
It Is Not Just in the Genes


Research Division Experimental and Molecular Psychiatry, Department of Psychiatry, Psychotherapy and Preventive Medicine, LWL University Hospital, Ruhr-University Bochum, 44780 Bochum, Germany
Symmetry 2021, 13(10), 1815; https://doi.org/10.3390/sym13101815
Submission received: 4 September 2021
/
Revised: 22 September 2021
/
Accepted: 23 September 2021
/
Published: 29 September 2021
(This article belongs to the Special Issue Cognitive and Neurophysiological Models of Brain Asymmetry)
{kind=link}
{kind=link}
Abstract
:Asymmetries in the functional and structural organization of the nervous system are widespread in the animal kingdom and especially characterize the human brain. Although there is little doubt that asymmetries arise through genetic and nongenetic factors, an overarching model to explain the development of functional lateralization patterns is still lacking. Current genetic psychology collects data on genes relevant to brain lateralizations, while animal research provides information on the cellular mechanisms mediating the effects of not only genetic but also environmental factors. This review combines data from human and animal research (especially on birds) and outlines a multi-level model for asymmetry formation. The relative impact of genetic and nongenetic factors varies between different developmental phases and neuronal structures. The basic lateralized organization of a brain is already established through genetically controlled embryonic events. During ongoing development, hemispheric specialization increases for specific functions and subsystems interact to shape the final functional organization of a brain. In particular, these developmental steps are influenced by environmental experiences, which regulate the fine-tuning of neural networks via processes that are referred to as ontogenetic plasticity. The plastic potential of the nervous system could be decisive for the evolutionary success of lateralized brains.
1. The Functional Organization of Brain Asymmetries and Its Development
“A number of embryonic events make up an integrated overture to the post-hatching expression of lateralization”Lesley Rogers [1]
1.1. Lateralization Patterns of Neuronal Systems across the Animal Kingdom
A fundamental organizational principle of our brain is its asymmetries, which encompass both structural and functional differences between the two hemispheres. This characteristic has led to numerous hypotheses and research projects, which have attempted to elucidate the evolutionary and developmental origins of this specific trait [2,3]. However, lateralization of the brain is not specific to humans, but is present in many species across the animal kingdom. Not only vertebrates, but also many invertebrates, such as flies, bees, octopuses or nematodes, show left–right differences in neural organization and behavior [3,4,5,6,7,8,9], which suggests that lateralization is a common feature of metazoan nervous systems [10]. Neuronal asymmetries can be observed in all areas of information processing, including perception, cognition, emotion, homeostatic regulation or motor control and are based on neuroanatomical as well as physiological left–right differences [7,11,12]. Lateralization can be present at the individual level, with left-sided dominance for a certain function in half of a population and right-sided dominance in the other half. In other cases, the direction of a lateralized function within a population is aligned, so that lateralization is present at population level [2,11,13]. Comparative studies indicate that some aspects of functional brain lateralizations share a common evolutionary history [3,7,8,14]. It has been suggested that the vertebrate brain is characterized by specific functional dichotomy, with the left hemisphere more strongly involved in routine and approach behavior, while the right hemisphere dominates detection and response to unexpected, novel and potentially pivotal stimuli [15,16,17]. For example, several species of fish, amphibians and birds react faster when a predator approaches from the left, indicating that right-hemispheric networks are specialized for the detection of potential dangers, while foraging is controlled by the left hemisphere [3,7,8,18]. The processing of social stimuli, such as faces, is also dominated by the right hemisphere [19] in humans [20], sheep [21,22] and chicks [23]. On the other hand, at least in mammals, communicative signals, such as spoken language in humans [24] or other forms of conspecific vocalizations [25,26], are typically processed within the left hemisphere. A widespread behavioral indicator of hemispheric lateralization is the preferred use of one extremity, which has been documented in a variety of vertebrate and invertebrate species at individual and population levels [5,27,28,29,30,31]. In humans, handedness is the most obvious asymmetry with about 90% of individual preferring to use their right hand for complex manual tasks like fine-tuned object manipulation or writing [32]. Handedness is related to other behavioral asymmetries, like preferential foot use [33] or cradling bias [34], while deviations from the typical pattern are associated with psychiatric or developmental disorders [2,35,36,37]. Therefore, handedness is used as the favorite measure for correlating functional lateralization with structural left–right differences and genetic variations ([e.g., [12,32,36,38]).
1.2. Understanding Ontogeny of Neuronal Asymmetries—An Unfinished Business
Despite increasing knowledge about the relationships between different functional lateralizations and their structural foundations, our understanding of the underlying ontogenetic mechanisms is still limited. The presence of population-level lateralizations and cross-species similarities makes it likely that neuronal asymmetries have developed under phylogenetic pressure and, therefore, have a genetic basis [7,8,39,40]. However, human and animal research currently differ in the approaches and methods used in investigating the mechanisms guiding the development of a functional lateralized brain and, therefore, there has only been limited integration of knowledge between research approaches [41]. In some animal models, the genetically controlled events that drive the development of neuronal asymmetries have been studied in detail. In the nervous system of the nematode Caenorhabditis elegans, for example, there are pairs of chemoperceptive neurons, which are characterized by molecular left–right differences and different connectivity patterns that are related to their differential functional embedding [42,43]. Molecular genetic studies identified a complex regulation network comprising transcription factors, microRNAs, chromatin regulators and intercellular signals, which determine the asymmetric features of these specific neurons [44]. A second well-studied example is the epithalamus of vertebrate brains, which connects limbic regions of the forebrain with hindbrain motor circuits and which is characterized by evolutionarily conserved asymmetries within the pineal complex and the adjacent habenular nuclei. The molecular pathways that control these asymmetries have mainly been elucidated in studies with larvae of the zebrafish. Here, too, it is a chain of gene expression cascades that underlie the development of lateralization in this area [42,45,46]. Other genes are persistently expressed asymmetrically within the adult forebrain of zebrafish [47].
In human research, however, the first popular models, such as the right shift [48] or dextral chance [49] theory, suggested a single gene origin for human brain lateralization and attributed the left-hemispheric dominance for language processing and hand use to a common genetic factor. Their predictions fit data on the prevalence of handedness and language lateralization, but they did not explain the nature, as well as the action, of such a factor. However, recent meta-analysis studies have shown that the associations between language lateralization and motor asymmetries are much weaker than previously assumed [50]. Currently, research concentrates on the identification of genes that regulate functional and structural lateralization using large-scale heritability and genome-wide association (GWAS), or single nucleotide (SNP) variation studies to find associations between gene variants and phenotypic lateralizations. These studies have reported an increasing number of genes and their variants related to lateralization pattern. One recent study even identified multifaceted gene networks associated with different aspects of anatomical brain asymmetries [51]. It has also been suggested that the impact of single genes is small and functional lateralizations are polygenic traits [38,52,53,54,55,56]. A recent study, for instance, detected 41 gene loci associated with left-handedness and seven associated with ambidexterity [52]. This also suggests that different manifestations of a trait can be controlled by different types of genes, which are either relevant during different phases of development or which influence discrete differentiation processes of the underlying neural networks.
In general, additive genetic effects account for less than one quarter of the variance in human handedness data, while nonshared environmental factors explain the remaining variance [52,57]. This is not surprising since neuronal systems always differentiate in close interactions with environmental experiences and genes alone do not explain the functional organization of neuronal systems [58]. This implies that the emergence of a functional lateralization pattern can only be understood by elucidating how genes and the environment interact to shape the functional organization of a lateralized brain [2,3,14,59,60,61,62,63]. It must also be considered how noncoding microRNA [64], or epigenetic mechanisms, which affect gene activity and expression by modifying DNA accessibility or chromatin structure, mediate long-term effects of gene–environment interactions [37,63].
Research has reported a potpourri of environmental factors influencing lateralization patterns in humans, including sex hormones [65], stress experience [66], sensory input, learning, birthweight, location and season of birth, breast feeding and cultural constraints [32,53]. These influences underline, on the one hand, the general role of environmental factors, while on the other hand, the lack of specificity of some factors suggests that deviations from genetically controlled patterns simply reflect random stochastic asymmetry [67].
1.3. Structural Foundations of Functional Asymmetries
Since neuronal functions are based on the organization of specific neuronal networks or cells, it is necessary to clarify how exactly structural and functional asymmetries are related. An increasing number studies have reported structure–function associations, but have also provided an inconsistent pattern. However, it is important to differentiate that left–right differences in the structure of neural circuits can be realized on different organizational levels, from the cellular to the macroscopic level.
An obvious global shape asymmetry of the human brain is the so called “cerebral torque”, which refers to a counter-clockwise twist of the whole brain along the anterior–posterior axis [68,69]. At macroscopic level, the left hemisphere has a thicker cortex but a smaller surface area relative to the right hemisphere [70]. Region-specific left–right differences are reported in size and shape [70,71,72] and connectivity [73,74], as well the cellular and molecular organization [75,76]. Similar cortical asymmetries are also present in chimpanzees [77,78,79]. The left-hemispheric dominance of language processing is related to left–right differences in the microcircuitry of cortical columns in the posterior part of the superior temporal gyrus [80]. Moreover, there are function-related asymmetries in the hippocampus and subcortical structures in humans [71] and other mammals [81]. Handedness for instance is related to asymmetries within the nigrostriatal dopaminergic system in humans [82] and rodents [83,84,85].
Cortical left–right differences emerge early during development in humans [86,87], but also in nonhuman primates [88]. The cortical torque can be detected by the second trimester of gestation [68,89], while asymmetry of perisylvian language-related cortical regions appears during the third trimester [90,91,92]. Motor asymmetries can be observed even earlier. Human fetuses tend to make more movements with their right arms and preferentially suck the right thumb from the 12th gestational week onwards [93]. These motor asymmetries are related to postnatal handedness [94]. In relation to this behavioral lateralization, the fetal spinal cord segments innervating hands and arms display asymmetries in gene expression and DNA methylation at the end of the first trimester [95].
In sum, average left–right differences of global brain anatomy, which emerge early during development, suggest a developmental program that is genetically determined [96]. However, when analyzing specific cognitive functions, gene–structure interrelations are less detectable. Twin studies, for instance, indicate that pre- and postnatal events can affect asymmetry during development of the planum temporal [97,98]. Accordingly, a recent large-scale study did not find significant associations between cortical asymmetries and language lateralization [99]. There is also no significant relation between cortical asymmetries and handedness [70]. The lack of correlations may not come as a surprise since the macroscopic cortical features do not necessarily represent the internal microscopic organization. It is conceivable that functional asymmetries only emerge on the cellular, synaptic or neurophysiological level. This means that it is necessary to understand how neuronal asymmetries arise at precisely this cellular level. To this end, findings from developmental neurosciences have to be integrated into models of asymmetry formation. Experiments with animal models have shown that activity-dependent processes triggered by internal or external signals are decisive for the functional maturation of neural networks [100,101,102,103]. In the following, I will, therefore, first summarize what is known about the role of genetic factors for asymmetry formation during different developmental phases. I then illustrate the possible effects of environmental factors as suggested by the light-dependent development of visual asymmetries in birds.
