
Effect of Active Coatings Containing Lippa citriodora Kunth. Essential Oil on Bacterial Diversity and Myofibrillar Proteins Degradation in Refrigerated Large Yellow Croaker


Abstract
:1. Introduction
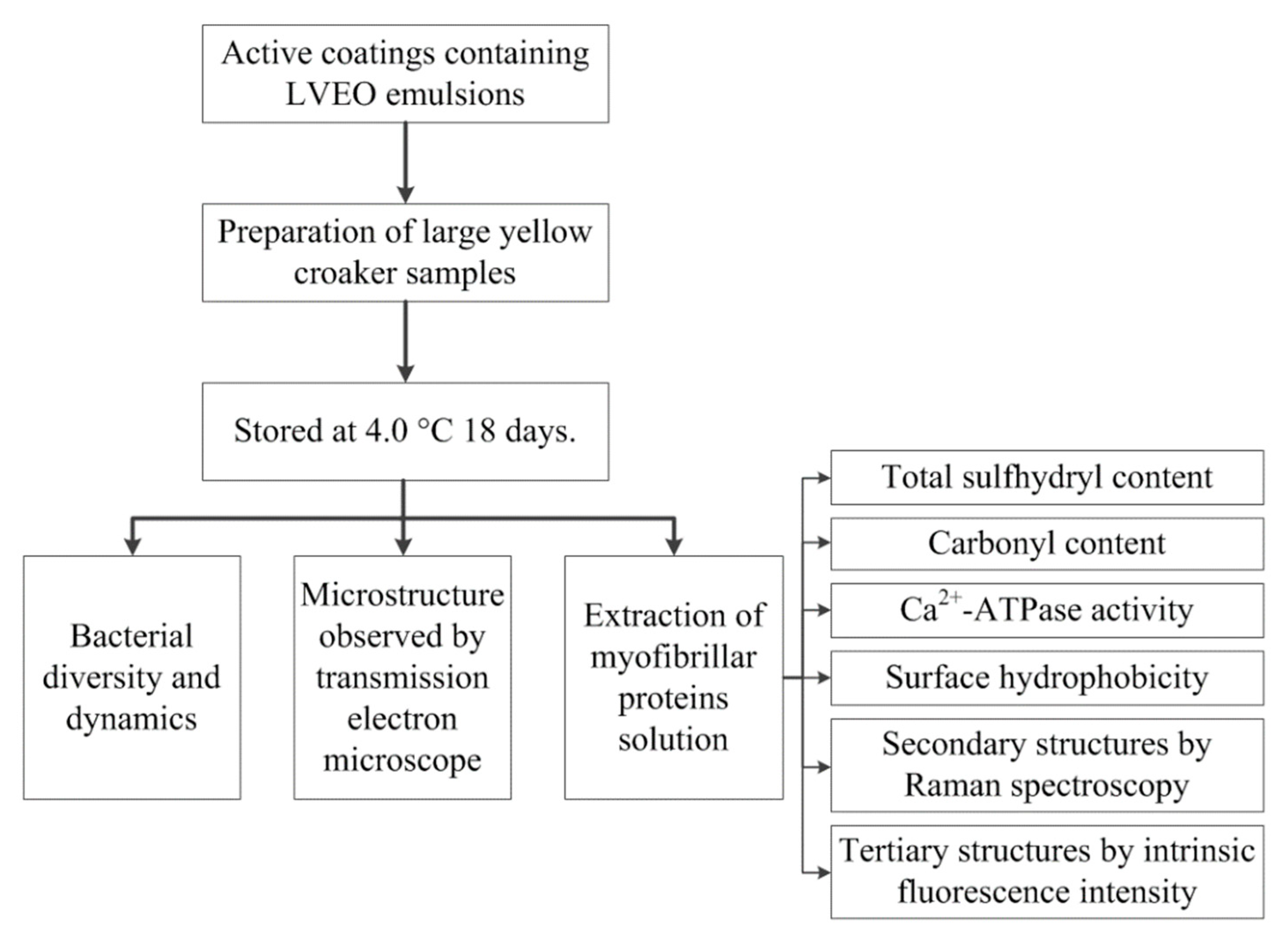
2. Materials and Methods
2.1. Preparation of LBG-SA Active Coating Solutions Containing LVEO Emulsions
2.2. Preparation of Large Yellow Croaker Samples
2.3. Analysis of Bacterial Diversity and Dynamics
2.4. Extraction of MPs Solution
2.4.1. Determination of Total Sulfhydryl Content
2.4.2. Determination of Free Carbonyl Compounds Contents
2.4.3. Determination of Ca2+-ATPase Activity
2.4.4. Determination of Surface Hydrophobicity
2.4.5. Raman Spectroscopy
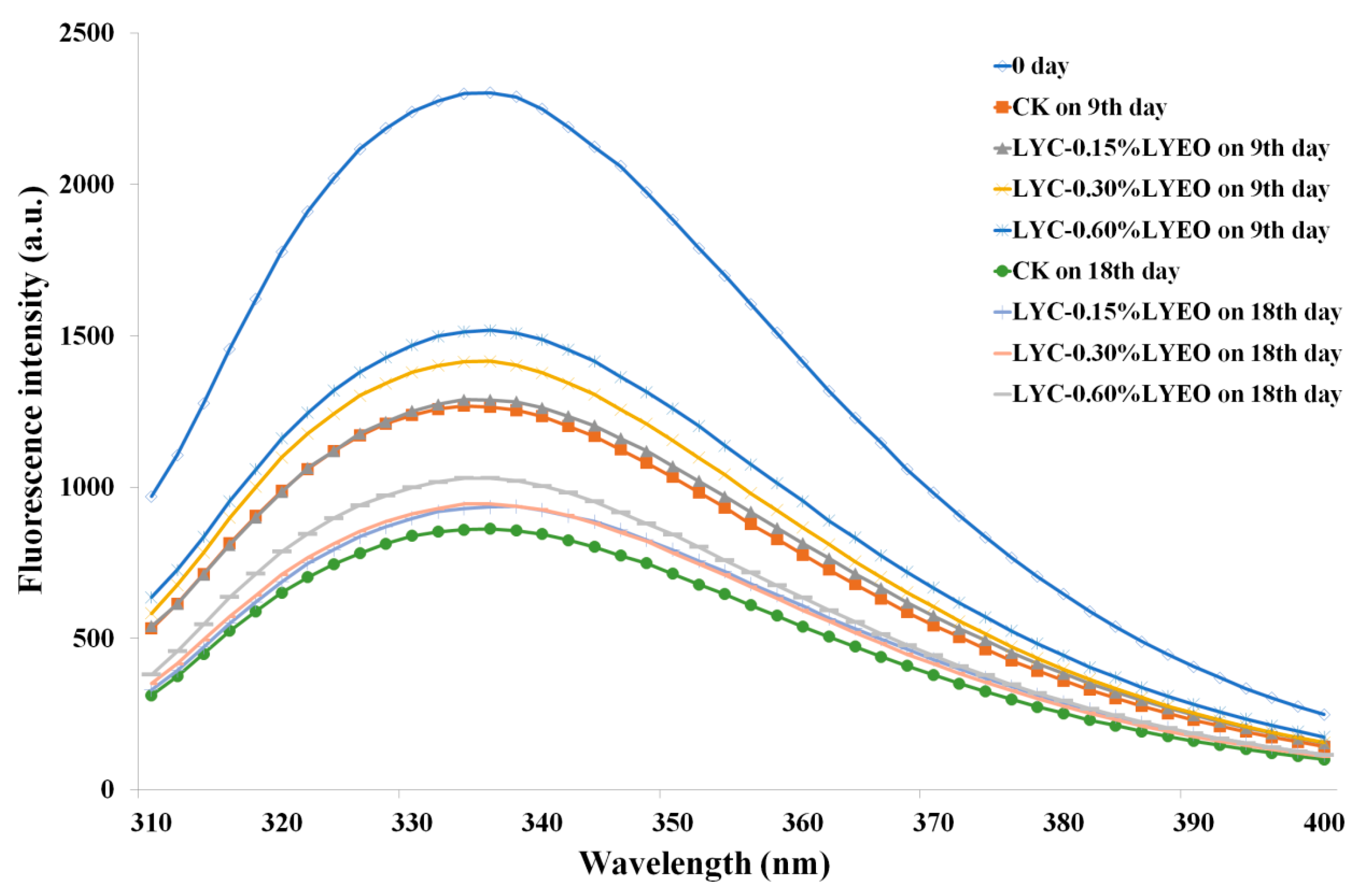
2.4.6. Determination of Intrinsic Fluorescence Intensity (IFI)
2.5. Microstructure Observed by Transmission Electron Microscope (TEM)
2.6. Statistical Analysis
3. Results and Discussions
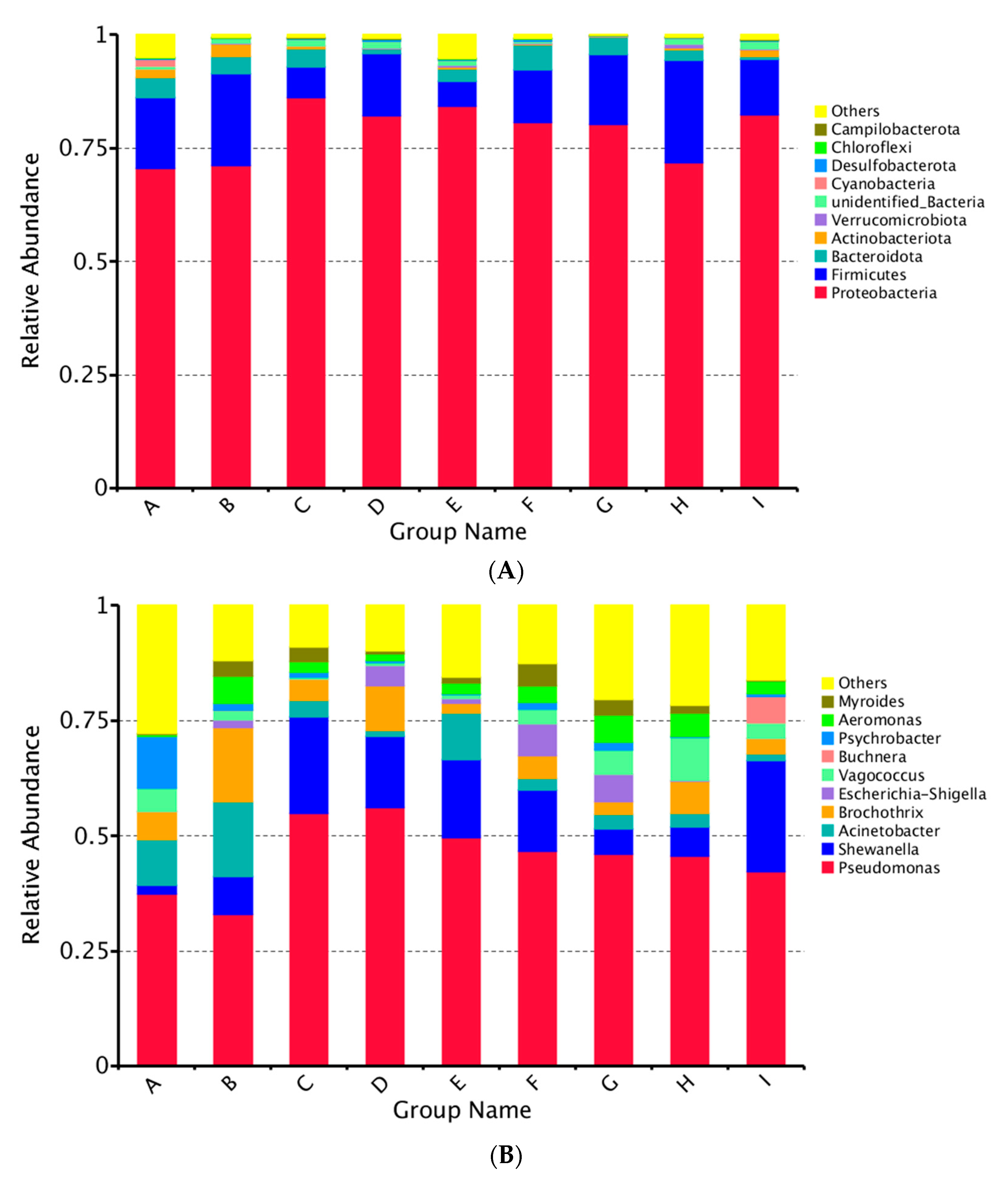
3.1. Microbiome Analysis
3.1.1. Community Abundance and Diversity
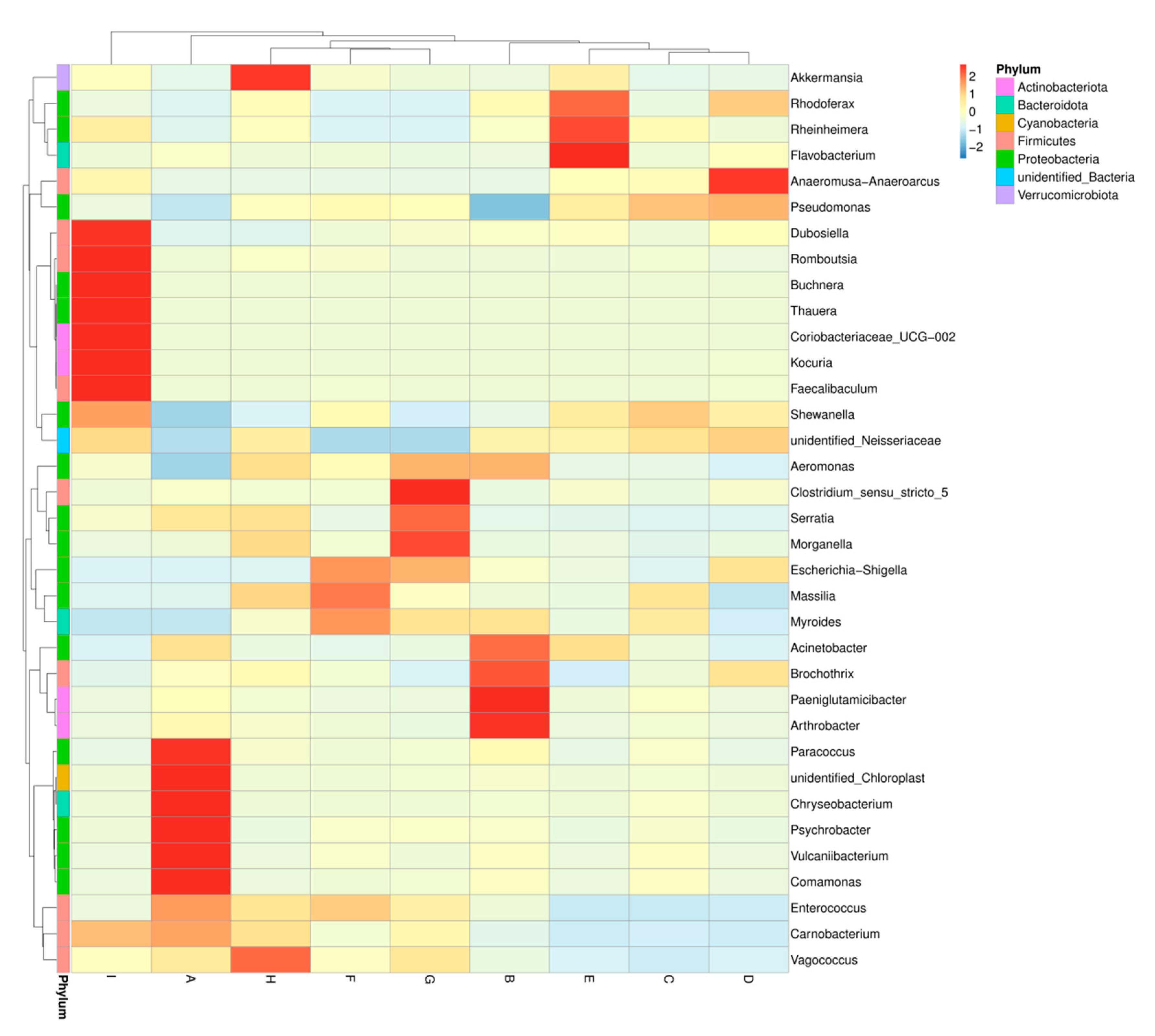
3.1.2. Relative Abundance of the Major Phyla and Genera
3.1.3. Changes of Bacterial Diversity in Large Yellow Croaker Samples during Refrigerated Storage
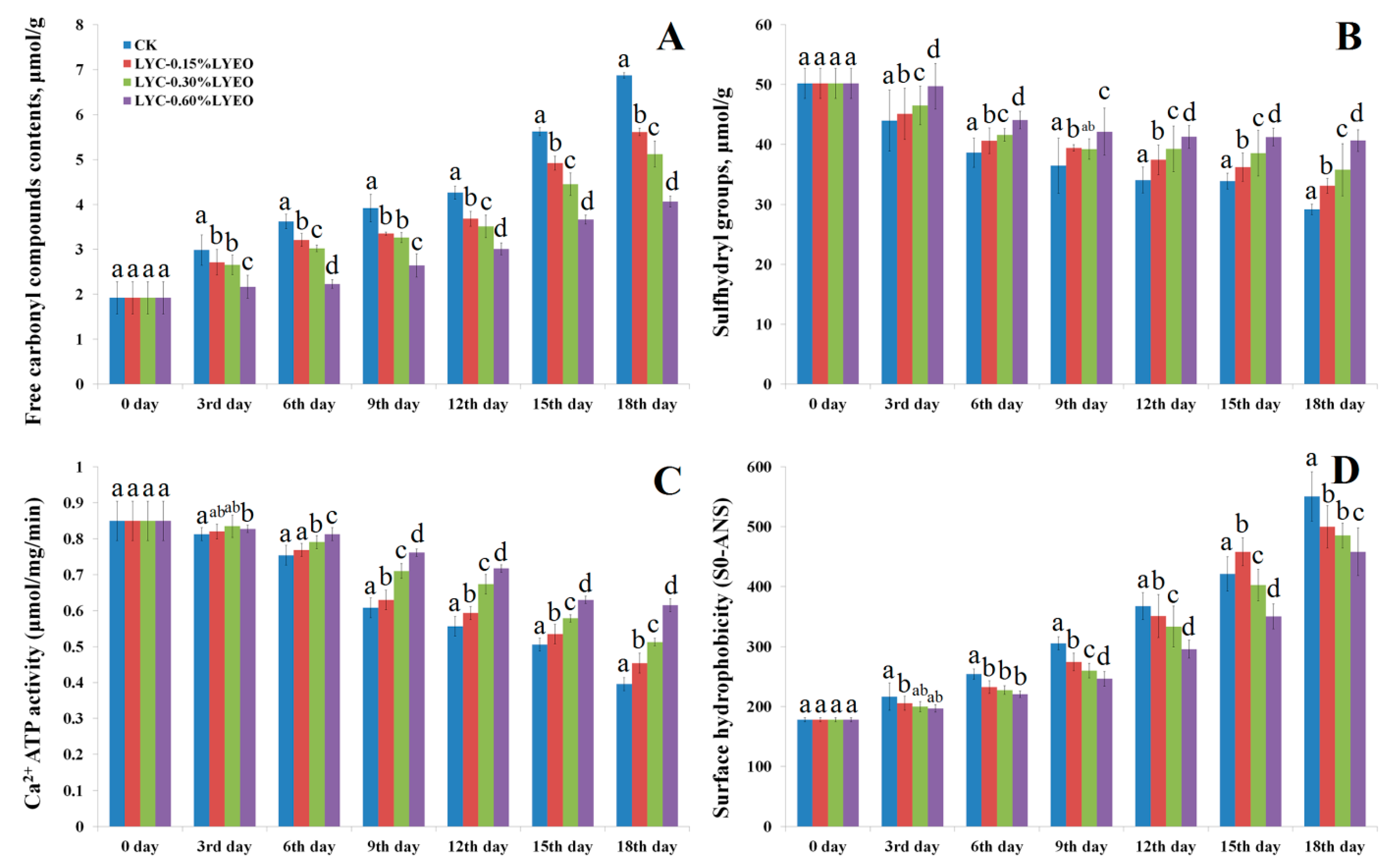
3.2. Changes in Free Carbonyl Compounds Contents
3.3. Changes in Total Sulfhydryl Contents
3.4. Changes in Ca2+-ATPase Activity
3.5. Changes in Surface Hydrophobicity
3.6. Secondary Structure Changes Analysed by Raman Spectroscopy
3.7. Tertiary Structure Changes Analysed by IFI
3.8. Morphological Changes Detected by TEM
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hao, T.; Li, J.; Liu, Q.; Cui, K.; Chen, Q.; Xu, D.; Liu, Y.; Zhao, Y.; Mai, K.; Ai, Q. Fatty acid translocase (FAT/CD36) in large yellow croaker (Larimichthys crocea): Molecular cloning, characterization and the response to dietary fatty acids. Aquaculture 2020, 528, 735557. [Google Scholar] [CrossRef]
- Liang, P.; Li, R.; Sun, H.; Zhang, M.; Cheng, W.; Chen, L.; Cheng, X.; Akoh, C.C. Phospholipids composition and molecular species of large yellow croaker (Pseudosciaena crocea) roe. Food Chem. 2018, 245, 806–811. [Google Scholar] [CrossRef]
- Fu, L.; Wang, C.; Ruan, X.; Li, G.; Zhao, Y.; Wang, Y. Preservation of large yellow croaker (Pseudosciaena crocea) by Coagulin L1208, a novel bacteriocin produced by Bacillus coagulans L1208. Int. J. Food Microbiol. 2018, 266, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.T.; Ge, Y.J.; Li, Y.; Xiang, Y.C.; Jiang, Y.Y.; Hu, Y.Q. Quality enhancement of large yellow croaker treated with edible coatings based on chitosan and lysozyme. Int. J. Biol. Macromol. 2018, 120, 1072–1079. [Google Scholar] [CrossRef] [PubMed]
