, A.; Page, M.D.; Casero, D.; Gallaher, S.D.; Merchant, S.S. Remodeling of membrane lipids in iron starved Chlamydomonas. J. Biol. Chem. 2013, 288, 30246–30258. [Google Scholar] [CrossRef] [Green Version]
- Carpio, R.B.; De Leon, R.L.; Martinez-Goss, M.R. Growth, lipid content, and lipid profile of the green alga, Chlorella vulgaris Beij., under different concentrations of Fe and CO2. J. Eng. Sci. Technol. 2015, 6, 19–30. [Google Scholar]
- Wang, S.K.; Stiles, A.R.; Guo, C.; Liu, C.Z. Harvesting microalgae by magnetic separation: A review. Algal Res. 2015, 9, 178–185. [Google Scholar] [CrossRef]
- Guillard, R.R.L.; Ryther, J.H. Studies of Marine Planktonic Diatoms: I. Cyclotella nana Hustedt and Detonula confervacea (Cleve) Gran. Can. J. Microbiol. 1962, 8, 229–239. [Google Scholar] [CrossRef] [PubMed]
- Henriques, M.; Silva, A.; Rocha, J. Extraction and quantification of pigments from a marine microalga: A simple and reproducible method. Commun. Curr. Res. Educ. Top. Trends Appl. Microbiol. Formatex 2007, 2, 586–593. [Google Scholar]
- Chen, Y.; Wang, J.; Liu, T.; Gao, L. Effects of initial population density (IPD) on growth and lipid composition of Nannochloropsis sp. J. Appl. Phycol. 2012, 24, 1623–1627. [Google Scholar] [CrossRef]
- Kail, B.W.; Link, D.D.; Morreale, B.D. Determination of Free Fatty Acids and Triglycerides by gas Chromatography Using Selective Esterification Reactions. J. Chromatogr. Sci. 2012, 50, 934–939. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Medium Composition | F/2-P | F/2-N | F/2-Fe | F/2-Mn | Control |
|---|---|---|---|---|---|
| μ (d−1) | 0.274 | 0.215 | 0.286 | 0.253 | 0.278 |
| Xmax (g L−1) | 0.29 | 0.26 | 0.29 | 0.25 | 0.31 |
| Px (mg L−1 d−1) | 18.55 | 16.44 | 18.15 | 15.37 | 20.01 |
| Y (%DCW) | 23.7 | 41.5 | 33.4 | 34.5 | 37.5 |
| PL (mg L−1 d−1) | 4.0 | 6.4 | 5.7 | 5.0 | 6.8 |
| Chl (μg mL−1) | 0.95 | 1.41 | 4.85 | 2.76 | 2.70 |
| Medium Composition | F/2-P | F/2-N | F/2-Fe | F/2-Mn | Control |
|---|---|---|---|---|---|
| Total FAs content 1 | 10.9 ± 0.5 | 7.3 ± 0.8 | 11.7 ± 0.4 | 11.7 ± 0.6 | 9.22 ± 0.25 |
| % Ratio to total FA | |||||
| Saturated Fatty Acids (SFAs) | |||||
| C12:0 | 1.74 ± 0.2 | 1.64 ± 0.1 | 2.15 ± 0.3 | 2.66 ± 0.2 | 2.39 ± 0.2 |
| C14:0 | 7.15 ± 0.6 | 10.26 ± 0.8 | 8.24 ± 0.7 | 9.09 ± 0.5 | 8.46 ± 0.4 |
| C15:0 | 1.37 ± 0.1 | 1.78 ± 0.3 | 1.97 ± 0.2 | 2.06 ± 0.3 | 1.84 ± 0.3 |
| C16:0 | 36.21 ± 0.7 | 41.04 ± 0.8 | 35.19 ± 0.7 | 33.45 ± 0.9 | 37.64 ± 1.1 |
| C18:0 | 5.68 ± 0.4 | 4.79 ± 0.2 | 4.46 ± 0.4 | 5.75 ± 0.5 | 3.90 ± 0.5 |
| Sum SFAs | 53.35 ± 1.2 | 60.60 ± 0.9 | 53.56 ± 0.8 | 54.72 ± 0.9 | 55.75 ± 1.1 |
| Monounsaturated Fatty Acids (MUFAs) | |||||
| C16:1 n-7 | 26.31 ± 0.8 | 22.16 ± 0.5 | 23.86 ± 0.6 | 21.10 ± 0.7 | 23.43 ± 0.5 |
| C18:1 n-9 | 13.93 ± 0.4 | 10.12 ± 0.5 | 7.81 ± 0.2 | 7.20 ± 0.5 | 10.95 ± 0.4 |
| Sum MUFAs | 40.24 ± 1.1 | 32.38 ± 0.8 | 31.67 ± 0.8 | 28.30 ± 1.6 | 34.38 ± 0.6 |
| Polyunsaturated Fatty Acids (PUFAs) | |||||
| C18:2 n-6 | 1.74 ± 0.2 | 1.78 ± 0.3 | 2.40 ± 0.5 | 2.83 ± 0.4 | 2.17 ± 0.3 |
| C20:4 n-3 (ETA) | 1.65 ± 0.3 | 1.92 ± 0.3 | 3.52 ± 0.5 | 4.03 ± 0.1 | 2.49 ± 0.3 |
| C20:5 n-3 (EPA) | 3.02 ± 0.4 | 3.42 ± 0.2 | 8.84 ± 0.3 | 10.12 ± 0.7 | 5.21 ± 0.4 |
| Sum PUFAs | 6.42 ± 0.8 | 7.11 ± 0.5 | 14.76 ± 0.2 | 16.98 ± 0.6 | 9.87 ± 0.4 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Savvidou, M.G.; Boli, E.; Logothetis, D.; Lymperopoulou, T.; Ferraro, A.; Louli, V.; Mamma, D.; Kekos, D.; Magoulas, K.; Kolisis, F.N. A Study on the Effect of Macro- and Micro- Nutrients on Nannochloropsis oceanica Growth, Fatty Acid Composition and Magnetic Harvesting Efficiency. Plants 2020, 9, 660. https://doi.org/10.3390/plants9050660
Savvidou MG, Boli E, Logothetis D, Lymperopoulou T, Ferraro A, Louli V, Mamma D, Kekos D, Magoulas K, Kolisis FN. A Study on the Effect of Macro- and Micro- Nutrients on Nannochloropsis oceanica Growth, Fatty Acid Composition and Magnetic Harvesting Efficiency. Plants. 2020; 9(5):660. https://doi.org/10.3390/plants9050660
Chicago/Turabian StyleSavvidou, Maria G., Elenitsa Boli, Dimitrios Logothetis, Theopisti Lymperopoulou, Angelo Ferraro, Vasiliki Louli, Diomi Mamma, Dimitris Kekos, Kostis Magoulas, and Fragiskos N. Kolisis. 2020. "A Study on the Effect of Macro- and Micro- Nutrients on Nannochloropsis oceanica Growth, Fatty Acid Composition and Magnetic Harvesting Efficiency" Plants 9, no. 5: 660. https://doi.org/10.3390/plants9050660