
Asymmetry of Plant Cell Divisions under Salt Stress



1
Plant Cell Biology Laboratory, All-Russia Research Institute of Agricultural Biotechnology, Timiryazevskaya 42, 127550 Moscow, Russia
2
N.V. Tsitsin Main Botanical Garden of Russian Academy of Sciences, 127276 Moscow, Russia
*
Author to whom correspondence should be addressed.
Symmetry 2021, 13(10), 1811; https://doi.org/10.3390/sym13101811
Submission received: 1 August 2021
/
Revised: 22 September 2021
/
Accepted: 24 September 2021
/
Published: 28 September 2021
(This article belongs to the Special Issue Molecular Mechanisms Regulating Asymmetry in Crop Plants Embryogenesis and Growth)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Salt stress causes several damaging effects in plant cells. These commonly observed effects are the results of oxidative, osmotic, and toxic stresses. To ensure normal growth and development of tissues, the cellular compartments of multicellular plants have a unique system that provides the specified parameters of growth and differentiation. The cell shape and the direction of division support the steady development of the organism, the habit, and the typical shape of the organs and the whole plant. When dividing, daughter cells evenly or unevenly distribute the components of cytoplasm. Factors such as impaired osmotic regulation, exposure to toxic compounds, and imbalance in the antioxidant system cause disorders associated with the moving of organelles, distribution transformations of the endoplasmic reticulum, and the vacuolar compartment. In some cases, one can observe a different degree of plasmolysis manifestation, local changes in the density of cytoplasm. Together, these processes can cause disturbances in the direction of cell division, the formation of a phragmoplast, the formation of nuclei of daughter cells, and a violation of their fine structural organization. These processes are often accompanied by significant damage to the cytoskeleton, the formation of nonspecific structures formed by proteins of the cytoskeleton. The consequences of these processes can lead to the death of some cells or to a significant change in their morphology and properties, deformation of newly formed tissues and organs, and changes in the plant phenotype. Thus, as a result of significant violations of the cytoskeleton, causing critical destabilization of the symmetric distribution of the cell content, disturbances in the distribution of chromosomes, especially in polyploid cells, may occur, resulting in the appearance of micronuclei. Hence, the asymmetry of a certain component of the plant cell is a marker of susceptibility to abiotic damage.
1. Introduction
A living cell has special properties that provide the possibility of its dynamic development, providing the very possibility of the existence of multicellular organisms—tropism, or, in other words, sensitivity [1]. A stable system of a living organism, starting from a single cell, tends towards an ideally expedient and effective form—a sphere. However, such organisms are found mainly when we study single cells in a liquid medium [2,3]. Under such conditions, the sensitivity to a number of environmental factors, such as geotropism, phototropism, or chemotropism, is reduced or not noticeable. Even under these conditions, we are still faced with a change in the rounded shape to oval or cylindrical, and, in some cases, more complex, for example, spiral formations [4]. However, the life of plants, as multicellular organisms, is associated with the peculiarities of increased sensitivity to tropisms of various kinds as a result of the existence on the border of two environments, and, in some cases, even three environments simultaneously (air, water, and land). The stability and inviolability of an optimal habitat for plants is very deceptive and is regulated by the availability of resources necessary for their development and/or their limitation. Thus, we are faced with the justified need to modify the body of a plant to ensure its existence and reproduction [5]. It would seem that a rounded cell should divide endlessly in an ideal environment, ensuring sequential division with perpendicular primary and subsequent divisions, and form a multicellular sphere with a conditionally infinite number of symmetry planes. However, in reality, we observe the formation of a wide range of various organs, with a very different specialization, possessing, in different applications, both bilateral (for example, leaf, petiole) and organs with central symmetry (root, stem, primary and secondary meristems), while the entire body of a plant can possess different properties and types of symmetry and asymmetry at different levels and differ at different stages of ontogenesis [6].
This situation has a logical explanation, as access to resources for cells located inside the “ideal sphere” will be severely limited, and such cells will be unviable at some stage. Since, in the case of cell death and destruction, there is a high probability of toxic damage and the attack of pathogens, evolutionarily, it has been found to be more expedient to acquire a new skill—the mechanism of programmed cell death. This can be observed, for example, during the formation of aerenchyma in root systems with central symmetry, where parts of parenchyma cells undergo death in order to provide roots an important resource—air for respiration (the most important process ensuring the transformation of chemical energy accumulated by a plant as a result of photosynthesis, which is a source of maintenance of non-photosynthetic organs of plants) [7,8]. Moreover, specific processes of targeted death provide the formation of vascular tissues, changes in cap cells, falling leaves and the dying-off of roots, and the death of individual maternal tissues in the process of generative development [9]. Thus, various types of asymmetry, along with symmetry, are observed in the cell, tissue, organ, and the whole plant, which is constantly exposed to gravity and other environmental factors, including stress factors (as a special case). These factors constantly change and provide a balance between the processes that ensure the maintenance conditional order (symmetry) and conditional chaos (asymmetry). Significant disturbances in this balance will lead to irreversible consequences, fraught with the death of an individual organism, or the extinction of a species or population. It should be understood that, unlike the cells that make up the bodies of animals, plant cells are much more interconnected due to their specific structure. Almost all plant cells, with the exception of cells that have passed to terminal differentiation, are interconnected by specific formations—plasmodesmata. For example, due to plasmodesmata, the rate of calcium signal transduction may be multiplied and comparable to the transduction rate in the animal nervous system [10,11]. Thus, we can consider that the plant organism is a single system—a coenocyte [12,13]. However, the presence of significant limitations in the transmission of regulatory molecules through the symplast of the cytoplasm is constrained by the sequence of arrangement of plasmodesmata. Thus, the cells in a one-cell layer can have a large number of plasmodesmata distributed over the entire surface area of neighboring cells in one layer, but may not have plasmodesmata with adjacent cells of underlying tissue. Moreover, a number of tissues have a special modification of cell walls that prevents contact through the apoplast—for example, endoderm cells with waterproof Casparian strips [14].
The cytoskeleton is a unique and sensitive system of cells capable of qualitatively providing trophic reactions (in the form of a specific enhancement or limitation, depending on the interaction of an individual organ or cell fragment). Similar to other components of cells, it exists in the range of constant fluctuations of temperature, humidity, pressure, helio- and gravitropism, constant and variable magnetic fields, as well as other wave phenomena, such as light and sound, which a researcher, due to the limited systems of perception and recognition during the performance of experiments, takes into account only partially [15,16,17]. In plant cells, the functions of the cytoskeleton, represented by actin filaments and tubulin microtubules, generally coincide with functions in other eukaryotes. However, there are significant differences; for example, in plant cells there is no centriole associated with microtubules (it is replaced by a cell center with an indefinite structure), with the exception of unique plants that acquire the ability to form similar structures when the flagellum of the male gametophyte is laid. Another feature of the plant cytoskeleton (corresponding to the interphase stage) is its participation in the formation of cell walls, both during their formation in the late telophase in the form of a phragmoplast, and when modifying the cell wall of the vascular bundle cells, in which it becomes necessary to stimulate the formation of protrusions [18]. Other important functions of the cytoskeleton (manifested during the transition to mitosis and directly during mitosis and meiosis) are the formation of a preprophase band that determines the location of the division spindle, as well as securing the centromeric region of chromosomes through the kinetochore and the distribution and movement of chromosomes during the distribution of material between daughter cells [19,20]. Disturbances of the tubulin cytoskeleton can lead to both reversible and irreversible consequences, depending on the duration, phase, and intensity (dose) of the active factor [21,22]. The actin cytoskeleton promotes the fast and efficient intracellular transport and movement of organelles, which, in the event of damage, leads to the displacement or complete blockage of associated processes.
Salt stress is one of the simplest model systems for studying impairing impacts on plants. Salinity effects are well studied and described in detail both from the point of view of plant physiology and from the point of view of changes in individual cellular organelles and their ultrastructure [23,24,25]. Cells of various plant tissues and organs were found to be highly sensitive to the stress effects accompanying the action of salt, causing osmotic, oxidative, and toxic stresses. All cellular compartments undergo significant modification, which causes a change in the growth rate, violation of division, modification of the fine structure, and redistribution of organelles and compartments, including vacuoles [23]. In addition, for the early stages of salt exposure, the effect of plasmolysis has been described, which is caused by the action of external osmotic pressure and causes a reversible compression of the cell cytoplasm [26]. Most important, however, is the apparent disturbance in the growth, habitus, and shape of the organs of the exposed plants. For example, seedlings under a salt influence can form roots with a pronounced spiral structure. A number of these processes are accompanied and ensured by specific or nonspecific modification of the fine structure, affecting the regulation of expansion processes, as well as cell division [27,28]. Obviously, the sensitivity of an individual element can lead to a disturbance in the symmetry and asymmetry ratio, as a kind of algorithm of a given order. On the other hand, assessing the value (as adaptive potential) and danger (as an adverse effect) of such violations presents certain difficulties, except for the differences between death and survival.
In this work, only those effects will be demonstrated that, in our opinion, have a significant effect on the balance of symmetry and asymmetry, which ensures the maintenance of organ (tissue) habitus and development and leads to significant, but not fatal, damage to cell systems at the meso- and ultrastructure levels. The goal of this study is to demonstrate the processes caused by a shift in the symmetry/asymmetry ratio towards asymmetry while maintaining the viability of the organism, and borderline states when the viability is preserved but stable development and the transition to the generative phase can be limited by significant injuries and delaying or cessation of development.
2. Materials and Methods
2.1. Plant Material
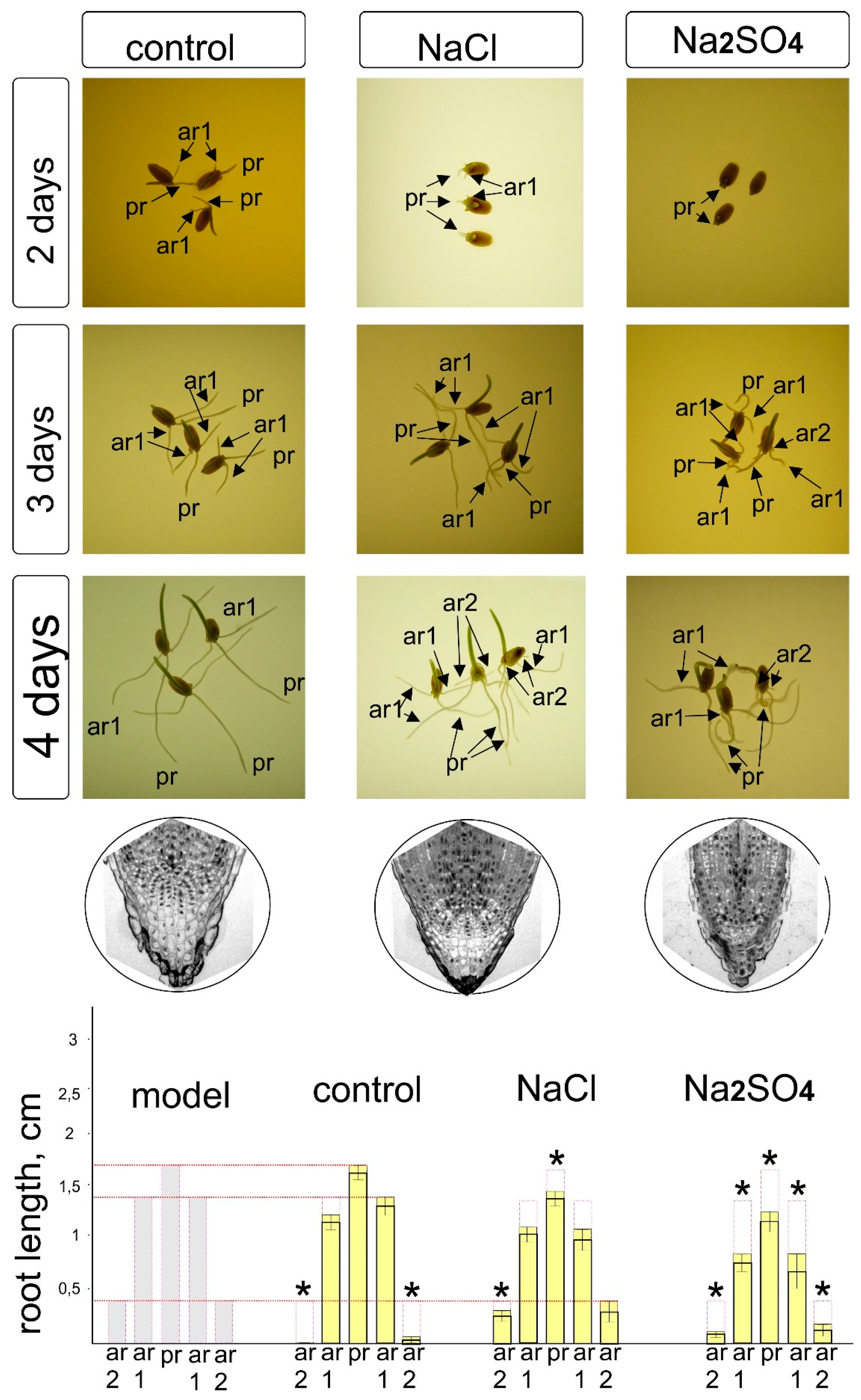
Grains (kernels) of winter wheat Triticum aestivum L. cv Moskovskaya 39 were imbibed in salt solutions or water between two layers of moistened filter paper in Petri dishes (25 kernels per dish) in four replicates. The seeds were preliminarily analyzed and the largest kernels without obvious mechanical damage were selected with uniform right and left moieties, uniform color and pubescence, and without traces of damage by insects or fungi. For soaking and germination, solutions of the following composition—0.1 M NaCl, 0.1 M Na2SO4—and water as a control were used. Each variant of treatment was carried out in four replicates, with 25 kernels per replicate. The germination temperature was 18–20 °C. On days 2, 3, and 4, plant seedlings were photographed and the length of the main and lateral roots (if any) was measured.
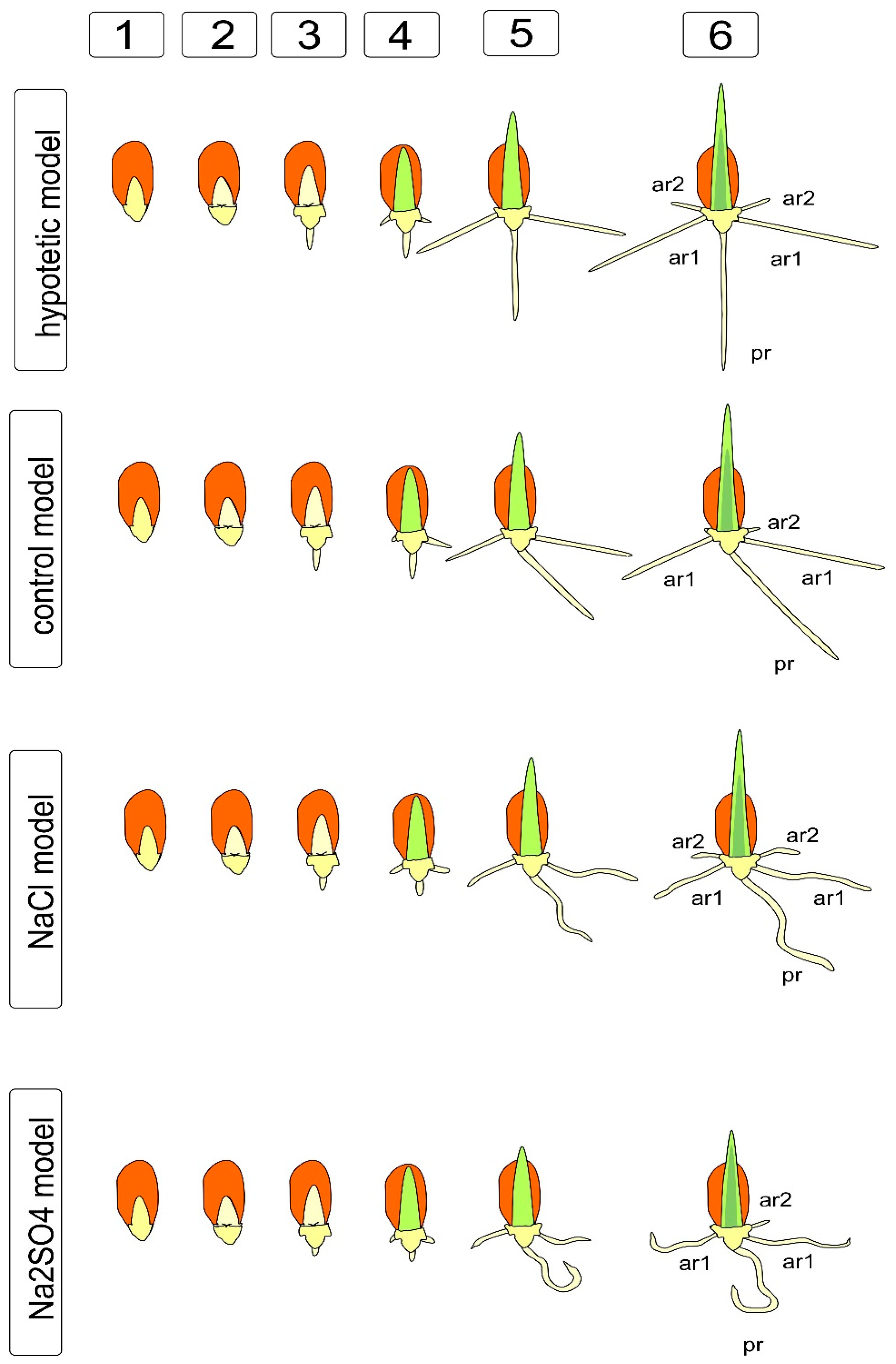
2.2. Arraying of an Ideal Model of Juvenile Seedling
To identify changes in symmetry/asymmetry in the development of wheat seedlings, we made an assumption that the maximum elongation is an option for the optimal implementation of the germplasm potential. For comparison, the maximum indicators achieved in the experiment were used.
2.3. Immunodetection of Cytoskeleton Transformations
The main root of 4-day-old seedlings was used as a model to reveal the characteristic lesions of the microtubular cytoskeleton. For immunofluorescent identification of microtubules, a double staining method was used: antibodies to mouse β-tubulin (clone DM1A) and FITC-conjugated anti-mouse IgG, Alexa-conjugated anti-rat IgG (Life Technologies, Bourgoin-Jallieum, France) [29]. To fix and obtain the preparations of macerated cells, 0.5 cm of the root of each plant was cut off and fixed in a solution of 4% paraformaldehyde in PHEM buffer pH 6.9 (60 mM PIPES, 25 mM HEPES, 10 mM EGTA, 2 mM MgCl2) for 2 h at room temperature (18–20 °C). Then, the fixative was washed in the same buffer. The meristem zone (2–3 mm) was separated and placed in a macerating solution containing 2% cellulase solution (Sigma-Aldrich, St. Louis, MO, USA) in Na–acetate buffer pH 5.0 for 30 s, after which the treated roots were accumulated in PHEM-buffer. The preparations were obtained by dividing them into individual cells using a metal dissecting needle, placed in 0.2% gelatin and dried in air in a refrigerator at +4 °C. Then, the preparations were placed in 0.5% Triton X-100, prepared on PHEM containing 5% DMSO, for 30 min, washed in YutM Mg-Tris-Cl buffer (pH 7.6) and incubated overnight at room temperature with monoclonal antibodies to β-tubulin, mouse (clone DM1A) in a humid chamber to prevent drying out. After this, the samples were washed in 10 mM Mg-Tris-Cl buffer (pH 7.6) and incubated in 20 mM Mg-Tris-Cl buffer containing 0.1% BSA (pH 8.2). Root cell preparations were then incubated with FITC-conjugated anti-mouse IgG, Alexa-conjugated anti-rat IgG, TRITC-conjugated anti-rat IgG for 45 min at 37 °C. Cells were stained with DAPI (4′,6-diamidino-2-phenylindole) and placed into the Vectashield mounting medium (Vector Laboratories, Burlingame, CA, USA). The obtained preparations were analyzed in an Olympus light microscope equipped with epifluorescent lighting, and a standard set of filters ×40 and ×100 objectives were used. The images were recorded using a DC digital camera, the Kodak 260, and analyzed using Corel 12.0 software. According to the deviation in the orientation of microtubules in the cortical cytoskeleton of cells for 4-day-old seedlings, the deviation in the preservation of the assumed and observed parameters of symmetry/asymmetry due to the location and presence of deformities was assessed.
