
Potential Use of Lactic Acid Bacteria with Pathogen Inhibitory Capacity as a Biopreservative Agent for Chorizo


1
Departamento de Biología, Universidad del Valle, Calle 13 No. 100-00, Ciudad Universitaria Meléndez, Santiago de Cali 76001, Valle del Cauca, Colombia
2
Escuela de Ingenieria de alimentos, Universidad del Valle, Calle 13 No. 100-00, Ciudad Universitaria Meléndez, Santiago de Cali 76001, Valle del Cauca, Colombia
*
Author to whom correspondence should be addressed.
Processes 2021, 9(9), 1582; https://doi.org/10.3390/pr9091582
Submission received: 24 June 2021
/
Revised: 13 July 2021
/
Accepted: 20 July 2021
/
Published: 3 September 2021
(This article belongs to the Special Issue Microbial Biotechnology for Environmental Remediation and Restoration)
Abstract
:The biopreservation of meat products is of great interest due to the demand for products with low or minimal chemical additives. Lactic acid bacteria (LAB) have been used as protective cultures for many centuries. The objective of this work was to characterize 10 native LAB isolated from meat masses with biopreservative potential for meat products. The isolates were subjected to viability tests with different concentrations of NaCl, nitrite, and nitrate salts, pHs, and temperature conditions. Antibiotic resistance and type of lactic acid isomer were tested. In addition, the isolates were tested against seven pathogens, and inhibitory substances were identified by diffusion in agar wells. Finally, two isolates, Lb. plantarum (SB17) and Lb. sakei (SB3) were tested as protective cultures of chorizo in a model. As a result, the viability at different concentrations of NaCl and nitrate and nitrate salts were obtained. pH and temperature exerted a negative effect on the growth of some of the isolates. Pathogens were inhibited mainly by the presence of organic acids; P. aurius was the most susceptible, and S. typhimurium and S. marcescens were the most resistant. The strains SB17 and SB3 had similar effects on chorizo, and time exerted a deleterious effect on microbiological quality and pH. The results indicated that the 10 isolates show promising characteristics for the preservation of cooked meat products, with the strain Lb. plantarum (SB17) being the most promising.
1. Introduction
Lactic acid bacteria (LAB) are generally considered safe and have been associated with the production of fermented foods for centuries [1,2]. This group of bacteria can inhibit the growth of pathogenic organisms in different foods of plant or animal origin [3,4], and it is possible to isolate native strains from fermented dairy, meat and vegetable products, and alcoholic beverages. LAB improve characteristics such as aromas, flavor, acidity, and protein digestibility and can benefit the appearance of the product [5,6]. The use of LAB as protective cultures is based on the production of secondary metabolites such as lactic acid and bacteriocins, substances that are considered biopreservatives [1,7]. In addition to their ability to survive or resist extreme and diverse conditions, they are thermostable and have the competitive characteristics necessary for their use as bioprotective and biopreservative cultures [1,8,9].
Meat products are prone to bacterial contamination because they contain compounds that favor bacterial growth [2,10]. These microorganisms are considered spoilage bacteria (that damage or degrade the product) or pathogenic bacteria (that cause symptoms) [11,12]. Foodborne pathogens include Listeria monocytogenes, Escherichia coli, Salmonella typhimurium, Serratia marcescens, Staphylococcus aureus, Pseudomonas aeruginosa, and Proteus vulgaris, which can grow in refrigerated foods [11,13,14].
One method of preservation that has long been used is the application of chemical preservatives, such as nitrite and nitrate salts, which have a negative effect on human health [15,16,17,18]. Currently, there is great consumer demand for high-quality food that is free of chemical preservatives; therefore, there is a need to find alternative preservation methods. One potential preservation method is the use of LAB as a biopreservative; in Colombia, the use of LAB is mostly focused on dairy products and is restricted in fermented meat products [11,14,19]. For this reason, this work aims to characterize native LAB isolated from the meat masses of chorizo and sausage-type products that have biopreservative potential for meat products.
2. Materials and Methods
For this work, 10 native strains of LAB isolated from chorizo and sausage-type meat masses were selected and tested. The strains were deposited in the microorganism, and filamentous fungi strain bank of the Microorganism and Applied Biotechnology Research Group (MIBIA) of the Universidad del Valle, where the tests were carried out, Table 1 shows the identification of the isolates.
2.1. Viability at Different Concentrations of NaCl, Nitrate and Nitrate Salts and Different pH Values and Temperatures
The LAB were tested for viability under different conditions by modifying the MRS broth as follows: pHs of 6, 4, and 3 from acetic acid (3M); NaCl concentrations of 1, 2, and 3%; and nitrite and nitrate salts at 100, 81, 60, and 30 ppm. The medium was inoculated with 1 mL of the bacterial strain and incubated for 16–24 h at 35 °C. The number of viable microorganisms was estimated by plate counting with serial dilutions; this test was performed in triplicate for each strain used, and only plates with counts between 30 and 300 colonies were considered. The number of colonies was multiplied by the inverse of the dilution to obtain the CFU/mL (Jurado & Gúzman, 2015). Using the same method, the strains were incubated for 24 h at 35 °C in MRS broth and subjected to temperatures of 40, 50, 60, and 80 °C for 15 min, followed by microorganism counting.
ANOVAs were performed for pH, NaCl concentrations, and nitrite and nitrate salts with heterogeneous variances, and single-factor ANOVA was performed for the temperature variable.
2.2. Antibiotic Resistance
Antibiotic resistance was determined using the Kirby Bauer method, for which 4 antibiotics (Sigma) were used: amoxicillin (30 µg), ampicillin (10 µg), penicillin (10 IU), and tetracycline (30 µg). The bacterial strains were cultured in 9 mL of MRS broth and then diluted to 10−3 (Macfarland 0.5) and inoculated by depletion on MRS agar. The discs were then overlayed with the antibiotics and incubated at 35 °C for 24 h. After the incubation period, the discs were examined to determine the presence or absence of inhibition halos (the translucent zone formed around the antibiotic discs) [20].
2.3. Lactic Acid Isomers
LAB are characterized by the production of different lactic acid isomers through the fermentation of glucose. During this process, they can produce L(+) dextrorotatory lactic acid, D(−) levorotatory lactic acid, or a racemic mixture of both D and L. The strains were determined by the type of lactic acid isomers they produced, following the method recommended by Betancourt Botero et al., 2013 [21]. This method consisted of an ester-specific enzymatic oxidation-reduction reaction catalyzed by the enzyme lactate dehydrogenase, which oxidizes lactate to pyruvate by simultaneously reducing the L(+) and D(−) isomer to a NAD (NADH) form, which produces a coupling reaction that reduces the tetrazolium salt under diaphoresis, forming a reddish, insoluble compound. With this method, it was possible to differentiate the strains that produced D(−) lactic acid, which had intense reddish halos, from those that produced a racemic mixture (DL), which had less intense reddish halos. The strains that exclusively produced L(+) isomers were distinguished by the absence of color and were recommended for human consumption.
2.4. Antimicrobial Activity Test
The antimicrobial activity of the LAB isolates was determined using the modified disk diffusion method. The commercial strains used were Listeria monocytogenes ATCC 19111, Escherichia coli ATCC 25922, Salmonella typhimurium ATCC 14028, Serratia marcescens ATCC 13880, Staphylococcus aureus ATCC 25923, Pseudomonas aeruginosa ATCC 9027, and Proteus vulgaris ATCC 9484, all of which were activated according to the manufacturer’s instructions.
The LAB strains were cultured in MRS broth for 24 h at 35 °C and then diluted in 9 mL of water with 0.1% peptone to 10−3 (adjusted to the MacFarland scale 0.5). This last dilution was seeded in MRS agar by surface seeding with a sterile swab and incubated at 35 °C for 16 h. Additionally, pathogenic bacteria were cultured in 9 mL of brain heart infusion (BHI) broth for 24 h at 35 °C and diluted to 10−3 in peptone water. A total of 100 microliters of this dilution was removed and sown on Mueller Hinton agar with a sterile swab. From the LAB cultures, which were incubated for 16 h, discs with diameters of 5 mm were extracted and placed in Mueller Hinton boxes inoculated with pathogens at evenly spaced locations and incubated for 24 h at 35 °C. This test was performed in duplicate. A disc of MRS agar without inoculum was also placed as a negative control. The antimicrobial activity was determined according to the diameter of the clear zones surrounding the LAB discs [13]. An ANOVA using a generalized linear model with heterogeneous variances was performed in R version 3.32.
2.5. Identification of Inhibitory Substances
2.5.1. Identification of the Inhibitory Substance by the Agar Well Diffusion Method
To determine the production of inhibitory substances during the growth of the isolates, the method of Angmo et al. 2016 was modified [22]. Isolates were grown in 20 mL of MRS broth for 48 h at 35 °C and then refrigerated for 3 days. Once the incubation time was complete, the samples were centrifuged (10,000× g for 10 min at 4 °C), and the supernatant was sterilized by filtration using 0.22 µm syringe filters (Millipore, MERCK, Burlington, MA, USA) and placed in sterile containers. The cell-free extract (CFE) was treated as follows: fraction A: CFE of the 10 isolates; fraction B: CFE-N with the pH neutralized (pH ≥ 6) by 1 M calcium bicarbonate, which was used to determine inhibition by acids; fraction C: MRS broth as control.
Next, 5 mm-deep wells were made in Müller Hinton plates inoculated with the S. aureus, S. marcescens, and L. monocytogenes strains previously cultured in 9 mL of BHI, and 90 µL aliquots of fractions A, B, and C were added to the wells. The wells were incubated for 48 h at 35 °C, and observations were performed at 24 and 48 h to examine clear zones of inhibition around the wells.