- Lan, W.Q.; Sun, Y.Q.; Zhang, N.N.; Xie, J. Effects of epsilon-polylysine and rosemary extract on quality attributes and microbial communities in vacuum-packaged large yellow croaker (Pseudosciaena crocea) during ice storage. Food Sci. Biotechnol. 2021. [Google Scholar] [CrossRef] [PubMed]
- Lan, W.Q.; Liu, J.L.; Wang, M.; Xie, J. Effects of apple polyphenols and chitosan-based coatings on quality and shelf life of large yellow croaker (Pseudosciaena crocea) as determined by low field nuclear magnetic resonance and fluorescence spectroscopy. J. Food Saf. 2021, e12887. [Google Scholar] [CrossRef]
- Su, H.M.; Chen, W.; Fu, S.L.; Wu, C.; Li, K.; Huang, Z.Y.; Wu, T.; Li, J.R. Antimicrobial effect of bayberry leaf extract forthe preservation of large yellow croaker (Pseudosciaena crocea). J. Sci. Food Agric. 2014, 94, 935–942. [Google Scholar] [CrossRef] [PubMed]
- Zheng, D.; Chen, L.; Yuman, G.; Ishfaq, A.; Ramesh Pavase, T.; Liangtao, L.; Zhenxing, L.; Hong, L. Characterization of new active packaging based on PP/LDPE composite films containing attapulgite loaded with Allium sativum essence oil and its application for large yellow croaker (Pseudosciaena crocea) fillets. Food Packag. Shelf 2019, 20, 100320. [Google Scholar] [CrossRef]
- Lu, W.; Cui, R.; Zhu, B.; Qin, Y.; Cheng, G.; Li, L.; Yuan, M. Influence of clove essential oil immobilized in mesoporous silica nanoparticles on the functional properties of poly(lactic acid) biocomposite food packaging film. J. Mater. Res. Technol. 2021, 11, 1152–1161. [Google Scholar] [CrossRef]
- Aloui, H.; Khwaldia, K.; Licciardello, F.; Mazzaglia, A.; Muratore, G.; Hamdi, M.; Restuccia, C. Efficacy of the combined application of chitosan and Locust Bean Gum with different citrus essential oils to control postharvest spoilage caused by Aspergillus flavus in dates. Int. J. Food Microbiol. 2014, 170, 21–28. [Google Scholar] [CrossRef]
- Mostafavi, F.S.; Kadkhodaee, R.; Emadzadeh, B.; Koocheki, A. Preparation and characterization of tragacanth-locust bean gum edible blend films. Carbohyd. Polym. 2016, 139, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Licciardello, F.; Kharchoufi, S.; Muratore, G.; Restuccia, C. Effect of edible coating combined with pomegranate peel extract on the quality maintenance of white shrimps (Parapenaeus longirostris) during refrigerated storage. Food Packag. Shelf 2018, 17, 114–119. [Google Scholar] [CrossRef]
- Tian, H.; Xiang, D.; Li, C. Tea polyphenols encapsulated in W/O/W emulsions with xanthan gum–locust bean gum mixture: Evaluation of their stability and protection. Int. J. Biol. Macromol. 2021, 175, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Yerramathi, B.B.; Kola, M.; Annem Muniraj, B.; Aluru, R.; Thirumanyam, M.; Zyryanov, G.V. Structural studies and bioactivity of sodium alginate edible films fabricated through ferulic acid crosslinking mechanism. J. Food Eng. 2021, 301, 110566. [Google Scholar] [CrossRef]