2. Potential Roles of Genetic Factors for Asymmetry Formation
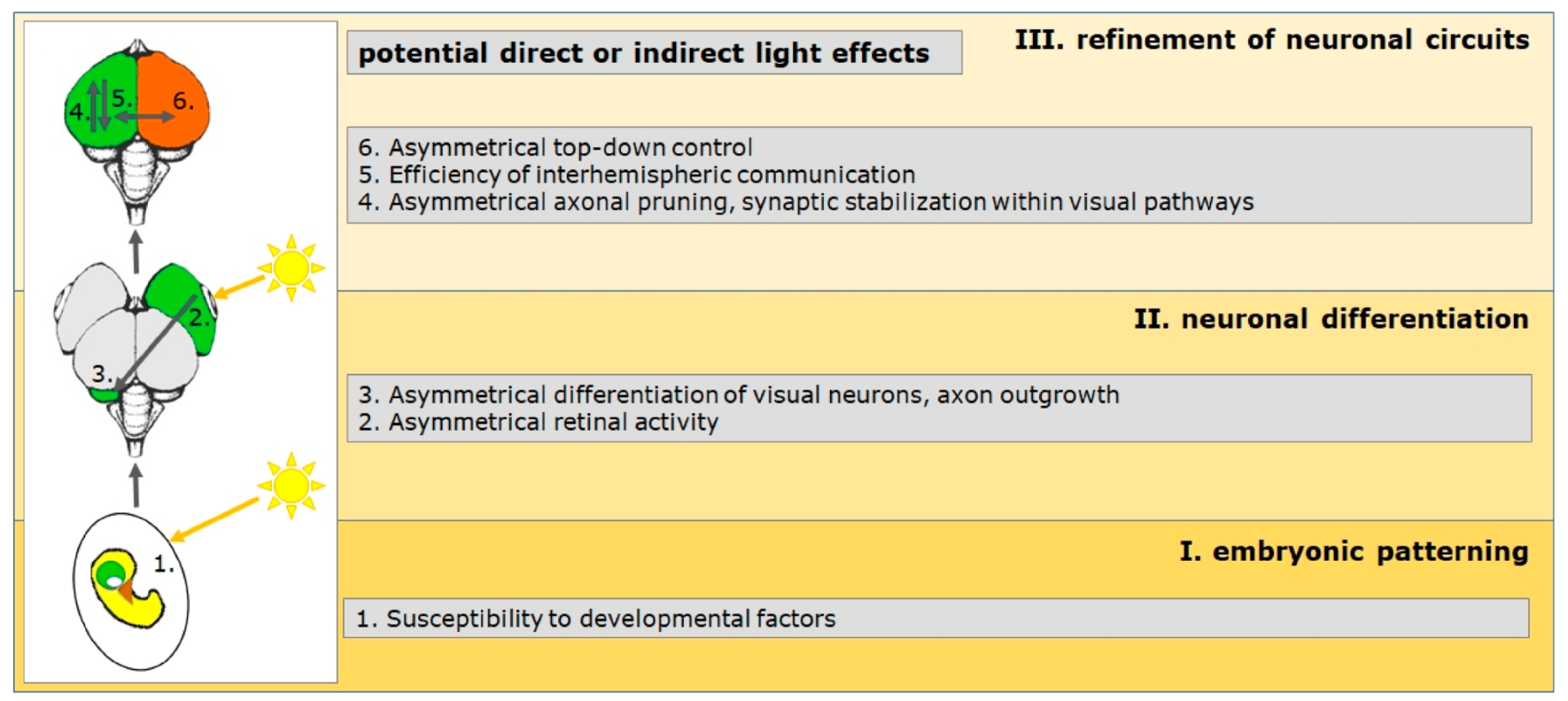
The relative importance of genes and the environment depends on the species examined, the specific neuronal function and their developmental trajectories, as well as the level of analysis [61]. This means that we have to differentiate the action of gene–environment interactions depending on the development phase. The development of the nervous system can be roughly divided into three phases, during which the degree of hemispheric specialization increases (Figure 1). The first phase comprises the earliest embryological steps, in which the axes of the body plan are determined. The second phase includes the differentiation of neural systems and networks, while processes mediating the refinement of neural connections dominate the third phase. During these phases, different cellular processes dominate development and genes can influence the action of epigenetic factors in different ways, which affect the developing organism (Figure 1):
- Specific genes can account for left–right differences in the amount of neuronal substrate. An asymmetrical number of neuronal and/or glial precursor cells can result in gross morphological asymmetries and can be related to differences in cellular identity or cell-type-specific proteins.
- Specific genes can control left–right differences in developmental dynamics. Differences in the maturation of left- or right-hemispheric neurons or networks could lead to asymmetries in the susceptibility to epigenetic factors like hormones, sensory input or motor activity.
- Specific genes can regulate asymmetrical morphogenetic events leading to asymmetric body positions or craniofacial asymmetries, which bias sensory experiences.
- Specific genes can control asymmetrical differentiation of neuronal elements like growth or arborization of axons and dendrites or development of synapses.
- Specific genes involved in synaptogenesis or signal transmission can lead to left–right differences in the degree of neuronal plasticity.
In the meantime, a number of genes have been discovered that mediate at least one of these actions during asymmetry formation:
2.1. Embryonic Patterning
Asymmetry formation within neuronal systems starts with breaking the symmetry of the body plan during early embryogenesis in all bilaterian animals, when the primary axes and tissue layers form. Complex cascades of genetic and epigenetic interactions lead to an asymmetrical placement of internal organs, but also induce asymmetries of paired organs like the lungs or the nervous system [104,105,106,107,108,109]. Determination of the left–right body axis is coordinated by a midline structure called the node. In several species, including humans, symmetry is broken by the rotation of motile cilia, which generate a directed flow that acts as a signal for the asymmetrical expression of a gene cascade, the Nodal signaling pathway. This pathway is remarkably conserved within bilaterian evolution [10,106,108].
This implies that asymmetry formation of body and brain starts with the action of cilia and, therefore, genes controlling generation and motility of cilia could play an early role in the development of neuronal asymmetries [38,110]. Some studies have actually provided evidence for the involvement of cilial genes for handedness—however, only in specific humans populations [38,54,111].
A second critical mechanism during this early phase is the lateralized action of the Nodal pathway. One key player in this signaling cascade is PCSK6, which cleaves the Nodal protoprotein into its biologically active form [10,110,112]. PCSK6 polymorphism has been associated with human handedness [38,113], but also with structural asymmetries in temporal cortical areas, indicating a potential role of PCSK6 not only for motoric but also language networks [114].
However, when symmetry breaking processes of visceral and neuronal structures share the same developmental route, one should assume that individuals with reversed visceral organization also display reversed brain asymmetries. A test case involves individuals with situs inversus, where the visceral organs are organized as a mirror image of the default organ position. Situs inversus can occur in, but does not depend on, ciliary dyskinesia [115]. While the typical gross morphological asymmetry of the human brain–cerebral torque is actually reversed in situs inversus, functional and cortical lateralizations are not [115,116,117,118,119], although atypical functional segregation can be more frequent in participants with visceral reversal [115,120]. Similarly, in less complex animals, such as the nematode C. elegans, motor lateralization is independent from left–right body asymmetry [121] and zebrafish with situs inversus develop reversed lateralization of some but not all structural and behavioral lateralizations [122]. This suggests that early embryonic patterning processes regulate, to some degree, the establishment of basic brain asymmetries, but lateralization of specific functional modules are presumably shaped by specific cellular mechanisms later during development [119,123].
2.2. Regionalization of Neuronal Substrate
When the neuronal anlage starts to differentiate region-specific differences, genes playing a role in symmetry breaking of the embryo are also involved in the generation of specific brain asymmetries. The best known example is the Nodal pathway, whereby asymmetrical left-sided Nodal signaling within the developing dorsal diencephalon is required for determining the direction of epithalamic asymmetries [42,45,112,124]. It is conceivable that laterality signals result in asymmetrical expression of neuron-type-specific gene batteries, which are responsible for cell-type-specific structural and functional properties [125].
2.3. Differential Developmental Dynamics
One consequence of the early left–right patterning is that the left and right hemispheres develop at different speeds. In human embryos, the right hemisphere tends to develop a little earlier than the left one [86] and the lateralized gradient of brain development might contribute to the development of the cerebral torque [69]. Differences in developmental speed of cortical subareas are indicated by specific lateralized gene expression profiles from the fifth week postconception onwards [126]. The early appearance of asymmetrical arm movements in human fetuses can be explained by left–right differences in the differentiation of spinal neurons, since the cortex and spinal cord are not connected at this age [127,128]. As a result of the asymmetrical developmental gradients of the two hemispheres, it is possible that a nongenetic factor, which acts on the developing organism at a certain point in time, differentially influences left- and right-hemispherical neuronal structures. There is, for instance, some evidence that the right hemisphere of human fetuses is generally less subject to external influences than the left one [86].
2.4. Differentiation of Hemisphere-Specific Neuronal Elements
When the nervous tissue starts to differentiate region-specific neurons and connections, specific genes regulate proliferation, migration and growth of axonal and/or dendritic fibers. Therefore, asymmetrical expression of these genes can account for the asymmetrical differentiation of specific brain regions.
Sun et al. [129,130] identified a couple of genes in perisylvian regions of the human cortex, which are asymmetrically expressed at the end of the first trimester and, therefore, before a neuroanatomical asymmetry of this area can be detected [90,91]. Intriguingly, most of these asymmetrically expressed genes function in signal transduction and gene expression regulation [129,130].
One of these genes is the transcription factor LMO4, which is consistently more highly expressed in the right perisylvian cortex of 12–16-week human fetuses and, hence, during a period of high proliferation and migration rate [129,130]. LMO4 displays higher expression level also in the right forebrain of zebrafish [47], while in the mouse cortex, LMO4 expression is not constantly lateralized to one side [129,130]. Expression of LMO4 is confined to postmitotic neurons [131] and regulates key aspects of neuronal differentiation, radial migration of newborn nerve cells and acquisition of neuronal identities [132,133].
Another example is the transcription factor forkhead box P2 gene FOXP2, which is involved in neural development and, in particular, in regulating neurogenesis of the embryonal cortex. It is expressed in distinct brain areas from gestational week six onwards and is related to speech development [134]. Intriguingly, FOXP2 polymorphism is associated with the interindividual variability in hemispheric asymmetries for speech perception [135].