2.5.2. Qualitative and Quantitative Estimation of Organic Acids Produced by the Isolates through HPLC
The isolates were cultured in 20 mL of MRS broth for 48 h at 35 °C. The supernatant was sterilized by filtration using 0.22 µm syringe filters (Millipore, Merck) and deposited in sterile vials at 4 °C for further analysis. The concentration of organic acids in the samples was determined using the high-performance liquid chromatography (HPLC) technique in a Waters® Breeze HPLC chromatograph (Waters, Milford, MA, USA), which performed dual detection by means of 2 detectors: 1 for the refractive index (RI) and the other for ultraviolet (UV). An Aminex® HPX-87 (Bio-Rad, Hercules, CA, USA) column and a mobile phase of 5 mM H2SO4 were used. Organic acids such as citric, succinic, lactic, acetic, and propionic acids were detected [23].
2.5.3. Purification and Enzyme Assay of Crude Cell Extracts
To obtain proteins, the cell-free supernatant with neutralized pH (fraction B) was precipitated with ammonium sulfate reaching 80% saturation, left to dialyze for 12 h at 4 °C and then centrifuged (4000 rpm for 30 min at 4 °C). The precipitate was dissolved in 1 M phosphate buffer (pH 6.0), and the presence and concentration of proteins was estimated by the Lowry method with the standard curve of albumin. Enzyme activity was determined by the agar diffusion method described previously [22,24].
2.6. Meat Mass Testing
Production of Cooked Chorizo
The manufacturing chorizo was carried out at the MIBIA laboratory following the industrial formulation for meat sausage products; see Table 2. In this test, 6 treatments were carried out using two inoculums, SB3 and SB17. For these treatments, 30 g of chorizo-type meat mass was inoculated with 3% of the bacterial strain, with a viability of 106 CFU per mL. This mass was subjected to 80 °C for 15 min, followed by a thermal shock at 4 °C. Once this process was completed, microbiological control was carried out, the pH of the meat product was measured, and 1 g of the sausage was removed and homogenized in 9 mL of water with a Stomacher MiniMix.
The pH was measured at room temperature using an Orion 710-A instrument (Bozkurt and Erkmen, 2002). From these mixtures, 10−4 dilutions were made. A total of 100 µL of the dilution was inoculated in Petri dishes with culture medium, potato dextrose agar (PDA), plate count agar (PCA), MRS with aniline blue, MacConkey agar, and violet red bile agar (VRBA). The cultures were incubated at 28 and 35 °C for 24 h in triplicate. The chorizo was stored at 4 °C under anaerobic conditions and monitored for 40 days.
Microbiological control was carried out according to Colombian Food Industries Technical Standard NTC 1325 (2008) for noncanned processed meat products and took into account the method recommended by the Colombian Technical Standards (NTC 1325, 2008; NTC 4092, 2009) for microbiological counting. The data were analyzed by means of a one-factor ANOVA model in R version 3.32.
This test had 2 controls and 2 treatments, as shown in Table 2.
3. Results
3.1. Viability at Different Concentrations of NaCl, Nitrite and Nitrate Salts, pH Values and Temperatures
3.1.1. Tolerance of LAB to Different NaCl Concentrations
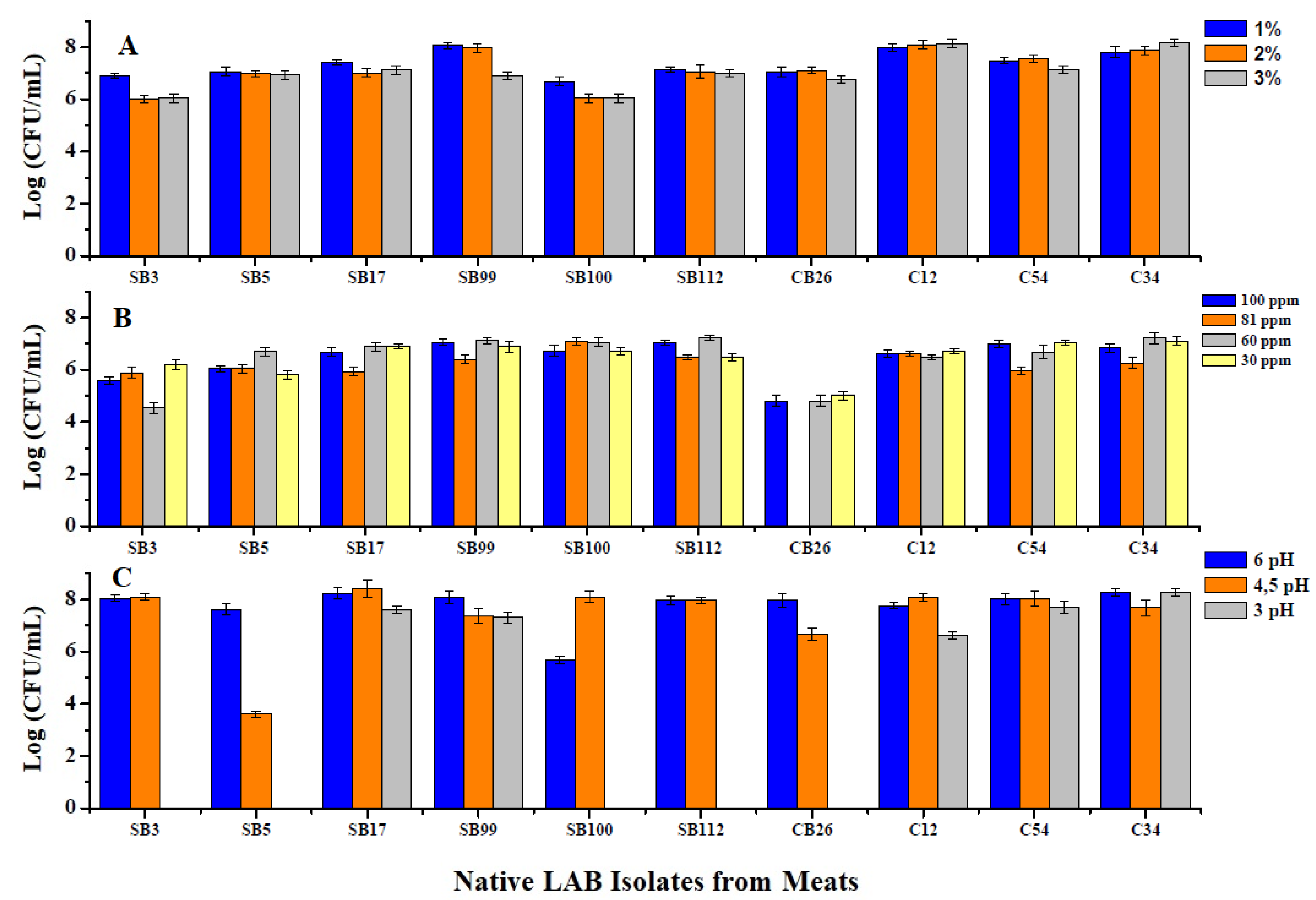
The isolates were able to grow and survive in concentrations of 1, 2, and 3% NaCl; the results are shown in Figure 1A. Statistically, there were significant differences between strains in their responses to salt concentrations (p < 0.0001): strains SB3 and SB100 presented the lowest tolerance of NaCl, and strain C12 presented the highest tolerance.
3.1.2. Tolerance of LAB to Different Concentrations of Nitrate and Nitrate Salts
Regarding the concentration of nitrate and nitrate salts, growth was influenced by nitrate salt, but there were no significant differences in the tolerance of different concentrations (p = 0.9138). However, there were significant differences (p = 0.001) between strains in the tolerance of different concentrations of nitrite and nitrate salts. Strain SB112 presented higher growth at the concentrations evaluated. Strain CB26 was inhibited at the nitrite salt concentration of 81 ppm. The results are shown in Figure 1B.
3.1.3. Viability at Different pH Concentrations
The isolates presented better growth at a pH of 6, although the results were not significantly different from those at a pH 4 (p = 0.1453). Growth under these two treatment conditions differed from growth at pH 3 (p < 0.005), at which five of the isolates were not viable, as shown in Figure 1C.
3.1.4. Viability at Different Temperatures
When the strains were subjected to heat shock for 15 min at 50 °C, strains C12, C54, C34, CB26, SB99, and SB112 were viable and showed no significant differences in growth (p = 0.154) in growth. None of the strains were viable at temperatures of 60 and 80 °C.
3.1.5. Antibiotic Resistance and Type of Lactic Acid
Table 3 shows the type of lactic acid and the antibiotic resistance and susceptibility of the isolates, highlighting the resistance of strains SB3, SB112, and CB26 to tetracycline.