2.4. Arraying of an Ideal Model of Cytoskeleton
To identify changes in symmetry/asymmetry in the dynamics of transformations of the microtubular cytoskeleton of wheat seedlings, the routine description of transformations of the cytoskeleton in the cell cycle was used. Assumption that a significant violation should be considered a chaotic arrangement that differs from the expected norm by an increase/decrease in the length and/or shape of visible elements, as well as a significant violation of the location relative to that observed and described by the ideal model, in which elements are noted that do not correspond to a typical cytoskeleton for a given stage cycle, duplicating elements that are not characteristic, the presence of heterogeneous chaotically located elements that are not characteristic of the ideal and observed in control models. For comparison, we used the maximum indices differing from the “norm” achieved in the experiment. Due to the fact that the number of studied cells in anaphase and prometaphase was insufficient, the results for these phases should be considered preliminary (provisionally correct).
2.5. Transmission Electron Microscopy of Root and Shoot Meristem Cells
To analyze the tissue ultrastructure, qualitative analysis was used. In this case, central longitudinal ultrathin sections obtained from at least five roots of independent plants for each of the experimental treatment were used. On the second day after soaking in salt solutions, the shoot apexes and the tips of the main root, prepared under a light microscope, were fixed with 2.5% glutaraldehyde in a phosphate buffer with the addition of sucrose (pH 7.2) for 24 h at 4 °C, followed by additional fixation with 1% OsO4 for 2 h. After dehydration in ethanol solutions, the samples were placed in propylene oxide and embedded in an epon-araldite (Merck, Darmstadt, Germany) mixture. After adding the catalyst and removing the residual solvent, the preparations were polymerized in two stages in a thermostat at a temperature of 45 °C (24 h) and 60 °C (24 h). Ultrathin sections were prepared using at least 3 independent samples using an LKB-3 (LKB, Stockholm, Sweden) microtome, placed on copper grids covered with formvar film and contrasted with uranyl acetate and lead citrate. Preparations were analyzed and photographed using a H-500 electron microscope (Hitachi, Ibaraki, Japan).
2.6. Arraying of an Ideal Model of Ultrastructure Elements
To identify changes in symmetry/asymmetry in the development of wheat seedlings, we made an assumption that the maximum growth in length is a variant of the optimal realization of the inherent potential. For comparison, the maximum indicators achieved in the experiment were used.
2.7. Statistical Analysis
The mean values were obtained, including three replicates from four biological replicates, and each biological replicate comprised 25 seedlings. The differences were assessed by comparing the sample means. Moreover, the significance of these differences using the Student’s test at the 5% significance level in Microsoft Excel software was assessed. The measure of discrepancy between empirical and theoretical data (ideal model) was performed using the Pearson chi-square test.
3. Results and Discussion
Peculiarities of the asymmetry of seedling development in normal conditions and during salt stress impact in monocotyledonous plants, as exemplified by soft wheat, were studied. Bread or spring wheat belongs to the widespread genus Triticum, which is the most widespread object of cultivation and consumption of products obtained from it. Belonging to an extensive family of cereals, it has unique characteristics of high yield, yield stability, and high resistance to adverse factors. The development of this culture, as with most agricultural crops, is associated with its predictability and manufacturability, since, as a result of selection, the height of the plants changes insignificantly even with a lack of moisture, the arrangement of ears is characterized by the absence of tiering, and the seeds ripen at the same time and have a slight difference in size and, accordingly, in composition. This also determines the fact that its seeds also consistently provide parameters important for the initial stages of development: uniformity of development, starting from swelling, pecking, and seedling development. It is obvious that such indicators can involve a rigid correction of unwanted deviations from the given (desired) standard. Thus, this culture is a convenient object for studying the relationship between symmetry and asymmetry in plants.
To identify the effects of asymmetry, we found it useful to use the salt effect, which, on the one hand, is a well-known damaging factor that causes the manifestations of oxidative, osmotic, and toxic effects [30,31], accompanied by various instruments for correcting injuries. On the other hand, the effect of salts is characterized as positive, with the property of leveling the uneven germination, imparting nonspecific resistance to other abiotic stresses, due to the nonspecific protective response [32] and the priming effect, which ensures more uniform seed development at an early stage [33]. To simplify the study, we considered the earliest, juvenile stage of development of wheat plants: seedlings before the first leaf emerges from the coleoptile and before the appearance of third-order adventitious lateral roots characteristic of bread wheat [34]. Observations of the development of seeds in water and aqueous solutions of NaCl and Na2SO4 revealed the absence of symmetry in the appearance of roots, unevenness in appearance, as well as not only the presence of a significant growth lag in the presence of salts, but also a change in the shape of the main and adventitious roots (Figure 1).
In addition, it was shown that salts cause a change in the development of the root cap (which, under the action of Na2SO4, had fewer layers and was deformed), which probably contributes to a visible disturbance of gravitropism, due to the redistribution of sugars and or providing the growth of auxins [24,35]. The same effect apparently causes serious disturbances in the morphology and branching of roots at a later stage. We have previously described similar effects in comparative experiments on alfalfa with NaCl, Na2SO4, and mannitol [36]. Changes in the growth, branching, and shape of roots are quite often detected under the action of abiotic factors, particularly salinity. The histograms (Figure 1) show significant differences in the length of the main and lateral roots under the influence of salts, confirming the effects of using the balancing effect during priming in brewing and preparing seeds for sowing [37], while it is obvious that this effect is associated with the inhibition of growth and, in the case of Na2SO4, with significant deformation of the root shape. The differences in the real picture of germination shown in Figure 1 are reflected in the diagram, where it becomes obvious that the ideal hypothetical model differs from the actually observed changes in morphology. Even in distilled water, the seedlings showed a rather interesting deviation from the ideal model (Figure 2), indicating that, at least for this model and this experimental approach, the asymmetry of development is the norm and should probably be provided by the uneven formation of the seed and embryo, as well as ways of transporting nutrients from the endosperm through the scutellum and further to the primary and embryonic roots. Interestingly, we noted the presence of an angle between the radicle and the first of the developing adventitious roots, which persists for some time after germination, as well as a violation of the tropism and orientation of part of the roots. Such disturbances result from osmotic damage to cells and most likely lead to a disruption of cell expansion and division, which are the constituents of tissue growth. This disorder is a consequence of the cytoskeleton converting, the fusion of vacuoles and the transformation of the endoplasmic reticulum and the Golgi apparatus (dictyosomes). In addition, there are violations in the division of DNA-containing cell compartments, such as nuclei, plastids and mitochondria.
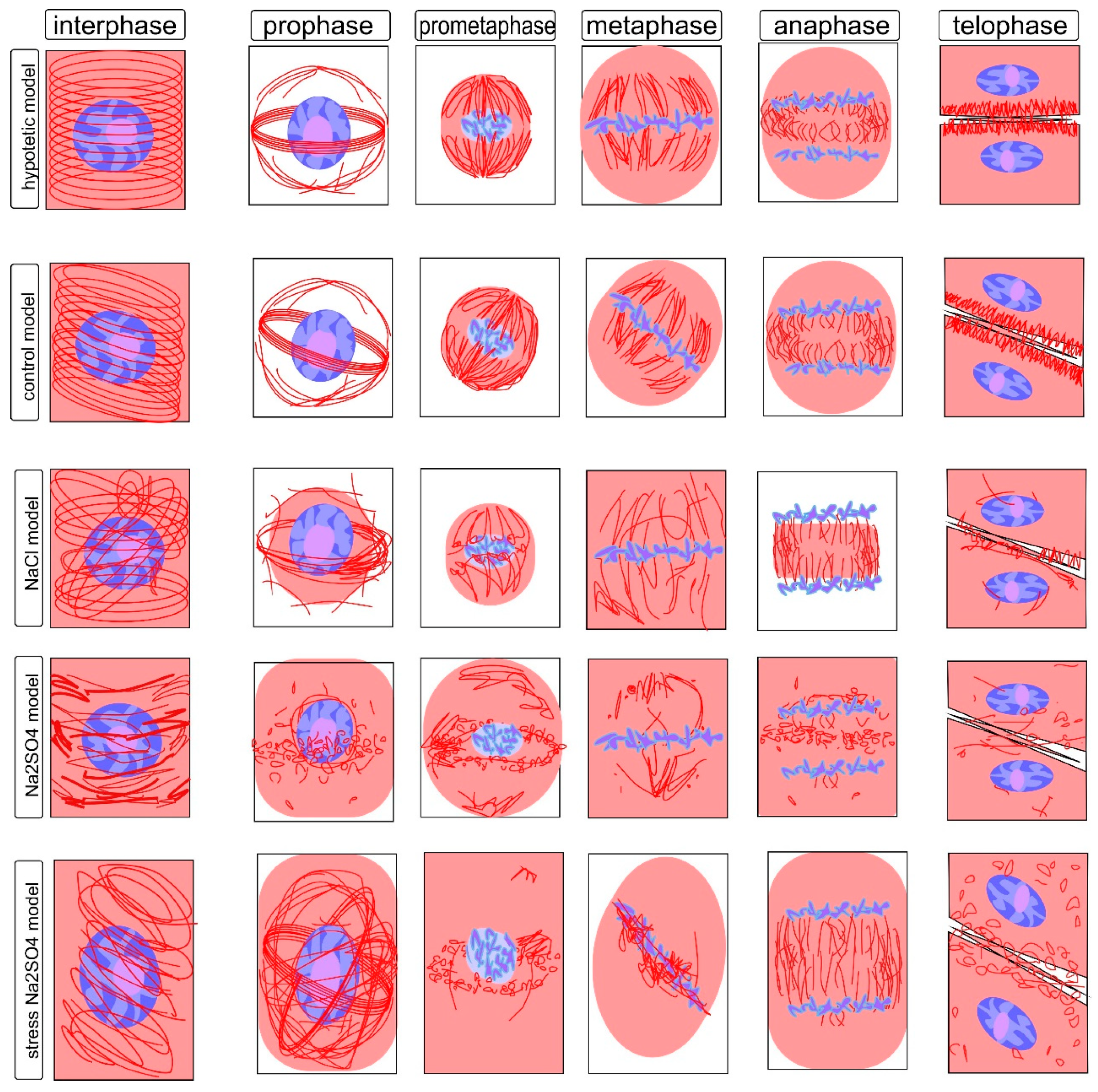
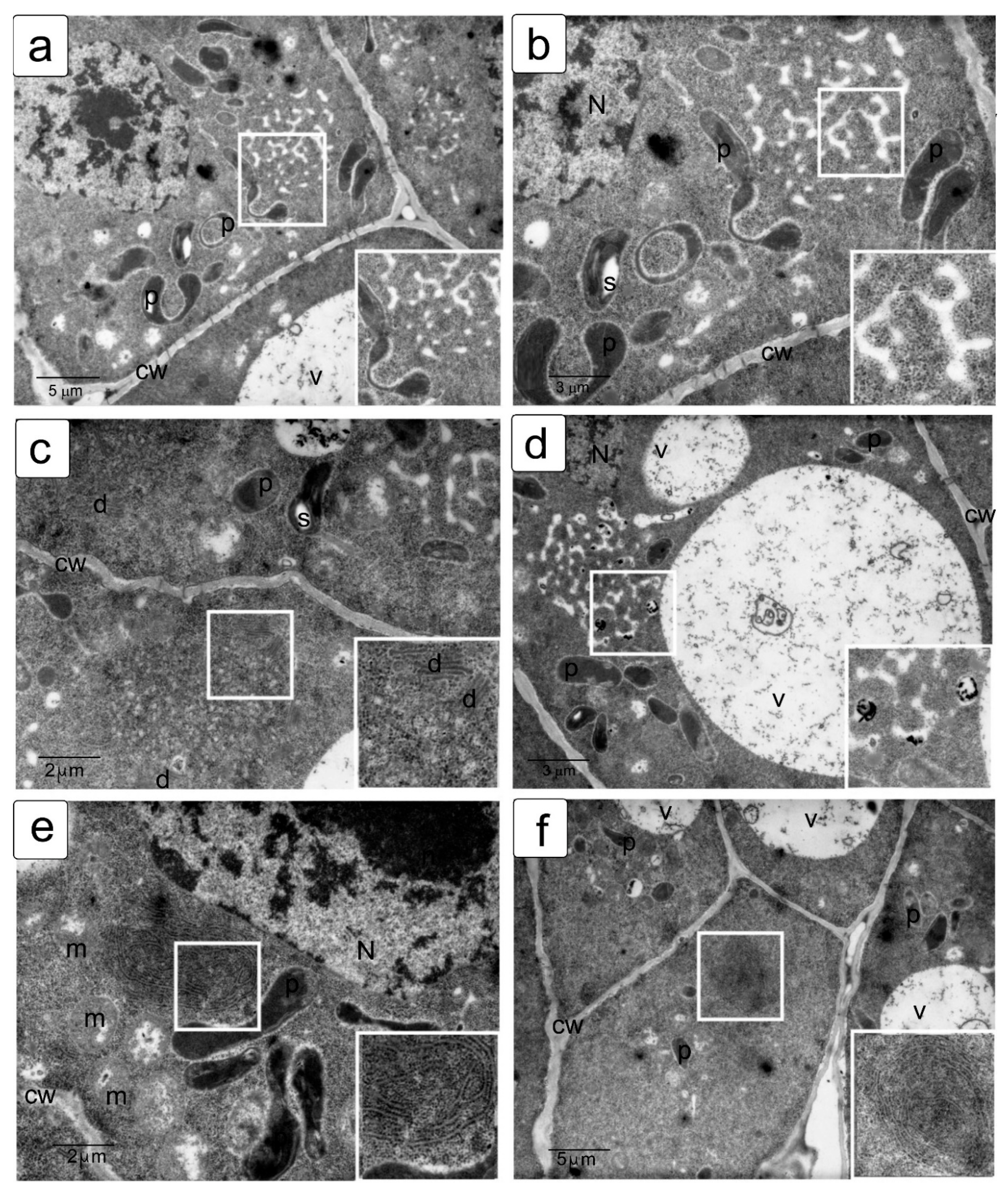
The cytoskeleton is a well-studied cellular system with high dynamics and clear reproducibility of transformations, of which the most well-studied feature is the transformation of the microtubular cytoskeleton in meristematic cells, which are characterized by high division activity. The structure of the cytoskeleton is a clearly reproducible value that ensures both growth and development and, to some extent, the fate of plant cells, since, at the initial stages of differentiation, the direction of division sets the parameters that determine which tissue and which layer a given cell will belong to, and this process is regulated by the displacement of the axes’ symmetry, which is directly related to the rearrangements of the cytoskeleton in mitotic division. As a sensitive model, we used dividing and interphase cells of the root meristem of the main root of wheat seedlings. These cells are a standard object for comparative analysis of the organization of the mitotic spindle corresponding to different phases of mitosis and interphase cytoskeleton, providing growth by stretching under normal conditions and, under damaging abiotic influences, causing disruption of the assembly, polymerization, and depolymerization of microtubule systems and their interaction with each other and with the cell wall [38,39]. In most interphase cells, microtubules are grouped into a system of highly ordered parallel structures located along the cell wall and usually located perpendicular to its long axis; this arrangement is typical for sequentially dividing cells (Figure 3; Figure S1).
The tubulin cytoskeleton, being a highly mobile and extremely sensitive cell structure (Figure S1), can differ significantly from cell to cell. However, some stable states are well described and characteristic of certain phases of the cell cycle and therefore can be easily identified by the state of the chromosomal material and the location of microtubules. The transformation of tubulin and the formation of microtubules is a dynamic process. This is reflected in the fact that homogeneous staining, characteristic of the uniform distribution of tubulin in the interphase, is accompanied by the identification of microtubules associated with the cell wall (Figure S1a,b). They are located parallel to the axis of future fission under control conditions without salt exposure. The action of sodium chloride causes a displacement of the angles of microtubule inclination and a disturbance in the uniformity of tubulin distribution, with the formation of a network around the nucleus (Figure S1d,e). Sodium sulfate, in addition, contributes to the significant shortening of microtubules (Figure S1g,h,j). Obviously, such a violation will also affect the formation of the cytoskeleton in preparation for division and directly affect its passage, which is summarized in Figure 3.
Changes in the slope can be noted in the zone of the meristem initials and can usually suggest a change in the direction of division; however, in the presence of stressful influences, such violations are found everywhere, although they still retain some similarity in the peripheral location. One can observe a large number of deformed bundles devoid of parallel arrangement, the formation of conglomerates, and the formation of perinucleolar clusters and globules (especially in the presence of NaCl); fragmentation and thinning of individual zones can also be noted, against the background of the formation of multidirectional clusters with Na2SO4. These formations are generally characteristic of damage to the interphase cytoskeleton. Earlier, we showed the manifestation of similar phenomena at the ultrastructural level in plants, leading to the formation of clusters of microtubules and their crystal-like clustering. It can be assumed that these consequences are characteristic of cells that have undergone mitotic transformations with disturbances, which are probably accompanied by a violation of the formation of cell walls and the transport of water and nutrients, particularly sugars, accompanying stress and leading to metabolic disturbances characteristic of salt stress. Thus, although cell elongation as a whole is ensured, but significantly slowed down, growth impairment is likely accompanied by increased chaos in the organization of the interphase cytoskeleton structures, which affects the impairments in the direction of interaction and growth, leading to asymmetry in root development (Figure 3; Figure S1). During the transition from interphase (G2 phase) to prophase, processes associated with the onset of mitosis are triggered, and despite the transformations inside the nucleus and the preservation of the nuclear envelope, the structure of the cytoskeleton changes significantly (Figure 3; Figure S2). Disturbance of the symmetry axis and of the thin network of microtubules on the outer surface of the nucleus, which are detected under control conditions (Figure S2a,b), lead to significant changes. These changes are tracked by the violation of the division axis, deformation of the preprophase ring, and disruption of the near-nuclear cytoskeleton. Moreover, the changes are most pronounced under sodium sulfate exposure (Figure S2g,h,j).