2.5. Ontogenetic Plasticity
After the establishment of the basic brain organization, neuronal networks typically sharpen their functional efficiency. Growth, stabilization or reduction of synaptic contacts or cell death occur in an activity-dependent manner and are triggered by sensory experience [100,136]. This critical period is likely to amplify expression of genes and proteins that mediate synaptic plasticity. Accordingly, genes that are involved in regulating ontogenetic plasticity can affect the asymmetrical development during specific sensitive phases. Asymmetrical expression of these genes can result in a differential sensitivity of left- and right-hemispheric circuits towards stimulation. Karlebach and Francks [137], for instance, identified several asymmetrically expressed genes in the human cortex that are likely to fine-tune electrophysiological and neurotransmission properties of cortical circuits during different phases of development. Additionally, in the rat hippocampus, a dynamic pattern of asymmetrically expressed genes has been identified during the first postnatal weeks, with a large percentage of genes being associated with synaptic function [138]. One example could be the transmembrane molecule LRRTM1 (leucine-rich repeat transmembrane neuronal 1). It interacts at synapses with the extracellular matrix as a regulator of neuronal plasticity [139]. Gene variations have been associated with handedness [53,140,141].
Crucial mediators of ontogenetic plasticity are neurotrophic factors like BDNF (brain-derived neurotrophic factor), which mediates activity-dependent synaptic stabilization, axo-dendritic growth, arborization and cell survival [142,143]. It is, therefore, intriguing that BDNF is asymmetrically expressed in the hippocampus of rats, specifically during the first two weeks after birth when neurogenesis rate is high [144]. BDNF might also mediate stress effects in the brain and could, therefore, regulate the well-known action of stress hormones onto brain lateralization [145].
To sum it up, neuronal development is controlled at very different levels of differentiation by genes that are either asymmetrically expressed or whose variants are associated with specific phenotypes. The same function (e.g., handedness) can, therefore, be regulated during different developmental phases by different types of genes. Asymmetrical expression of single genes can be confined to specific developmental phases, while other genes are lateralized up until adulthood. At all levels, nongenetic factors can modulate genetic effect and thereby change the direction and/or degree of lateralization. However, little is yet understood about the neuronal processes through which environmental factors can influence the differentiation of the complex functional organization of lateralized brains. One of the few models in which the influence of a specific environmental factor has been examined in more detail is the visual system of birds. Research on chicks and pigeons has delineated a chain of events that begins with asymmetrical photic stimulation of the embryo in the egg and ends in a lateralized organization of visual processing and cognition [1,2,3,14,40,59,60,61,146,147]. This model suggests critical steps for the formation of asymmetries that can serve as a blueprint for a better understanding of the ontogenesis of brain asymmetries in general. These developmental steps are summarized below (Figure 2) and are complemented by findings in other species, especially in humans.
3. The Avian Visual System as a Model for Ontogenetic Plasticity
The visual system of chicks and pigeons is lateralized with a pattern that is similar to the lateralization of the human brain. The left hemisphere dominates the discrimination of small optic details, rule learning, categorization and visuomotor control [59,60,61,147,148,149]. The right hemisphere on the contrary, is in charge of spatial attention [150] and aspects of social cognition [23]. These hemispherical specializations can be identified very easily by temporarily occluding one eye with an opaque cap. Since the optic nerves cross virtually completely in birds, information from the left eye is primarily directed to the right hemisphere and vice versa. A comparison of monocular and binocular testing, therefore, enables the investigation of hemispherical differences in performances or analysis strategies. Behavioral asymmetries are accompanied by anatomical left–right differences within the ascending visual pathways. In both pigeons and chicks, for example, differences in the projection strength between the two hemispheres can be observed. Major aspects of these asymmetries develop in response to asymmetrical visual stimulation during development. Therefore, light deprivation before and after hatching prevents or modifies visual lateralizations. The comparison of structural and behavioral lateralizations of light-exposed or light-deprived birds makes it possible to unravel critical neuronal processes that mediate light-dependent development (Figure 2) [1,2,3,14,40,59,60,61,146,147,151,152].
3.1. Mechanisms during Embryonic Patterning (Phase I)
As in all vertebrates, asymmetry formation in birds starts during embryonic body patterning [153,154], whereby symmetry breaking is independent from motile cilia [105,106]. At this point of development, light cannot directly affect visual lateralization patterns but there are at least three routes serving as starting points for the induction of asymmetries in the visual system:
- Differences in left–right identity presumably determine asymmetries in the developing nervous system, which result in functional lateralizations when no other factors modify these predetermined ones. Early asymmetrical differentiation is indicated by a rightward torsion of the neuronal tube. Some evidence suggests that bending is caused by differential growth of the left and right neuronal tube side but physical mechanisms in relation to asymmetrical heart bending might also play a role [154]. This bending also occurs in mammalian embryos [155] and could contribute to the emergence of the cortical torque in the human brain. Presumably dependent on these early asymmetrical developmental processes, some visual lateralization develops independent from asymmetrical light stimulation. In chicks, visual choice to approach a social partner [156,157], uni-hemispheric sleep [158] or structural asymmetries of forebrain areas [159,160,161,162] are present in birds that are not exposed to biased visual stimulation. Interocular transfer of conditioned information [163] or lateralized visuospatial attention [164] emerge without embryonic light stimulation in pigeons but, interestingly, not in chicks [165,166]. It is currently unknown which genetic factors and which neuronal processes control the emergence of these visual asymmetries. However, endogenous asymmetries can be modulated by later visual experience [159,163,164,167].
- As a consequence of the primarily Nodal-dependent left–right determination of the body, morphogenetic processes lead to bending of the head region, which turns to the left so that the beak points to the right and the right side of the head is facing the egg [168]. Due to the fact that the size of the embryo during the last embryonic phase does not allow free head rotations anymore, the left eye arrests on the body while the right eye is close to the egg shell and can be stimulated by light shining through the egg shell (Figure 2). This biased environmental stimulation triggers the second step in asymmetry formation, inducing asymmetrical differentiation processes, which involve neuronal mechanisms well known for ontogenetic plasticity [59,60,152] (see phase II). Such a rightward torsion occurs in all amniotes [168] including human embryos, which display a right-turn of their head during the last gestational phase from week 38 onwards [169]. During this time, human fetuses are already responsive to sensory stimulation. They are able to memorize auditory stimuli from the external world by the last trimester of pregnancy, with a particular sensitivity to melody contour in both music and language [170,171]. Differential auditory input to the left and right ear because of postural asymmetries, therefore, might affect the development of language lateralizations [172,173,174].
- Although visual systems are not developed, there is some evidence that during this phase, light stimulation already affects the establishment of some aspects of lateralization in both chickens and zebrafish [166,175,176,177] (Figure 2). Transduction mechanisms mediating these light effects are unknown but might include epigenetic mechanisms [166,177]. It is also possible that some genes unfold their actions only after photostimulation [178].
3.2. Mechanisms during Neuronal Differentiation (Phase II)
It is well known that the differentiation of visual networks is critically influenced by visual stimulation (e.g., [100]], and it is therefore not surprising that unbalanced light stimulation differentially affects left- and right-hemispheric developmental processes during species-specific sensitive phases [61,179]. Some behaviors and anatomical asymmetries only develop after embryonic light stimulation [180,181,182] and can be reversed by altered visual experience before (chicks, [183]) or after (pigeons [184,185]) hatching. In chicks, the outgrowth of visual fibers is influenced by light stimulation, resulting in a transiently stronger innervation of the right visual forebrain. Thereby, the action of light is modified by corticosterone, testosterone and estradiol [1,14,146,147,149,186]. The modulatory action of steroid hormones is in line with the often described sex- and stress-effects on human and nonhuman lateralization patterns [66,187,188]. In pigeons, left–right differences in cell size and projection strength differentiate in response to asymmetric photic stimulation [180,182,184,185,189]. Posthatch experimental manipulations have shown that starting with asymmetrical retinal activity [190], asymmetrical differentiation within the ascending visual system is mediated partly by BDNF-dependent processes [191,192].
The avian models exemplify how an environmental factor shapes the generation of neuronal asymmetries by modifying specific bottom-up systems. In a similar way, left–right differences in spectrotemporal selectivity of neurons in the auditory cortex of mice develop depending on hearing experience, which is related to the left-hemispheric dominance for the analysis of vocalization features [193]. In humans, visual experience can affect handedness [173], head turning preference [194] or lateralized face-processing competence [195,196].
However, an asymmetrical sensory trigger, such as light, not only enhances differentiation of the stronger stimulated hemisphere but also modifies the balance of left- and right-hemispheric development. A detailed analysis of light- and dark-incubated pigeons, for instance, revealed that light induces a left-hemispheric increase in visuoperceptual skills but simultaneously decreases visuomotor speed within the right hemisphere [182]. At the neuroanatomical level, embryonic light stimulation does not increase the bilateral innervation of the more strongly stimulated left brain side, but rather decreases input to the right side [180].
Presumably, interdependent left- and right-hemispheric developmental processes also play a role in the experience-dependent specialization of the human cortex, as indicated by the distribution of hemispheric language and face recognition processing. While the visual word form area in the left hemisphere becomes specialized while learning to read, the right hemisphere develops face recognition dominance. This suggests that the hemispheric organization of face recognition and of word recognition does not develop independently, and that word lateralization may precede and drive later face lateralization [196,197].
3.3. Consolidation of Functional Asymmetries (Phase III)
The ontogeny of visual asymmetries in birds is profoundly triggered within the developing ascending visual pathways but cognitive asymmetries emerge only at a higher (forebrain) processing level [60,148]. This means that asymmetries, which are induced within bottom-up systems, have to be transferred onto higher brain structures. At this level, they might interact with inherent or light-independent asymmetries (see above) and thereby sculpt and stabilize the final functional organization of the visual brain. In the pigeon, these processes mainly take place after hatching, when light input is normally symmetrical. During this phase, lateralization can still be modified by manipulating the visual experience [59,60,152,184,190]. It is likely that top-down as well as commissural mechanisms play a critical role in these stabilization processes [59,60,185,189,198,199]. As a consequence, relevant top-down and/or commissural systems develop their own asymmetrical properties for controlling asymmetrical decision-making and behaviors, but also for determining the degree of interhemispheric crosstalk. For example, left-hemispheric dominance for conflict choices is related to the asymmetrical action of top-down projections from the forebrain [198]. Light-dependent efficiency of interhemispheric integration has been shown in chicks, where only light-stimulated individuals can efficiently allocate food searching to the left and predator vigilance to the right hemisphere [200]. Also, only light-exposed chicks can use object (left-hemisphere)- as well as position (right-hemispheric)-dependent cues in food searching tasks [201,202]. A study with pigeons showed that only light-stimulated birds integrate hemispheric-specific knowledge for solving a task that cannot be correctly answered with information of one hemisphere alone [199]. Relevance of interhemispheric mechanisms for the generation and modulation of hemispheric-specific functions is in line with studies exploring the role of the corpus callosum for brain lateralizations [203,204]. The avian model suggests that top-down and commissural systems unfold their effects mainly at the end of asymmetry formation and modulate the interaction of more or less strongly lateralized neuronal networks in the left and right hemispheres [148]. To this regard, these processes shape the final functional organization of lateralized cognitive modules.