3.2. Inhibition Testing of Selected Isolates
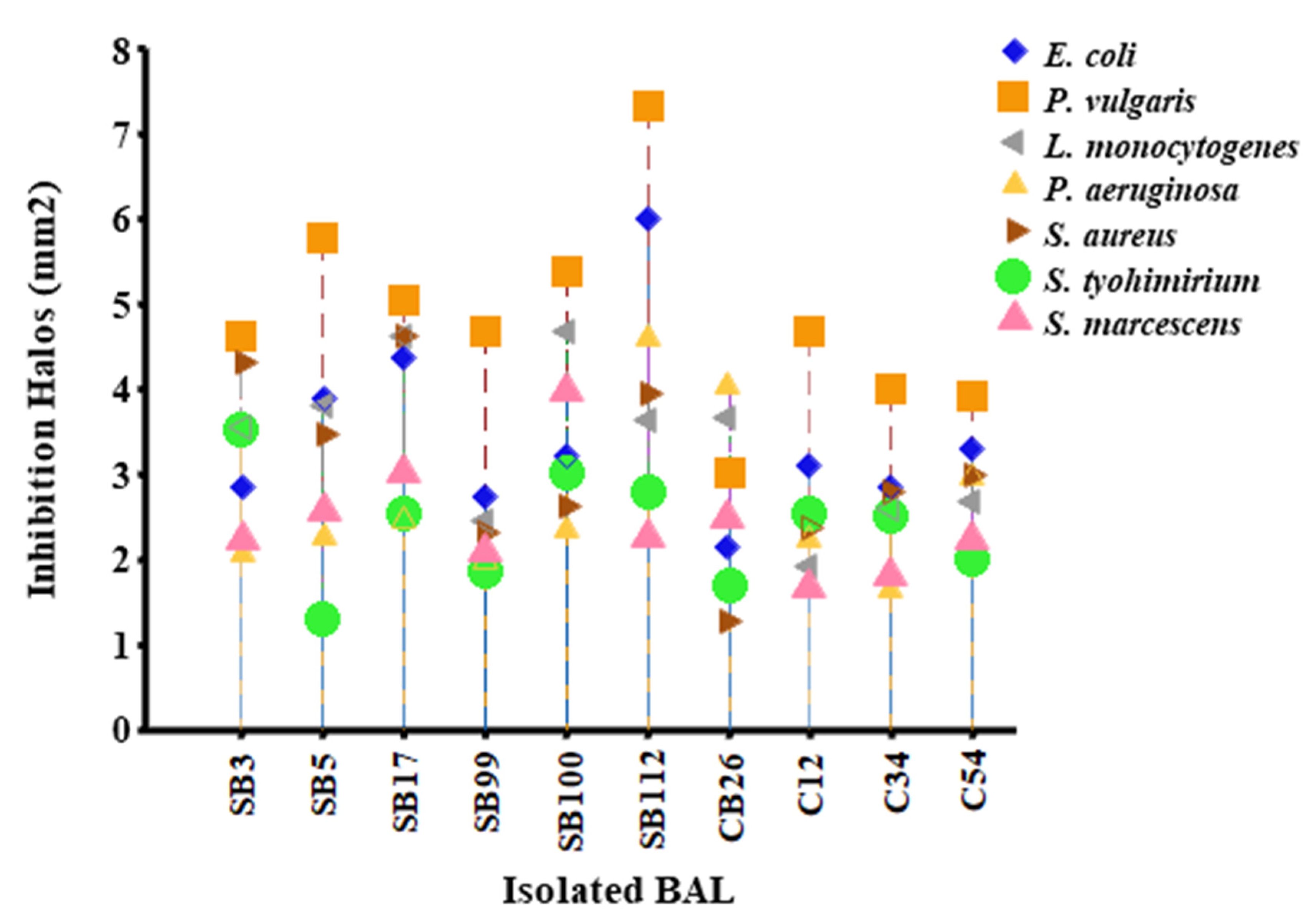
When the 10 isolates were tested against the pathogenic strains, the most susceptible was P. vulgaris, which presented the largest inhibition halos, and the most resistant strains were S. typhimurium and S. marcescens, which had the smallest inhibition halos. The strains with the highest inhibition were SB112, SB17, and SB5, and those with the lowest inhibition were SB99, C54, and C34; these results are shown in Figure 2. Statistically significant differences were found in the inhibition of the pathogen (p < 0.0001) depending on the isolate and the pathogen used.
3.3. Identification of Inhibitory Substances
3.3.1. Identification of Inhibitory Substances Using the Agar Well Diffusion Method
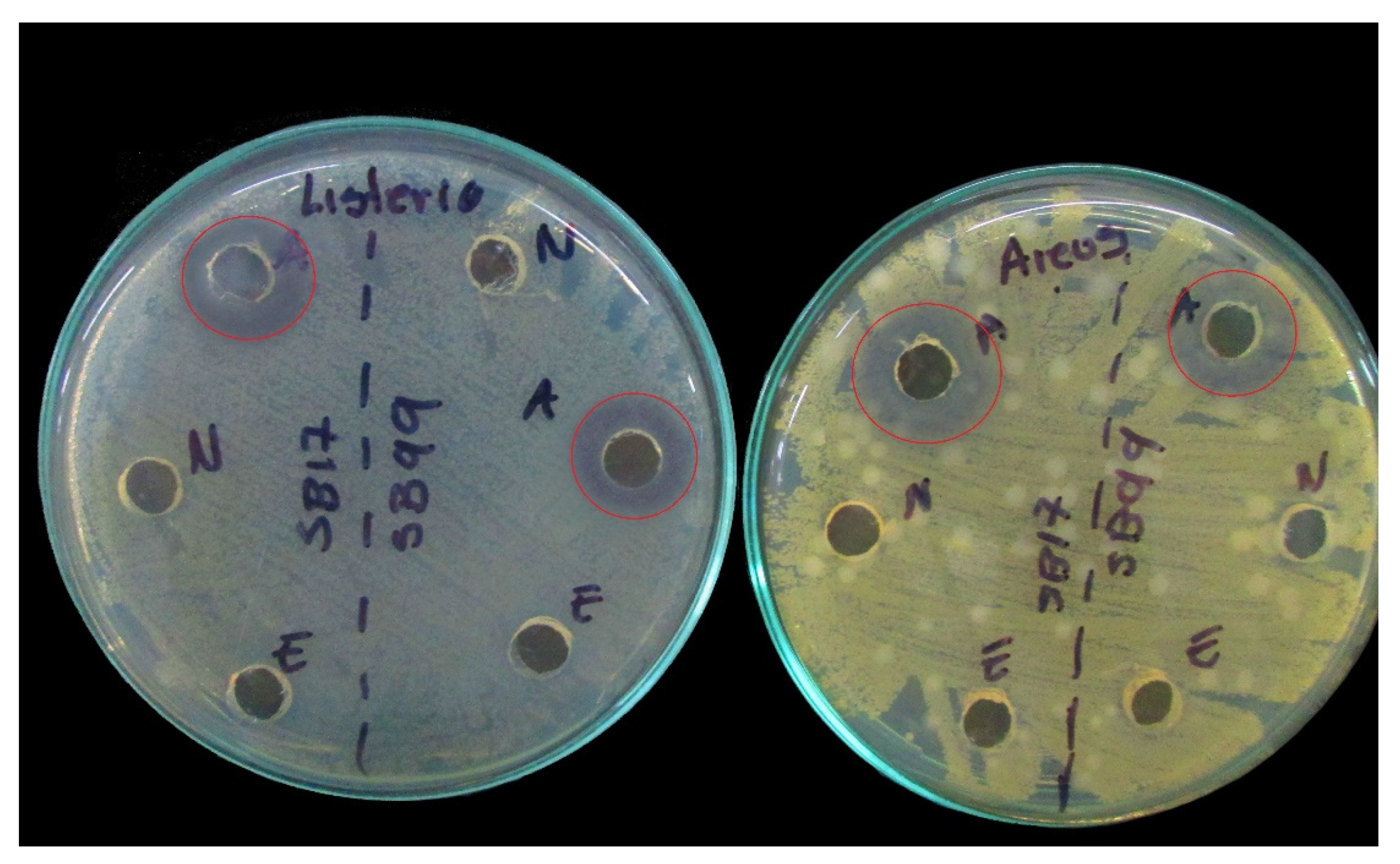
Fraction C (MRS broth) was negative for inhibiting the three pathogens tested; strain SB99, identified as Lactococcus lactis, presented inhibition of Serratia and Listeria with fraction B. Fraction A was positive for all of the strains, which shows that the inhibition was due to the action of the acids present in the CFE; see Figure 3 and Table 4, which show the pH values of fraction A.
3.3.2. Qualitative and Quantitative Estimation of Organic Acids Produced by the Isolates Detected by HPLC
Table 5 details the concentration of organic acids and shows that lactic acid was the predominant metabolite produced. Its concentration presented high variability, ranging from 1871.67 to 13613.97 mg/L. Succinic acid presented the next highest variability; with values between 100 and 163 mg/L, it showed less variation.
3.3.3. Purification and Enzyme Assay of Crude Cell Extracts
The activity of the crude extracts of bacterial cells after precipitation with ammonium sulfate was negative for all the bacterial isolates tested. When protein quantification was enhanced, there was a reduction of proteins in the fermented culture medium, indicating consumption of proteins by the isolates tested. The results are shown in Table 6 and Figure 4.