- Farias, N.S.d.; Silva, B.; de Oliveira Costa, A.C.; Müller, C.M.O. Alginate based antioxidant films with yerba mate (Ilex paraguariensis St. Hil.): Characterization and kinetics of phenolic compounds release. Food Packag. Shelf 2021, 28, 100548. [Google Scholar] [CrossRef]
- Aloui, H.; Deshmukh, A.R.; Khomlaem, C.; Kim, B.S. Novel composite films based on sodium alginate and gallnut extract with enhanced antioxidant, antimicrobial, barrier and mechanical properties. Food Hydrocoll. 2021, 113, 106508. [Google Scholar] [CrossRef]
- Prajapati, V.D.; Jani, G.K.; Moradiya, N.G.; Randeria, N.P.; Maheriya, P.M.; Nagar, B.J. Locust bean gum in the development of sustained release mucoadhesive macromolecules of aceclofenac. Carbohyd. Polym. 2014, 26, 113–148. [Google Scholar] [CrossRef] [PubMed]
- Draget, K.I.; Taylor, C. Chemical, physical and biological properties of alginates and their biomedical implications. Food Hydrocoll. 2011, 25, 251–256. [Google Scholar] [CrossRef]
- Yaghoubi, M.; Ayaseh, A.; Alirezalu, K.; Nemati, Z.; Pateiro, M.; Lorenzo, J.M. Effect of chitosan coating incorporated with Artemisia fragrans essential oil on fresh chicken meat during refrigerated storage. Polymers 2021, 13, 716. [Google Scholar] [CrossRef]
- Anis, A.; Pal, K.; Al-Zahrani, S.M. Essential oil-containing polysaccharide-based edible films and coatings for food security applications. Polymers 2021, 13, 575. [Google Scholar] [CrossRef]
- Mei, J.; Ma, X.; Xie, J. Review on natural preservatives for extending fish shelf life. Foods 2019, 8, 490. [Google Scholar] [CrossRef] [Green Version]
- Loke, X.-J.; Chang, C.-K.; Hou, C.-Y.; Cheng, K.-C.; Hsieh, C.-W. Plasma-treated polyethylene coated with polysaccharide and protein containing cinnamaldehyde for active packaging films and applications on tilapia (Orechromis niloticus) fillet preservation. Food Control. 2021, 125, 108016. [Google Scholar] [CrossRef]
- Homayonpour, P.; Jalali, H.; Shariatifar, N.; Amanlou, M. Effects of nano-chitosan coatings incorporating with free /nano-encapsulated cumin (Cuminum cyminum L.) essential oil on quality characteristics of sardine fillet. Int. J. Food Microbiol. 2021, 341, 109047. [Google Scholar] [CrossRef] [PubMed]
- Ansari, M.; Larijani, K.; Saber-Tehrani, M. Antibacterial activity of Lippa citriodora herb essence against MRSA Staphylococcus aureaus. Afr. J. Microbiol. Res. 2012, 6, 16–19. [Google Scholar] [CrossRef]
- Solano-Báez, A.R.; Venegas-Portilla, A.; Rodríguez-Mendoza, J.; Camacho-Tapia, M.; Leyva-Mir, S.G.; Márquez-Licona, G. First report of neoerysiphe aloysiae causing powdery mildew on Lemon verbena (Aloysia citrodora) in Mexico. Plant Dis. 2020, 104, 2293. [Google Scholar] [CrossRef] [Green Version]
- Dianat, M.; Saharkhiz, M.J.; Tavassolian, I. Salicylic acid mitigates drought stress in Lippia citriodora L.: Effects on biochemical traits and essential oil yield. Biocatal. Agric. Biotechnol. 2016, 8, 286–293. [Google Scholar] [CrossRef]