4. Conclusions
Studies on the genetic basis and/or environmental influences on the formation of asymmetries in humans and other animals have shown that the development of a lateralized functional architecture of the brain is to be understood as an example of ontogenetic plasticity. Genes and environmental factors play different but intertwined and complementary roles that can be specific to certain processing modules. The final functional lateralization pattern is then the result of hierarchical processes that build on one another. Genetically controlled early embryonic developmental steps set the framework for hemispherical differences and can be indicated by gross morphological asymmetries in volume and/ or shape of gray and white matter. Epigenetic processes lead to increasing hemispherical specialization and control dynamics of interhemispheric communication. This means that no factor alone can explain the variance of lateralization patterns in a population; it is the sum of individual experiences, which shape individual brain lateralization. It is possible to identify general roles of single genes or environmental factors, but only their interplay within a specific environment determines the functional outcome. Consequently, single factors can only explain limited variance in the lateralization pattern within a population.
This flexibility enables fluctuating lateralization patterns within a population depending on the ecological requirements. Recent field studies showed, for instance, that factors such as predator pressure, environmental pollutants or seasonal conditions can modify brain asymmetries [205,206,207]. Humans have cultural constraints affecting, for example, the prevalence of left-handedness [208]. It is conceivable that the specific ecological or social conditions account for population-level lateralization in humans, which is absent in other animals species [208]. Ontogenetic plasticity, however, can be a general mechanism that enhances the evolutionary benefit of brain asymmetries [61,208].
Funding
This work was funded by the DFG (grant number MA4485/2-2).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The author declares no conflict of interest.
References
- Rogers, L.J. Factors Influencing Development of Lateralization. Cortex 2006, 42, 107–109. [Google Scholar] [CrossRef]
- Güntürkün, O.; Ocklenburg, S. Ontogenesis of Lateralization. Neuron 2017, 94, 249–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Güntürkün, O.; Ströckens, F.; Ocklenburg, S. Brain Lateralization: A Comparative Perspective. Physiol. Rev. 2020, 100, 1019–1063. [Google Scholar] [CrossRef]
- Frasnelli, E.; Vallortigara, G.; Rogers, L.J. Left–right asymmetries of behaviour and nervous system in invertebrates. Neurosci. Biobehav. Rev. 2012, 36, 1273–1291. [Google Scholar] [CrossRef]
- Niven, J.E.; Frasnelli, E. Insights into the evolution of lateralization from the insects. Prog. Brain Res. 2018, 238, 3–31. [Google Scholar] [CrossRef]
- Rogers, L.J.; Vallortigara, G.; Andrew, R.J. Divided Brains: The Biology and Behaviour of Brain Asymmetries; University Press: Cambridge, UK, 2013. [Google Scholar]
- Vallortigara, G.; Rogers, L.J. Survival with an asymmetrical brain: Advantages and disadvantages of cerebral lateralization. Behav. Brain Sci. 2005, 28, 575–589. [Google Scholar] [CrossRef]
- Vallortigara, G.; Rogers, L.J. A function for the bicameral mind. Cortex 2020, 124, 274–285. [Google Scholar] [CrossRef]
- Vallortigara, G.; Versace, E. Laterality at the neural, cognitive, and behavioral levels. In APA Handbook of Comparative Psychology: Basic Concepts, Methods, Neural Substrate, and Behavior; American Psychological Association: Washington, DC, USA, 2017; Volume 1, pp. 557–577. [Google Scholar] [CrossRef]
- Heger, P.; Zheng, W.; Rottmann, A.; Panfilio, K.A.; Wiehe, T. The genetic factors of bilaterian evolution. eLife 2020, 9. [Google Scholar] [CrossRef]
- Manns, M. Hemispheric Specialization. In Encyclopedia of Animal Cognition and Behavior; Vonk, J., Shackelford, T., Eds.; Springer International Publishing: Cham, Germany, 2019; pp. 1–10. [Google Scholar]
- Manns, M.; El Basbasse, Y.; Freund, N.; Ocklenburg, S. Paw preferences in mice and rats: Meta-analysis. Neurosci. Biobehav. Rev. 2021, 127, 593–606. [Google Scholar] [CrossRef]
- Frasnelli, E.; Vallortigara, G. Individual-Level and Population-Level Lateralization: Two Sides of the Same Coin. Symmetry 2018, 10, 739. [Google Scholar] [CrossRef]
- Rogers, L.J. Asymmetry of brain and behavior in animals: Its development, function, and human relevance. Genesis 2014, 52, 555–571. [Google Scholar] [CrossRef] [PubMed]
- Lippolis, G.; Joss, J.; Rogers, L. Australian Lungfish (Neoceratodus forsteri): A Missing Link in the Evolution of Complementary Side Biases for Predator Avoidance and Prey Capture. Brain Behav. Evol. 2009, 73, 295–303. [Google Scholar] [CrossRef] [PubMed]
- MacNeilage, P.F.; Rogers, L.J.; Vallortigara, G. Origins of the Left & Right Brain. Sci. Am. 2009, 301, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Vallortigara, G. Comparative Neuropsychology of the Dual Brain: A Stroll through Animals’ Left and Right Perceptual Worlds. Brain Lang. 2000, 73, 189–219. [Google Scholar] [CrossRef]
- Rogers, L.J.; Andrew, R. Comparative Vertebrate Lateralization; Cambridge University Press: Cambridge, UK, 2002. [Google Scholar]
- Salva, O.R.; Regolin, L.; Mascalzoni, E.; Vallortigara, G. Cerebral and Behavioural Asymmetries in Animal Social Recognition. Comp. Cogn. Behav. Rev. 2012, 7, 110–138. [Google Scholar] [CrossRef] [Green Version]
- Gainotti, G. Laterality effects in normal subjects’ recognition of familiar faces, voices and names. Perceptual and representational components. Neuropsychologia 2013, 51, 1151–1160. [Google Scholar] [CrossRef]
- Kendrick, K.M. Brain asymmetries for face recognition and emotion control in sheep. Cortex 2006, 42, 96–98. [Google Scholar] [CrossRef]
- Versace, E.; Morgante, M.; Pulina, G.; Vallortigara, G. Behavioural lateralization in sheep (Ovis aries). Behav. Brain Res. 2007, 184, 72–80. [Google Scholar] [CrossRef]
- Daisley, J.N.; Mascalzoni, E.; Salva, O.R.; Rugani, R.; Regolin, L. Lateralization of social cognition in the domestic chicken (Gallus gallus). Philos. Trans. R. Soc. B Biol. Sci. 2008, 364, 965–981. [Google Scholar] [CrossRef] [Green Version]
- Ocklenburg, S.; Beste, C.; Arning, L.; Peterburs, J.; Güntürkün, O. The ontogenesis of language lateralization and its relation to handedness. Neurosci. Biobehav. Rev. 2014, 43, 191–198. [Google Scholar] [CrossRef]
- Levy, R.B.; Marquarding, T.; Reid, A.P.; Pun, C.M.; Renier, N.; Oviedo, H.V. Circuit asymmetries underlie functional lateralization in the mouse auditory cortex. Nat. Commun. 2019, 10, 2783. [Google Scholar] [CrossRef] [Green Version]
- Ocklenburg, S.; Ströckens, F.; Güntürkün, O. Lateralisation of conspecific vocalisation in non-human vertebrates. Laterality 2013, 18, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Bell, A.T.A.; Niven, J.E. Individual-level, context-dependent handedness in the desert locust. Curr. Biol. 2014, 24, R382–R383. [Google Scholar] [CrossRef] [Green Version]
- Bell, A.T.A.; Niven, J.E. Strength of forelimb lateralization predicts motor errors in an insect. Biol. Lett. 2016, 12, 20160547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Byrne, R.A.; Kuba, M.J.; Meisel, D.V.; Griebel, U.; Mather, J.A. Does Octopus vulgaris have preferred arms? J. Comp. Psychol. 2006, 120, 198–204. [Google Scholar] [CrossRef] [PubMed]
- Ströckens, F.; Güntürkün, O.; Ocklenburg, S. Limb preferences in non-human vertebrates. Laterality 2013, 18, 536–575. [Google Scholar] [CrossRef] [PubMed]
- Versace, E.; Vallortigara, G. Forelimb preferences in human beings and other species: Multiple models for testing hypotheses on lateralization. Front. Psychol. 2015, 6, 233. [Google Scholar] [CrossRef] [Green Version]
- Papadatou-Pastou, M.; Ntolka, E.; Schmitz, J.; Martin, M.; Munafò, M.R.; Ocklenburg, S.; Paracchini, S. Human handedness: A meta-analysis. Psychol. Bull. 2020, 146, 481–524. [Google Scholar] [CrossRef]
- Packheiser, J.; Schmitz, J.; Berretz, G.; Carey, D.P.; Paracchini, S.; Papadatou-Pastou, M.; Ocklenburg, S. Four meta-analyses across 164 studies on atypical footedness prevalence and its relation to handedness. Sci. Rep. 2020, 10, 1–21. [Google Scholar] [CrossRef]
- Packheiser, J.; Schmitz, J.; Berretz, G.; Papadatou-Pastou, M.; Ocklenburg, S. Handedness and sex effects on lateral biases in human cradling: Three meta-analyses. Neurosci. Biobehav. Rev. 2019, 104, 30–42. [Google Scholar] [CrossRef]
- Grimshaw, G.M.; Ecarmel, D. An asymmetric inhibition model of hemispheric differences in emotional processing. Front. Psychol. 2014, 5, 489. [Google Scholar] [CrossRef]
- Ocklenburg, S.; Beste, C.; Güntürkün, O. Handedness: A neurogenetic shift of perspective. Neurosci. Biobehav. Rev. 2013, 37, 2788–2793. [Google Scholar] [CrossRef]
- Schmitz, J.; Metz, G.A.; Güntürkün, O.; Ocklenburg, S. Beyond the genome—Towards an epigenetic understanding of handedness ontogenesis. Prog. Neurobiol. 2017, 159, 69–89. [Google Scholar] [CrossRef] [PubMed]