For this condition, one can observe the disappearance of most of the cortical microtubules, the movement of part of the tubulin into the perinucleolar zone, and the formation of a special structure—the preprophase band. This formation coincides with the place of formation of the future phragmoplast and sets the location of the future cell wall [40]. Subject to symmetry, the preprophase band is located perpendicular to the long axis, strictly in the central part of the cell, and is a large accumulation of parallel microtubules surrounding the nucleus, resembling a set of rings. For a given violation of symmetry, the angle of the ring can be significantly displaced. The same effect is observed when the root shape is disturbed. In addition, it has been described that salinity, drought, cold, and other stresses have significant effects on these structures [41,42]. The action of NaCl and Na2SO4 is accompanied by a violation of the axis of the band location, a change in the location of perinucleolar microtubules, and the appearance of globular and cluster structures, sometimes accompanied by the formation of a second ring perpendicular to the first ring and/or its fragment. One can expect a significant decrease in the symmetry and order of the cells in the future. Many works pay attention to this stage in connection with stresses affecting the morphology and growth of plants [43,44].
Transformations of the prometaphase cytoskeleton are a process that precedes the creation of a super-efficient mechanism that guarantees the masterly division of daughter chromosomes between daughter cells (Figure 3; Figure S3). There is a loss of the nuclear envelope, and the chromosomes are located in the free central zone of the nucleus. For the effective formation of separation, disassembly of the preprophase band is required, as well as the creation of a specialized system, including poles located in the center of the long axis of the dividing cell and covering the chromosomes of the microtubule stretching to the poles, forming a typical division spindle. Disturbances at this stage indicate a kind of confusion in the transformation of the structures of the cytoskeleton; for example, under the action of NaCl, weak formation of the spindle and a violation of its departure to the poles were observed. Na2SO4 promoted the preservation of sections of the preprophase band and the formation of clusters of microtubules around the nucleus (Figure S3). Similar structures were observed by a number of authors as disturbances in cytoskeleton perturbations under a number of other influences [45,46].
The metaphase is of significant interest for the identification of cytoskeleton damage (Figure 3; Figure S4) and may indicate significant disturbances in the division processes, as it is known that the future process of microtubule divergence is provided by a special arrangement of polar zones, a kind of pole formed by kinetochoric beams of microtubules, which are significantly expanded in comparison with a prometaphase spindle; this structure allows for the clear and error-free separation of the genetic material of future daughter cells [47]. Violation of the structural organization can lead to an essential violation at the stage of chromosome distribution, as well as leading to a violation of their divergence and the formation of non-identical aneuploid cells or the formation of deformed nuclei with a violation of the formation of cell walls. The action of salts prevents the formation of a metaphase plate, which is probably caused by a violation of the processes that provide the polymerization/depolymerization of microtubules, which ensure the movement of chromosomes to the division axis. Disorganization of the cytoskeleton is accompanied by a homogeneous distribution of tubulin throughout the cell, and the formation of uncharacteristic bundles and clusters chaotically distributed throughout the cell volume. At the same time, in some cells, the structure of the metaphase spindle is still formed and disorganization is more pronounced under the action of Na2SO4. It can be assumed that this explains many disturbances in morphology under edaphic stresses [48,49]. However, in this case, it is also possible to associate the disturbance with chaotization rather than with a given and controlled asymmetry, which is why such damage can hardly be completely leveled out in further growth.
In anaphase of the cell cycle, the process of chromosome separation in future daughter cells is completed (Figure 3; Figure S5). Chromosomes retain their distinct structure and a high degree of condensation; no nucleus is formed. The cytoskeleton is a shortened bundle of microtubules located perpendicular to the division axis between the accumulations of chromosomes passing to the formation of nuclei. In anaphase, the most noticeable are the critically dangerous consequences of disruption of the earlier stages of cytoskeleton reformation, in particular, leading to the formation of abnormal nuclei and violations of the distribution of chromosomes in the metaphase, the so-called chromosomal bridges characteristic of many effectors damaging division [50,51] (for example, Figure S5g–i) under the influence of Na2SO4. It is also possible to observe disturbances in the distribution of tubulin not associated with microtubules in the cytoplasm and changes in the location and length of the bundles between the dispersed chromosomes, with the formation of a less uniform and ordered cluster structure, which is probably an obstacle to ensuring the normal flow of cytoskeleton transformation processes in anaphase, as in other phases, there is an increase in the density of the cytoplasm and a decrease in mobility due to the presence of osmotically active substances, such as sugars, proline, and polyamines, the amount of which, according to many authors, is significantly increased under the action of salts and osmotics [52,53].
Telophase is the final stage of mitosis, accompanied by two important processes that determine the shape of cells, the interaction between cells, and the assembly of the nuclear membrane around the nuclei of daughter cells. Disorders that accompanied the restructuring of the mitotic cytoskeleton at this stage affect the formation of the phragmoplast, the shape and structures of the cell wall, and the distribution of plasmodesmata, and at this stage, the consequences of the violation of the distribution of chromosomes, affecting the shape of the nuclei and their structure, become apparent. The observed disturbances are accompanied by phenomena probably explained by the delay in the assembly/disassembly of the cytoskeleton structures, causing the preservation of elements characteristic of other phases of the transformation of tubulin-containing structures [54]. NaCl causes the preservation of the distribution of fragments of globules and bundles throughout the volume of the cytoplasm, fragments of asymmetric structures in the phragmoplast zone (Figure 3; Figure S6d–f). Na2SO4 is characterized by the fragmented preservation of bundle clusters between daughter nuclei and in individual cells by the globularly diffuse distribution of tubulin with fragments randomly located along the periphery of the cell and in the phragmoplast zone by short microtubules (Figure S6i–k). Cytoskeleton structure reorganization accompanies all phases of mitosis, and although individual disorders can be identified in normal conditions, they are, nevertheless, single. The disturbances presented here are the most typical and indicate significant disorders of division processes caused by the action of NaCl and Na2SO4 (Figures S1–S6) and are characteristic of osmotically harmful stress consequences [52].
Significant disturbances in the transformations of the cytoskeleton under the action of NaCl and Na2SO4 are associated with the formation of specific structures accompanying and providing the process of mitotic division and distribution of chromosomes between daughter nuclei. These disorders are laid down and persist at different stages of mitosis and are associated with a violation of the correct, close to symmetric distribution and transformation. Disturbances are associated with a violation of the characteristic pattern of parallelism of cytoskeleton fragments: in interphase and mitosis, angles and places of divergence; in prometaphase and metaphase, the distribution of tubulin not localized in microtubules; at different stages and under various influences, the displacement and duplication or clustering of elements that are characterized by parallelism or a high degree of organization (mirror symmetry), especially during the formation of a preprophase band; in prophase, a spindle of division in prometaphase and metaphase, and disturbances in the arrangement and fragmentation of the organization of the arrangement of microtubules during the transition to the initiation and formation of a phragmoplast and cell walls in anaphase and telophase (Figure 3).
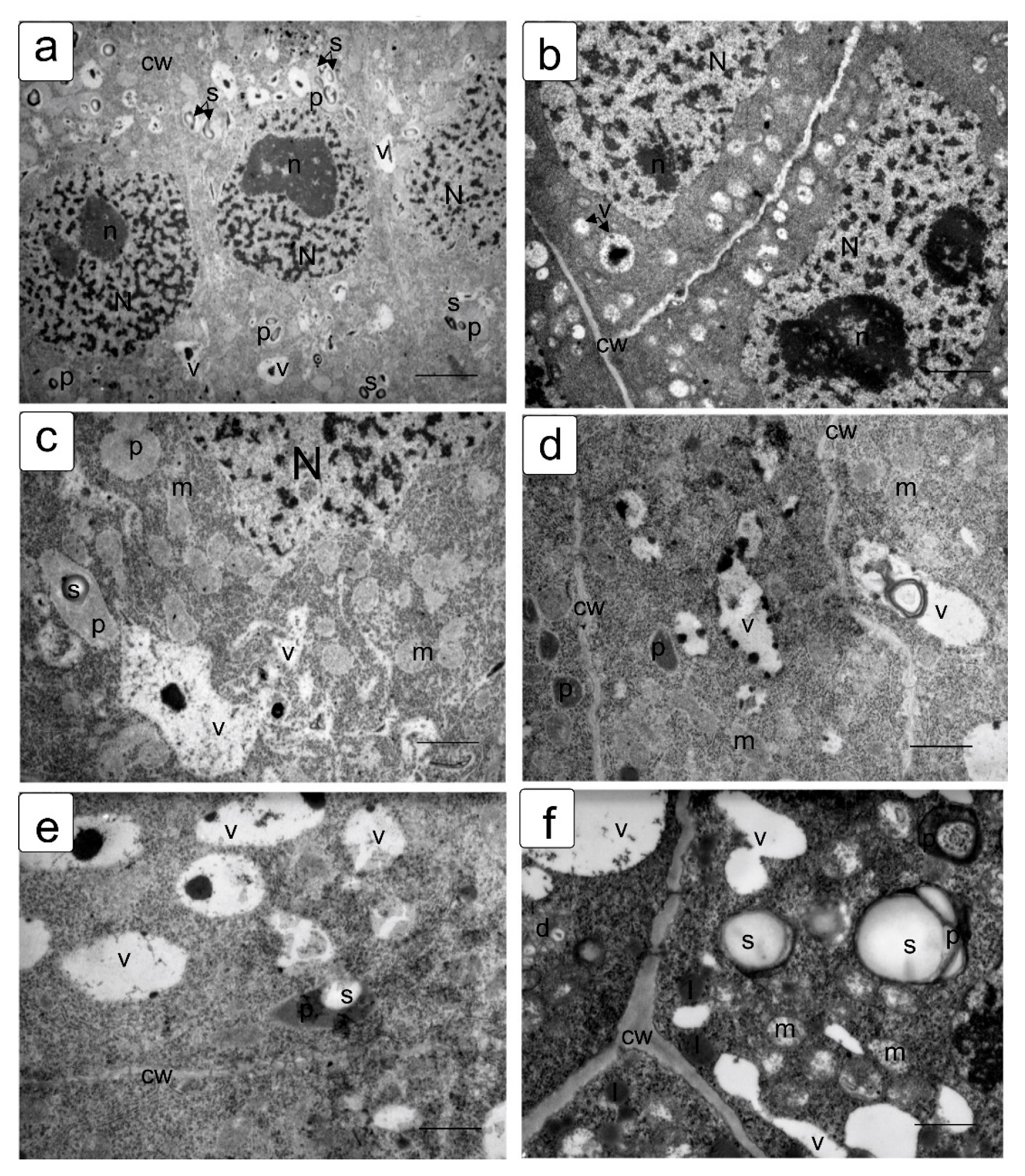
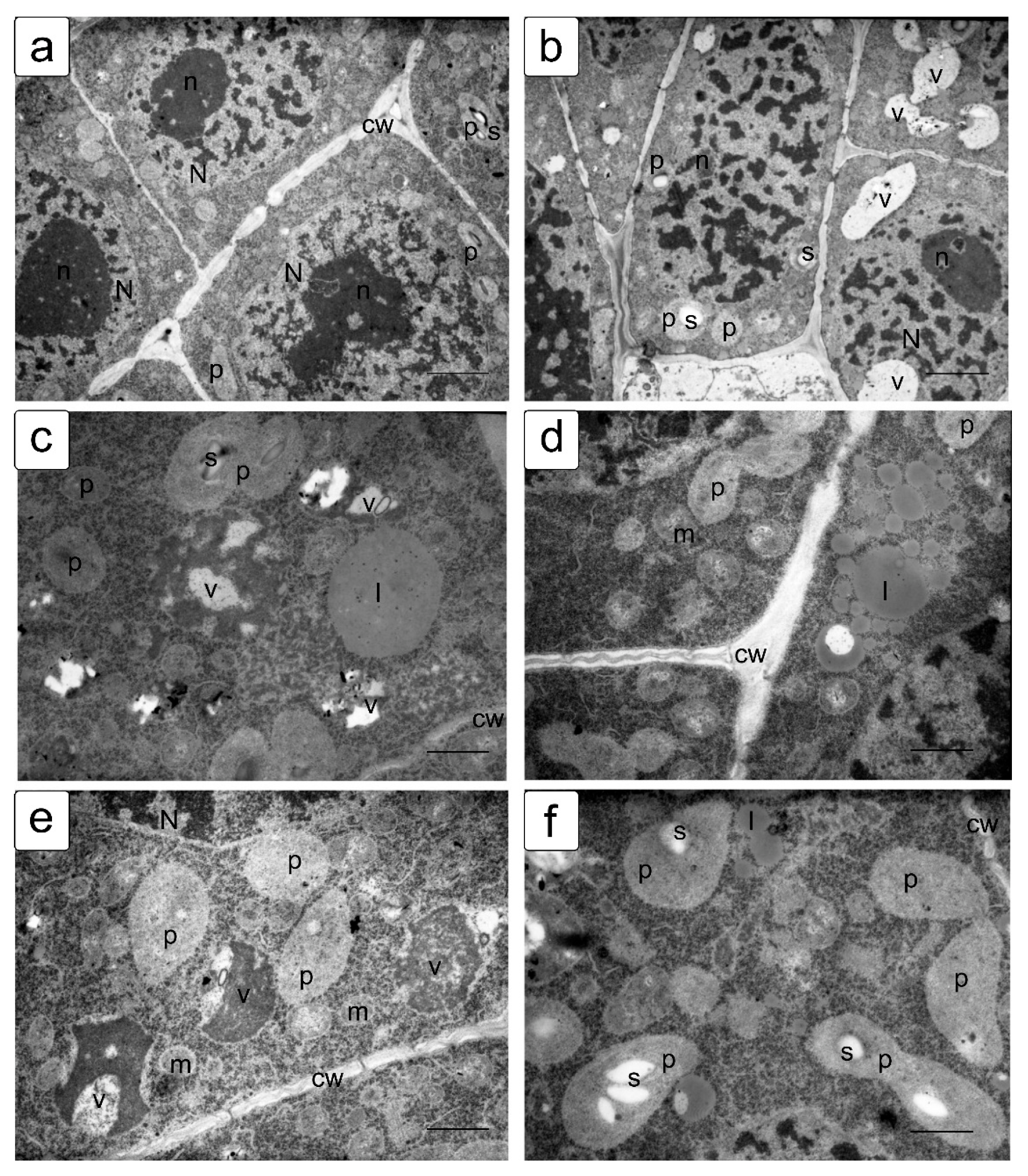
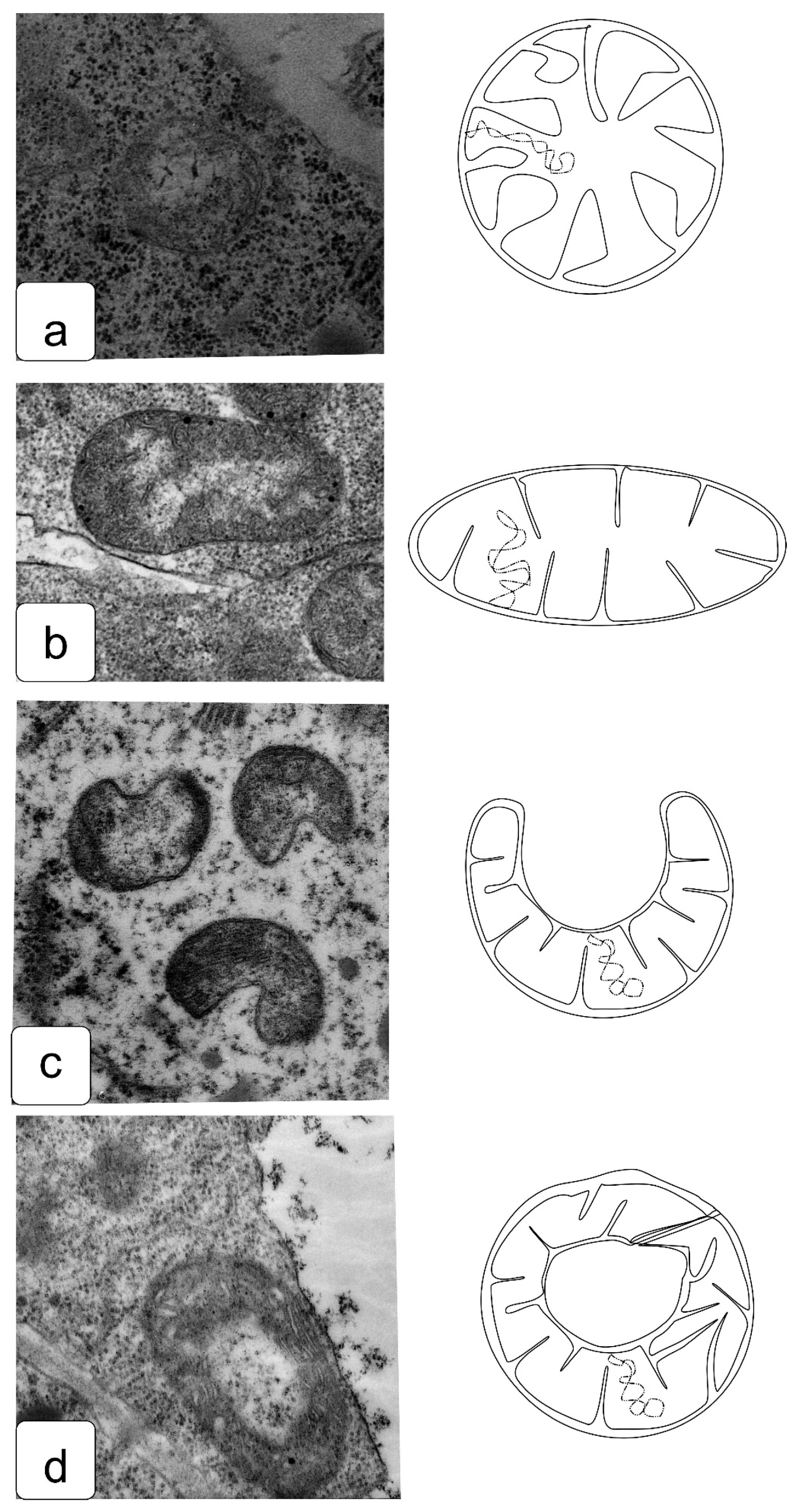
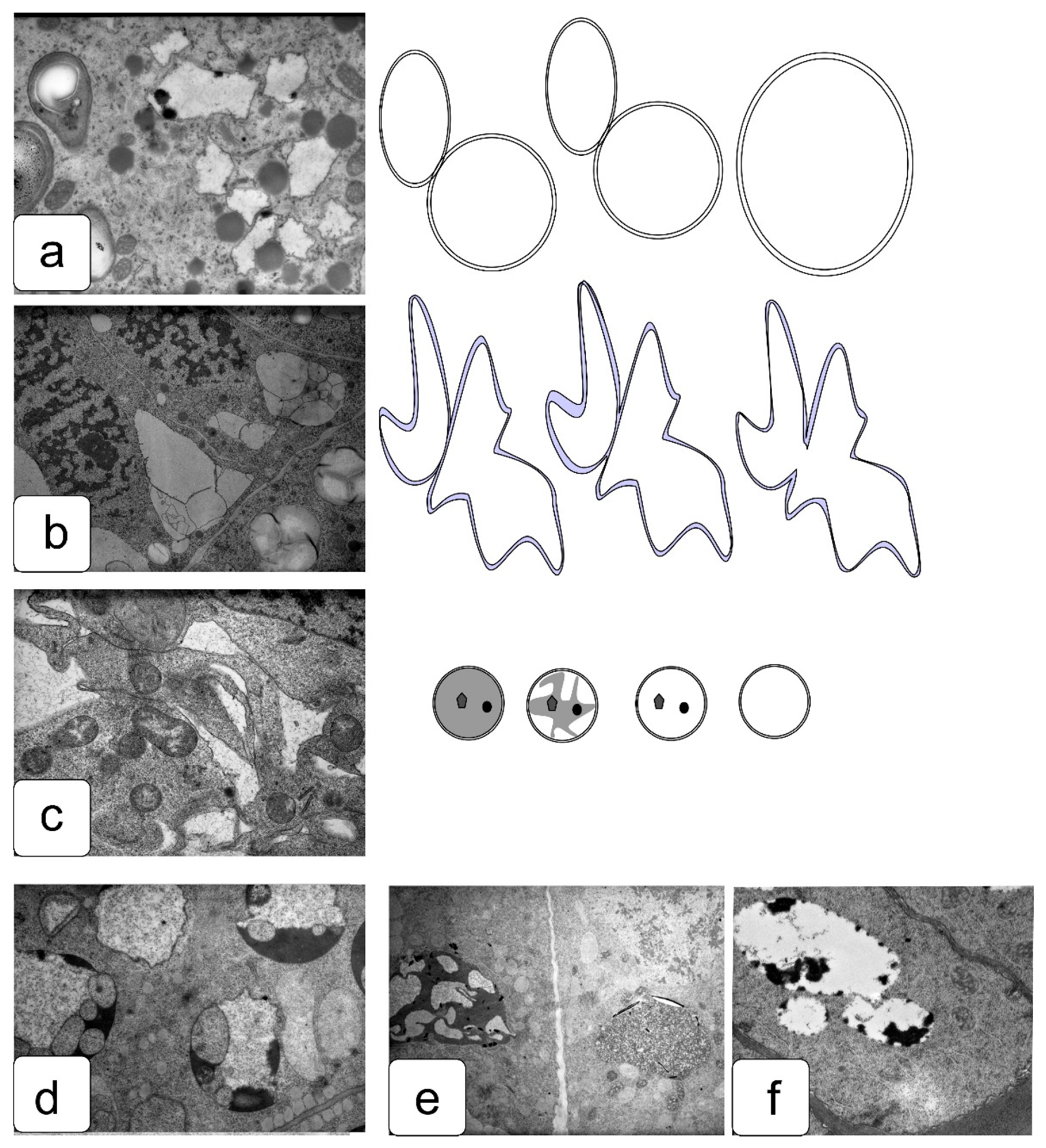
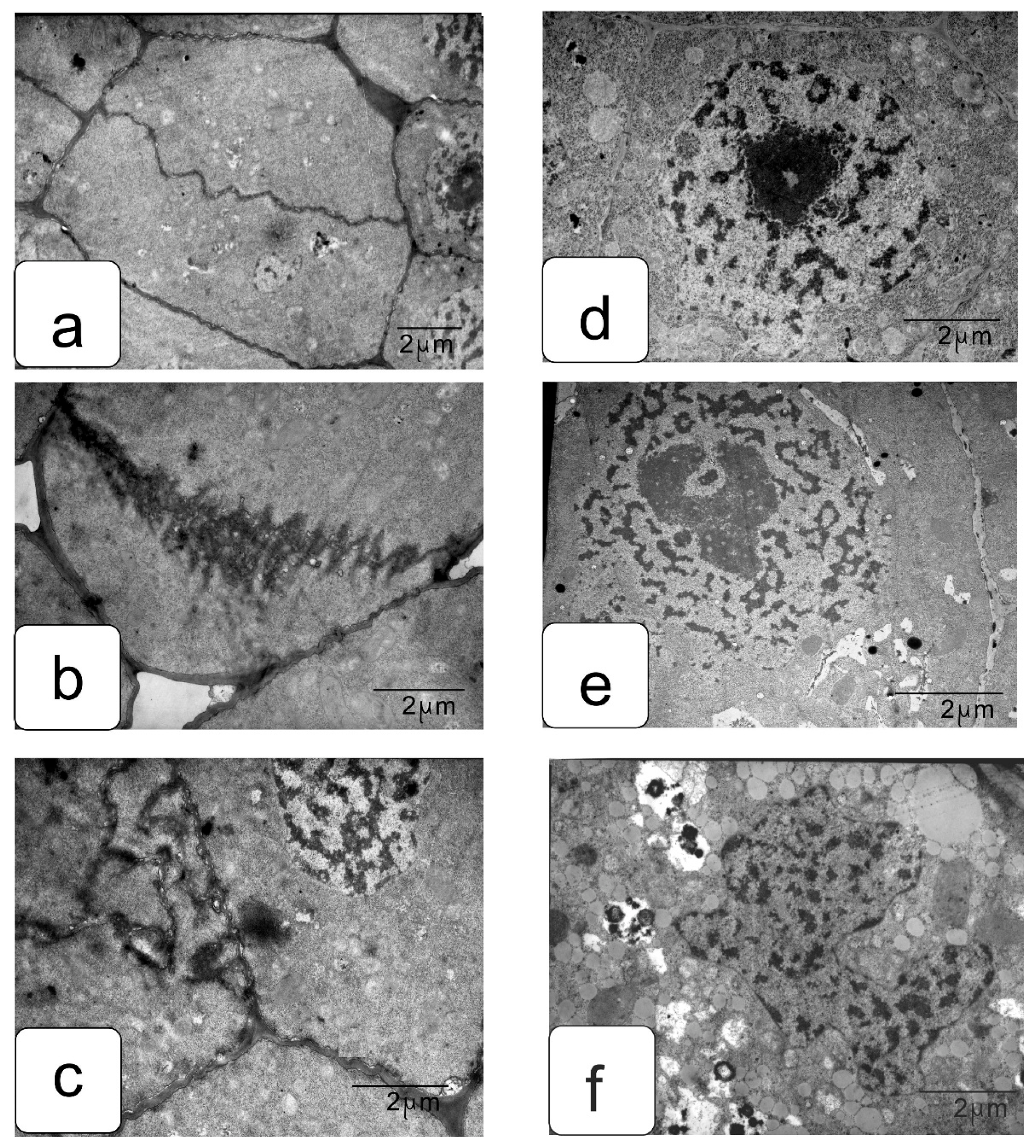
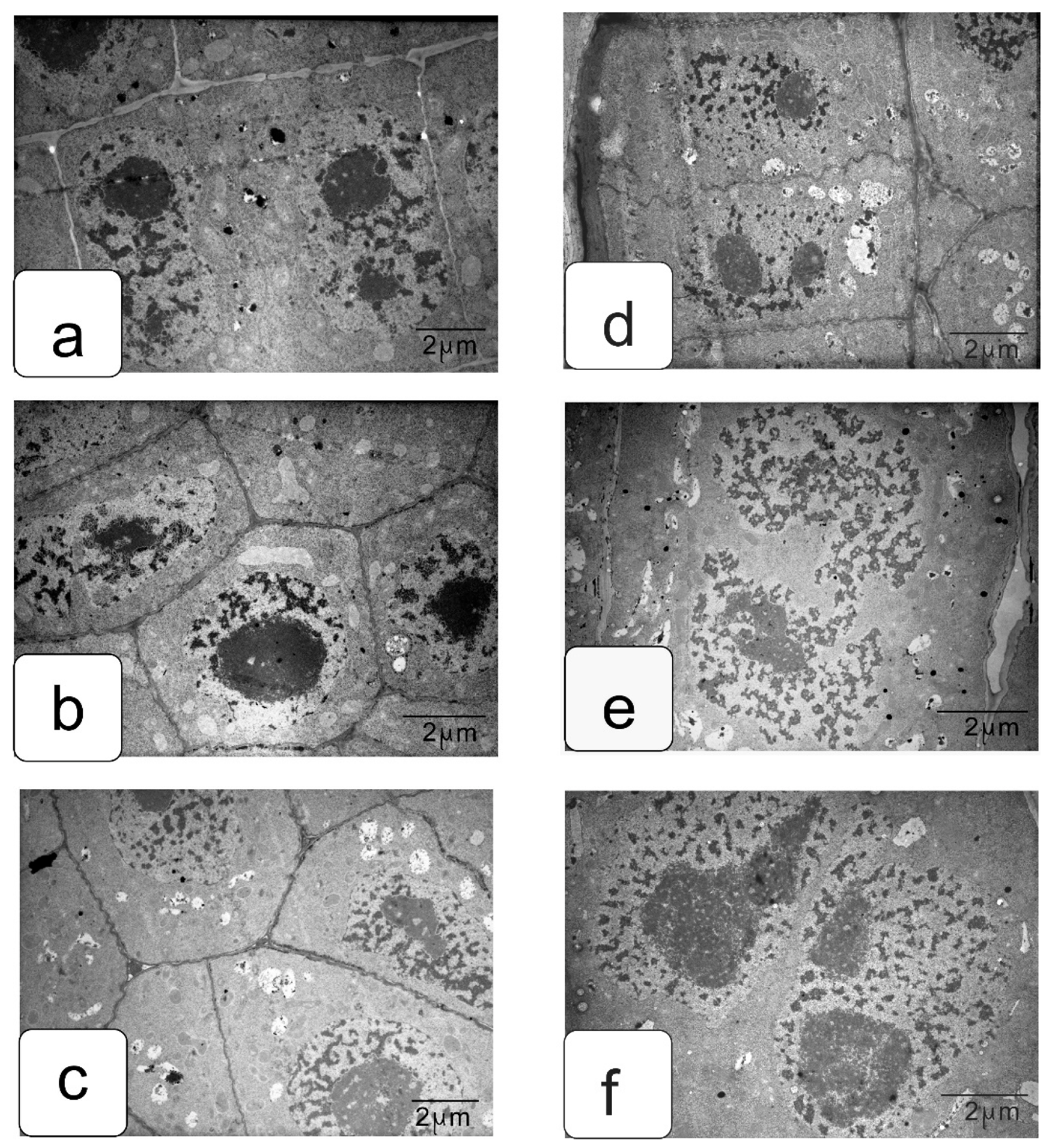
In actively proliferating cells, the root and shoot meristems are usually characterized by a large nucleus with a central location. The nuclei have a large nucleolus, and a high density of ribosomes can be observed in the cytoplasm, which corresponds to the efficient process of the production, assembly, and production of ribosomes characteristic of meristematic tissues (Figure 4a,b). Plastids and mitochondria are usually evenly distributed in the cytoplasm. The production of small vacuoles and their fusion does not lead to the formation of large ones; inside them, dark inclusions of presumably protein nature are often noticeable (Figure 4c–f). Individual cells contain plastids with large starch grains (Figure 4f).
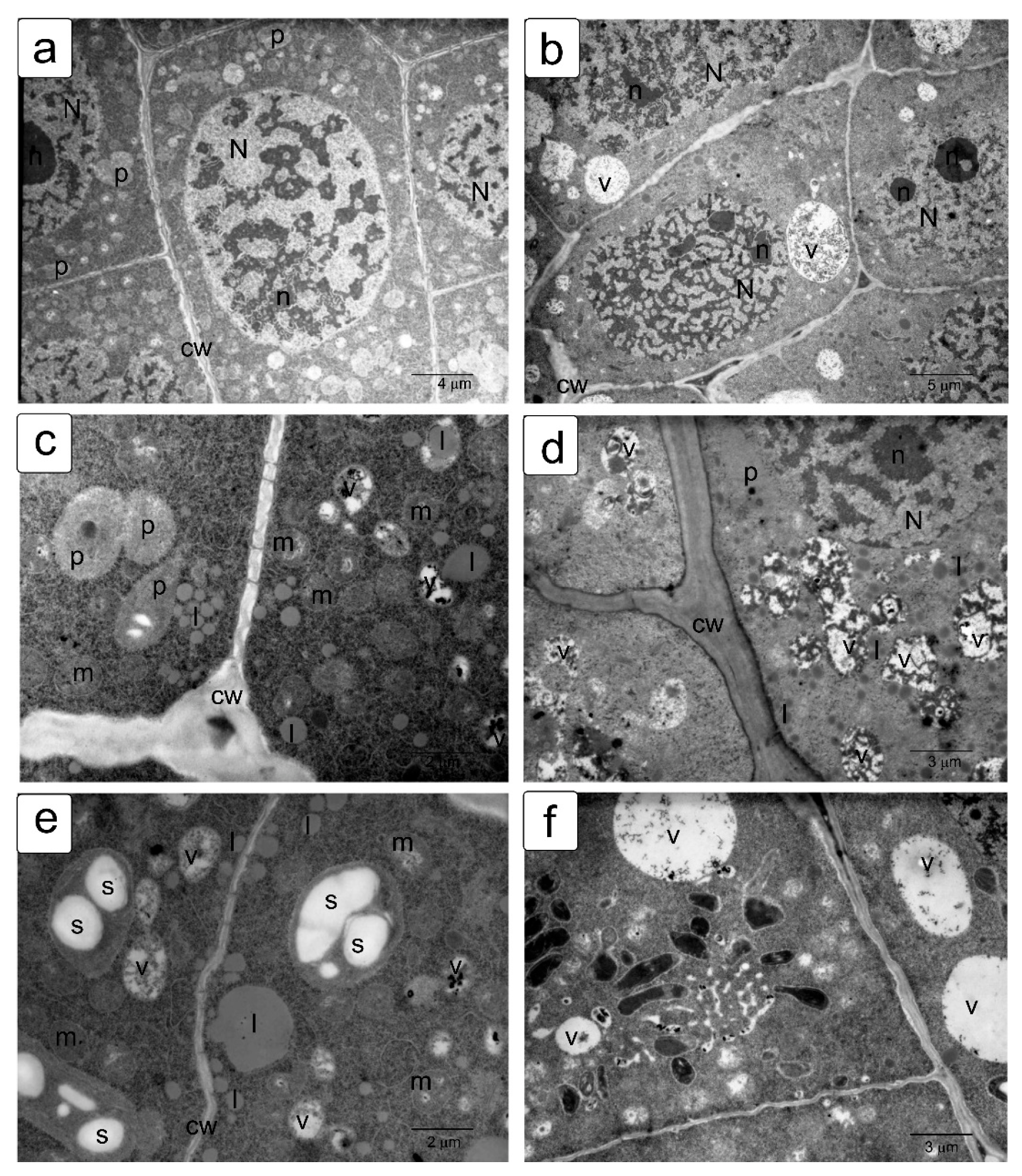
When germinating in the presence of NaCl, the cells of the apex of the root and shoot have a characteristic structure with a large nucleus and nucleolus, and dense cytoplasm saturated with ribosomes. However, lumps of condensed chromatin can be seen inside the nucleolus; the size of large lumps of condensed chromatin in the cytoplasm also increases, which indicates the inhibition of a number of synthetic processes, including the activity of the nucleolus (Figure 5a,b). There is a noticeable retention of storage substances such as lipids and protein inclusions in the vacuoles (Figure 5c,d). One can also note a decrease in density and an increase in the volume of plastids—they become more rounded, and one can observe their active division, for example, in shoot cells (Figure 5d,f).
Na2SO4 causes a marked increase in separation between condensed and decondensed chromatin (Figure 6a,b), more pronounced in shoot cells. Processes of slowing down lipid utilization can be observed, especially in root cells (Figure 5c,e), and proteins in individual shoot cells (Figure 5d). In the initial cells of the parenchyma of the shoot, vacuoles become more rounded and contain special structures of the endoplasmic reticulum located only in one zone of the cell (Figure 5f).
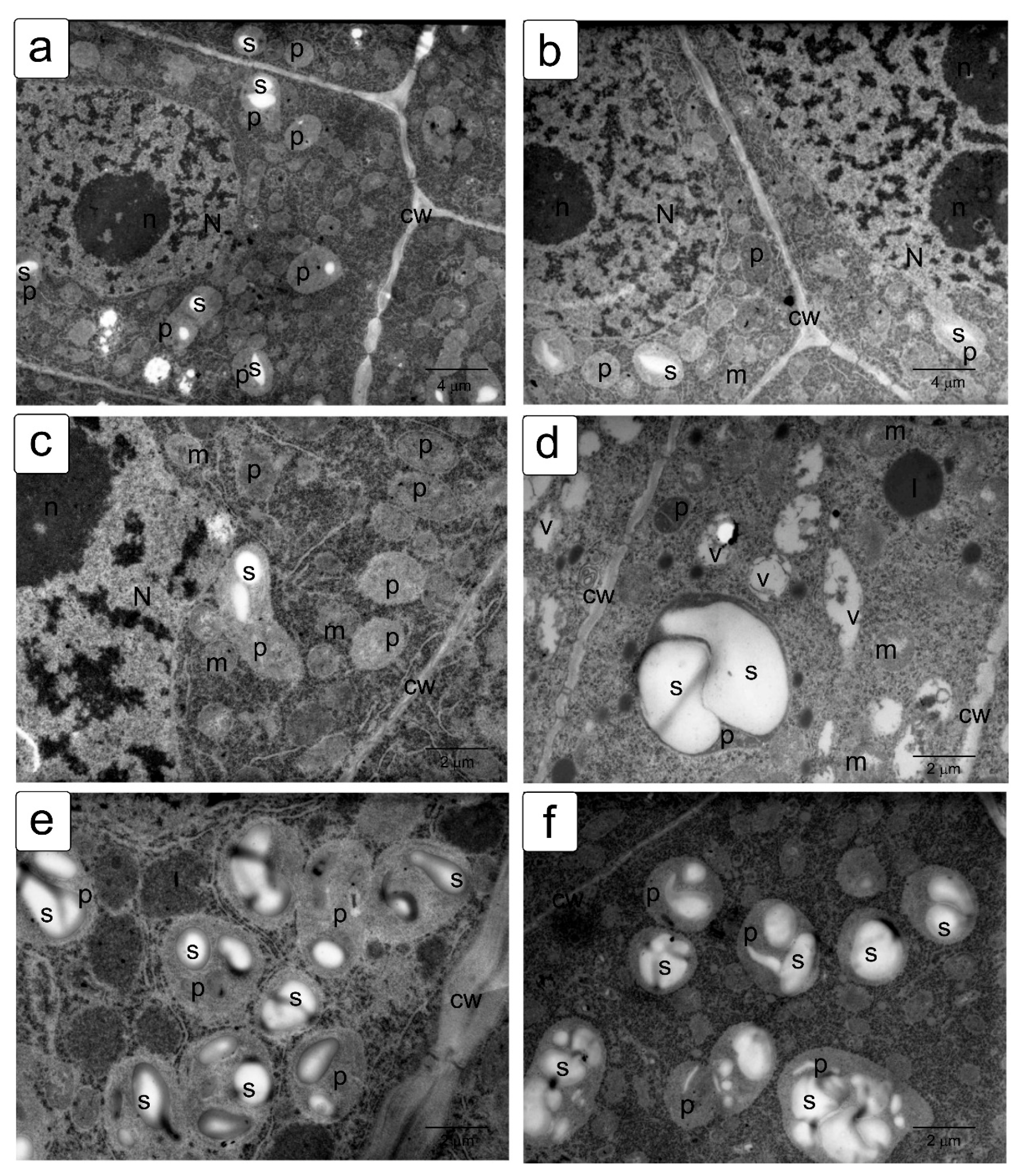
The effects of delayed utilization of storage materials are well documented in a number of crops, such as wheat, flax, cashew, and others [55,56,57]. In the cells of juvenile seedlings, a delay in the utilization of starch, protein reserves, and a violation of lipid metabolism [36,57,58,59] were previously noted, accompanying a significant decrease in growth and development. This can be explained by blocking of the functioning of enzymes, impaired absorption capacity, and the ability to transport and synthesize metabolites [60,61].
To identify osmotic effects, we also analyzed the effect of mannitol at the appropriate iso-osmotic concentration (Figure 7). It can be assumed that the osmotic does not give rise to the toxic or oxidative stress components of salinity stress. In general, the effects of delayed lipid and protein utilization were observed (Figure 7c–e). It was also possible to note a hypertrophied large amount of starch in plastid inclusions (Figure 7e,f), which was not observed under other influences.
In addition, expansion growth processes are accompanied by characteristic movements of cell organelles. In meristematic cells, the nucleus is located in the center of the cell, and the plastids, mitochondria, and endoplasmic reticulum are located in the cytoplasm between the nuclear envelope and plasma membrane.
However, their arrangement can be considered uniform only conditionally, which is primarily due to the fact that the number of organelles is not the same, and even with a relatively equal osmotic pressure of the cytoplasm, the osmotic pressure in each individual intracellular compartment is relatively higher, including the nucleus, mitochondria, and plastids.
This is easy to understand, due to their preservation of the “ideal” rounded shape. With growth or osmotic abnormalities, the processes presented in Figure S7 can be significantly impaired. Normally, an increase in the volume of the vacuole leads to a displacement of the nucleus and organelles, together with the surrounding cytoplasm, to the edge—the periphery of the cell volume. Deformations involving the impaired processes of fusion, division, and locomotion are associated with uneven osmotic pressure and improper cytoskeleton rearrangements under stress (Figure 4, Figure 5, Figure 6, Figure 7 and Figure 8).
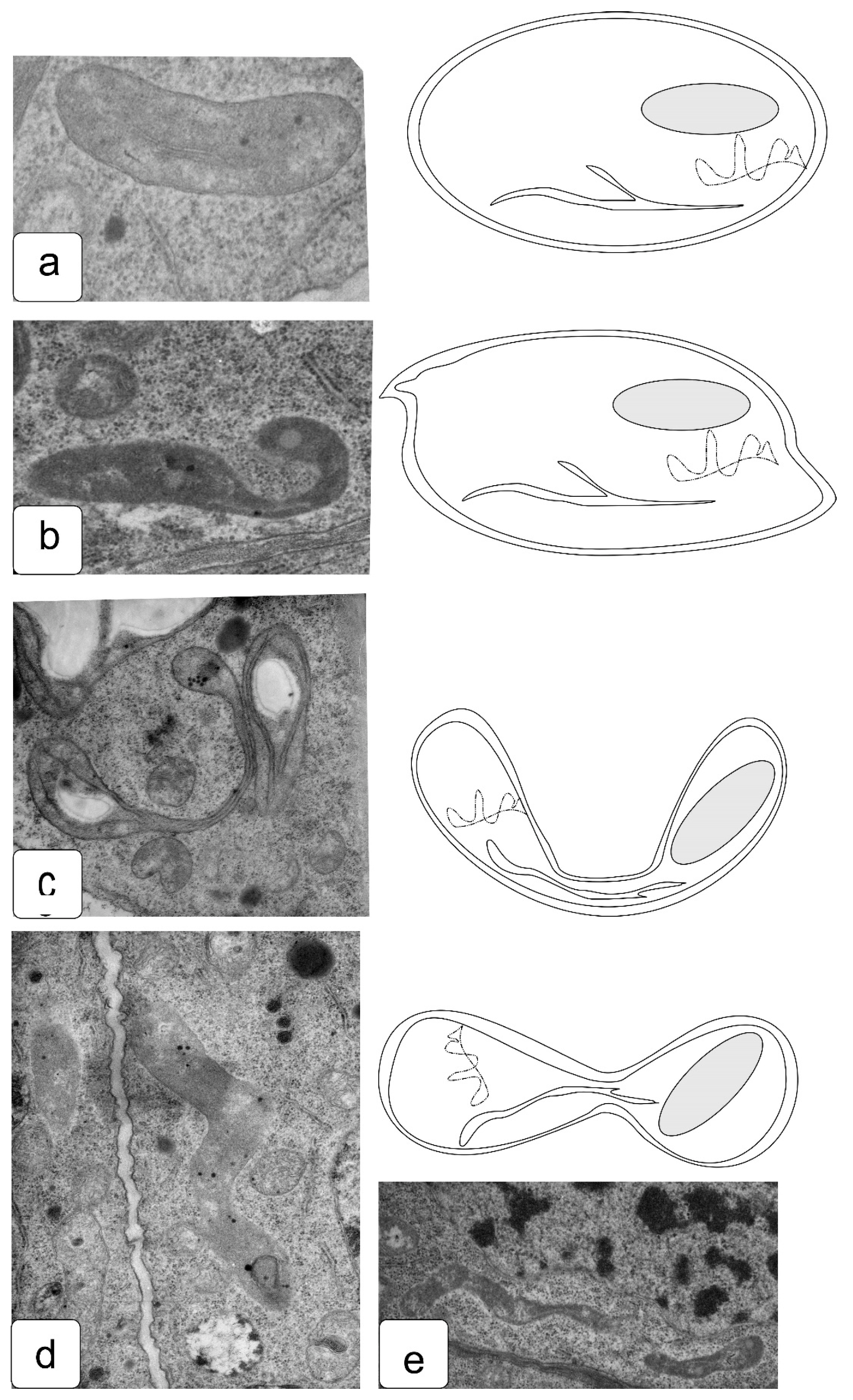
Features of changes in mitochondrial morphology in apex cells under salt-related stress often include morphology changes associated with acceleration of development or presumably similar to damage characteristic of oxidative stress [62] (Figure 9). Being the most important plant organelle in terms of energy, mitochondria can change significantly depending on the environmental conditions. When cells move from the meristematic zone, the most frequent change is a change in shape from round to oval (Figure 9b).
In cells that develop under salt action, the change in shape occurs more rapidly. Thus, a frequent case is a change in the morphology of cristae and their density [63]. One may also observe a change in shape with the formation of invaginations or even cupped mitochondria, which are torus-shaped in a longitudinal section (Figure 9c,d).
Figure 9.
Ultrastructure of stress-induced changes in mitochondria of wheat root cells: usual configuration (a), oval shape (b), transverse cup-shaped (c) and longitudinal cup-shaped mitochondria (d). The effects of changes in the shape and ultrastructure of plastids and mitochondria reveal the high sensitivity of these organelles to the action of stress, and often indicate disturbances in the process of division, which lead to the appearance of large organelles of uncharacteristic size [64].
Figure 9.
Ultrastructure of stress-induced changes in mitochondria of wheat root cells: usual configuration (a), oval shape (b), transverse cup-shaped (c) and longitudinal cup-shaped mitochondria (d). The effects of changes in the shape and ultrastructure of plastids and mitochondria reveal the high sensitivity of these organelles to the action of stress, and often indicate disturbances in the process of division, which lead to the appearance of large organelles of uncharacteristic size [64].