- Bahramsoltani, R.; Rostamiasrabadi, P.; Shahpiri, Z.; Marques, A.M.; Rahimi, R.; Farzaei, M.H. Aloysia citrodora Paláu (Lemon verbena): A review of phytochemistry and pharmacology. J. Ethnopharmacol. 2018, 222, 34–51. [Google Scholar] [CrossRef]
- Vergis, J.; Gokulakrishnan, P.; Agarwal, R.K.; Kumar, A. Essential oils as natural food antimicrobial agents: A review. Crit. Rev. Food Sci. 2015, 55, 1320–1323. [Google Scholar] [CrossRef]
- Rezaeifar, M.; Mehdizadeh, T.; Mojaddar Langroodi, A.; Rezaei, F. Effect of chitosan edible coating enriched with lemon verbena extract and essential oil on the shelf life of vacuum rainbow trout (Oncorhynchus mykiss). J. Food Saf. 2020, 40, e12781. [Google Scholar] [CrossRef]
- Hosseini, M.; Jamshidi, A.; Raeisi, M.; Azizzadeh, M. Effect of sodium alginate coating containing clove (Syzygium Aromaticum) and lemon verbena (Aloysia Citriodora) essential oils and different packaging treatments on shelf life extension of refrigerated chicken breast. J. Food Process. Pres. 2021, 45, e14946. [Google Scholar] [CrossRef]
- Rahmanzadeh Ishkeh, S.; Asghari, M.; Shirzad, H.; Alirezalu, A.; Ghasemi, G. Lemon verbena (Lippia citrodora) essential oil effects on antioxidant capacity and phytochemical content of raspberry (Rubus ulmifolius subsp. sanctus). Sci. Hortic. 2019, 248, 297–304. [Google Scholar] [CrossRef]
- Neri, F.; Mari, M.; Brigati, S.; Bertolini, P. Control of Neofabraea alba by plant volatile compounds and hot water. Postharvest Biol. Technol. 2009, 51, 425–430. [Google Scholar] [CrossRef]
- Liu, W.; Mei, J.; Xie, J. Effect of locust bean gum-sodium alginate coatings incorporated with daphnetin emulsions on the quality of Scophthalmus maximus at refrigerated condition. Int. J. Biol. Macromol. 2021, 170, 129–139. [Google Scholar] [CrossRef]
- Yang, F.; Jia, S.; Liu, J.; Gao, P.; Yu, D.; Jiang, Q.; Xu, Y.; Yu, P.; Xia, W.; Zhan, X. The relationship between degradation of myofibrillar structural proteins and texture of superchilled grass carp (Ctenopharyngodon idella) fillet. Food Chem. 2019, 301, 125278. [Google Scholar] [CrossRef]
- Sun, S.; Wang, S.Q.; Lin, R.; Cheng, S.S.; Yuan, B.; Wang, Z.X.; Tan, M.Q. Effect of different cooking methods on proton dynamics and physicochemical attributes in spanish mackerel assessed by low-field NMR. Foods 2020, 9, 364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.-B.; Pan, S.-K.; Wu, S.-J. Chitooligosaccharides suppress the freeze-denaturation of actomyosin in Aristichthys nobilis surimi protein. Int. J. Biol. Macromol. 2014, 63, 104–106. [Google Scholar] [CrossRef]
- Zhang, Z.; Xiong, Z.; Lu, S.; Walayat, N.; Hu, C.; Xiong, H. Effects of oxidative modification on the functional, conformational and gelling properties of myofibrillar proteins from Culter alburnus. Int. J. Biol. Macromol. 2020, 162, 1442–1452. [Google Scholar] [CrossRef]