- Brandler, W.M.; Morris, A.P.; Evans, D.M.; Scerri, T.S.; Kemp, J.P.; Timpson, N.J.; Pourcain, B.S.; Smith, G.D.; Ring, S.M.; Stein, J.; et al. Common Variants in Left/Right Asymmetry Genes and Pathways Are Associated with Relative Hand Skill. PLoS Genet. 2013, 9, e1003751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laland, K.N. Exploring gene–culture interactions: Insights from handedness, sexual selection and niche-construction case studies. Philos. Trans. R. Soc. B Biol. Sci. 2008, 363, 3577–3589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rogers, L.J.; Vallortigara, G. When and Why Did Brains Break Symmetry? Symmetry 2015, 7, 2181–2194. [Google Scholar] [CrossRef] [Green Version]
- Ocklenburg, S.; Berretz, G.; Packheiser, J.; Friedrich, P. Laterality 2020: Entering the next decade. Laterality 2021, 26, 265–297. [Google Scholar] [CrossRef]
- Concha, M.L.; Bianco, I.; Wilson, S. Encoding asymmetry within neural circuits. Nat. Rev. Neurosci. 2012, 13, 832–843. [Google Scholar] [CrossRef]
- Hobert, O.; Johnston, R.J.; Chang, S. Left–right asymmetry in the nervous system: The Caenorhabditis elegans model. Nat. Rev. Neurosci. 2002, 3, 629–640. [Google Scholar] [CrossRef]
- Hobert, O. Development of left/right asymmetry in the Caenorhabditis elegans nervous system: From zygote to postmitotic neuron. Genesis 2014, 52, 528–543. [Google Scholar] [CrossRef]
- Concha, M.L.; Signore, I.A.; Colombo, A. Mechanisms of directional asymmetry in the zebrafish epithalamus. Semin. Cell Dev. Biol. 2009, 20, 498–509. [Google Scholar] [CrossRef] [PubMed]
- Roberson, S.; Halpern, M.E. Development and connectivity of the habenular nuclei. Semin. Cell Dev. Biol. 2018, 78, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Messina, A.; Boiti, A.; Vallortigara, G. Asymmetric distribution of pallial-expressed genes in zebrafish (Danio rerio). Eur. J. Neurosci. 2021, 53, 362–375. [Google Scholar] [CrossRef] [PubMed]
- Annett, M. Handedness and Cerebral Dominance. J. Neuropsychiatry Clin. Neurosci. 1998, 10, 459–469. [Google Scholar] [CrossRef]
- McManus, I.C. Handedness, language dominance and aphasia: A genetic model. Psychol. Med. Monogr. Suppl. 1985, 8, 1–40. [Google Scholar] [CrossRef] [Green Version]
- Packheiser, J.; Schmitz, J.; Arning, L.; Beste, C.; Güntürkün, O.; Ocklenburg, S. A large-scale estimate on the relationship between language and motor lateralization. Sci. Rep. 2020, 10, 1–10. [Google Scholar] [CrossRef]
- Sha, Z.; Schijven, D.; Carrion-Castillo, A.; Joliot, M.; Mazoyer, B.; Fisher, S.E.; Crivello, F.; Francks, C. The genetic architecture of structural left–right asymmetry of the human brain. Nat. Hum. Behav. 2021, 1–14. [Google Scholar] [CrossRef]
- Cuellar-Partida, G.; Tung, J.Y.; Eriksson, N.; Albrecht, E.; Aliev, F.; Andreassen, O.A.; Barroso, I.; Beckmann, J.S.; Boks, M.P.; Boomsma, D.I.; et al. Genome-wide association study identifies 48 common genetic variants associated with handedness. Nat. Hum. Behav. 2021, 5, 59–70. [Google Scholar] [CrossRef]
- De Kovel, C.G.F.; Carrión-Castillo, A.; Francks, C. A large-scale population study of early life factors influencing left-handedness. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef] [Green Version]
- De Kovel, C.G.F.; Francks, C. The molecular genetics of hand preference revisited. Sci. Rep. 2019, 9, 1–9. [Google Scholar] [CrossRef] [Green Version]
- McManus, C. Half a century of handedness research: Myths, truths; fictions, facts; backwards, but mostly forwards. Brain Neurosci. Adv. 2019, 3, 2398212818820513. [Google Scholar] [CrossRef] [Green Version]
- McManus, I.C.; Davison, A.; Armour, J. Multilocus genetic models of handedness closely resemble single-locus models in explaining family data and are compatible with genome-wide association studies. Ann. N. Y. Acad. Sci. 2013, 1288, 48–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medland, S.E.; Duffy, D.L.; Wright, M.; Geffen, G.M.; Hay, D.A.; Levy, F.; Van-Beijsterveldt, C.E.; Willemsen, G.; Townsend, G.C.; White, V.; et al. Genetic influences on handedness: Data from 25,732 Australian and Dutch twin families. Neuropsychology 2009, 47, 330–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, W.J.; Fine, I. New insights into cortical development and plasticity: From molecules to behavior. Curr. Opin. Physiol. 2020, 16, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Güntürkün, O.; Manns, M. The Embryonic Development of Visual Asymmetry in the Pigeon. In The Two Halves of the Brain; Hugdahl, K., Westerhausen, R., Eds.; The MIT Press: Cambridge, MA, USA, 2010; pp. 121–142. [Google Scholar]
- Manns, M.; Güntürkün, O. Dual coding of visual asymmetries in the pigeon brain: The interaction of bottom-up and top-down systems. Exp. Brain Res. 2009, 199, 323–332. [Google Scholar] [CrossRef] [PubMed]
- Manns, M.; Ströckens, F. Functional and structural comparison of visual lateralization in birds-similar but still different. Front. Psychol. 2014, 5, 206. [Google Scholar] [CrossRef] [Green Version]
- Ocklenburg, S.; Güntürkün, O. Hemispheric Asymmetries: The Comparative View. Front. Psychol. 2012, 3, 5. [Google Scholar] [CrossRef] [Green Version]
- Schmitz, J.; Güntürkün, O.; Ocklenburg, S. Building an Asymmetrical Brain: The Molecular Perspective. Front. Psychol. 2019, 10, 982. [Google Scholar] [CrossRef] [Green Version]
- Miao, N.; Lai, X.; Zeng, Z.; Cai, W.; Chen, W.; Sun, T. Differential expression of microRNAs in the human fetal left and right cerebral cortex. Mol. Biol. Rep. 2020, 47, 6573–6586. [Google Scholar] [CrossRef]
- Richards, G.; Beking, T.; Kreukels, B.P.; Geuze, R.H.; Beaton, A.A.; Groothuis, T. An examination of the influence of prenatal sex hormones on handedness: Literature review and amniotic fluid data. Horm. Behav. 2021, 129, 104929. [Google Scholar] [CrossRef]
- Berretz, G.; Wolf, O.T.; Güntürkün, O.; Ocklenburg, S. Atypical lateralization in neurodevelopmental and psychiatric disorders: What is the role of stress? Cortex 2020, 125, 215–232. [Google Scholar] [CrossRef]
- McManus, C. Is any but a tiny fraction of handedness variance likely to be due to the external environment? Laterality 2021, 26, 310–314. [Google Scholar] [CrossRef]
- Kong, X.-Z.; Postema, M.; Schijven, D.; Castillo, A.C.; Pepe, A.; Crivello, F.; Joliot, M.; Mazoyer, B.; Fisher, S.E.; Francks, C. Large-Scale Phenomic and Genomic Analysis of Brain Asymmetrical Skew. Cereb. Cortex 2021, 31, 4151–4168. [Google Scholar] [CrossRef] [PubMed]
- Xiang, L.; Crow, T.; Roberts, N. Cerebral torque is human specific and unrelated to brain size. Brain Struct. Funct. 2019, 224, 1141–1150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, X.-Z.; Mathias, S.R.; Guadalupe, T.; Glahn, D.C.; Franke, B.; Crivello, F.; Tzourio-Mazoyer, N.; Fisher, S.E.; Thompson, P.M.; Francks, C.; et al. Mapping cortical brain asymmetry in 17,141 healthy individuals worldwide via the ENIGMA Consortium. Proc. Natl. Acad. Sci. USA 2018, 115, E5154–E5163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guadalupe, T.; Zwiers, M.P.; Teumer, A.; Wittfeld, K.; Vasquez, A.A.; Hoogman, M.; Hagoort, P.; Fernandez, G.; Buitelaar, J.; Hegenscheid, K.; et al. Measurement and genetics of human subcortical and hippocampal asymmetries in large datasets. Hum. Brain Mapp. 2014, 35, 3277–3289. [Google Scholar] [CrossRef] [Green Version]
- Maingault, S.; Tzourio-Mazoyer, N.; Mazoyer, B.; Crivello, F. Regional correlations between cortical thickness and surface area asymmetries: A surface-based morphometry study of 250 adults. Neuropsychology 2016, 93, 350–364. [Google Scholar] [CrossRef]
- Ocklenburg, S.; Friedrich, P.; Güntürkün, O.; Genç, E. Intrahemispheric white matter asymmetries: The missing link between brain structure and functional lateralization? Rev. Neurosci. 2016, 27, 465–480. [Google Scholar] [CrossRef]
- De Schotten, M.T.; Dell’Acqua, F.; Forkel, S.; Simmons, A.; Vergani, F.; Murphy, D.; Catani, M. A lateralized brain network for visuospatial attention. Nat. Neurosci. 2011, 14, 1245–1246. [Google Scholar] [CrossRef]
- Chance, S.A. The cortical microstructural basis of lateralized cognition: A review. Front. Psychol. 2014, 5, 820. [Google Scholar] [CrossRef] [Green Version]
- Schmitz, J.; Fraenz, C.; Schlüter, C.; Friedrich, P.; Jung, R.E.; Güntürkün, O.; Genç, E.; Ocklenburg, S. Hemispheric asymmetries in cortical gray matter microstructure identified by neurite orientation dispersion and density imaging. NeuroImage 2019, 189, 667–675. [Google Scholar] [CrossRef]
- Cheng, L.; Zhang, Y.; Li, G.; Wang, J.; Sherwood, C.; Gong, G.; Fan, L.; Jiang, T. Connectional asymmetry of the inferior parietal lobule shapes hemispheric specialization in humans, chimpanzees, and rhesus macaques. eLife 2021, 10, e67600. [Google Scholar] [CrossRef] [PubMed]
- Graïc, J.-M.; Peruffo, A.; Corain, L.; Centelleghe, C.; Granato, A.; Zanellato, E.; Cozzi, B. Asymmetry in the Cytoarchitecture of the Area 44 Homolog of the Brain of the Chimpanzee Pan troglodytes. Front. Neuroanat. 2020, 14, 55. [Google Scholar] [CrossRef] [PubMed]
- Spocter, M.A.; Sherwood, C.C.; Schapiro, S.J.; Hopkins, W.D. Reproducibility of leftward planum temporale asymmetries in two genetically isolated populations of chimpanzees Pan troglodytes. Proc. R. Soc. B Boil. Sci. 2020, 287, 20201320. [Google Scholar] [CrossRef] [PubMed]
- Galuske, R.A.W.; Schlote, W.; Bratzke, H.; Singer, W. Interhemispheric Asymmetries of the Modular Structure in Human Temporal Cortex. Science 2000, 289, 1946–1949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, G.; Yang, X.; Yuan, T.-F. Hippocampal Asymmetry: Differences in Structures and Functions. Neurochem. Res. 2013, 38, 453–460. [Google Scholar] [CrossRef]