3.4. Meat Mass Testing
Determination of the Microbiological Characteristics of Chorizo
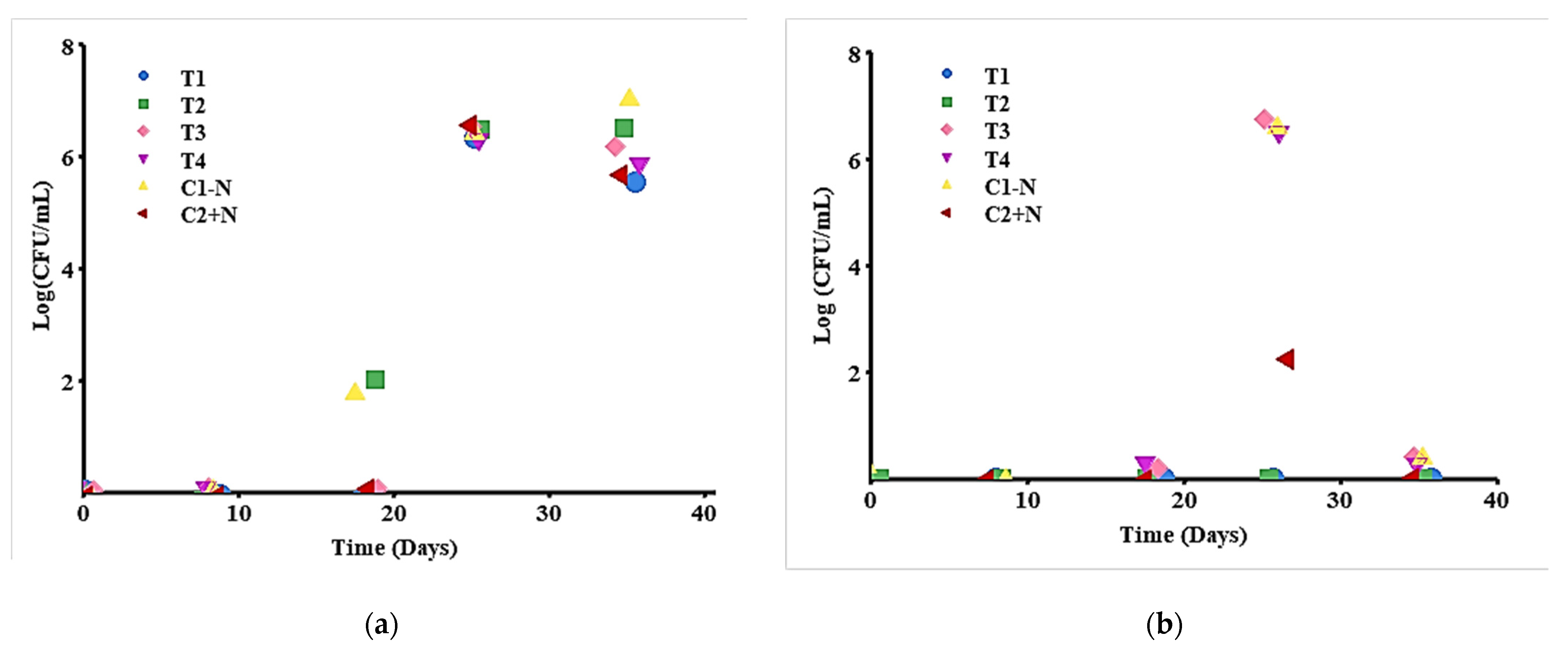
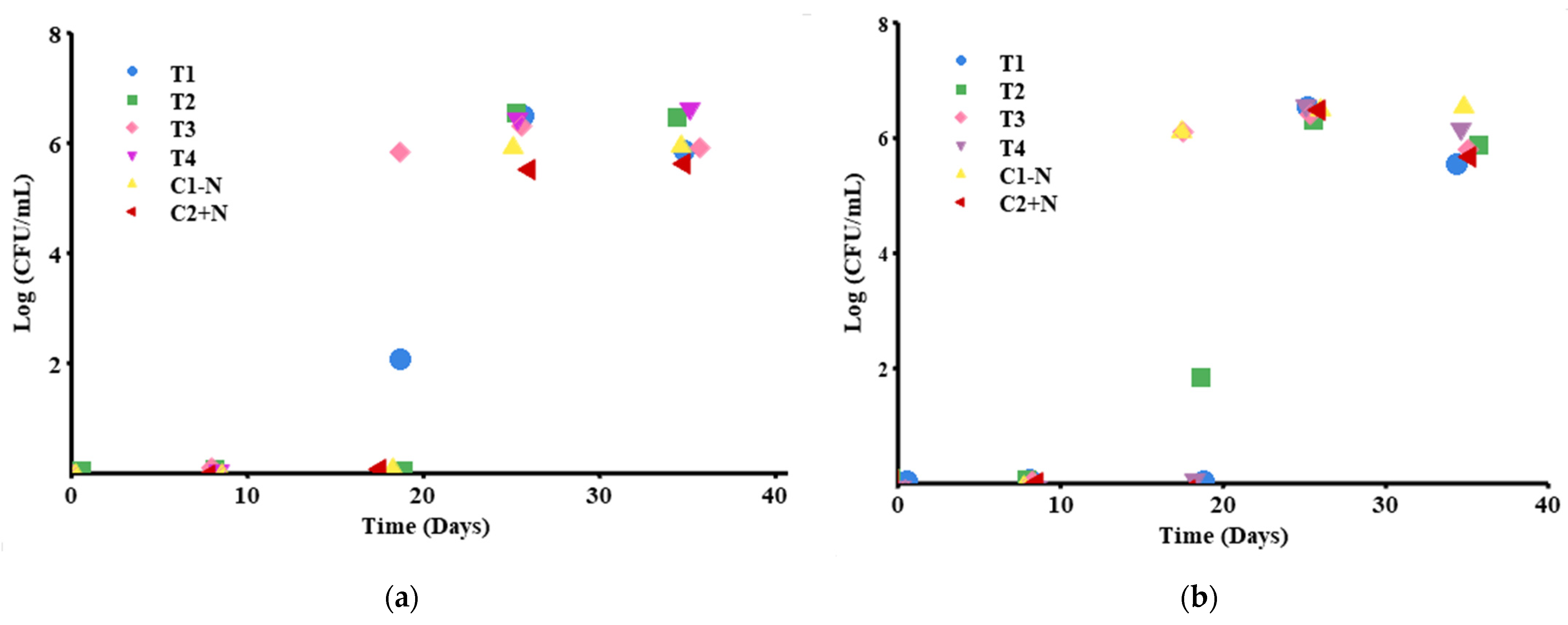
Mold and yeast counts are shown in Figure 5a. Statistically, there were significant differences between the first three days of measurement and the last two days of measurement, p < 0.005; the first days of measurement were zero, but the last two presented values between 5.6–6.5 log CFU/mL. However, no significant differences were found between treatments, p > 0.282, even though T2 (strain SB3 + nitrite and nitrate salts) and the control without nitrite salt were already showing growth of molds and yeasts at day 18.
The counts of enteric organisms such as Escherichia coli and Staphylococcus were negative. The colonies that appeared on day 26 were colorless and grew in both the treatments and the control without nitrite salt (T3, T4, and C1 − N) (see Figure 5b) and did not present significant differences (p > 0.05). T1 and T2 did not present microbiological growth during the 35-day follow-up, although the control presented growth on day 26. In this case, it was found that day 26 was statistically different (p < 0.005) from the other days of measurement, where no microbiological growth was present, p = 0.999.
Figure 6a shows the results of LAB monitoring, indicating significant differences (p = 0.0024) between T3 on day 18 and the other treatments. It should be noted that T1 presented growth on the same day, but it was not statistically significant. The treatments presented strain SB17 (Lb. plantarum) in both the absence and presence of nitrite and nitrate salts. On days 26 and 35, there were no significant differences between the treatments and controls.
In terms of time, the measurement taken on the first three days did not differ significantly (p > 0.005), but the last two measurements showed significant differences (p < 0.005). This is due to the fact that on the first days, there was zero or little growth of LAB, while in the last days, the growth was 5.49 log CFU/mL at minimum.
The presence of mesophilic microorganisms began on day 18, as shown in Figure 6b when treatment T3 (BS17 without nitrite and nitrate salts) and control C1 − N (chorizo without nitrite salts) showed differences from the rest of the treatments (p < 0.005) but not from each other. On days 26 and 35, there was no significant difference between the controls and the treatments (p > 0.005), which presented counts between 6.45 and 6.5 log CFU/mL.
In terms of time, the measurements taken on day 18 were statistically different from those taken on the other days, p < 0.005, which explains the growth of mesophilic organisms. Additionally, the measurements taken on the first two days were significantly different from those taken on the last two days, p < 0.005, since on the first days, the growth was zero, and on the last days, it was 6.5 log CFU/mL; however, there was no difference between the first two days and the last two days.
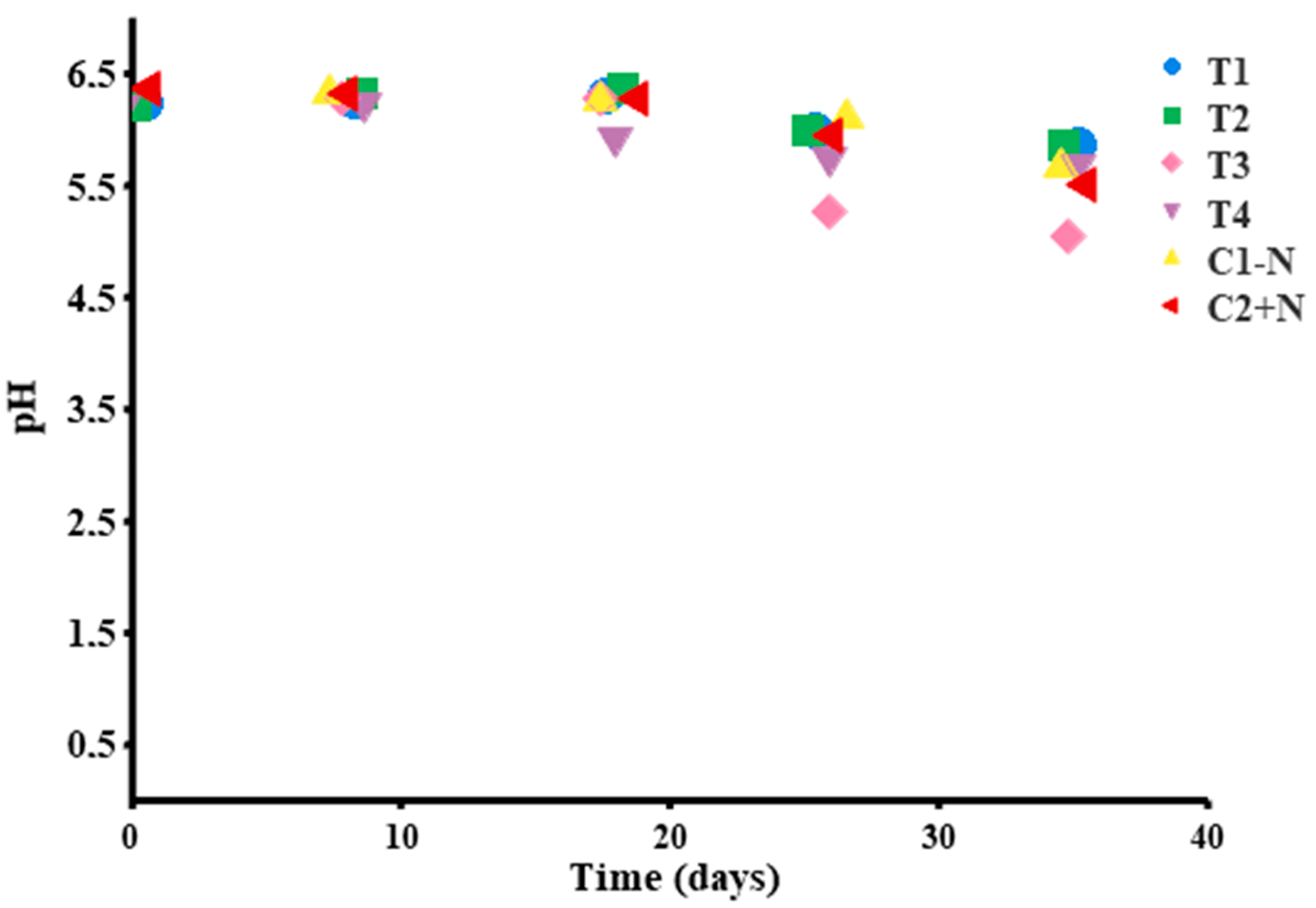
The effects of pH in the controls and treatments are shown in Figure 7. A change was evidenced during storage time, which was associated with the presence of microorganisms: The initial pH value was between 6.25 and 6.37, and the final pH values were between 5.86 and 5.52. Statistically, there was no difference in pH value between treatments (p = 0.7485); however, the pH value on day 35 was significantly different from the value on other days, p < 0.005.