- Cao, M.; Cao, A.; Wang, J.; Cai, L.; Regenstein, J.; Ruan, Y.; Li, X. Effect of magnetic nanoparticles plus microwave or far-infrared thawing on protein conformation changes and moisture migration of red seabream (Pagrus Major) fillets. Food Chem. 2018, 266, 498–507. [Google Scholar] [CrossRef]
- Tan, M.; Li, P.; Yu, W.; Wang, J.; Xie, J. Effects of glazing with preservatives on the quality changes of squid during frozen storage. Appl. Sci. 2019, 9, 3847. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Chen, X.; Ding, Y.; Ke, Z.; Zhou, X.; Zhang, J. Diversity and succession of the microbial community and its correlation with lipid oxidation in dry-cured black carp (Mylopharyngodon piceus) during storage. Food Microbiol. 2021, 98, 103686. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Pu, H.; Sun, D.-W. Novel techniques for evaluating freshness quality attributes of fish: A review of recent developments. Trends Food Sci. Technol. 2019, 83, 259–273. [Google Scholar] [CrossRef]
- Sterniša, M.; Bucar, F.; Kunert, O.; Možina, S.S. Targeting fish spoilers Pseudomonas and Shewanella with oregano and nettle extracts. Int. J. Food Microbiol. 2020, 328, 108664. [Google Scholar] [CrossRef] [PubMed]
- Maillet, A.; Denojean, P.; Bouju-Albert, A.; Scaon, E.; Leuillet, S.; Dousset, X.; Jaffres, E.; Combrisson, J.; Prevost, H. Characterization of bacterial communities of cold-smoked salmon during storage. Foods 2021, 10, 362. [Google Scholar] [CrossRef] [PubMed]
- Thielmann, J.; Theobald, M.; Wutz, A.; Krolo, T.; Buergy, A.; Niederhofer, J.; Welle, F.; Muranyi, P. Litsea cubeba fruit essential oil and its major constituent citral as volatile agents in an antimicrobial packaging material. Food Microbiol. 2021, 96, 103725. [Google Scholar] [CrossRef] [PubMed]
- Ghavam, M.; Manconi, M.; Manca, M.L.; Bacchetta, G. Extraction of essential oil from Dracocephalum kotschyi Boiss. (Lamiaceae), identification of two active compounds and evaluation of the antimicrobial properties. J. Ethnopharmacol. 2021, 267, 113513. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Liu, X.; Jia, S.; Zhang, L.; Luo, Y. The effect of essential oils on microbial composition and quality of grass carp (Ctenopharyngodon idellus) fillets during chilled storage. Int. J. Food Microbiol. 2018, 266, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Zhou, Y.; Zhao, L.; Chen, L.; He, Y.; Yang, H. Vacuum impregnation of fish gelatin combined with grape seed extract inhibits protein oxidation and degradation of chilled tilapia fillets. Food Chem. 2019, 294, 316–325. [Google Scholar] [CrossRef]
- Mozaffarzogh, M.; Misaghi, A.; Shahbazi, Y.; Kamkar, A. Evaluation of probiotic carboxymethyl cellulose-sodium caseinate films and their application in extending shelf life quality of fresh trout fillets. LWT 2020, 126, 109305. [Google Scholar] [CrossRef]