- De la Fuente-Fernández, R.; Kishore, A.; Calne, D.B.; Ruth, T.J.; Stoessl, A. Nigrostriatal dopamine system and motor lateralization. Behav. Brain Res. 2000, 112, 63–68. [Google Scholar] [CrossRef]
- Budilin, S.Y.; Midzyanovskaya, I.S.; Shchegolevskii, N.V.; Ioffe, M.E.; Bazyan, A.S. Asymmetry in dopamine levels in the nucleus accumbens and motor preference in rats. Neurosci. Behav. Physiol. 2008, 38, 991–994. [Google Scholar] [CrossRef]
- Nielsen, D.M.; Visker, K.E.; Cunningham, M.J.; Keller, R.W.; Glick, S.D.; Carlson, J.N. Paw Preference, Rotation, and Dopamine Function in Collins HI and LO Mouse Strains. Physiol. Behav. 1997, 61, 525–535. [Google Scholar] [CrossRef]
- Schwarting, R.; Nagel, J.A.; Huston, J.P. Asymmetries of brain dopamine metabolism related to conditioned paw usage in the rat. Brain Res. 1987, 417, 75–84. [Google Scholar] [CrossRef]
- Bisiacchi, P.; Cainelli, E. Structural and functional brain asymmetries in the early phases of life: A scoping review. Brain Struct. Funct. 2021, 1–18, Online ahead of print. [Google Scholar] [CrossRef]
- Vasung, L.; Rollins, C.K.; Velasco-Annis, C.; Yun, H.J.; Zhang, J.; Warfield, S.K.; Feldman, H.A.; Gholipour, A.; Grant, P.E. Spatiotemporal Differences in the Regional Cortical Plate and Subplate Volume Growth during Fetal Development. Cereb. Cortex 2020, 30, 4438–4453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, J.; Wang, F.; Wu, Z.; Wang, L.; Zhang, C.; Shen, D.; Li, G. Mapping hemispheric asymmetries of the macaque cerebral cortex during early brain development. Hum. Brain Mapp. 2019, 41, 95–106. [Google Scholar] [CrossRef] [PubMed]
- Weinberger, D.R.; Luchins, D.J.; Morihisa, J.; Wyatt, R.J. Asymmetrical volumes of the right and left frontal and occipital regions of the human brain. Ann. Neurol. 1982, 11, 97–100. [Google Scholar] [CrossRef] [PubMed]
- Chi, J.G.; Dooling, E.C.; Gilles, F.H. Left-Right Asymmetries of the Temporal Speech Areas of the Human Fetus. Arch. Neurol. 1977, 34, 346–348. [Google Scholar] [CrossRef]
- Hering-Hanit, R.; Achiron, R.; Lipitz, S.; Achiron, A. Asymmetry of fetal cerebral hemispheres: In utero ultrasound study. Arch. Dis. Child.-Fetal Neonatal Ed. 2001, 85, 194–196. [Google Scholar] [CrossRef] [Green Version]
- Kasprian, G.; Langs, G.; Brugger, P.C.; Bittner, M.; Weber, M.; Arantes, M.; Prayer, D. The Prenatal Origin of Hemispheric Asymmetry: An In Utero Neuroimaging Study. Cereb. Cortex 2011, 21, 1076–1083. [Google Scholar] [CrossRef]
- Hepper, P.G. The developmental origins of laterality: Fetal handedness. Dev. Psychobiol. 2013, 55, 588–595. [Google Scholar] [CrossRef]
- Hepper, P.G.; Wells, D.L.; Lynch, C. Prenatal thumb sucking is related to postnatal handedness. Neuropsychologia 2005, 43, 313–315. [Google Scholar] [CrossRef]
- Ocklenburg, S.; Schmitz, J.; Moinfar, Z.; Moser, D.; Klose, R.; Lor, S.; Kunz, G.; Tegenthoff, M.; Faustmann, P.; Francks, C.; et al. Epigenetic regulation of lateralized fetal spinal gene expression underlies hemispheric asymmetries. eLife 2017, 6, e22784. [Google Scholar] [CrossRef]
- Francks, C. Exploring human brain lateralization with molecular genetics and genomics. Ann. N. Y. Acad. Sci. 2015, 1359, 1–13. [Google Scholar] [CrossRef]
- Eckert, M.A.; Leonard, C.M.; Molloy, E.A.; Blumenthal, J.; Zijdenbos, A.; Giedd, J.N. The Epigenesis of Planum Temporale Asymmetry in Twins. Cereb. Cortex 2002, 12, 749–755. [Google Scholar] [CrossRef] [Green Version]
- Steinmetz, H.; Herzog, A.; Schlaug, G.; Huang, Y.; Jäncke, L. Brain (A)Symmetry in Monozygotic Twins. Cereb. Cortex 1995, 5, 296–300. [Google Scholar] [CrossRef]
- Tzourio-Mazoyer, N.; Crivello, F.; Mazoyer, B. Is the planum temporale surface area a marker of hemispheric or regional language lateralization? Brain Struct. Funct. 2017, 223, 1–12. [Google Scholar] [CrossRef]
- Cohen-Cory, S. The Developing Synapse: Construction and Modulation of Synaptic Structures and Circuits. Science 2002, 298, 770–776. [Google Scholar] [CrossRef] [Green Version]
- Katz, L.C.; Shatz, C.J. Synaptic Activity and the Construction of Cortical Circuits. Science 1996, 274, 1133–1138. [Google Scholar] [CrossRef] [Green Version]
- Pratt, K.G.; Hiramoto, M.; Cline, H.T. An Evolutionarily Conserved Mechanism for Activity-Dependent Visual Circuit Development. Front. Neural Circuits 2016, 10, 79. [Google Scholar] [CrossRef] [Green Version]
- Sur, M.; Rubenstein, J.L.R. Patterning and Plasticity of the Cerebral Cortex. Science 2005, 310, 805–810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilmour, D.; Rembold, M.; Leptin, M. From morphogen to morphogenesis and back. Nat. Cell Biol. 2017, 541, 311–320. [Google Scholar] [CrossRef] [PubMed]
- Grimes, D.T. Making and breaking symmetry in development, growth and disease. Development 2019, 146, dev170985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grimes, D.T.; Burdine, R.D. Left–Right Patterning: Breaking Symmetry to Asymmetric Morphogenesis. Trends Genet. 2017, 33, 616–628. [Google Scholar] [CrossRef]
- Levin, M. Left–right asymmetry in embryonic development: A comprehensive review. Mech. Dev. 2005, 122, 3–25. [Google Scholar] [CrossRef]
- Nakamura, T.; Hamada, H. Left-right patterning: Conserved and divergent mechanisms. Development 2012, 139, 3257–3262. [Google Scholar] [CrossRef] [Green Version]
- Zinski, J.; Tajer, B.; Mullins, M.C. TGF-β Family Signaling in Early Vertebrate Development. Cold Spring Harb. Perspect. Biol. 2018, 10, a033274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brandler, W.M.; Paracchini, S. The genetic relationship between handedness and neurodevelopmental disorders. Trends Mol. Med. 2014, 20, 83–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmitz, J.; Fraenz, C.; Schlüter, C.; Friedrich, P.; Kumsta, R.; Moser, D.; Güntürkün, O.; Genç, E.; Ocklenburg, S. Schizotypy and altered hemispheric asymmetries: The role of cilia genes. Psychiatry Res. Neuroimaging 2019, 294, 110991. [Google Scholar] [CrossRef] [PubMed]
- Signore, I.A.; Palma, K.; Concha, M.L. Nodal signalling and asymmetry of the nervous system. Philos. Trans. R. Soc. B: Biol. Sci. 2016, 371, 20150401. [Google Scholar] [CrossRef] [Green Version]
- Arning, L.; Ocklenburg, S.; Schulz, S.; Ness, V.; Gerding, W.M.; Hengstler, J.G.; Falkenstein, M.; Epplen, J.T.; Güntürkün, O.; Beste, C. PCSK6 VNTR Polymorphism Is Associated with Degree of Handedness but Not Direction of Handedness. PLoS ONE 2013, 8, e67251. [Google Scholar] [CrossRef] [Green Version]
- Berretz, G.; Arning, L.; Gerding, W.M.; Friedrich, P.; Fraenz, C.; Schlüter, C.; Epplen, J.T.; Güntürkün, O.; Beste, C.; Genç, E.; et al. Structural Asymmetry in the Frontal and Temporal Lobes Is Associated with PCSK6 VNTR Polymorphism. Mol. Neurobiol. 2019, 56, 7765–7773. [Google Scholar] [CrossRef] [PubMed]
- Postema, M.C.; Carrion-Castillo, A.; Fisher, S.E.; Vingerhoets, G.; Francks, C. The genetics of situs inversus without primary ciliary dyskinesia. Sci. Rep. 2020, 10, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Ihara, A.; Hirata, M.; Fujimaki, N.; Goto, T.; Umekawa, Y.; Fujita, N.; Terazono, Y.; Matani, A.; Wei, Q.; Yoshimine, T.; et al. Neuroimaging study on brain asymmetries in situs inversus totalis. J. Neurol. Sci. 2010, 288, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, D.N.; O’Craven, K.M.; Ticho, B.S.; Goldstein, A.M.; Makris, N.; Henson, J.W. Structural and functional brain asymmetries in human situs inversus totalis. Neurology 1999, 53, 1260. [Google Scholar] [CrossRef] [PubMed]
- Schuler, A.-L.; Kasprian, G.; Schwartz, E.; Seidl, R.; Diogo, M.C.; Mitter, C.; Langs, G.; Prayer, D.; Bartha-Doering, L. Mens inversus in corpore inverso? Language lateralization in a boy with situs inversus totalis. Brain Lang. 2017, 174, 9–15. [Google Scholar] [CrossRef]
- Vingerhoets, G.; Li, X.; Hou, L.; Bogaert, S.; Verhelst, H.; Gerrits, R.; Siugzdaite, R.; Roberts, N. Brain structural and functional asymmetry in human situs inversus totalis. Brain Struct. Funct. 2018, 223, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Vingerhoets, G.; Gerrits, R.; Bogaert, S. Atypical brain functional segregation is more frequent in situs inversus totalis. Cortex 2018, 106, 12–25. [Google Scholar] [CrossRef]
- Downes, J.C.; Birsoy, B.; Chipman, K.C.; Rothman, J.H. Handedness of a Motor Program in C. elegans Is Independent of Left-Right Body Asymmetry. PLoS ONE 2012, 7, e52138. [Google Scholar] [CrossRef] [Green Version]
- Barth, K.A.; Miklosi, A.; Watkins, J.; Bianco, I.H.; Wilson, S.W.; Andrew, R.J. fsi Zebrafish Show Concordant Reversal of Laterality of Viscera, Neuroanatomy, and a Subset of Behavioral Responses. Curr. Biol. 2005, 15, 844–850. [Google Scholar] [CrossRef] [Green Version]
- McManus, C. Reversed Bodies, Reversed Brains, and (Some) Reversed Behaviors: Of Zebrafish and Men. Dev. Cell 2005, 8, 796–797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halpern, M. Leaning to the left: Laterality in the zebrafish forebrain. Trends Neurosci. 2003, 26, 308–313. [Google Scholar] [CrossRef]
- Hobert, O. Homeobox genes and the specification of neuronal identity. Nat. Rev. Neurosci. 2021, 22, 627. [Google Scholar] [CrossRef] [PubMed]
- De Kovel, C.G.F.; Lisgo, S.N.; Fisher, S.; Francks, C. Subtle left-right asymmetry of gene expression profiles in embryonic and foetal human brains. Sci. Rep. 2018, 8, 12606. [Google Scholar] [CrossRef] [Green Version]
- Hepper, P.G.; Mccartney, G.R.; Shannon, E. Lateralised behaviour in first trimester human foetuses. Neuropsychologia 1998, 36, 531–534. [Google Scholar] [CrossRef]