4. Discussion
Ten LAB strains isolated from meat masses were used in this study for the preparation of cooked chorizo and sausages. The LAB were identified as Lb. sakei (SB3; SB5), Lb. plantarum (SB17), Lactococcus lactis (SB99), Weissella confusa (SB100; CB26), Lb. buchneri (SB112), Enterococcus faecalis (C12), Pediococcus acidilactici (C34) and Lactococcus garvieae (C54). These species have previously been isolated from different types of meats and their derived products [25,26]. Species such as Lb. Plantarum, P. acidilactici, and Lb. sakei are used commercially as starter cultures for the production of meat products since they are competitive and dominate the fermentation process [18,27,28], while E. faecalis and L. garvieae are found in the first stages of the production of artisanal fermented products [17,18,27], results that coincide with those found in this study.
The selection of LAB as bioprotective cultures should be based on the cultures’ ability to survive under stress conditions and their competitive power against indigenous microbiota. To determine this, the strains were subjected to different concentrations of NaCl. All isolates were found to be viable, although there was an interaction effect between the salt concentration and the strain used. Similar results were reported by Castro et al., 2011 [29]; Swetwiwathana and Visessanguan, 2015 [30]; Chavez-Martinez et al., 2016 [31]; Das, Khowala and Biswas, 2016 [32]; Todorov et al., 2017 [33]; and Patil, Disouza and Pawar, 2019 [34] and are due to the characteristics of each bacterial species and their resistance mechanisms.
Nitrite and nitrate salts are used to control Clostridium botulinum, and have long been credited with antimicrobial characteristics [10,18,19,35,36]. The results showed that the concentration of the salts did not exert a significantly different effect on the growth of the LAB evaluated; however, a relationship was found between the concentration and the strain evaluated, although it should be noted that the lowest growth was 4.5 log (CFU/mL). Therefore, nitrite and nitrate salts did not exert microbiological control; their use is more likely related to the organoleptic qualities that they contribute to the products [10,18,37].
When pH was evaluated, the strains presented their highest growth at pH values of 6 and 4; pH 3 affected the growth of five strains (Figure 1C). In LAB, a pH < 4 is considered an inhibitory factor that affects viability due to the reduction of intracellular pH, which diffuses through the cell membrane, causing a collapse of the electrochemical gradient that has a bacteriostatic or bactericidal effect [5,30,32]. However, there are LAB that can survive at a pH < 4 and are capable passing through the digestive tract, which is a probiotic characteristic [8,16,30,34,38].
Another important aspect to evaluate is temperature, a factor that affects the viability and survival of bacteria. The isolates were viable at temperatures up to 40 °C; at 50 °C, four isolates did not present growth, and at 60 °C, none of the isolates presented growth. Similar results were reported by Todotov et al., 2017 [33], in which at 40 °C, growth was moderate, and at 50 °C, it was weak or absent. The growth and tolerance to temperature changes shown by six of the isolates were extensive since the optimal growth temperature for LAB is between 35–37 °C [1,39,40].
In the antibiotic sensitivity test, the isolates W. confusa (CB26), Lb. buchneri (SB112), and Lb. Sakei (SB3) were resistant to tetracycline (30 µg), and the remaining eight strains were susceptible to all of the tested antibiotics. Biologically, it has been shown that bacteria transfer their resistance genes to other nearby microorganisms, contributing to a growing problem worldwide [19,24,39,41,42]. Such resistance is of great clinical interest worldwide since it confers to microorganisms the ability to colonize and cause diseases in hospital patients [2,43,44,45].
Another factor of importance for human health is the type of lactic acid isomer produced by the isolates. In infants, the use of type D (−) lactic acid, which causes acidosis, is rejected [21]. For this reason, the lactic acid produced by the isolates was identified. The two strains identified as Lb. sakei (SB3; SB5) presented the racemic mixture (DL), and the remaining ones produced the L(+) type, which has greater inhibitory power than the D type and has not been associated with any pathology [46].
When testing the antimicrobial activity of the 10 isolates against the seven test pathogens, a significant difference was found depending on the strains and the pathogen used. The pathogen with the highest susceptibility was P. vulgaris, and the most resistant pathogens were S. typhimurium and S. marcescens. The effect of LAB against Gram-negative bacteria is mainly attributed to the action of organic acids, which modify pH and hydrogen peroxide activity [19,47]. It has been found that bacteriocins are usually not very effective on this group due to the nature of their outer membrane, which acts as a permeable barrier [6,19,29,48,49].
To identify the inhibitory substances, the agar well diffusion method was performed (see Table 4). The results indicated that the inhibition was mainly due to the action of the acids present in the CFE (fraction A) since in CFE-N (fraction B), only the isolate Lactococcus lactis (SB99) inhibited S. marcescens and L. monocytogenes. These findings coincide with those reported by Angmo et al., 2016 [22] and Rahmeh et al., 2019 [2] but differ with those of Bungenstock et al., 2020–2021 [26,50] and Yazgan et al., 2021 [28], in which CFE-N presented antimicrobial activity, although not for all of the pathogenic isolates tested, which led them to conclude that crude CFE presents greater antimicrobial activity.
The organic acids produced by LAB were identified (Table 5) as hetero-fermentative due to their production of different acids and ethanol [1,19,51,52], the production of which modifies the pH and has an antimicrobial effect. Lactic acid is considered to enhance the nutritional and sensory characteristics of fermented products [40], and acetic and propionic acid function as strong inhibitory agents by acting on the cell membrane and controlling the growth of molds and yeasts [1,22,53]. Succinic acid exhibits an inhibitory power similar to that of acetic acid, although it usually acts as a buffer solution, keeping the pH stable [2,54,55,56].
As previously mentioned, the Lactococcus lactis strain (SB99) was the only one that showed activity with fraction B. Precipitation was performed with ammonium sulfate, which would allow the identification of the presence of protein compounds such as bacteriocins, but the precipitate did not show antimicrobial activity in any of the isolates. These results differ from those reported by Lü et al., 2014 [57]; Castilho et al., 2019 [58]; Rahmeh et al., 2019 [2], and Bungenstock, Abdulmawjood, and Reich, 2020 [26], in which the precipitates presented inhibitory capacity, but not for all of the evaluated pathogens. The efficiency of the ammonium sulfate precipitation technique depends on the characteristics of the peptide and the amount of antagonistic substances produced [26,59]. It is possible that the activity of SB99 was due to the presence of hydrogen peroxide, another inhibitory substance found in LAB [18,22].
When testing the effects of the isolates Lb. sakei (SB3) and Lb. plantarum (SB17) on chorizo-type sausage with and without nitrite salt, the microbiological control of molds, yeasts, and enteric organisms (Section 3.4) generally did not differ significantly between the treatments used, which is promising since the microorganisms are giving equal results to the treatments with nitro salts. But there was a significant difference in the presence of microorganisms over time, with the difference being related to packaging—specifically, whether the products were refrigerated or vacuum packed. It has been reported that the loss of vacuum sealing allows the entry of oxygen, which allows the growth of yeasts and enteric bacteria [60,61,62]; however, in this study, there were no molds, E. coli or Staphylococcus in the product.
Regarding the monitoring of LAB and mesophilic organisms, there were significant differences between the treatments and the controls. It was expected that the heat treatment of chorizo (80 °C) would eliminate most of the mesophilic organisms, including LAB, but the treatments containing the SB17 strain showed growth in both the mesophilic organisms and LAB count (T3 in both; T1 in LAB). It has been found that the natural flora and starter cultures do not present a uniform distribution but are immobilized in the cavities of the sausage mixture, which act as growth nests, multiplying over time [60]; likewise, emulsion offers protection to the inoculums when they are located in the center and thus allows them to survive the thermal process. Bacteria are also known to temporarily inactivate under unfavorable conditions (latency) [37,52,60,63], which may have occurred in the third measurement performed on day 18 in treatments T3 and T1. Meanwhile, the differences over time (between the first three measurements and the last two) in the mesophilic organism and LAB counts were similar to those found in Laszkiewicz et al., 2021 [52], in which microbiological growth is associated with the deterioration of the packaging and the contamination of the product after cooking.
Finally, the pH of the control and treatments showed a continuous decrease between evaluations over time, with no difference between treatments, which verifies the presence of microorganisms in the products. The T3 treatment, which was inoculated with the Lb. plantarum strain (SB17) at pH 5.5; Abhari et al., 2018 [62] point out that cooked products with pH values < 5.8 are expired or damaged, causing organoleptic changes that are not to the consumer’s taste. However, in fermented products, such pH levels are the optimum conditions for avoiding contamination by pathogenic or spoilage organisms [4,37,58].
5. Conclusions
The LAB isolated in this work expressed biopreservative characteristics such as pathogen inhibitory capacity and resistance to different pH, NaCl, nitrate, and nitrite salt concentrations, and temperatures.