- Saheem, A. Protein oxidation: An overview of metabolism of sulphur containing amino acid, Cysteine. Front. Biosci. 2017, 9, 71–87. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.; Benjakul, S. The combined effect of squid pen chitooligosaccharides and high voltage cold atmospheric plasma on the shelf-life extension of Asian sea bass slices stored at 4 °C. Innov. Food Sci. Emerg. 2020, 64, 102339. [Google Scholar] [CrossRef]
- Farvin, K.; Grejsen, H.D.; Jacobsen, C. Potato peel extract as a natural antioxidant in chilled storage of minced horse mackerel (Trachurus trachurus): Effect on lipid and protein oxidation. Food Chem. 2012, 131, 843–851. [Google Scholar] [CrossRef]
- Zhang, R.R.; Xiong, S.B.; You, J.; Hu, Y.; Liu, R.; Yin, T. Effects of ozone treatments on the physicochemical changes of myofibrillar proteins from silver Carp (Hypophthalmichthys molitrix) during frozen storage. J. Food Qual. 2017, 9506596. [Google Scholar] [CrossRef] [Green Version]
- Xia, X.; Kong, B.; Qian, L.; Jing, L. Physicochemical change and protein oxidation in porcine longissimus dorsi as influenced by different freeze-thaw cycles. Meat Sci. 2009, 83, 239–245. [Google Scholar] [CrossRef]
- Prodpran, T.; Benjakul, S.; Phatcharat, S. Effect of phenolic compounds on protein cross-linking and properties of film from fish myofibrillar protein. Int. J. Biol. Macromol. 2012, 51, 774–782. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Lv, W.; Wang, J.; Li, J.; Liu, X.; Zhu, J. Effects of tea polyphenols on the post-mortem integrity of large yellow croaker (Pseudosciaena crocea) fillet proteins. Food Chem. 2013, 141, 2666–2674. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Yang, H. Effects of calcium ion on gel properties and gelation of tilapia (Oreochromis niloticus) protein isolates processed with pH shift method. Food Chem. 2019, 277, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Hong, H.; Zhang, L.; Zhang, C.; Luo, Y. Antioxidant and cryoprotective effects of hydrolysate from gill protein of bighead carp (Hypophthalmichthys nobilis) in preventing denaturation of frozen surimi. Food Chem. 2019, 298, 124868. [Google Scholar] [CrossRef] [PubMed]
- Kong, B.; Guo, Y.; Xia, X.; Liu, Q.; Li, Y.; Chen, H. Cryoprotectants reduce protein oxidation and structure deterioration induced by freeze-thaw cycles in common carp (Cyprinus carpio) surimi. Food Biophys. 2013, 8, 104–111. [Google Scholar] [CrossRef]
- Kong, C.; Wang, H.; Li, D.; Zhang, Y.; Pan, J.; Zhu, B.; Luo, Y. Quality changes and predictive models of radial basis function neural networks for brined common carp (Cyprinus carpio) fillets during frozen storage. Food Chem. 2016, 201, 327–333. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Xiong, G.; Peng, Y.B.; Wu, W.; Li, X.; Wang, J.; Qiao, Y.; Liao, L.; Ding, A. The cryoprotective effect of different konjac glucomannan (KGM) hydrolysates on the glass carp (Ctenopharyngodon idella) myofibrillar during frozen storage. Food Bioprocess Tech. 2014, 7, 3398–3406. [Google Scholar] [CrossRef]