- McCartney, G.; Hepper, P. Development of lateralized behaviour in the human fetus from 12 to 27 weeks’ gestation. Dev. Med. Child Neurol. 1999, 41, 83–86. [Google Scholar] [CrossRef]
- Sun, T.; Collura, R.V.; Ruvolo, M.; Walsh, C.A. Genomic and Evolutionary Analyses of Asymmetrically Expressed Genes in Human Fetal Left and Right Cerebral Cortex. Cereb. Cortex 2006, 16, i18–i25. [Google Scholar] [CrossRef] [Green Version]
- Sun, T.; Patoine, C.; Abu-Khalil, A.; Visvader, J.; Sum, E.; Cherry, T.J.; Orkin, S.H.; Geschwind, D.H.; Walsh, C.A. Early Asymmetry of Gene Transcription in Embryonic Human Left and Right Cerebral Cortex. Science 2005, 308, 1794–1798. [Google Scholar] [CrossRef] [Green Version]
- Cederquist, G.Y.; Azim, E.; Shnider, S.J.; Padmanabhan, H.; Macklis, J.D. Lmo4 Establishes Rostral Motor Cortex Projection Neuron Subtype Diversity. J. Neurosci. 2013, 33, 6321–6332. [Google Scholar] [CrossRef] [Green Version]
- Asprer, J.S.T.; Lee, B.; Wu, C.S.; Vadakkan, T.; Dickinson, M.E.; Lu, H.-C.; Lee, S.-K. LMO4 functions as a co-activator of neurogenin 2 in the developing cortex. Development 2011, 138, 2823–2832. [Google Scholar] [CrossRef] [Green Version]
- Kashani, A.H.; Qiu, Z.; Jurata, L.; Lee, S.-K.; Pfaff, S.; Goebbels, S.; Nave, K.-A.; Ghosh, A. Calcium Activation of the LMO4 Transcription Complex and Its Role in the Patterning of Thalamocortical Connections. J. Neurosci. 2006, 26, 8398–8408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nudel, R.; Newbury, D.F. Foxp2. Wiley Interdiscip. Rev. Cogn. Sci. 2013, 4, 547–560. [Google Scholar] [CrossRef] [PubMed]
- Ocklenburg, S.; Arning, L.; Gerding, W.M.; Epplen, J.T.; Güntürkün, O.; Beste, C. FOXP2 variation modulates functional hemispheric asymmetries for speech perception. Brain Lang. 2013, 126, 279–284. [Google Scholar] [CrossRef] [PubMed]
- Wong, R.O.L.; Ghosh, A. Activity-dependent regulation of dendritic growth and patterning. Nat. Rev. Neurosci. 2002, 3, 803–812. [Google Scholar] [CrossRef] [PubMed]
- Karlebach, G.; Francks, C. Lateralization of gene expression in human language cortex. Cortex 2015, 67, 30–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moskal, J.R.; Kroes, R.A.; Otto, N.J.; Rahimi, O.; Claiborne, B.J. Distinct patterns of gene expression in the left and right hippocampal formation of developing rats. Hippocampus 2006, 16, 629–634. [Google Scholar] [CrossRef] [PubMed]
- Ribic, A.; Biederer, T. Emerging Roles of Synapse Organizers in the Regulation of Critical Periods. Neural Plast. 2019, 2019, 1–9. [Google Scholar] [CrossRef]
- Beste, C.; Arning, L.; Gerding, W.M.; Epplen, J.T.; Mertins, A.; Röder, M.C.; Bless, J.J.; Hugdahl, K.; Westerhausen, R.; Güntürkün, O.; et al. Cognitive Control Processes and Functional Cerebral Asymmetries: Association with Variation in the Handedness-Associated Gene LRRTM. Mol. Neurobiol. 2018, 55, 2268–2274. [Google Scholar] [CrossRef] [PubMed]
- Francks, C.; Maegawa, S.; Laurén, J.; Abrahams, B.S.; Velayos-Baeza, A.; Medland, S.; Colella, S.; Groszer, M.; McAuley, E.Z.; Caffrey, T.M.; et al. LRRTM1 on chromosome 2p12 is a maternally suppressed gene that is associated paternally with handedness and schizophrenia. Mol. Psychiatry 2007, 12, 1129–1139. [Google Scholar] [CrossRef]
- Berardi, N.; Maffei, L. From visual experience to visual function: Roles of neurotrophins. J. Neurobiol. 1999, 41, 119–126. [Google Scholar] [CrossRef]
- Vicario-Abejón, C.; Owens, D.; McKay, R.; Segal, M. Role of neurotrophins in central synapse formation and stabilization. Nat. Rev. Neurosci. 2002, 3, 965–974. [Google Scholar] [CrossRef] [PubMed]
- Sardar, R.; Zandieh, Z.; Namjoo, Z.; Soleimani, M.; Shirazi, R.; Hami, J. Laterality and sex differences in the expression of brain-derived neurotrophic factor in developing rat hippocampus. Metab. Brain Dis. 2021, 36, 133–144. [Google Scholar] [CrossRef]
- Marrocco, J.; McEwen, B.S. Sex in the brain: Hormones and sex differences. Dialog-Clin. Neurosci. 2016, 18, 373–383. [Google Scholar] [CrossRef]
- Deng, C.; Rogers, L.J. Factors affecting the development of lateralization in chicks. In Comparative Vertebrate Lateralization; Rogers, L.J., Andrew, R., Eds.; Cambridge University Press: Cambridge, UK, 2002; pp. 206–246. [Google Scholar]
- Rogers, L.J. Development and function of lateralization in the avian brain. Brain Res. Bull. 2008, 76, 235–244. [Google Scholar] [CrossRef] [PubMed]
- Manns, M.; Otto, T.; Salm, L. Pigeons show how meta-control enables decision-making in an ambiguous world. Sci. Rep. 2021, 11, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Rogers, L.J. A Matter of Degree: Strength of Brain Asymmetry and Behaviour. Symmetry 2017, 9, 57. [Google Scholar] [CrossRef] [Green Version]
- Diekamp, B.; Regolin, L.; Güntürkün, O.; Vallortigara, G. A left-sided visuospatial bias in birds. Curr. Biol. 2005, 15, R372–R373. [Google Scholar] [CrossRef]
- Manns, M. The riddle of nature and nurture-lateralization has an epigenetic trait. Behav. Brain Sci. 2005, 28, 602–603. [Google Scholar] [CrossRef]
- Manns, M. The epigenetic control of asymmetry formation: Lessons from the avian visual system. In Behavioral and Morphological Asymmetries in Vertebrates; Malashichev, Y., Deckel, W., Eds.; Landes Bioscience: Georgetown, TX, USA, 2006; pp. 13–23. [Google Scholar]
- Monsoro-Burq, A.H.; Levin, M. Avian models and the study of invariant asymmetry: How the chicken and the egg taught us to tell right from left. Int. J. Dev. Biol. 2018, 62, 63–77. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Guo, Q.; Dai, E.; Forsch, N.; Taber, L.A. How the embryonic chick brain twists. J. R. Soc. Interface 2016, 13, 20160395. [Google Scholar] [CrossRef] [Green Version]
- Fujinaga, M.; Hoffman, B.B.; Baden, J.M. Axial rotation in rat embryos: Morphological analysis and microsurgical study on the role of the allantois. Teratology 1995, 51, 94–106. [Google Scholar] [CrossRef]
- Andrew, R.J.; Johnston, A.N.B.; Robins, A.; Rogers, L.J. Light experience and the development of behavioural lateralisation in chicks. II. Choice of familiar versus unfamiliar model social partner. Behav. Brain Res. 2004, 155, 67–76. [Google Scholar] [CrossRef]
- Deng, C.; Rogers, L.J. Social recognition and approach in the chick: Lateralization and effect of visual experience. Anim. Behav. 2002, 63, 697–706. [Google Scholar] [CrossRef] [Green Version]
- Mascetti, G.G.; Vallortigara, G. Why do birds sleep with one eye open? Light exposure of the chick embryo as a determinant of monocular sleep. Curr. Biol. 2001, 11, 971–974. [Google Scholar] [CrossRef] [Green Version]
- Costalunga, G.; Kobylkov, D.; Rosa-Salva, O.; Vallortigara, G.; Mayer, U. Light-incubation effects on lateralisation of single unit responses in the visual Wulst of domestic chicks. Brain Struct. Funct. 2021, 1–17. [Google Scholar] [CrossRef]
- Johnston, A.; Rogers, L.; Dodd, P. [3H]MK-801 binding asymmetry in the IMHV region of dark-reared chicks is reversed by imprinting. Brain Res. Bull. 1995, 37, 5–8. [Google Scholar] [CrossRef]
- Lorenzi, E.; Mayer, U.; Rosa-Salva, O.; Morandi-Raikova, A.; Vallortigara, G. Spontaneous and light-induced lateralization of immediate early genes expression in domestic chicks. Behav. Brain Res. 2019, 368, 111905. [Google Scholar] [CrossRef] [PubMed]
- Morandi-Raikova, A.; Danieli, K.; Lorenzi, E.; Rosa-Salva, O.; Mayer, U. Anatomical asymmetries in the tectofugal pathway of dark-incubated domestic chicks: Rightwards lateralization of parvalbumin neurons in the entopallium. Laterality 2021, 26, 163–185. [Google Scholar] [CrossRef] [PubMed]
- Letzner, S.; Patzke, N.; Verhaal, J.; Manns, M. Shaping a lateralized brain: Asymmetrical light experience modulates access to visual interhemispheric information in pigeons. Sci. Rep. 2014, 4, 4253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Letzner, S.; Güntürkün, O.; Lor, S.; Pawlik, R.J.; Manns, M. Visuospatial attention in the lateralised brain of pigeons—A matter of ontogenetic light experiences. Sci. Rep. 2017, 7, 15547. [Google Scholar] [CrossRef] [Green Version]
- Chiandetti, C. Pseudoneglect and embryonic light stimulation in the avian brain. Behav. Neurosci. 2011, 125, 775–782. [Google Scholar] [CrossRef]
- Chiandetti, C.; Galliussi, J.; Andrew, R.J.; Vallortigara, G. Early-light embryonic stimulation suggests a second route, via gene activation, to cerebral lateralization in vertebrates. Sci. Rep. 2013, 3, 2701. [Google Scholar] [CrossRef] [Green Version]
- Bobbo, D.; Galvani, F.; Mascetti, G.G.; Vallortigara, G. Light exposure of the chick embryo influences monocular sleep. Behav. Brain Res. 2002, 134, 447–466. [Google Scholar] [CrossRef]
- Zhu, L.; Marvin, M.J.; Gardiner, A.; Lassar, A.B.; Mercola, M.; Stern, C.D.; Levin, M. Cerberus regulates left–right asymmetry of the embryonic head and heart. Curr. Biol. 1999, 9, 931–938. [Google Scholar] [CrossRef] [Green Version]
- Ververs, I.A.; De Vries, J.I.; Van Geijn, H.P.; Hopkins, B. Prenatal head position from 12–38 weeks. I. Developmental aspects. Early Hum. Dev. 1994, 39, 83–91. [Google Scholar] [CrossRef]
- Mampe, B.; Friederici, A.D.; Christophe, A.; Wermke, K. Newborns’ Cry Melody Is Shaped by Their Native Language. Curr. Biol. 2009, 19, 1994–1997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Partanen, E.; Kujala, T.; Näätänen, R.; Liitola, A.; Sambeth, A.; Huotilainen, M. Learning-induced neural plasticity of speech processing before birth. Proc. Natl. Acad. Sci. USA 2013, 110, 15145–15150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fagard, J. The nature and nurture of human infant hand preference. Ann. N. Y. Acad. Sci. 2013, 1288, 114–123. [Google Scholar] [CrossRef]