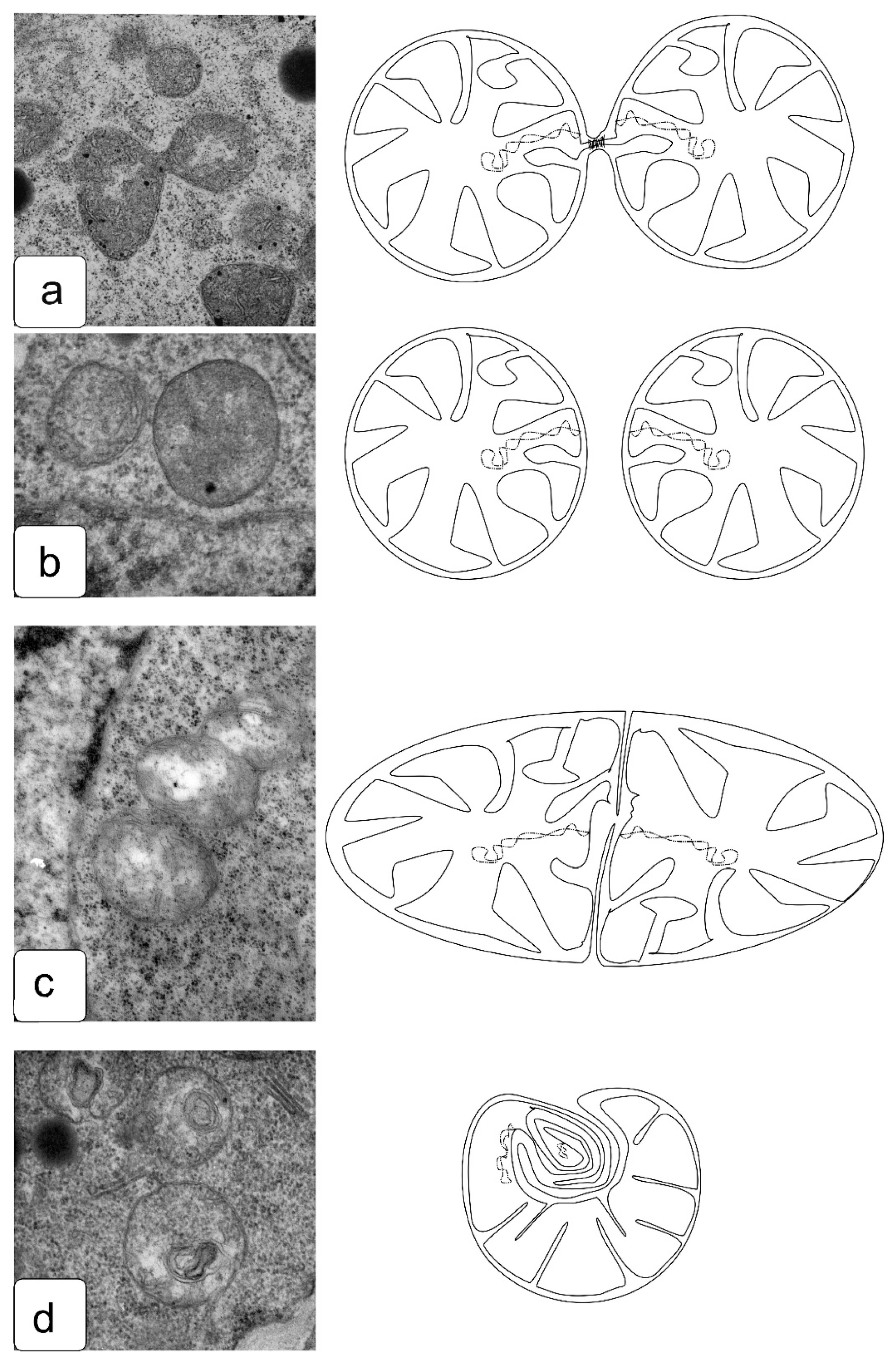
The development of plastids, the accumulation of starch, and the formation of membrane structures in them in non-photosynthetic tissues are currently insufficiently studied. Changes in their shape are a frequent occurrence accompanying various stress effects, including edaphic stresses. One can note the appearance of strong invaginations, the identification of a large number of plastids of an irregular or cupped shape (Figure 10c), as well as elongated plastids (Figure 10d,e).
The processes of organelle division are as necessary as the processes of nuclear division, since, if this process is disturbed or blocked, the cells newly formed after division must receive their portion of organelles that will be able to provide energy and metabolic processes. Meanwhile, the division of organelles and their distribution between daughter cells are often disturbed under the influence of stress factors [65]. We observed typical patterns of cell division and some features of division in the presence of salts, which indicate a violation of the process. Here, we demonstrate that, in the most common process of mitochondrial division with the formation of a constriction, the cells undergo a slight change in turgor and do not maintain an ideal round shape (Figure 11a). In some cases, such a violation is accompanied by differences in the ultrastructure of daughter mitochondria (Figure 11c), which are reflected in changes in the density of the matrix and in the location of the cristae. Another less common type of division is preserved, in which the constriction is not formed, but one can observe the budding of one mitochondrion from another while maintaining a larger interaction area and a common outer membrane (Figure 11c), which is rarely found in the mitochondria of apex cells under normal conditions. In a number of cells formed in the presence of salts, it is possible to observe a violation of division with the formation of micromitochondria within the maternal mitochondria (Figure 11d). This effect has been described for some animal cells with significant disturbances in energy processes associated with growth and development under damaging influences [66].
The division of plastids is also an important process that ensures the development of plant cells and the body as a whole [67]. The toxic effects of salts can affect the separation of plastids after division (Figure 12). The type of plastid division with an increase in osmotic pressure can also change significantly, since, in some initial cells of various tissues and in the proliferation zone, non-diverging conglomerates of plastids can form, inside which one can observe the divergence of plastids from a single center (Figure 12c,d). This phenomenon is not typical and has not been previously described in the literature, since it was considered the simple accumulation of plastids. However, if we consider the types of divisions of cyanobacteria, then, in some works a similar structure is described [68,69]. The alga Coleochaete orbicularis shows alternations of radial and circumferential cell divisions, and this is apparently achieved through centrosomes that migrate to the respective cellular poles [70,71].
Thus, it is obvious that the action of salts can cause a change in the type of symmetry from a stable equilibrium system with a parallel, axial, and rotary model of division and growth to a more retarded (in development) and uneven model. In this case, the processes of strengthening the conditional order and alignment of the length of the lateral adventitious roots under the influence of NaCl can be noted (Figure 1 and Figure 2). On the other hand, the processes of “conditional chaos” are clearly intensifying—for example, manifested in the curvature of the growth directions of the main and lateral roots under the influence of NaCl and Na2SO4, which looks like a disturbance of gravitropism (Figure 1 and Figure 2).
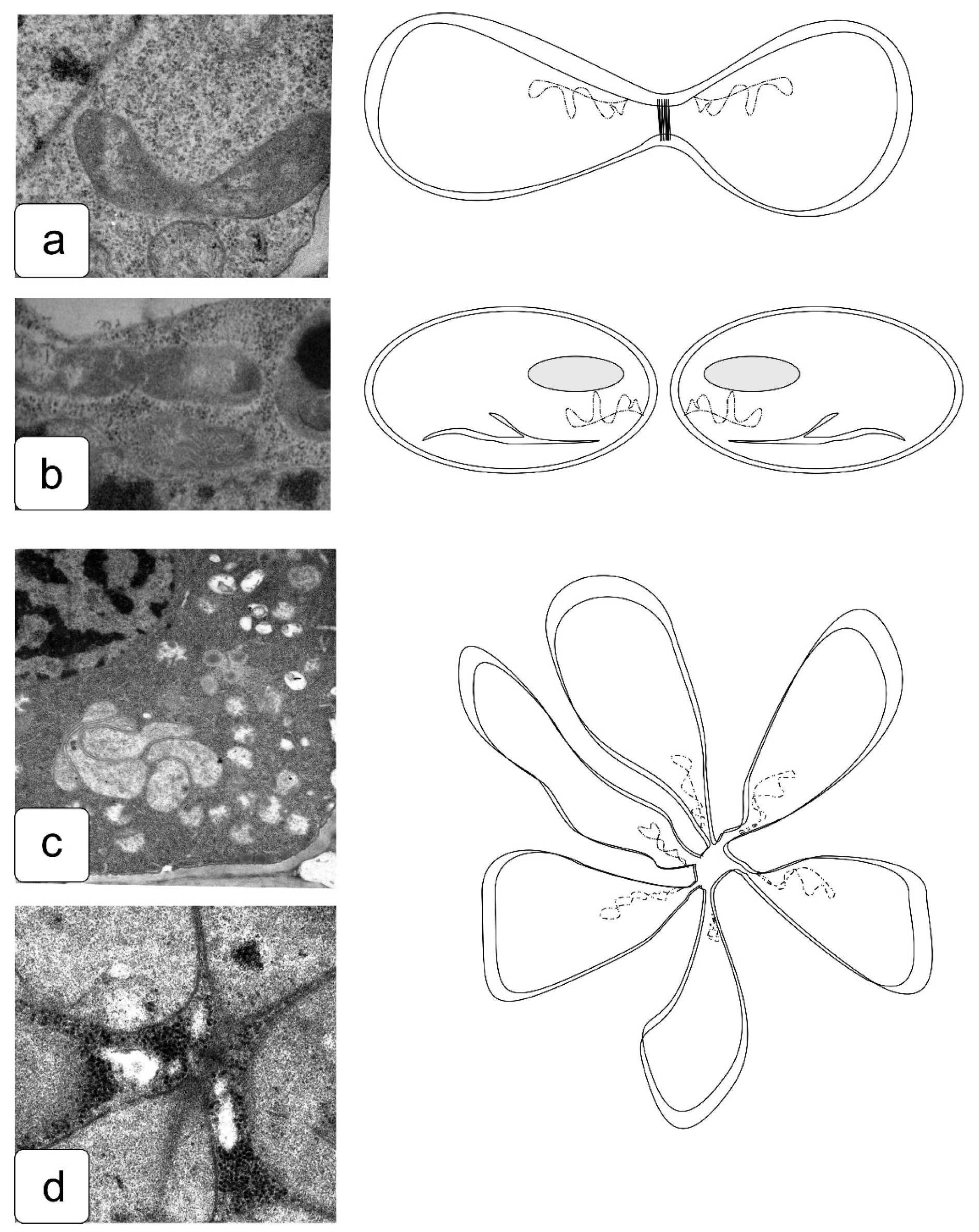
The growth of plants during the transition of developing tissues from the division zone is accompanied by the development of new vacuolar membranes and the accumulation of solutions with various functions in them, which, in the case of salinity, will contain not only water but also often accumulate excess ions to eliminate their toxic effects; as a result of this, the storage and lytic functions of the vacuolar compartment can be disrupted (Figure 13). In particular, when the balanced system of membrane transporters was disturbed or malfunctioned, vacuoles with uneven edges were observed (Figure 13a,c,d). Vacuoles provide the regulated exchange of fluids and ions between the cytoplasm and the intravacuolar solution. The osmotic pressure can change within a vacuole, which is normally higher in vacuoles than in the cytoplasm, due to which they have an “ideally” round or nearly round shape (Figure 13f).
With an increase in cell size, this pressure provides a displacement of the cytoplasm and organelles to the periphery of the cell and their location along the cell wall, in contrast to the central location of the nucleus typical for meristematic tissues and the relatively uniform arrangement of organelles along the periphery (Figure S7). If the fusion is disturbed, part of the membranes can be non-functional; the fusion of the membranes into a single structure can be disturbed, as in the case of the action of Na2SO4 (Figure 13b). If the fusion of membranes is not disturbed by the action of NaCl, but the membrane permeability is disturbed or the pressure of the external solution is insufficient, cells with a highly branched vacuole can be observed, which remains without the proper pressure of intravacuolar solute and probably prevents the cell from expansion (Figure 13c). The utilization of storage substances in this case can be severely impaired (Figure 13d–f). Vacuoles formed and developing in cells at early stages of development are functionally similar to the vacuoles of storage organs and may contain different protein fractions, phenolic compounds, and crystalline inclusions [72,73]. Disruption of enzymatic systems and the activity of lysing systems leads to the partial retention of a variety of intravacuolar inclusions, characteristic of salt and osmotic stress [58]. Such processes can be caused by changes in the system of neutralization of ions and biogenesis of aquaporins caused by downregulation of the expression of genes associated with these processes or by other factors [74,75].
In the cells of the initials of the root and shoot, the vacuolar system is not developed. Moreover, the process of cell wall formation is influenced by the above-described disturbances in the cytoskeleton structure at different stages of mitosis. They cause a change in the normal process of cell wall formation at the site of the phragmoplast after telophase (Figure 14a). Significant disturbances occurring in individual cells under the action of NaCl (Figure 14b) and Na2SO4 (Figure 14c) cause significant disorganization and randomness in the formation of the cell wall as a result of disturbances in the formation of the phragmoplast, changing the morphology of a part of the cells. Such disturbances are well described in the literature and are accompanied by patterns of cytoskeleton disturbance, growth damage, and disorganization of the root and shoot shape—for example, under the action of herbicides, toxicants, and edaphic influences [76,77,78,79]—accompanying significant disturbances of symmetry characteristic of growth.
Division pathologies also affect the shape of the nucleus and nuclear envelope. Normally, the nucleus is almost perfectly round (Figure 15d). The appearance of local invagination (Figure 15e) or a complete change in the shape of the core (Figure 15f) was observed under the influence of NaCl (Figure 15e) and Na2SO4 (Figure 15f). The shape of the nucleus is an important feature and is associated with the fact that its internal pressure is maintained by the cell to be higher than that of the cytoplasm due to a complex double membrane with selective throughput [80]. Therefore, disruption of the shape is a qualitative characteristic of damage, at least in the regulation of the osmotic potential of cell compartments.
Thus, if relative asymmetry is a variant of the norm, then a clear violation probably indicates significant damage that will not remain without consequences.
A peculiar balance between phenomena that can be attributed to the manifestations of symmetry (with a characteristic effect of the repetition of phases, stages, states of individual elements or cell structures) and asymmetry (in the manifestation of which the coincidence, analogies, and repetition of phenomena become unclear, unobvious, or cannot be identified at all) determines the development of multicellular organisms and, in particular, plants. The clearest correlation of phenomena associated with the new formation or uneven development of a leaf, root, shoot, or flower is currently associated with the peculiarities of the coordinated auxin-driven reorganization of the cytoskeleton. The reorganization controls asymmetric cell expansion and division during embryonic and postembryonic organogenesis [81], changes in root or shoot growth, and the establishment of new organs due to changes in cell polarity [82]. For a long time, the connection between the impact and changes in the amount of auxins and, in particular, IAA was not clear. However, when it was found that the amount of IAA significantly increases in the places of bends, and a connection with gravitropism was established [83], it became clear that this process is related to the transport of sugars and deposition of starch grains by analogy with the “leading” role of columella in the direction of root growth [84].
In this study, we used the early stage of plant development, when the bulk of metabolites are not yet produced in cells, but are supplied from seed reserves, in general, providing excessive energy and intensive primary growth. An increase in osmotic pressure during germination inhibits enzymes, which decompose the storage reserves and transport the products of utilization through the scutellum to the developing tissues of the seedling, which limits the growth and development of plantlets [85]. This process is well studied on both dicotyledonous and monocotyledonous plants and, despite the fact that the reserves are concentrated in different types of cells, in general, it affects the disruption of the utilization of vacuolar protein bodies, starch inclusions of plastids, and lipid droplets [36,58,86,87,88,89]. The role of plant hormones in ensuring the growth of new organs is obvious and correlates with the supply of starch to the sites of growth from the sites of its utilization. Thus, the effect in starch mobilization and root elongation is in some part regulated due to treatment by plant hormones under lower salt stress during germination in rice seeds [90].
A change in the size and shape of cells always implies a modification or transformation of the cytoskeleton [91,92]. Salt effects induce significant disturbances in the interaction between the elements of the cytoskeleton, in some cases caused by the toxic effect of ions and in some cases by osmotic or oxidative damage affecting both the regulation of ion exchange and the fine mechanisms of the regulation and biosynthesis of various membrane and non-membrane proteins with the functions of transporters or enzymes [93,94]. The consequences of modifications of the cytoskeleton can affect cell growth, slowing down or enhancing the expansion, as well as changing the direction of future divisions. Moreover, this process can occur both locally in individual cells or tissues, and in all cells of proliferating tissues. In some cases, the damages to the cytoskeleton become irreversible, since its components can form disturbed structures, including crystalline ones, the assembly of which is not possible, and they remain in daughter cells after division [21,95].
Plant cells, for the most part, can withstand the negative effects of salts only at relatively low concentrations. In this case, the development of plants not only slows down, which could not have a significant effect on the ratio of symmetrical and asymmetric manifestations of growth. Salinity cannot cause the fundamentally different development of a certain genotype, but only introduces various inhibitory effects [31]. At the same time, multiple effects have been observed that cause disturbances, which intensify the asymmetry of the processes. To explain a significant change in development, as well as the appearance of the effect of disturbance of gravitropism, it can be speculated that an increase in chaotic elements in the structures of cytoskeleton, cell nuclei, mitochondria, plastids, and endoplasmic reticulum, as well as an unequal decrease in the amount of storage substances in cells, testifies to the fact that the processes accompanying growth and development, and providing metabolism, as well as rearrangement of the cytoskeleton, have different degrees of sensitivity to influences. Thus, while some processes proceed relatively normally, others are accompanied by an increase in asymmetry, which manifests itself in a shape change in compartments and structures. In addition, some processes can lead to the inhibition of dynamic transformations—for example, to the utilization of reserve substances. Moreover, in a number of cases, asymmetry leads to the impossibility of the logical completion of the beginning process of division of nuclei or organelles, which results in the appearance of cells that are uncharacteristic for a given organism—for example, differing in ploidy. The ploidy of plant cells is heterogeneous and this parameter is also sensitive to stress [96,97]. This effect was noted for a long time and was used in targeted breeding. In particular, it allows larger fruits to be obtained than wild forms and an increase in resistance to stress [98]. Salt-induced dysfunctions in some cells have caused abnormalities in the formation of nuclei. This was accompanied by the appearance of atypical polyploid or aneuploid cells and the appearance of invaginations in the nuclei under the action of salts, which probably indicates excessive production of the membrane material and the impossibility of maintaining the characteristic osmotic parameters in the nuclear compartment, since the “ideal” round shape that supports normal transport routes within the nucleus and between the nucleus and cytoplasm becomes unattainable [99]. It is also possible to note the violation of fusion, which provides a number of processes, including the formation of a large vacuole during growth by expansion.
4. Conclusions
The effect of salinity on germinating seedlings has significant effects on the growth and development of roots and their shape. In addition, there is a clear change in the uniformity of development, since the main and lateral roots are not equally suppressed. Disturbances in gravitropism also differ in the main and adventitious roots. The change in uniformity is the first sign of the manifestation of the intensification of the effects of asymmetry, which are characteristic of plant development, but do not have an inhibitory effect, at least in our understanding of the norm. This study demonstrates that the effects, manifested in the level of changes in size and shape, are accompanied by deep patterns of symmetry disorder at the level of such cellular structures as the cell membrane, nucleus, plastids, mitochondria, and cytoplasmic reticulum, exerting a mutual influence on all stages of the cytoskeleton transformation corresponding to different phases of the cell cycle. The result of uneven inhibition with a delay in some transformations in the presence of a stressor leads to a shift in the general habitual picture, which we conventionally consider as a norm or conditional symmetry, towards an increase in a certain “chaos” inherent to asymmetry. It is likely that, under the influence of these stressors, plant cells are displaced to the area of a kind of bifurcation point, as a result of which we can observe multiple and significant disturbances in the orderliness of cell structures, which are reflected in the processes of further development. Indeed, the unevenness of the normal course of differentiation processes and division, which provide visible effects of growth, provoked, probably, by the action of a stress factor, induces the effect of uneven development through the uneven disturbance of the dynamic transformations of plant compartments.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/sym13101811/s1, Figure S1: Features of the structure of the interphase cytoskeleton and distribution of αtubulin in wheat seedlings during germination in water and in the presence of NaCl and Na2SO4; Figure S2: Features of the structure of the preprophase cytoskeleton and distribution of α-tubulin in wheat seedlings during germination in water and in the presence of NaCl and Na2SO4; Figure S3: Features of the structure of the prometaphase cytoskeleton and distribution of α-tubulin in wheat seedlings during germination in water and in the presence of NaCl and Na2SO4; Figure S4: Features of the structure of the metaphase cytoskeleton and distribution of α-tubulin in wheat seedlings during germination in water and in the presence of NaCl and Na2SO4; Figure S5: Features of the structure of the anaphase cytoskeleton and distribution of α-tubulin in wheat seedlings during germination in water and in the presence of NaCl and Na2SO4; Figure S6: Structure features of the telophase cytoskeleton and distribution of α-tubulin in cells of wheat seedlings during germination in water and in the presence of NaCl and Na2SO4; Figure S7: Changes in the root cell shape occurring during growth in the meristem zone and differentiation as the volume of the cell vacuole in the extension zone increases.
Author Contributions
Conceptualization, E.N.B.; methodology, E.N.B.; software, E.N.B.; validation, A.A.G. and E.N.B.; formal analysis, E.N.B. and A.A.G.; investigation, E.N.B. and A.A.G.; resources, E.N.B. and A.A.G.; data curation, E.N.B. and A.A.G.; writing—original draft preparation, E.N.B.; writing—review and editing, E.N.B. and A.A.G.; visualization, E.N.B. and A.A.G.; supervision, A.A.G.; project administration, E.N.B.; funding acquisition, E.N.B. and A.A.G. Both authors have read and agreed to the published version of the manuscript.
Funding
The reported study was supported by assignments 0574-2019-0002 (All-Russia Research Institute of Agricultural Biotechnology) and 18-118021490111-5 (Tsitsin Main Botanical Garden of Russian Academy of Sciences) of the Ministry of Science and Higher Education of the Russian Federation.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data sharing is not applicable to this article.
Acknowledgments
The authors are thankful to Yu. V. Polyakov and Elena Kalinina-Terner for their kind help in setting up the experiment.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Mazzolai, B. Growth and tropism. In Living Machines: A Handbook of Research in Biomimetics and Biohybrid Systems; Prescott, T.J., Lepora, N., Verschure, P.F.M.J., Eds.; Oxford University Press Scholarship Online: Oxford, UK, 2018; pp. 99–104. [Google Scholar]
- Lucké, B.; McCutcheon, M. The living cell as an osmotic system and its permeability to water. Physiol. Rev. 1932, 12, 68–139. [Google Scholar] [CrossRef]
- Wehner, F.; Olsen, H.; Tinel, H.; Kinne-Saffran, E.; Kinne, R.K. Cell volume regulation: Osmolytes, osmolyte transport, and signal transduction. Rev. Physiol. Biochem. Pharmacol. 2003, 148, 1–80. [Google Scholar] [PubMed]
- Thery, M.; Bornens, M. Cell shape and cell division. Curr. Opin. Cell Biol. 2006, 18, 648–657. [Google Scholar] [CrossRef] [PubMed]
- Paluch, E.; Heisenberg, C.P. Biology and physics of cell shape changes in development. Curr. Biol. 2009, 19, R790–R799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, M.; Roeder, A.H. Plants are better engineers: The complexity of plant organ morphogenesis. Curr. Opin. Genet. Dev. 2020, 63, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Evans, D.E. Aerenchyma formation. New Phytol. 2004, 161, 35–49. [Google Scholar] [CrossRef]
- Kordyum, E.L.; Shevchenko, G.V.; Brykov, V.O. Cytoskeleton during aerenchyma formation in plants. Cell Biol. Int. 2019, 43, 991–998. [Google Scholar] [CrossRef]
- Daneva, A.; Gao, Z.; Van Durme, M.; Nowack, M.K. Functions and regulation of programmed cell death in plant development. Ann. Rev. Cell Dev. Biol. 2016, 32, 441–468. [Google Scholar] [CrossRef] [PubMed]
- Gamalei, Y.V. Supercellular plant organization. Russ. J. Plant Physiol. 1997, 44, 706–730. [Google Scholar]
- Brenner, E.D.; Stahlberg, R.; Mancuso, S.; Vivanco, J.; Baluška, F.; Van Volkenburgh, E. Plant neurobiology: An integrated view of plant signaling. Trends Plant Sci. 2006, 11, 413–419. [Google Scholar] [CrossRef] [PubMed]