- Leelapongwattana, K.; Benjakul, S.; Visessanguan, W.; Howell, N.K. Physicochemical and biochemical changes during frozen storage of minced flesh of lizardfish (Saurida micropectoralis). Food Chem. 2005, 90, 141–150. [Google Scholar] [CrossRef] [Green Version]
- Xiao, H.; Wu, Y.; Liu, J.; Zhou, L.; Zeng, X.; Zhao, X. Potential mechanism of different gelation properties of white and red muscle fibre from crocodile (Crocodylus siamensis) meat: Study of myofibrillar protein. LWT 2021, 143, 111045. [Google Scholar] [CrossRef]
- Cai, L.; Zhang, W.; Cao, A.; Cao, M. Effects of different thawing methods on the quality of largemouth bass (Micropterus salmonides). LWT 2020, 120, 108908. [Google Scholar] [CrossRef]
- Xu, X.L.; Han, M.Y.; Fei, Y.; Zhou, G.H. Raman spectroscopic study of heat-induced gelation of pork myofibrillar proteins and its relationship with textural characteristic. Meat Sci. 2011, 87, 159–164. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Zhao, Y.Y.; Kang, Z.L.; Wang, P.; Han, M.Y.; Xu, X.L.; Zhou, G.H. Reduced functionality of PSE-like chicken breast meat batter resulting from alterations in protein conformation. Poultry Sci. 2015, 94, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Martínez, M.; Velazquez, G.; Cando, D.; Núnez-Flores, R.; Borderías, A.; Moreno, H.M. Effects of high pressure processing on protein fractions of blue crab (Callinectes sapidus) meat. Innov. Food Sci. Emerg. 2017, 41, 323–329. [Google Scholar] [CrossRef]
- Wei, P.; Zhu, K.; Cao, J.; Dong, Y.; Li, M.; Shen, X.; Duan, Z.; Li, C. The inhibition mechanism of the texture deterioration of tilapia fillets during partial freezing after treatment with polyphenols. Food Chem. 2021, 335, 127647. [Google Scholar] [CrossRef] [PubMed]
- Zhao, N.; Zou, H.; Sun, S.; Yu, C. The interaction between sodium alginate and myofibrillar proteins: The rheological and emulsifying properties of their mixture. Int. J. Biol. Macromol. 2020, 161, 1545–1551. [Google Scholar] [CrossRef]
- Li, F.; Wang, B.; Kong, B.; Shi, S.; Xia, X. Decreased gelling properties of protein in mirror carp (Cyprinus carpio) are due to protein aggregation and structure deterioration when subjected to freeze-thaw cycles. Food Hydrocoll. 2019, 97, 105223. [Google Scholar] [CrossRef]
- Zhang, M.; Li, F.; Diao, X.; Kong, B.; Xia, X. Moisture migration, microstructure damage and protein structure changes in porcine longissimus muscle as influenced by multiple freeze-thaw cycles. Meat Sci. 2017, 133, 10–18. [Google Scholar] [CrossRef]