It is concluded that the inhibitory activity of the 10 isolates was mainly due to the presence of organic acids, although the strain Lactococcus lactis (SB99) presented an inhibitory substance that could not be identified.
The addition of Lb. plantarum (SB17) and Lb. sakei (SB3) strains to the sausage had similar effects on microbiological control and pH, which is why it is necessary to measure a greater number of variables in the product.
6. Recommendations
For further studies, the performance of encapsulation tests with the isolates is recommended to achieve greater thermostability and guarantee the survival of the inocula.
An additional test of the sausages should be performed to measure variables such as color, texture, water activity, and relative humidity, which are necessary for quality-checking the product. Due to adverse conditions, it was not possible to perform these measurements in the present study.
Author Contributions
Conceptualization, C.R.-T., G.A.B. and R.S.L.-B.; methodology, R.S.L.-B. and C.R.-T.; validation, R.S.L.-B.; formal analysis, R.S.L.-B.; investigation, R.S.L.-B. and C.R.-T.; resources, C.R.-T. and G.A.B.; data curation, R.S.L.-B.; writing original draft preparation, R.S.L.-B.; writing review and editing, G.A.B., C.R.-T. and R.S.L.-B.; visualization, G.A.B. and C.R.-T.; supervision, G.A.B. and C.R.-T.; project administration, G.A.B. and C.R.-T.; funding acquisition, G.A.B. and C.R.-T. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by COLCIENCIAS (Call 745) and funded by the Universidad del Valle.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
To the Universidad del Valle for the financing of the project C.I 71112 and COLCIENCIAS; to the research group and the Laboratory of Microbiology and Applied Biotechnology (MIBIA) for the use of the facilities and advice during the execution of the project. To the imaging laboratory of the biology department for photography and photo editing services; also to the postgraduate degree in biological sciences from the Universidad del Valle for the services and consultancies provided.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Akbar, A.; Ali, I.; Anal, A.K. Industrial Perspetives of Lactic Acid Bateria for Biopreservation and food safety. J. Anim. Plant Sci. 2016, 26, 938–948. [Google Scholar]
- Rahmeh, R.; Akbar, A.; Kishk, M.; Al-Onaizi, T.; Al-Azmi, A.; Al-Shatti, A.; Shajan, A.; Al-Mutairi, S.; Akbar, B. Distribution and antimicrobial activity of lactic acid bacteria from raw camel milk. New Microbes New Infect. 2019, 30, 100560. [Google Scholar] [CrossRef] [PubMed]
- Trias, R.; Bañeras, L.; Montesinos, E.; Badosa, E. Lactic acid bacteria from fresh fruit and vegetables as biocontrol agents of phytopathogenic bacteria and fungi. Int. Microbiol. 2008, 11, 231–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yépez, A.; Luz, C.; Meca, G.; Vignolo, G.; Mañes, J.; Aznar, R. Biopreservation potential of lactic acid bacteria from Andean fermented food of vegetal origin. Food Control 2017, 78, 393–400. [Google Scholar] [CrossRef]
- Velázquez-López, A.; Covatzin-Jirón, D.; Toledo-Meza, M.D.; Gutierrez, G.V. Bebida fermentada elaborada con bacterias ácido lácticas aisladas del pozol tradicional chiapaneco Fermented drink elaborated with lactic acid bacteria isolated from chiapaneco traditional pozol. CienciaUAT 2018, 13, 165–178. [Google Scholar] [CrossRef] [Green Version]
- Jones, R.J.; Hussein, H.M.; Zagorec, M.; Brightwell, G.; Tagg, J.R. Isolation of lactic acid bacteria with inhibitory activity against pathogens and spoilage organisms associated with fresh meat. Food Microbiol. 2008, 25, 228–234. [Google Scholar] [CrossRef]
- Federici, S.; Ciarrocchi, F.; Campana, R.; Ciandrini, E.; Blasi, G.; Baffone, W. Identification and functional traits of lactic acid bacteria isolated from Ciauscolo salami produced in Central Italy. Meat Sci. 2014, 98, 575–584. [Google Scholar] [CrossRef]
- Prabhurajeshwar, C.; Chandrakanth, K. Evaluation of antimicrobial properties and their substances against pathogenic bacteria in-vitro by probiotic Lactobacilli strains isolated from commercial yoghurt. Clin. Nutr. Exp. 2019, 23, 97–115. [Google Scholar] [CrossRef] [Green Version]
- Gänzle, M. Lactic metabolism revisited: Metabolism of lactic acid bacteria in food fermentations and food spoilage. Curr. Opin. Food Sci. 2015, 2, 106–117. [Google Scholar] [CrossRef]
- Rahman, U.U.; Khan, M.I.; Sohaib, M.; Sahar, A.; Ishaq, A. Exploiting microorganisms to develop improved functional meat sausages: A review. Food Rev. Int. 2016, 33, 195–215. [Google Scholar] [CrossRef]
- Lopez, M.C.V.; Canencio, J.A.O.; Acuña, P.A.G.; Durango, A.C. Bacterias Acido Lácticas (Bal) Como Alternativa de Conservación de la Carne de Res Empacada en Atmósfera Modificada. Aliment. Hoy 2011, 20, 18–27. [Google Scholar]
- Zannini, E.; Waters, D.M.; Coffey, A.; Arendt, E.K. Production, properties, and industrial food application of lactic acid bacteria-derived exopolysaccharides. Appl. Microbiol. Biotechnol. 2016, 100, 1121–1135. [Google Scholar] [CrossRef]
- Ayyash, M.; Abushelaibi, A.; Al-Mahadin, S.; Enan, M.; El-Tarabily, K.; Shah, N. In-vitro investigation into probiotic characterisation of Streptococcus and Enterococcus isolated from camel milk. LWT 2018, 87, 478–487. [Google Scholar] [CrossRef]
- Burgos, M.J.G.; López, R.L.; Aguayo, M.C.L.; Pulido, R.P.; Gálvez, A. Bioconservación de Alimentos Cárnicos. Real Acad. Cienc. Vet. Andal. Orient. 2011, 24, 111–123. [Google Scholar]
- Deshmukh, P.V.; Thorat, P.R. Bacteriocins: A new trend in antimicrobial food packaging. Int. J. Adv. Res. Eng. Appl. Sci. 2013, 2, 1–12. [Google Scholar]
- Jurado-Gámez, H.; Gúzman-Insuasty, M.; Jarrín-Jarrín, V. Determinación de la cinética, pruebas de crecimiento y efecto de inhibición in vitro de Lactobacillus lactis en Staphylococcus aureus, Staphylococcus epidermidis, Streptococcus agalactiae y Escherichia coli. Rev. Fac. Med. Vet. Zootec. 2015, 62, 23–39. [Google Scholar] [CrossRef]
- Blanco, L. Diseño de dos Modelos de Conservación en Productos Cárnicos, Fundamentados en la Sustitución Total o Parcial de Nitrito–Nitrato. Ph.D. Thesis, Universidad de La Sabana, Bogota, Colombia, 2016. [Google Scholar]
- García-Díez, J.; Saraiva, C. Use of Starter Cultures in Foods from Animal Origin to Improve Their Safety. Int. J. Environ. Res. Public Health 2021, 18, 2544. [Google Scholar] [CrossRef] [PubMed]
- Da Costa, R.J.; Voloski, F.L.S.; Mondadori, R.G.; Duval, E.H.; Fiorentini, Â.M. Preservation of Meat Products with Bacteriocins Produced by Lactic Acid Bacteria Isolated from Meat. J. Food Qual. 2019, 2019, 4726510. [Google Scholar] [CrossRef] [Green Version]
- Maye, B.R.; Guzman, M. El Antibiograma de discos. Normalización de la técnica de Kirby-bauer. Biomédica 1984, 4, 112–121. [Google Scholar] [CrossRef] [Green Version]
- Botero SP, B.; Gamboa DC, V.; Escobar GA, B.; Toro, C.R. Detección de isó-meros del ácido láctico: Metabolitos de bacterias ácido lácticas aisladas de masas ácidas fermentadas colom-bianas. Rev. Argent. Microbiol. 2013, 45, 205–206. Available online: http://www.scielo.org.ar/scielo.php?script=sci_arttext&pid=S0325-75412013000300012&lng=es&nrm=iso&tlng=es (accessed on 28 July 2021).