- Ocklenburg, S.; Bürger, C.; Westermann, C.; Schneider, D.; Biedermann, H.; Güntürkün, O. Visual experience affects handedness. Behav. Brain Res. 2010, 207, 447–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Previc, F.H. A general theory concerning the prenatal origins of cerebral lateralization in humans. Psychol. Rev. 1991, 98, 299–334. [Google Scholar] [CrossRef] [PubMed]
- Andrew, R.; Osorio, D.; Budaev, S. Light during embryonic development modulates patterns of lateralization strongly and similarly in both zebrafish and chick. Philos. Trans. R. Soc. B Biol. Sci. 2008, 364, 983–989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Budaev, S.; Andrew, R.J. Patterns of early embryonic light exposure determine behavioural asymmetries in zebrafish: A habenular hypothesis. Behav. Brain Res. 2009, 200, 91–94. [Google Scholar] [CrossRef] [PubMed]
- Chiandetti, C.; Vallortigara, G. Distinct effect of early and late embryonic light-stimulation on chicks’ lateralization. Neuroscience 2019, 414, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Caputto, B.L.; Guido, M.E. Immediate early gene expression within the visual system: Light and circadian regulation in the retina and the suprachiasmatic nucleus. Neurochem. Res. 2000, 25, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Ströckens, F.; Freund, N.; Manns, M.; Ocklenburg, S.; Güntürkün, O. Visual asymmetries and the ascending thalamofugal pathway in pigeons. Brain Struct. Funct. 2012, 218, 1197–1209. [Google Scholar] [CrossRef]
- Letzner, S.; Manns, M.; Güntürkün, O. Light-dependent development of the tectorotundal projection in pigeons. Eur. J. Neurosci. 2020, 52, 3561–3571. [Google Scholar] [CrossRef] [PubMed]
- Rogers, L.J. Light experience and asymmetry of brain function in chickens. Nat. Cell Biol. 1982, 297, 223–225. [Google Scholar] [CrossRef]
- Skiba, M.; Diekamp, B.; Güntürkün, O. Embryonic light stimulation induces different asymmetries in visuoperceptual and visuomotor pathways of pigeons. Behav. Brain Res. 2002, 134, 149–156. [Google Scholar] [CrossRef] [Green Version]
- Rogers, L.J. Light input and the reversal of functional lateralization in the chicken brain. Behav. Brain Res. 1990, 38, 211–221. [Google Scholar] [CrossRef]
- Manns, M.; Güntürkün, O. Monocular deprivation alters the direction of functional and morphological asymmetries in the pigeon’s (Columba livia) visual system. Behav. Neurosci. 1999, 113, 1257–1266. [Google Scholar] [CrossRef]
- Manns, M.; Güntürkün, O. ‘Natural’ and artificial monocular deprivation effects on thalamic soma sizes in pigeons. NeuroReport 1999, 10, 3223–3228. [Google Scholar] [CrossRef] [Green Version]
- Rogers, L.; Deng, C. Corticosterone treatment of the chick embryo affects light-stimulated development of the thalamofugal visual pathway. Behav. Brain Res. 2005, 159, 63–71. [Google Scholar] [CrossRef]
- Hausmann, M. Why sex hormones matter for neuroscience: A very short review on sex, sex hormones, and functional brain asymmetries. J. Neurosci. Res. 2017, 95, 40–49. [Google Scholar] [CrossRef] [Green Version]
- Hirnstein, M.; Hugdahl, K.; Hausmann, M. Cognitive sex differences and hemispheric asymmetry: A critical review of 40 years of research. Laterality 2019, 24, 204–252. [Google Scholar] [CrossRef]
- Manns, M.; Güntürkün, O. Light experience induces differential asymmetry pattern of GABA- and parvalbumin-positive cells in the pigeon’s visual midbrain. J. Chem. Neuroanat. 2003, 25, 249–259. [Google Scholar] [CrossRef]
- Prior, H.; Diekamp, B.; Güntürkün, O.; Manns, M. Post-hatch activity-dependent modulation of visual asymmetry formation in pigeons. NeuroReport 2004, 15, 1311–1314. [Google Scholar] [CrossRef] [PubMed]
- Manns, M.; Freund, N.; Leske, O.; Güntürkün, O. Breaking the balance: Ocular BDNF-injections induce visual asymmetry in pigeons. Dev. Neurobiol. 2008, 68, 1123–1134. [Google Scholar] [CrossRef] [PubMed]
- Manns, M.; Güntürkün, O.; Heumann, R.; Blöchl, A. Photic inhibition of TrkB/Ras activity in the pigeon’s tectum during development: Impact on brain asymmetry formation. Eur. J. Neurosci. 2005, 22, 2180–2186. [Google Scholar] [CrossRef] [PubMed]
- Oviedo, H.V.; Bureau, I.; Svoboda, K.; Zador, A.M. The functional asymmetry of auditory cortex is reflected in the organization of local cortical circuits. Nat. Neurosci. 2010, 13, 1413–1420. [Google Scholar] [CrossRef]
- Nava, E.; Güntürkün, O.; Röder, B. Experience-dependent emergence of functional asymmetries. Laterality 2013, 18, 407–415. [Google Scholar] [CrossRef] [Green Version]
- Le Grand, R.; Mondloch, C.J.; Maurer, D.; Brent, H.P. Early visual experience and face processing. Nat. Cell Biol. 2001, 410, 890. [Google Scholar] [CrossRef]
- Röder, B.; Ley, P.; Shenoy, B.H.; Kekunnaya, R.; Bottari, D. Sensitive periods for the functional specialization of the neural system for human face processing. Proc. Natl. Acad. Sci. USA 2013, 110, 16760–16765. [Google Scholar] [CrossRef] [Green Version]
- Dehaene, S.; Pegado, F.; Braga, L.W.; Ventura, P.; Filho, G.N.; Jobert, A.; Dehaene-Lambertz, G.; Kolinsky, R.; Morais, J.; Cohen, L. How Learning to Read Changes the Cortical Networks for Vision and Language. Science 2010, 330, 1359–1364. [Google Scholar] [CrossRef] [Green Version]
- Dundas, E.M.; Plaut, D.C.; Behrmann, M. An ERP investigation of the co-development of hemispheric lateralization of face and word recognition. Neuropsychology 2014, 61, 315–323. [Google Scholar] [CrossRef] [Green Version]
- Freund, N.; Valencia-Alfonso, C.E.; Kirsch, J.; Brodmann, K.; Manns, M.; Güntürkün, O. Asymmetric top-down modulation of ascending visual pathways in pigeons. Neuropsychology 2016, 83, 37–47. [Google Scholar] [CrossRef]
- Manns, M.; Römling, J. The impact of asymmetrical light input on cerebral hemispheric specialization and interhemispheric cooperation. Nat. Commun. 2012, 3, 696. [Google Scholar] [CrossRef] [PubMed]
- Dharmaretnam, M.; Rogers, L. Hemispheric specialization and dual processing in strongly versus weakly lateralized chicks. Behav. Brain Res. 2005, 162, 62–70. [Google Scholar] [CrossRef] [PubMed]
- Chiandetti, C.; Regolin, L.; Rogers, L.J.; Vallortigara, G. Effects of light stimulation of embryos on the use of position-specific and object-specific cues in binocular and monocular domestic chicks (Gallus gallus). Behav. Brain Res. 2005, 163, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Gazzaniga, M.S. Forty-five years of split-brain research and still going strong. Nat. Rev. Neurosci. 2005, 6, 653–659. [Google Scholar] [CrossRef]
- Witelson, S.; Nowakowski, R. Left out axoms make men right: A hypothesis for the origin of handedness and functional asymmetry. Neuropsychology 1991, 29, 327–333. [Google Scholar] [CrossRef]
- Besson, M.; Gache, C.; Bertucci, F.; Brooker, R.; Roux, N.; Jacob, H.; Berthe, C.; Sovrano, V.A.; Dixson, D.L.; Lecchini, D. Exposure to agricultural pesticide impairs visual lateralization in a larval coral reef fish. Sci. Rep. 2017, 7, 1–9. [Google Scholar] [CrossRef]
- Broder, E.D.; Angeloni, L.M. Predator-induced phenotypic plasticity of laterality. Anim. Behav. 2014, 98, 125–130. [Google Scholar] [CrossRef]
- Jozet-Alves, C.; Hébert, M. Embryonic exposure to predator odour modulates visual lateralization in cuttlefish. Proc. R. Soc. B Boil. Sci. 2013, 280, 20122575. [Google Scholar] [CrossRef] [Green Version]
- Manns, M. Laterality for the next decade: Costs and benefits of neuronal asymmetries–putting lateralization in an evolutionary context. Laterality 2021, 26, 315–318. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Model of the hierarchical development of brain lateralization—the three main phases of neuronal development are dominated by different cellular processes, which lead to an increasingly lateralized functional organization of the two hemispheres (indicated by the green and orange triangles). In each phase, certain types of genes regulate differentiation and, thus, asymmetry formation.
Figure 1.
Model of the hierarchical development of brain lateralization—the three main phases of neuronal development are dominated by different cellular processes, which lead to an increasingly lateralized functional organization of the two hemispheres (indicated by the green and orange triangles). In each phase, certain types of genes regulate differentiation and, thus, asymmetry formation.

Figure 2.
The developing visual systems of chicks and pigeons exemplifies how one environmental factor—in this case light—affects the development of brain asymmetries during the three main phases of neuronal development (see text for details).
Figure 2.
The developing visual systems of chicks and pigeons exemplifies how one environmental factor—in this case light—affects the development of brain asymmetries during the three main phases of neuronal development (see text for details).

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Manns, M. It Is Not Just in the Genes. Symmetry 2021, 13, 1815. https://doi.org/10.3390/sym13101815
AMA Style
Manns M. It Is Not Just in the Genes. Symmetry. 2021; 13(10):1815. https://doi.org/10.3390/sym13101815
Chicago/Turabian StyleManns, Martina. 2021. "It Is Not Just in the Genes" Symmetry 13, no. 10: 1815. https://doi.org/10.3390/sym13101815
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.