- Cao, Y.; Xiong, Y.L. Chlorogenic acid-mediated gel formation of oxidatively stressed myofibrillar protein. Food Chem. 2015, 180, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Roco, T.; Torres, M.J.; Briones-Labarca, V.; Reyes, J.E.; Tabilo-Munizaga, G.; Stucken, K.; Lemus-Mondaca, R.; Pérez-Won, M. Effect of high hydrostatic pressure treatment on physical parameters, ultrastructure and shelf life of pre- and post-rigor mortis palm ruff (Seriolella violacea) under chilled storage. Food Res. Int. 2018, 108, 192–202. [Google Scholar] [CrossRef] [PubMed]
- Mahto, R.; Ghosh, S.; Das, M.K.; Das, M. Effect of gamma irradiation and frozen storage on the quality of fresh water prawn (Macrobrachium rosenbergii) and tiger prawn (Penaeus monodon). LWT 2015, 61, 573–582. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Treatments | OTUs 1 | Shannon 2 | Simpson | Chao1 3 | ACE 4 | Good’s Coverage 5 |
|---|---|---|---|---|---|---|
| A 6 | 322 | 4.811 | 0.888 | 425.722 | 459.919 | 0.990 |
| B | 247 | 4.045 | 0.874 | 374.918 | 398.132 | 0.990 |
| C | 265 | 3.568 | 0.795 | 421.004 | 442.147 | 0.989 |
| D | 234 | 3.180 | 0.733 | 373.916 | 391.066 | 0.990 |
| E | 258 | 3.715 | 0.799 | 386.996 | 410.779 | 0.990 |
| F | 253 | 3.805 | 0.819 | 367.052 | 403.958 | 0.990 |
| G | 141 | 3.681 | 0.809 | 178.549 | 186.535 | 0.996 |
| H | 195 | 3.987 | 0.834 | 281.772 | 293.864 | 0.993 |
| I | 251 | 3.822 | 0.832 | 395.197 | 408.786 | 0.990 |
| Samples | α-Helices | β-Sheets | β-Turns | Random Coils |
|---|---|---|---|---|
| Fresh | 20.8% | 12.5% | 28.5% | 38.2% |
| CK on 9th day 1 | 27.6% | 13.3% | 20.4% | 38.7% |
| LYC-0.15%LVEO on 9th day | 8.4% | 15.5% | 20.3% | 55.8% |
| LYC-0.30%LVEO on 9th day | 21.0% | 12.0% | 26.6% | 40.4% |
| LYC-0.60%LVEO on 9th day | 17.0% | 20.6% | 33.0% | 29.4% |
| CK on 18th day | 7.6% | 10.1% | 21.7% | 60.6% |
| LYC-0.15%LVEO on 18th day | 8.3% | 15.6% | 20.6% | 55.5% |
| LYC-0.30%LVEO on 18th day | 10.2% | 15.4% | 23.5% | 50.9% |
| LYC-0.60%LVEO on 18th day | 10.9% | 15.6% | 22.3% | 51.2% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, B.; Wang, X.; Gao, X.; Mei, J.; Xie, J. Effect of Active Coatings Containing Lippa citriodora Kunth. Essential Oil on Bacterial Diversity and Myofibrillar Proteins Degradation in Refrigerated Large Yellow Croaker. Polymers 2021, 13, 1787. https://doi.org/10.3390/polym13111787
Li B, Wang X, Gao X, Mei J, Xie J. Effect of Active Coatings Containing Lippa citriodora Kunth. Essential Oil on Bacterial Diversity and Myofibrillar Proteins Degradation in Refrigerated Large Yellow Croaker. Polymers. 2021; 13(11):1787. https://doi.org/10.3390/polym13111787
Chicago/Turabian StyleLi, Bo, Xuesong Wang, Xin Gao, Jun Mei, and Jing Xie. 2021. "Effect of Active Coatings Containing Lippa citriodora Kunth. Essential Oil on Bacterial Diversity and Myofibrillar Proteins Degradation in Refrigerated Large Yellow Croaker" Polymers 13, no. 11: 1787. https://doi.org/10.3390/polym13111787