- Angmo, K.; Kumari, A.; Savitri; Bhalla, T.C. Probiotic characterization of lactic acid bacteria isolated from fermented foods and beverage of Ladakh. LWT 2016, 66, 428–435. [Google Scholar] [CrossRef]
- Cenicaña—Centro de Investigación de la Caña de Azúcar. Estación Experimetnal. Determinación de Ácidos Orgánicos, Glicerol y Etanol Empleando Cromatografía HPLC-UV/IR; Protocolo de Trabajo Cali; Cenicaña: Cali, Colombia, 2015. [Google Scholar]
- Padmavathi, T.; Bhargavi, R.; Priyanka, P.R.; Niranjan, N.R.; Pavitra, P.V. Screening of potential probiotic lactic acid bacteria and production of amylase and its partial purification. J. Genet. Eng. Biotechnol. 2018, 16, 357–362. [Google Scholar] [CrossRef]
- Ajao, O.; Banwo, K.; Ogunremi, O.; Sanni, A. Antimicrobial properties and probiotic potentials of lactic acid bacteria isolated from raw beef in ibadan, nigeria. J. Microbiol. Biotechnol. Food Sci. 2018, 8, 770–773. [Google Scholar] [CrossRef]
- Bungenstock, L.; Abdulmawjood, A.; Reich, F. Evaluation of antibacterial properties of lactic acid bacteria from traditionally and industrially produced fermented sausages from Germany. PLoS ONE 2020, 15, e0230345. [Google Scholar] [CrossRef]
- Leroy, F.; Verluyten, J.; De Vuyst, L. Functional meat starter cultures for improved sausage fermentation. Int. J. Food Microbiol. 2006, 106, 270–285. [Google Scholar] [CrossRef]
- Yazgan, H.; Kuley, E.; Gökmen, T.G.; Regenstein, J.M.; Özogul, F. The antimicrobial properties and biogenic amine production of lactic acid bacteria isolated from various fermented food products. J. Food Process. Preserv. 2021, 45, e15085. [Google Scholar] [CrossRef]
- Castro, M.; Palavecino, N.; Herman, C.; Garro, O.; Campos, C. Lactic acid bacteria isolated from artisanal dry sausages: Characterization of antibacterial compounds and study of the factors affecting bacteriocin production. Meat Sci. 2011, 87, 321–329. [Google Scholar] [CrossRef]
- Swetwiwathana, A.; Visessanguan, W. Potential of bacteriocin-producing lactic acid bacteria for safety improvements of traditional Thai fermented meat and human health. Meat Sci. 2015, 109, 101–105. [Google Scholar] [CrossRef] [PubMed]
- Martínez, A.C.; De Chihuahua, U.; Estrada-Gandarilla, M.; Monterrubio, A.L.R.; Acevedo, M.A.G. Prevalence of Lactic Acid Bacteria in Sliced Cooked Ham as an Indicator of Its Shelf Life. Rev. Vitae 2016, 23, 167–172. [Google Scholar] [CrossRef]
- Das, P.; Khowala, S.; Biswas, S. In vitro probiotic characterization of Lactobacillus casei isolated from marine samples. LWT 2016, 73, 383–390. [Google Scholar] [CrossRef]
- Todorov, S.D.; Stojanovski, S.; Iliev, I.; Moncheva, P.; Nero, L.A.; Ivanova, I.V. Technology and safety assessment for lactic acid bacteria isolated from traditional Bulgarian fermented meat product “lukanka”. Braz. J. Microbiol. 2017, 48, 576–586. [Google Scholar] [CrossRef]
- Patil, A.; DiSouza, J.; Pawar, S. Shelf life stability of encapsulated lactic acid bacteria isolated from sheep milk thrived in different milk as natural media. Small Rumin. Res. 2019, 170, 19–25. [Google Scholar] [CrossRef]
- Françoise, L. Occurrence and role of lactic acid bacteria in seafood products. Food Microbiol. 2010, 27, 698–709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Maso, M.; Talamini, R.; Bosetti, C.; Montella, M.; Zucchetto, A.; Libra, M.; Negri, E.; Levi, F.; La Vecchia, C.; Franceschi, S.; et al. Red meat and cancer risk in a network of case–control studies focusing on cooking practices. Ann. Oncol. 2013, 24, 3107–3112. [Google Scholar] [CrossRef] [PubMed]
- Szymański, P.; Łaszkiewicz, B.; Kern-Jędrychowska, A.; Siekierko, U.; Kołożyn-Krajewska, D. The Use of the Mixed Bacteria Limosilactobacillus fermentum and Staphylococcus carnosus in the Meat Curing Process with a Reduced Amount of Sodium Nitrite. Appl. Sci. 2021, 11, 904. [Google Scholar] [CrossRef]
- Singhal, N.; Singh, N.S.; Mohanty, S.; Singh, P.; Virdi, J.S. Evaluation of Probiotic Characteristics of Lactic Acid Bacteria Isolated from Two Commercial Preparations Available in Indian Market. Indian J. Microbiol. 2019, 59, 112–115. [Google Scholar] [CrossRef]
- Favaro, L.; Todorov, S. Bacteriocinogenic LAB Strains for Fermented Meat Preservation: Perspectives, Challenges, and Limitations. Probiotics Antimicrob. Proteins 2017, 9, 444–458. [Google Scholar] [CrossRef]
- Păcularu-Burada, B.; Georgescu, L.A.; Vasile, M.A.; Rocha, J.M.; Bahrim, G.-E. Selection of Wild Lactic Acid Bacteria Strains as Promoters of Postbiotics in Gluten-Free Sourdoughs. Microorganisms 2020, 8, 643. [Google Scholar] [CrossRef]
- Aquilanti, L.; Garofalo, C.; Osimani, A.; Silvestri, G.; Vignaroli, C.; Clementi, F. Isolation and Molecular Characterization of Antibiotic-Resistant Lactic Acid Bacteria from Poultry and Swine Meat Products. J. Food Prot. 2007, 70, 557–565. [Google Scholar] [CrossRef]
- Mejri, L.; Hassouna, M. Characterization and selection of Lactobacillus plantarum species isolated from dry fermented sausage reformulated with camel meat and hump fat. Appl. Biol. Chem. 2016, 59, 533–542. [Google Scholar] [CrossRef]
- del Campo, C.I.M.; Gómez, H.E. Bacteias Ácido Lácticas Con Capaciadad Antagónica y Actividad Bac-tiocinogénica Aisladas de Queos Fresco. e-Gnosis 2008, 6, 1–17. Available online: https://www.redalyc.org/articulo.oa?id=73011197005 (accessed on 28 July 2021).
- Georgieva, R.; Yocheva, L.; Tserovska, L.; Zhelezova, G.; Stefanova, N.; Atanasova, A.; Danguleva, A.; Ivanova, G.; Karapetkov, N.; Rumyan, N.; et al. Antimicrobial activity and antibiotic susceptibility of Lactobacillus and Bifidobacterium spp. intended for use as starter and probiotic cultures. Biotechnol. Biotechnol. Equip. 2014, 29, 84–91. [Google Scholar] [CrossRef]
- Abushelaibi, A.; Al-Mahadin, S.; El-Tarabily, K.; Shah, N.P.; Ayyash, M. Characterization of potential probiotic lactic acid bacteria isolated from camel milk. LWT 2017, 79, 316–325. [Google Scholar] [CrossRef]
- Ghanbari, M.; Jami, M. Lactic Acid Bacteria and Their Bacteriocins: A Promising Approach to Seafood Biopreservation. Intech 2013, 32, 381–404. [Google Scholar] [CrossRef] [Green Version]
- Bromberg, R.; Moreno, I.; Zaganini, C.L.; Delboni, R.R.; De Oliveira, J. Isolation of bacteriocin-producing lactic acid bacteria from meat and meat products and its spectrum of inhibitory activity. Braz. J. Microbiol. 2004, 35, 137–144. [Google Scholar] [CrossRef] [Green Version]
- Wayah, S.B.; Philip, K. Pentocin MQ1: A Novel, Broad-Spectrum, Pore-Forming Bacteriocin From Lactobacillus pentosus CS2 With Quorum Sensing Regulatory Mechanism and Biopreservative Potential. Front. Microbiol. 2018, 9, 564. [Google Scholar] [CrossRef]
- Agriopoulou, S.; Stamatelopoulou, E.; Sachadyn-Król, M.; Varzakas, T. Lactic Acid Bacteria as Antibacterial Agents to Extend the Shelf Life of Fresh and Minimally Processed Fruits and Vegetables: Quality and Safety Aspects. Microorganisms 2020, 8, 952. [Google Scholar] [CrossRef]
- Bungenstock, L.; Abdulmawjood, A.; Reich, F. Suitability of lactic acid bacteria and deriving antibacterial preparations to enhance shelf-life and consumer safety of emulsion type sausages. Food Microbiol. 2021, 94, 103673. [Google Scholar] [CrossRef] [PubMed]
- De Martinis, E.C.; Públio, M.R.; Santarosa, P.R.; Freitas, F.Z. Antilisterial activity of lactic acid bacteria isolated from vacuum-packaged brazilian meat and meat products. Braz. J. Microbiol. 2001, 32, 32–37. [Google Scholar] [CrossRef] [Green Version]
- Łaszkiewicz, B.; Szymański, P.; Zielińska, D.; Kołożyn-Krajewska, D. Application of Lactiplantibacillus plantarum SCH1 for the Bioconservation of Cooked Sausage Made from Mechanically Separated Poultry Meat. Appl. Sci. 2021, 11, 1576. [Google Scholar] [CrossRef]
- Huertas, R.A.P. Bacterias acido lacticas:papel funcional en los alimentos. Fac. Cienc. Agropecu. 2010, 8, 95–105. Available online: http://www.scielo.org.co/pdf/bsaa/v8n1/v8n1a12.pdf (accessed on 28 July 2021).
- Magala, M.; Kohajdová, Z.; Karovičová, J.; Greifová, M.; Greif, G. Application of Lactic Acid Bacteria as Starter Culture for Tarhana Fermentation. J. Microbiol. Biotechnol. Food Sci. 2014, 3, 498–504. [Google Scholar]
- Purohit, A.; Mohan, A. Antimicrobial effects of pyruvic and succinic acids on Salmonella survival in ground chicken. LWT 2019, 116, 108596. [Google Scholar] [CrossRef]
- Moradi, M.; Molaei, R.; Guimarães, J.T. A review on preparation and chemical analysis of postbiotics from lactic acid bacteria. Enzym. Microb. Technol. 2021, 143, 109722. [Google Scholar] [CrossRef]
- Lü, X.; Hu, P.; Dang, Y.; Liu, B. Purification and partial characterization of a novel bacteriocin produced by Lactobacillus casei TN-2 isolated from fermented camel milk (Shubat) of Xinjiang Uygur Autonomous region, China. Food Control 2014, 43, 276–283. [Google Scholar] [CrossRef]
- Castilho, N.P.A.; Colombo, M.; De Oliveira, L.L.; Todorov, S.D.; Nero, L.A. Lactobacillus curvatus UFV-NPAC1 and other lactic acid bacteria isolated from calabresa, a fermented meat product, present high bacteriocinogenic activity against Listeria monocytogenes. BMC Microbiol. 2019, 19, 63. [Google Scholar] [CrossRef] [Green Version]
- Vásquez, S.M.; Suárez, H.; Zapata, S. Utilización de sustancias antimicrobianas producidas por bacterias acido lácticas en la conservación de la carne. Rev. Chil. Nutr. 2009, 36, 64–71. [Google Scholar] [CrossRef]
- Pérez-Chabela, M.D.L.; Totosaus, A.; Guerrero, I. Evaluation of thermotolerant capacity of lactic acid bacteria isolated from commercial sausages and the effects of their addition on the quality of cooked sausages. Food Sci. Technol. 2008, 28, 132–138. [Google Scholar] [CrossRef] [Green Version]
- Kargozari, M.; Moini, S.; Basti, A.A.; Emam-Djomeh, Z.; Gandomi, H.; Martin, I.R.; Ghasemlou, M.; Carbonell-Barrachina, Á.A. Effect of autochthonous starter cultures isolated from Siahmazgi cheese on physicochemical, microbiological and volatile compound profiles and sensorial attributes of sucuk, a Turkish dry-fermented sausage. Meat Sci. 2014, 97, 104–114. [Google Scholar] [CrossRef] [PubMed]
- Abhari, K.; Jafarpour, D.; Shekarforoush, S. Effects of In-Package Pasteurization on Preventing Spoilage in Emulsion Vacuum Packaged Sausages during Refrigerated Storage. Food Raw Mater. 2018, 6, 40–46. [Google Scholar] [CrossRef]
- Arras, W.; Hussain, A.; Hausler, R.; Guiot, S. Mesophilic, thermophilic and hyperthermophilic acidogenic fermentation of food waste in batch: Effect of inoculum source. Waste Manag. 2019, 87, 279–287. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Viability of the 10 native LAB at different concentrations of NaCl (A), nitrite and nitrate salts (B), and different pH values (C).