- Baluška, F.; Volkmann, D.; Barlow, P.W. Eukaryotic cells and their cell bodies: Cell theory revised. Ann. Bot. 2004, 94, 9–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rudall, P.J.; Bateman, R.M. Coenocytic growth phases in land plant development: A paleo-evo-devo perspective. Int. J. Plant Sci. 2019, 180, 607–622. [Google Scholar] [CrossRef]
- Soukup, A.; Tylová, E. Apoplastic barriers: Their structure and function from a historical perspective. Concepts Cell Biol.-Hist. Evol. 2018, 23, 155–183. [Google Scholar]
- Fruleux, A.; Verger, S.; Boudaoud, A. Feeling stressed or strained? A biophysical model for cell wall mechanosensing in plants. Front. Plant Sci. 2019, 10, 757. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Mao, T. Understanding the functions and mechanisms of plant cytoskeleton in response to environmental signals. Curr. Opin. Plant Biol. 2019, 52, 86–96. [Google Scholar] [CrossRef] [PubMed]
- Blume, Y.B.; Krasylenko, Y.A.; Yemets, A.I. The role of the plant cytoskeleton in phytohormone signaling under abiotic and biotic stresses. Mech. Plant Horm. Signal. Stress 2017, 127–185. [Google Scholar]
- Szymanski, D.B.; Cosgrove, D.J. Dynamic coordination of cytoskeletal and cell wall systems during plant cell morphogenesis. Curr. Biol. 2009, 19, R800–R811. [Google Scholar] [CrossRef] [Green Version]
- Rasmussen, C.G.; Wright, A.J.; Müller, S. The role of the cytoskeleton and associated proteins in determination of the plant cell division plane. Plant J. 2013, 75, 258–269. [Google Scholar] [CrossRef] [Green Version]
- Smertenko, A.; Assaad, F.; Baluška, F.; Bezanilla, M.; Buschmann, H.; Drakakaki, G.; Žárský, V. Plant cytokinesis: Terminology for structures and processes. Trends Cell Biol. 2017, 27, 885–894. [Google Scholar] [CrossRef] [PubMed]
- Baranova, E.N.; Christov, N.K.; Kurenina, L.V.; Khaliluev, M.R.; Todorovska, E.G.; Smirnova, E.A. Formation of atypical tubulin structures in plant cells as a nonspecific response to abiotic stress. Bulg. J. Agric. Sci. 2016, 22, 987–992. [Google Scholar]
- Livanos, P.; Galatis, B.; Apostolakos, P. The interplay between ROS and tubulin cytoskeleton in plants. Plant Signal. Behav. 2014, 9, e28069. [Google Scholar] [CrossRef]
- Miake, H.; Mitsuya, S.; Rahman, M.S. Ultrastructural effects of salinity stress in higher plants. In Abiotic Stress Tolerance in Plants; Rai, A.K., Takabe, T., Eds.; Springer: Dordrecht, The Netherlands, 2006; pp. 215–226. [Google Scholar]
- Baranova, E.N.; Chaban, I.A.; Kononenko, N.V.; Gulevich, A.A.; Kurenina, L.V.; Smirnova, E.A. Ultrastructural changes of organelles in root cap cells of tobacco under salinity. Proc. Latv. Acad. Sci. Sec. B 2019, 73, 47–55. [Google Scholar] [CrossRef] [Green Version]
- Gao, H.J.; Yang, H.Y.; Bai, J.P.; Liang, X.Y.; Lou, Y.; Zhang, J.L.; Wang, D.; Zhang, J.L.; Niu, S.Q.; Chen, Y.L. Ultrastructural and physiological responses of potato (Solanum tuberosum L.) plantlets to gradient saline stress. Front. Plant Sci. 2015, 5, 787. [Google Scholar] [CrossRef] [PubMed]
- Oparka, K.J. Plasmolysis: New insights into an old process. New Phytol. 1994, 126, 571–591. [Google Scholar] [CrossRef]
- Hu, Y.; Fromm, J.; Schmidhalter, U. Effect of salinity on tissue architecture in expanding wheat leaves. Planta 2005, 220, 838–848. [Google Scholar] [CrossRef]
- Ogawa, A.; Kitamichi, K.; Toyofuku, K.; Kawashima, C. Quantitative analysis of cell division and cell death in seminal root of rye under salt stress. Plant Prod. Sci. 2006, 9, 56–64. [Google Scholar] [CrossRef]
- Polyakov, V.Y.; Lazareva, E.M.; Baranova, E.N.; Kharchenko, P.N. Method for Testing the Salt Tolerance of a Crop. RU 2520744 C1, 27 June 2014. [Google Scholar]
- Shi, D.; Sheng, Y. Effect of various salt–alkaline mixed stress conditions on sunflower seedlings and analysis of their stress factors. Environ. Exp. Bot. 2005, 54, 8–21. [Google Scholar] [CrossRef]
- Acosta-Motos, J.R.; Ortuño, M.F.; Bernal-Vicente, A.; Diaz-Vivancos, P.; Sanchez-Blanco, M.J.; Hernandez, J.A. Plant responses to salt stress: Adaptive mechanisms. Agronomy 2017, 7, 18. [Google Scholar] [CrossRef] [Green Version]
- Mahalingam, R. Consideration of combined stress: A crucial paradigm for improving multiple stress tolerance in plants. In Combined Stresses in Plants; Springer: Cham, Switzerland, 2015; pp. 1–25. [Google Scholar]
- Afzal, I.; Rauf, S.; Basra, S.M.A.; Murtaza, G. Halopriming improves vigor, metabolism of reserves and ionic contents in wheat seedlings under salt stress. Plant Soil Environ. 2008, 54, 382–388. [Google Scholar] [CrossRef] [Green Version]
- Figueroa-Bustos, V.; Palta, J.A.; Chen, Y.; Siddique, K.H. Characterization of root and shoot traits in wheat cultivars with putative differences in root system size. Agronomy 2018, 8, 109. [Google Scholar] [CrossRef] [Green Version]
- Marchant, A.; Kargul, J.; May, S.T.; Muller, P.; Delbarre, A.; Perrot-Rechenmann, C.; Bennett, M.J. AUX1 regulates root gravitropism in Arabidopsis by facilitating auxin uptake within root apical tissues. EMBO J. 1999, 18, 2066–2073. [Google Scholar] [CrossRef] [PubMed]
- Baranova, E.N.; Gulevich, A.A.; Polyakov, V.Y. Effect of NaCl, Na2SO4, and mannitol on utilization of storage starch and formation of plastids in the cotyledons and roots of alfalfa seedlings. Russ. J. Plant Physiol. 2007, 54, 50–57. [Google Scholar] [CrossRef]
- Ibrahim, E.A. Seed priming to alleviate salinity stress in germinating seeds. J. Plant Physiol. 2016, 192, 38–46. [Google Scholar] [CrossRef]
- Liu, B.; Palevitz, B.A. Organization of cortical microfilaments in dividing root cells. Cell Motil. Cytoskel. 1992, 23, 252–264. [Google Scholar] [CrossRef]
- He, F.; Chen, H.; Han, R. The plant cytoskeleton and crosslinking factors. CellBio 2020, 9, 85. [Google Scholar] [CrossRef]
- Galatis, B.; Apostolakos, P.; Palafoutas, D. Studies on the formation of ‘floating’guard cell mother cells in Anemia. J. Cell Sci. 1986, 80, 29–55. [Google Scholar] [CrossRef]
- Nick, P. Microtubules, signalling and abiotic stress. Plant J. 2013, 75, 309–323. [Google Scholar] [CrossRef]
- Smertenko, A.; Draber, P.; Viklický, V.; Opatrný, Z. Heat stress affects the organization of microtubules and cell division in Nicotiana tabacum cells. Plant Cell Environ. 1997, 20, 1534–1542. [Google Scholar] [CrossRef]
- Potters, G.; Pasternak, T.P.; Guisez, Y.; Palme, K.J.; Jansen, M.A. Stress-induced morphogenic responses: Growing out of trouble? Trends Plant Sci. 2007, 12, 98–105. [Google Scholar] [CrossRef]
- Guerriero, G.; Hausman, J.F.; Cai, G. No stress! Relax! Mechanisms governing growth and shape in plant cells. Int. J. Mol. Sci. 2014, 15, 5094–5114. [Google Scholar] [CrossRef] [Green Version]
- Wan, L.; Zhang, H. Cadmium toxicity: Effects on cytoskeleton, vesicular trafficking and cell wall construction. Plant Signal. Behav. 2012, 7, 345–348. [Google Scholar] [CrossRef] [Green Version]
- Örvar, B.L.; Sangwan, V.; Omann, F.; Dhindsa, R.S. Early steps in cold sensing by plant cells: The role of actin cytoskeleton and membrane fluidity. Plant J. 2000, 23, 785–794. [Google Scholar] [CrossRef] [PubMed]
- Lipka, E.; Herrmann, A.; Mueller, S. Mechanisms of plant cell division. Wiley Interdiscip. Rev. Dev. Biol. 2015, 4, 391–405. [Google Scholar] [CrossRef] [PubMed]
- Shoji, T.; Suzuki, K.; Abe, T.; Kaneko, Y.; Shi, H.; Zhu, J.K.; Rus, A.; Hasegawa, P.M.; Hashimoto, T. Salt stress affects cortical microtubule organization and helical growth in Arabidopsis. Plant Cell Physiol. 2006, 47, 1158–1168. [Google Scholar] [CrossRef] [PubMed]
- Sivaguru, M.; Baluska, F.; Volkmann, D.; Felle, H.H.; Horst, W.J. Impacts of aluminum on the cytoskeleton of the maize root apex. Short-term effects on the distal part of the transition zone. Plant Physiol. 1999, 119, 1073–1082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, K.L.; Hickson, I.D. New insights into the formation and resolution of ultra-fine anaphase bridges. In Seminars in Cell & Developmental Biology; Academic Press: Cambridge, MA, USA, 2011; Volume 22, pp. 906–912. [Google Scholar]
- Rasmussen, C.G.; Humphries, J.A.; Smith, L.G. Determination of symmetric and asymmetric division planes in plant cells. Ann. Rev. Plant Biol. 2011, 62, 387–409. [Google Scholar] [CrossRef] [Green Version]
- Komis, G.; Galatis, B. Altered patterns of tubulin polymerization in dividing leaf cells of Chlorophyton comosum after a hyperosmotic treatment. New Phytol. 2001, 149, 193–207. [Google Scholar] [CrossRef] [PubMed]
- Qin, R.; Wang, C.; Chen, D.; Björn, L.O.; Li, S. Copper-induced root growth inhibition of Allium cepa var. agrogarum L. involves disturbances in cell division and DNA damage. Environ. Toxicol. Chem. 2015, 34, 1045–1055. [Google Scholar] [CrossRef]
- Bashline, L.; Lei, L.; Li, S.; Gu, Y. Cell wall, cytoskeleton, and cell expansion in higher plants. Mol. Plant 2014, 7, 586–600. [Google Scholar] [CrossRef] [Green Version]
- Soltani, A.; Gholipoor, M.; Zeinali, E. Seed reserve utilization and seedling growth of wheat as affected by drought and salinity. Environ. Exp. Bot. 2006, 55, 195–200. [Google Scholar] [CrossRef]
- Sebei, K.; Debez, A.; Herchi, W.; Boukhchina, S.; Kallel, H. Germination kinetics and seed reserve mobilization in two flax (Linum usitatissimum L.) cultivars under moderate salt stress. J. Plant Biol. 2007, 50, 447–454. [Google Scholar] [CrossRef]
- Voigt, E.L.; Almeida, T.D.; Chagas, R.M.; Ponte, L.F.A.; Viégas, R.A.; Silveira, J.A.G. Source–sink regulation of cotyledonary reserve mobilization during cashew (Anacardium occidentale) seedling establishment under NaCl salinity. J. Plant Physiol. 2009, 166, 80–89. [Google Scholar] [CrossRef]
- Baranova, E.N.; Gulevich, A.A.; Kalinina-Turner, E.B.; Koslov, N.N. Effects of NaCl, Na2SO4 and mannitol on utilization of storage protein and transformation of vacuoles in the cotyledons and seedling roots of alfalfa (Medicago sativa L.). Russ. Agric. Sci. 2011, 37, 11–19. [Google Scholar] [CrossRef]
- Alencar, N.L.; Gadelha, C.G.; Gallão, M.I.; Dolder, M.A.; Prisco, J.T.; Gomes-Filho, E. Ultrastructural and biochemical changes induced by salt stress in Jatropha curcas seeds during germination and seedling development. Funct. Plant Biol. 2015, 42, 865–874. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, M.Y.; Afaf, R.; Qureshi, M.S.; Sarwar, G.; Naqvi, M.H. Salinity induced changes in α-amylase and protease activities and associated metabolism in cotton varieties during germination and early seedling growth stages. Acta Physiol. Plant. 2002, 24, 37–44. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, J.; Yang, F.; Zhou, W.; Mao, S.; Lin, J.; Yan, X. Untargeted LC-MS-based metabolomics revealed specific metabolic changes in cotyledons and roots of Ricinus communis during early seedling establishment under salt stress. Plant Physiol. Biochem. 2021, 163, 108–118. [Google Scholar] [CrossRef]
- Grigorova, B.; Vassileva, V.; Klimchuk, D.; Vaseva, I.; Demirevska, K.; Feller, U. Drought, high temperature, and their combination affect ultrastructure of chloroplasts and mitochondria in wheat (Triticum aestivum L.) leaves. J. Plant Interact. 2012, 7, 204–213. [Google Scholar] [CrossRef] [Green Version]
- Smith, M.M.; Hodson, M.J.; Öpik, H.; Wainwright, S.J. Salt-induced ultrastructural damage to mitochondria in root tips of a salt-sensitive ecotype of Agrostis stolonifera. J. Exp. Bot. 1982, 33, 886–895. [Google Scholar] [CrossRef]
- Mathur, J.; Mammone, A.; Barton, K.A. Organelle extensions in plant cells. J. Integr. Plant Biol. 2012, 54, 851–867. [Google Scholar] [CrossRef] [PubMed]
- Khona, D.K.; Shirolikar, S.M.; Gawde, K.K.; Hom, E.; Deodhar, M.A.; D’Souza, J.S. Characterization of salt stress-induced palmelloids in the green alga, Chlamydomonas reinhardtii. Algal Res. 2016, 16, 434–448. [Google Scholar] [CrossRef]
- Saprunova, V.B.; Bakeeva, L.E.; Yaguzhinskiy, L.S. Ultrastructure of mitochondrial mechanism of apoptotic rat cardiomyocytes induced by prolonged anoxia. Cytology 2003, 11, 1074–1082. [Google Scholar]
- Osteryoung, K.W.; Pyke, K.A. Division and dynamic morphology of plastids. Ann. Rev. Plant Biol. 2014, 65, 443–472. [Google Scholar] [CrossRef] [PubMed]
- Song, G.; Xiang, X.; Wang, Z.; Li, R. Polyphasic characterization of Stigonema dinghuense, sp. nov. (Cyanophyceae, Nostocophycidae, Stigonemaceae), from Dinghu Mountain, south China. Phytotaxa 2015, 213, 212–224. [Google Scholar] [CrossRef] [Green Version]
- Bisgrove, S.R.; Kropf, D.L. Cytokinesis in brown algae: Studies of asymmetric division in fucoid zygotes. Protoplasma 2004, 223, 163–173. [Google Scholar] [CrossRef]
- Buschmann, H.; Zachgo, S. The evolution of cell division: From streptophyte algae to land plants. Trends Plant Sci. 2016, 21, 872–883. [Google Scholar] [CrossRef] [PubMed]
- Brown, R.C.; Lemmon, B.E.; Graham, L.E. Morphogenetic plastid migration and microtubule arrays in mitosis and cytokinesis in the green-alga Coleochaete Orbicularis. Am. J. Bot. 1994, 81, 127–133. [Google Scholar] [CrossRef]
- Herman, E.M.; Larkins, B.A. Protein storage bodies and vacuoles. Plant Cell 1999, 11, 601–613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaughn, K. Protein Bodies/Vacuoles and Cytoplasm. In Immunocytochemistry of Plant Cells; Springer: Dordrecht, The Netherlands, 2013; pp. 123–127. [Google Scholar]
- Kapilan, R.; Vaziri, M.; Zwiazek, J.J. Regulation of aquaporins in plants under stress. Biol. Res. 2018, 51, 1–11. [Google Scholar] [CrossRef]
- Li, G.W.; Peng, Y.H.; Yu, X.; Zhang, M.H.; Cai, W.M.; Sun, W.N.; Su, W.A. Transport functions and expression analysis of vacuolar membrane aquaporins in response to various stresses in rice. J. Plant Physiol. 2008, 165, 1879–1888. [Google Scholar] [CrossRef] [PubMed]
- Kononenko, N.; Baranova, E.; Dilovarova, T.; Akanov, E.; Fedoreyeva, L. Oxidative damage to various root and shoot tissues of durum and soft wheat seedlings during salinity. Agriculture 2020, 10, 55. [Google Scholar] [CrossRef] [Green Version]
- Baranova, E.N.; Bakulina, A.V.; Shirokikh, I.G.; Shuplechova, O.N.; Chaban, I.A.; Gulevich, A.A.; Polyakov, V.Y. The influence of aluminum on the structural organization of the roots of barley in culture in vitro. Russ. Agric. Sci. 2011, 37, 358–363. [Google Scholar] [CrossRef]
- Dos Santos, N.M.C.; Amaral, W.G.; de Araújo, F.V.; Ferreira, E.A.; Castro, B.M.; Zanuncio, J.C.; Pereira, I.M.; Dos Santos, J.B. Effects of the herbicide trifluralin in the initial development of Piptadenia gonoacantha (Fabales: Fabaceae). Int. J. Phytoremed. 2019, 21, 1323–1328. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Feng, G.; Zhang, Z.; Huang, M.; Qi, W.; Ma, L. Geometrical and statistical analysis of dynamic crack morphology in shrink-swell soils with addition of maize roots or salinity (NaCl). Soil Till. Res. 2021, 212, 105057. [Google Scholar] [CrossRef]
- Chu, F.Y.; Haley, S.C.; Zidovska, A. On the origin of shape fluctuations of the cell nucleus. Proc. Natl. Acad. Sci. USA 2017, 114, 10338–10343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barro, A.V.; Stoeckle, D.; Thellmann, M.; Ruiz-Duarte, P.; Bald, L.; Louveaux, M.; von Born, P.; Denninger, P.; Goh, T.; Fukaki, H.; et al. Cytoskeleton dynamics are necessary for early events of lateral root initiation in Arabidopsis. Curr. Biol. 2019, 29, 2443–2454. [Google Scholar] [CrossRef] [Green Version]
- Dhonukshe, P.; Kleine-Vehn, J.; Friml, J. Cell polarity, auxin transport, and cytoskeleton-mediated division planes: Who comes first? Protoplasma 2005, 226, 67–73. [Google Scholar] [CrossRef]
- Sato, E.M.; Hijazi, H.; Bennett, M.J.; Vissenberg, K.; Swarup, R. New insights into root gravitropic signalling. J. Exp. Bot. 2015, 66, 2155–2165. [Google Scholar] [CrossRef]
- Zhang, Y.; He, P.; Ma, X.; Yang, Z.; Pang, C.; Yu, J.; Wang, G.; Friml, J.; Xiao, G. Auxin-mediated statolith production for root gravitropism. New Phytol. 2019, 224, 761–774. [Google Scholar] [CrossRef] [PubMed]
- Almansouri, M.; Kinet, J.M.; Lutts, S. Effect of salt and osmotic stresses on germination in durum wheat (Triticum durum Desf.). Plant Soil 2001, 231, 243–254. [Google Scholar] [CrossRef]
- David, A.; Yadav, S.; Bhatla, S.C. Sodium chloride stress induces nitric oxide accumulation in root tips and oil body surface accompanying slower oleosin degradation in sunflower seedlings. Physiol. Plant. 2010, 140, 342–354. [Google Scholar] [CrossRef] [PubMed]
- Yan, M.; Xue, C.; Xiong, Y.; Meng, X.; Li, B.; Shen, R.; Lan, P. Proteomic dissection of the similar and different responses of wheat to drought, salinity and submergence during seed germination. J. Proteom. 2020, 220, 103756. [Google Scholar] [CrossRef]
- Toosi, A.F.; Bakar, B.B.; Azizi, M. Effect of drought stress by using PEG 6000 on germination and early seedling growth of Brassica juncea Var. Ensabi. Sci. Pap. Ser. A Agron. 2014, 57, 360–363. [Google Scholar]
- Almaghrabi, O.A. Impact of drought stress on germination and seedling growth parameters of some wheat cultivars. Life Sci. J. 2012, 9, 590–598. [Google Scholar]
- Kim, S.K.; Son, T.K.; Park, S.Y.; Lee, I.J.; Lee, B.H.; Kim, H.Y.; Lee, S.C. Influences of gibberellin and auxin on endogenous plant hormone and starch mobilization during rice seed germination under salt stress. J. Environ. Biol. 2006, 27, 181. [Google Scholar]
- Smith, L.G. Cytoskeletal control of plant cell shape: Getting the fine points. Curr. Opin. Plant Biol. 2003, 6, 63–73. [Google Scholar] [CrossRef]
- Lian, N.; Wang, X.; Jing, Y.; Lin, J. Regulation of cytoskeleton-associated protein activities: Linking cellular signals to plant cytoskeletal function. J. Integr. Plant Biol. 2021, 63, 241–250. [Google Scholar] [CrossRef]
- Ali, A.S.; Elozeiri, A.A. Metabolic processes during seed germination. Adv. Seed Biol. 2017, 141–166. [Google Scholar]
- Gamalero, E.; Bona, E.; Todeschini, V.; Lingua, G. Saline and arid soils: Impact on bacteria, plants, and their interaction. Biology 2020, 9, 116. [Google Scholar] [CrossRef] [PubMed]
- Chun, H.J.; Baek, D.; Jin, B.J.; Cho, H.M.; Park, M.S.; Lee, S.H.; Lim, L.H.; Cha, Y.J.; Bae, D.W.; Kim, S.T.; et al. Microtubule dynamics plays a vital role in plant adaptation and tolerance to salt stress. Int. J. Mol. Sci. 2021, 22, 5957. [Google Scholar] [CrossRef]
- Kunakh, V.A. Genome variability in somatic plant cells. 7. Variability of population—Genetic parameters in the culture in vitro. Biopol. Cell 2002, 18, 377–393. [Google Scholar] [CrossRef] [Green Version]
- Van de Peer, Y.; Ashman, T.L.; Soltis, P.S.; Soltis, D.E. Polyploidy: An evolutionary and ecological force in stressful times. Plant Cell 2021, 33, 11–26. [Google Scholar] [CrossRef] [PubMed]
- Shah, Z.H.; Rehman, H.M.; Akhtar, T.; Daur, I.; Nawaz, M.A.; Ahmad, M.Q.; Rana, I.A.; Atif, R.M.; Yang, S.H.; Chung, G. Redox and ionic homeostasis regulations against oxidative, salinity and drought stress in wheat (A systems biology approach). Front. Genet. 2017, 8, 141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baranova, E.N.; Chaban, I.A.; Kononenko, N.V.; Khaliluev, M.R.; Christov, N.K.; Gulevich, A.A.; Todorovska, E.G. Ultrastructural organization of the domains in the cell nucleus of dicotyledonous and monocotyledonous plants under abiotic stress. Russ. Agric. Sci. 2017, 43, 199–206. [Google Scholar] [CrossRef]
Figure 1.
Formation of the primary and adventitious roots of the first and second order in wheat seedlings during germination in water and in the presence of NaCl and Na2SO4. Histogram of the differences in length of main root and lateral adventitious roots of the first and second orders. The idealized model (gray histogram) has been plotted using the maximum values of real experimental data (yellow histograms) without taking into account the experimental conditions. It can be seen that under the influence of NaCl, the growth of the primary root and adventive roots of the first order is inhibited. This is compensated by the growth of second-order adventive roots. Na2SO4 further inhibits the growth of main roots and adventitious roots of the first order. Thus, the red lines indicate the appropriateness between the values in the idealized histogram and the maximum values in real experimental patterns on the fourth germination day. Designations: pr—primary root, ar—adventitious root, *—indicates significant differences between ideal model and real experimental values at p = 0.05.
Figure 1.
Formation of the primary and adventitious roots of the first and second order in wheat seedlings during germination in water and in the presence of NaCl and Na2SO4. Histogram of the differences in length of main root and lateral adventitious roots of the first and second orders. The idealized model (gray histogram) has been plotted using the maximum values of real experimental data (yellow histograms) without taking into account the experimental conditions. It can be seen that under the influence of NaCl, the growth of the primary root and adventive roots of the first order is inhibited. This is compensated by the growth of second-order adventive roots. Na2SO4 further inhibits the growth of main roots and adventitious roots of the first order. Thus, the red lines indicate the appropriateness between the values in the idealized histogram and the maximum values in real experimental patterns on the fourth germination day. Designations: pr—primary root, ar—adventitious root, *—indicates significant differences between ideal model and real experimental values at p = 0.05.