Figure 1.
Viability of the 10 native LAB at different concentrations of NaCl (A), nitrite and nitrate salts (B), and different pH values (C).

Figure 2.
Inhibition halos of pathogenic bacteria vs. native LAB.

Figure 3.
Inhibition results by cell-free extract: the effect of fraction A. Red circles indicate the effect of organic acids on Listeria monocytogenes and Staphylococcus aureus strains.
Figure 3.
Inhibition results by cell-free extract: the effect of fraction A. Red circles indicate the effect of organic acids on Listeria monocytogenes and Staphylococcus aureus strains.

Figure 4.
Results for isolate supernatants treated with ammonium sulfate.

Figure 5.
Microbiological characteristics of biopreserved T1–T4 chorizo and controls: (a) Microbiological count of molds and yeasts (PDA); (b) enteric microbiological count (MacConKey).
Figure 5.
Microbiological characteristics of biopreserved T1–T4 chorizo and controls: (a) Microbiological count of molds and yeasts (PDA); (b) enteric microbiological count (MacConKey).

Figure 6.
Microbiological characteristics of biopreserved T1–T4 chorizo and controls: (a) microbiological count of LAB (MRS); (b) microbiological count of mesophiles (PCA).
Figure 6.
Microbiological characteristics of biopreserved T1–T4 chorizo and controls: (a) microbiological count of LAB (MRS); (b) microbiological count of mesophiles (PCA).

Figure 7.
pH monitoring for biopreserved chorizo treatments T1–T4 and controls.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Identification of the native LAB isolates used.
| LAB Strain | Identification | LAB Strain | Identification |
|---|---|---|---|
| SB3 | Lactobacillus sakei (MW073315) | SB112 | Lactobacillus buchneri (99.8%) 1 |
| SB5 | Lactobacillus sakei (MW073316) | CB26 | Weissella confusa (MW073317) |
| SB17 | Lactobacillus plantarum (MW073312) | C12 | Enterococcus faecalis (MW073313) |
| SB99 | Lactococcus lactis (96%) 1 | C34 | Pediococcus acidilactici (82%) 1 |
| SB100 | Weissella confusa (MW073314) | C54 | Lactococcus garvieae (MW073318) |
The numbers in parentheses indicate the GenBank code. 1 Identification according to biochemical test results (API 50CHL).
Table 2.
Formulation for the manufacture of chorizo-type sausage.
| INGREDIENT | Control (Nitrite Salt) (C2 + N) | Treatment 1 and 2 | Control (−Nitrite Salt) (C1 − N) | Treatments 3 and 4 |
|---|---|---|---|---|
| Water (ice) | 48.17 | 48.17 | 48.26 | 48.26 |
| Pulverized garlic | 4.82 | 4.82 | 4.83 | 4.83 |
| Cassava starch | 8.03 | 8.03 | 8.04 | 8.04 |
| Ascorbic acid | 0.96 | 0.96 | 0.97 | 0.97 |
| Beef (ground) | 289.02 | 289.02 | 289.58 | 289.58 |
| Scallions | 48.17 | 48.17 | 48.26 | 48.26 |
| Orange coloring | 0.32 | 0.32 | 0.32 | 0.32 |
| Chorizo seasoning | 6.74 | 6.74 | 6.76 | 6.76 |
| Monosodium glutamate | 0.80 | 0.80 | 0.80 | 0.80 |
| Pork fat | 81.89 | 81.89 | 82.05 | 82.05 |
| Liquid smoke (mL) | 1.45 | 1.45 | 1.45 | 1.45 |
| Nitrous salts ** | 0.96 | 0.96 | 0.00 | 0.00 |
| Soy protein | 6.42 | 6.42 | 6.44 | 6.44 |
| Refined salt | 2.25 | 2.25 | 2.25 | 2.25 |
| Inoculum | 0.00 | 15.00 | 0.00 | 15.00 |
Two controls were performed in the presence ** and absence of nitrite and nitrate salts; two treatments in the presence of LAB, T1 ** and T3 containing strain SB17 and T2 ** and T4 containing strain SB3 in the presence and absence of nitrite and nitrate salts, respectively.
Table 3.
Antibiotic resistance of bacterial isolates and type of lactic acid produced.
| Strain | Tetracycline | Amoxicillin | Penicillin | Ampicillin | LAT |
|---|---|---|---|---|---|
| SB3 | R | S | S | S | DL |
| SB5 | S | S | S | S | DL |
| SB17 | S | S | S | S | L (+) |
| SB99 | S | S | S | S | L (+) |
| SB100 | S | S | S | S | L (+) |
| SB112 | R | S | S | S | L (+) |
| C12 | S | S | S | S | L (+) |
| C34 | S | S | S | S | L (+) |
| C54 | S | S | S | S | L (+) |
| CB26 | R | S | S | S | L (+) |
R (resistant), S (susceptible), LAT (lactic acid type) and DL (racemic mixture).
Table 4.
Inhibition of three pathogens by isolate supernatants.
| Strain | Fraction A | Fraction B | ||
|---|---|---|---|---|
| L. monocytogenes | S. aureus | S. marcescens | ||
| SB112 | 3.77 | − | − | − |
| C12 | 4.53 | − | − | − |
| SB17 | 3.87 | − | − | − |
| SB5 | 4.29 | − | − | − |
| CB26 | 4.53 | − | − | − |
| SB99 | 4.72 | + | + | − |
| C54 | 4.39 | − | − | − |
| C34 | 4.48 | − | − | − |
| SB3 | 4.57 | − | − | − |
| SB100 | 4.37 | − | − | − |
Inhibition negative (−) and positive (+) cell-free extract (Fraction A) pH values.
Table 5.
Quantification of organic acids produced by LAB isolated from meat products.
| Sample | Citric (mg/L) | Succinic (mg/L) | Lactic (mg/L) | Acetic (mg/L) | Propionic (mg/L) | Ethanol (%) |
|---|---|---|---|---|---|---|
| SB3 | ND | 124.13 ± 0.03 | 7801.76 ± 0.01 | ND | ND | 0.22 |
| SB5 | ND | 151.47 ± 0.1 | 7704.21 ± 0.01 | ND | ND | 0.20 |
| SB17 | ND | 157.90 ± 0.1 | 1871.67 ± 0.01 | ND | 130.35 ± 0.002 | 0.67 |
| SB99 | ND | 160.89 ± 0.01 | 13,613.91 ± 0.01 | 94.36 ± 0.01 | ND | 0.23 |
| SB100 | ND | 156.00 ± 0.3 | 8828.47 ± 0.002 | ND | ND | 0.64 |
| C12 | ND | 163.49 ± 0.01 | 7663.85 ± 0.01 | 1885.77 ± 0.007 | ND | 0.49 |
| C34 | ND | 148.26 ± 0.6 | 5719.51 ± 0.03 | ND | ND | 0.22 |
| C54 | ND | 100.82 ± 0.15 | 5070.16 ± 0.06 | 4.71 ± 0.001 | ND | 0.22 |
| CB26 | ND | 159.54 ± 0.01 | 7823.55 ± 0.01 | 2000.29 ± 0.01 | ND | 0.57 |
Not detected (ND).
Table 6.
Quantification of proteins produced by LAB isolated from meat products.
| Strains | Protein Concentration (mg/L) |
|---|---|
| SB3 | 152.462 |
| SB5 | 155.54 |
| SB17 | 170.15 |
| SB99 | 142.46 |
| SB100 | 211.69 |
| CB26 | 159.89 |
| C12 | 168.61 |
| C34 | 157.85 |
| C54 | 147.85 |
| Control | 211.69 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Lucumi-Banguero, R.S.; Ramírez-Toro, C.; Bolívar, G.A. Potential Use of Lactic Acid Bacteria with Pathogen Inhibitory Capacity as a Biopreservative Agent for Chorizo. Processes 2021, 9, 1582. https://doi.org/10.3390/pr9091582
AMA Style
Lucumi-Banguero RS, Ramírez-Toro C, Bolívar GA. Potential Use of Lactic Acid Bacteria with Pathogen Inhibitory Capacity as a Biopreservative Agent for Chorizo. Processes. 2021; 9(9):1582. https://doi.org/10.3390/pr9091582
Chicago/Turabian StyleLucumi-Banguero, Ruby Stella, Cristina Ramírez-Toro, and German A. Bolívar. 2021. "Potential Use of Lactic Acid Bacteria with Pathogen Inhibitory Capacity as a Biopreservative Agent for Chorizo" Processes 9, no. 9: 1582. https://doi.org/10.3390/pr9091582
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.