Figure 2.
Scheme of seedling development during germination under hypothetical, “ideal” conditions, in water, and in the presence of NaCl and Na2SO4. One can note the asymmetric development of lateral roots, the unevenness of their development in the control model, the appearance of root bends in the presence of NaCl, and an enhancing of this effect, accompanied by gravitropism, under the influence of Na2SO4. Designations: (1)—dry kernel, (2)—swollen kernel, (3)—germinated kernel, where the primary root (radicle) emerged from the coleorhiza, (4)—a seedling with a primary and two lateral roots and a changed coleoptile, (5)—a seedling with a formed main and lateral roots and an enlarged coleoptile, (6)—seedling with a grown coleoptile, a visible and first leaf, the primary (main) root, and adventitious (lateral) roots of the first and second order. Designations: pr—primary root, ar1—adventitious root of first order, ar2—adventitious root of second order.
Figure 2.
Scheme of seedling development during germination under hypothetical, “ideal” conditions, in water, and in the presence of NaCl and Na2SO4. One can note the asymmetric development of lateral roots, the unevenness of their development in the control model, the appearance of root bends in the presence of NaCl, and an enhancing of this effect, accompanied by gravitropism, under the influence of Na2SO4. Designations: (1)—dry kernel, (2)—swollen kernel, (3)—germinated kernel, where the primary root (radicle) emerged from the coleorhiza, (4)—a seedling with a primary and two lateral roots and a changed coleoptile, (5)—a seedling with a formed main and lateral roots and an enlarged coleoptile, (6)—seedling with a grown coleoptile, a visible and first leaf, the primary (main) root, and adventitious (lateral) roots of the first and second order. Designations: pr—primary root, ar1—adventitious root of first order, ar2—adventitious root of second order.

Figure 3.
Scheme of transformations and characteristic disorders of the tubulin cytoskeleton and distribution of α-tubulin and material of the nucleus and nucleolus in wheat seedlings when germinating in water and in the presence of NaCl and Na2SO4. Designations: pink—α-tubulin in the cytoplasm, red—α-tubulin in the form of microtubules, blue—condensed chromatin, crimson—nucleolus, glaucous—decondensed chromatin, gray—nucleoplasm, lilac framed by glaucous—chromosomes.
Figure 3.
Scheme of transformations and characteristic disorders of the tubulin cytoskeleton and distribution of α-tubulin and material of the nucleus and nucleolus in wheat seedlings when germinating in water and in the presence of NaCl and Na2SO4. Designations: pink—α-tubulin in the cytoplasm, red—α-tubulin in the form of microtubules, blue—condensed chromatin, crimson—nucleolus, glaucous—decondensed chromatin, gray—nucleoplasm, lilac framed by glaucous—chromosomes.

Figure 4.
Ultrastructure of the apical meristem cells of the shoot and root of wheat seedlings when germinating in water. General view of the cells of juvenile cortex tissue, located beyond the initials in the root differentiation zone—(a); enlarged fragments of the cytoplasm containing the organelles (plastids, mitochondria and vacuoles)—(c,e). General view of the parenchyma cells of developing shoot in the differentiation zone—(b); enlarged fragments of the cytoplasm containing the organelles (plastids, mitochondria and vacuoles)—(d,f). Abbreviations: v—vacuole, cw—cell wall, s—starch, m—mitochondrion, p—plastid, N—nucleus, d—dictiosome, n—nucleolus.
Figure 4.
Ultrastructure of the apical meristem cells of the shoot and root of wheat seedlings when germinating in water. General view of the cells of juvenile cortex tissue, located beyond the initials in the root differentiation zone—(a); enlarged fragments of the cytoplasm containing the organelles (plastids, mitochondria and vacuoles)—(c,e). General view of the parenchyma cells of developing shoot in the differentiation zone—(b); enlarged fragments of the cytoplasm containing the organelles (plastids, mitochondria and vacuoles)—(d,f). Abbreviations: v—vacuole, cw—cell wall, s—starch, m—mitochondrion, p—plastid, N—nucleus, d—dictiosome, n—nucleolus.

Figure 5.
Ultrastructure of the apical meristem cells of the shoot and root of wheat seedlings during germination in the presence of NaCl. General view of the cells of juvenile cortex tissue, located beyond the initials in the root differentiation zone—(a); enlarged fragments of the cytoplasm containing the organelles (plastids, mitochondria and vacuoles)—(c,e). General view of the parenchyma cells of developing shoot in the differentiation zone—(b); enlarged fragments of the cytoplasm containing the organelles (plastids, mitochondria and vacuoles)—(d,f). Abbreviations: v—vacuole, cw—cell wall, s—starch, m—mitochondrion, p—plastid, N—nucleus, n—nucleolus, d—dictiosome, l—oil body.
Figure 5.
Ultrastructure of the apical meristem cells of the shoot and root of wheat seedlings during germination in the presence of NaCl. General view of the cells of juvenile cortex tissue, located beyond the initials in the root differentiation zone—(a); enlarged fragments of the cytoplasm containing the organelles (plastids, mitochondria and vacuoles)—(c,e). General view of the parenchyma cells of developing shoot in the differentiation zone—(b); enlarged fragments of the cytoplasm containing the organelles (plastids, mitochondria and vacuoles)—(d,f). Abbreviations: v—vacuole, cw—cell wall, s—starch, m—mitochondrion, p—plastid, N—nucleus, n—nucleolus, d—dictiosome, l—oil body.

Figure 6.
Ultrastructure of the apical meristem cells of the shoot and root of wheat seedlings during germination in the presence of Na2SO4. General view of the cells of juvenile cortex tissue, located beyond the initials in the root differentiation zone—(a); enlarged fragments of the cytoplasm containing the organelles (plastids, mitochondria and vacuoles)—(c,e). General view of the parenchyma cells of developing shoot in the differentiation zone—(b); enlarged fragments of the cytoplasm containing the organelles (plastids, mitochondria and vacuoles)—(d,f). Abbreviations: v—vacuole, cw—cell wall, s—starch, m—mitochondrion, p—plastid, N—nucleus, n—nucleolus, d—dictiosome, l—oil body.
Figure 6.
Ultrastructure of the apical meristem cells of the shoot and root of wheat seedlings during germination in the presence of Na2SO4. General view of the cells of juvenile cortex tissue, located beyond the initials in the root differentiation zone—(a); enlarged fragments of the cytoplasm containing the organelles (plastids, mitochondria and vacuoles)—(c,e). General view of the parenchyma cells of developing shoot in the differentiation zone—(b); enlarged fragments of the cytoplasm containing the organelles (plastids, mitochondria and vacuoles)—(d,f). Abbreviations: v—vacuole, cw—cell wall, s—starch, m—mitochondrion, p—plastid, N—nucleus, n—nucleolus, d—dictiosome, l—oil body.

Figure 7.
Ultrastructure of the apical meristem cells of the shoot and root of wheat seedlings during germination in the presence of mannitol. General view of the cells of juvenile cortex tissue, located beyond the initials in the root differentiation zone—(a); enlarged fragments of the cytoplasm containing the organelles (plastids, mitochondria and vacuoles)—(c,e). General view of the parenchyma cells of developing shoot in the differentiation zone—(b); enlarged fragments of the cytoplasm containing the organelles (plastids, mitochondria and vacuoles)—(d,f). Abbreviations: v—vacuole, cw—cell wall, s—starch, m—mitochondrion, p—plastid, N—nucleus, n—nucleolus, d—dictyosome, l—oil body.
Figure 7.
Ultrastructure of the apical meristem cells of the shoot and root of wheat seedlings during germination in the presence of mannitol. General view of the cells of juvenile cortex tissue, located beyond the initials in the root differentiation zone—(a); enlarged fragments of the cytoplasm containing the organelles (plastids, mitochondria and vacuoles)—(c,e). General view of the parenchyma cells of developing shoot in the differentiation zone—(b); enlarged fragments of the cytoplasm containing the organelles (plastids, mitochondria and vacuoles)—(d,f). Abbreviations: v—vacuole, cw—cell wall, s—starch, m—mitochondrion, p—plastid, N—nucleus, n—nucleolus, d—dictyosome, l—oil body.

Figure 8.
Ultrastructure of specific new forms of the endoplasmic reticulum in the apical meristem cells of the shoot and root of wheat seedlings during germination in the presence of Na2SO4. Fragments of the parenchyma cells of the developing shoot with changes in the morphology of endoplasmic reticulum (a–d); the expanded cisternae of the smooth endoplasmic reticulum are not surrounded by dictyosomes—(a,b,d); the forming area of the smooth endoplasmic reticulum is surrounded by dictyosomes—(c). Fragments of cells in the differentiation zone of the root cortex—(e,f); the rough endoplasmic reticulum is located near the nucleus—(e); the areas of the rough reticulum are clearly identified and not detected in other parts of cell—(f). Abbreviations: v—vacuole, cw—cell wall, s—starch, m—starch, N—nucleus, n—nucleolus, d—dictyosome, l—oil body.
Figure 8.
Ultrastructure of specific new forms of the endoplasmic reticulum in the apical meristem cells of the shoot and root of wheat seedlings during germination in the presence of Na2SO4. Fragments of the parenchyma cells of the developing shoot with changes in the morphology of endoplasmic reticulum (a–d); the expanded cisternae of the smooth endoplasmic reticulum are not surrounded by dictyosomes—(a,b,d); the forming area of the smooth endoplasmic reticulum is surrounded by dictyosomes—(c). Fragments of cells in the differentiation zone of the root cortex—(e,f); the rough endoplasmic reticulum is located near the nucleus—(e); the areas of the rough reticulum are clearly identified and not detected in other parts of cell—(f). Abbreviations: v—vacuole, cw—cell wall, s—starch, m—starch, N—nucleus, n—nucleolus, d—dictyosome, l—oil body.

Figure 10.
Ultrastructure of stress-induced changes in plastids of wheat root cells: normal configuration (a), altered shape with invaginations (b), cup-shaped plastids (c), highly elongated curved plastids preserving width (d,e). In addition, the shape of organelles and a change in their structure from a rounded shape characteristic of juvenile organelles to a complex one—curved or cup-shaped with other types of symmetry—accompany damage and suggest that developmental asymmetry accompanies adaptation to negative influences, allowing them to maintain vitality by sacrificing a harmonious, close to symmetric picture of development.
Figure 10.
Ultrastructure of stress-induced changes in plastids of wheat root cells: normal configuration (a), altered shape with invaginations (b), cup-shaped plastids (c), highly elongated curved plastids preserving width (d,e). In addition, the shape of organelles and a change in their structure from a rounded shape characteristic of juvenile organelles to a complex one—curved or cup-shaped with other types of symmetry—accompany damage and suggest that developmental asymmetry accompanies adaptation to negative influences, allowing them to maintain vitality by sacrificing a harmonious, close to symmetric picture of development.

Figure 11.
Ultrastructure of stress-induced changes in mitochondrial division in wheat root cells: division by constriction (a), differences between mitochondria (b), division of mitochondria without constriction (c), formation of mitochondrial microrudiments (d).
Figure 11.
Ultrastructure of stress-induced changes in mitochondrial division in wheat root cells: division by constriction (a), differences between mitochondria (b), division of mitochondria without constriction (c), formation of mitochondrial microrudiments (d).

Figure 12.
Stress-induced changes in the process of plastid division in wheat root cells: division by constriction (a), differences between plastids (b), division of plastids without constriction (c), formation of micro-buds in fissionable plastids (d).
Figure 12.
Stress-induced changes in the process of plastid division in wheat root cells: division by constriction (a), differences between plastids (b), division of plastids without constriction (c), formation of micro-buds in fissionable plastids (d).

Figure 13.
Stress-induced changes in the process of vacuole fusion and utilization of storage substances during germination in wheat root cells: reduced internal osmotic pressure of vacuoles (a), fusion of vacuoles with impaired fusion of the vacuolar membrane (b), excessive production of vacuolar membranes in case of impaired fusion and growth limitation of the volume of vacuoles (c), the initial stage of utilization of protein deposits in the norm, lipid droplets along the walls (d,e), delay in utilization of protein deposits in vacuoles (f), fusion of vacuoles in the cells of the root tip during germination.
Figure 13.
Stress-induced changes in the process of vacuole fusion and utilization of storage substances during germination in wheat root cells: reduced internal osmotic pressure of vacuoles (a), fusion of vacuoles with impaired fusion of the vacuolar membrane (b), excessive production of vacuolar membranes in case of impaired fusion and growth limitation of the volume of vacuoles (c), the initial stage of utilization of protein deposits in the norm, lipid droplets along the walls (d,e), delay in utilization of protein deposits in vacuoles (f), fusion of vacuoles in the cells of the root tip during germination.

Figure 14.
Significant asymmetry of the shape of the cell wall and nucleus under the influence of salinity in the cells of the wheat root. Formation of the cell wall after division is normal (a), with the toxic effects of salts ((b)—NaCl, (c)—Na2SO4); the structural organization of the nucleus in interphase root cells is normal (d), under the action of salts ((e)—NaCl, (f)—Na2SO4).
Figure 14.
Significant asymmetry of the shape of the cell wall and nucleus under the influence of salinity in the cells of the wheat root. Formation of the cell wall after division is normal (a), with the toxic effects of salts ((b)—NaCl, (c)—Na2SO4); the structural organization of the nucleus in interphase root cells is normal (d), under the action of salts ((e)—NaCl, (f)—Na2SO4).

Figure 15.
Symmetrical and asymmetric processes affect the cell shape, the formation of cell walls, and the processes of the formation of nuclei and their shape in wheat root cells. Description: two cells in telophase in the initial stage of cell formation in the root parenchyma (a); the asymmetry of cells of the root central cylinder normally allows the similarity (symmetry) between cells to be maintained (b); disturbance in cell growth, division, and improper formation of cell walls lead to the emergence of cells of irregular shape (c), late telophase stage with the completed stage of cell wall formation (d), consequences of chromosome separation and the formation of a common nuclear envelope with invaginations under the influence of NaCl (e), the formation of large asymmetric nuclei in disturbance of divisions caused by Na2SO4 (c), the nuclei look as the formed organelles—but there is no cell wall between them, which indicates significant disorders of the fission process (f).
Figure 15.
Symmetrical and asymmetric processes affect the cell shape, the formation of cell walls, and the processes of the formation of nuclei and their shape in wheat root cells. Description: two cells in telophase in the initial stage of cell formation in the root parenchyma (a); the asymmetry of cells of the root central cylinder normally allows the similarity (symmetry) between cells to be maintained (b); disturbance in cell growth, division, and improper formation of cell walls lead to the emergence of cells of irregular shape (c), late telophase stage with the completed stage of cell wall formation (d), consequences of chromosome separation and the formation of a common nuclear envelope with invaginations under the influence of NaCl (e), the formation of large asymmetric nuclei in disturbance of divisions caused by Na2SO4 (c), the nuclei look as the formed organelles—but there is no cell wall between them, which indicates significant disorders of the fission process (f).

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Baranova, E.N.; Gulevich, A.A. Asymmetry of Plant Cell Divisions under Salt Stress. Symmetry 2021, 13, 1811. https://doi.org/10.3390/sym13101811
AMA Style
Baranova EN, Gulevich AA. Asymmetry of Plant Cell Divisions under Salt Stress. Symmetry. 2021; 13(10):1811. https://doi.org/10.3390/sym13101811
Chicago/Turabian StyleBaranova, Ekaterina N., and Alexander A. Gulevich. 2021. "Asymmetry of Plant Cell Divisions under Salt Stress" Symmetry 13, no. 10: 1811. https://doi.org/10.3390/sym13101811
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.