Presence of Flavivirus Antibodies Does Not Lead to a Greater Number of Symptoms in a Small Cohort of Canadian Travelers Infected with Zika Virus

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Virus and Antibody Detection
2.3. Statistical Analysis
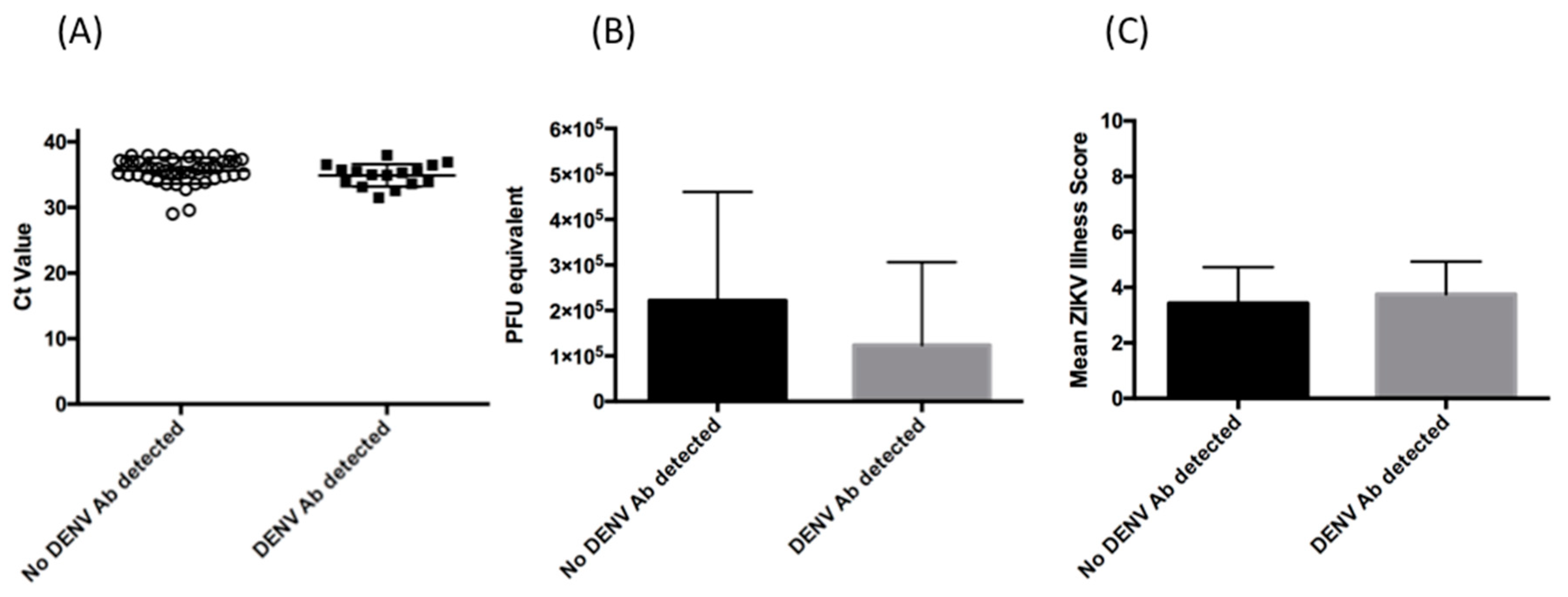
3. Results
3.1. Cohort Description
3.2. Relative Magnitude of Viremia
3.3. Clinical Presentation
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kass, D.E.; Merlino, M. Zika Virus. N. Engl. J. Med. 2016, 375, 294–295. [Google Scholar] [CrossRef]
- De Oliveira Garcia, M.H. Zika: The continuing threat. Bull. World Health Organ. 2019, 97, 6–7. [Google Scholar] [CrossRef]
- Patterson, J.; Sammon, M.; Garg, M. Dengue, Zika and Chikungunya: Emerging Arboviruses in the New World. West J. Emerg. Med. 2016, 17, 671–679. [Google Scholar] [CrossRef] [PubMed]
- Martin-Acebes, M.A.; Saiz, J.C.; Jimenez de Oya, N. Antibody-Dependent Enhancement and Zika: Real Threat or Phantom Menace? Front Cell Infect. Microbiol. 2018, 8, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katzelnick, L.C.; Gresh, L.; Halloran, M.E.; Mercado, J.C.; Kuan, G.; Gordon, A.; Balmaseda, A.; Harris, E. Antibody-dependent enhancement of severe dengue disease in humans. Science 2017, 358, 929–932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawiecki, A.B.; Christofferson, R.C. Zika Virus-Induced Antibody Response Enhances Dengue Virus Serotype 2 Replication In Vitro. J. Infect. Dis. 2016, 214, 1357–1360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Priyamvada, L.; Quicke, K.M.; Hudson, W.H.; Onlamoon, N.; Sewatanon, J.; Edupganti, S.; Pattanapanyasat, K.; Chokephaibulkit, K.; Mulligan, M.; Wilson, C.; et al. Human antibody responses after dengue virus infection are highly cross-reactive to Zika virus. Proc. Natl. Acad. Sci. USA 2016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rogers, T.F.; Goodwin, E.C.; Briney, B.; Sok, D.; Beutler, N.; Strubel, A.; Nedellec, R.; Le, K.; Brown, M.E.; Burton, D.R.; et al. Zika virus activates de novo and cross-reactive memory B cell responses in dengue-experienced donors. Sci. Immunol. 2017, 2. [Google Scholar] [CrossRef] [Green Version]
- Bardina, S.V.; Bunduc, P.; Tripathi, S.; Duehr, J.; Frere, J.J.; Brown, J.A.; Nachbagauer, R.; Foster, G.A.; Krysztof, D.; Tortorella, D.; et al. Enhancement of Zika virus pathogenesis by preexisting antiflavivirus immunity. Science 2017, 356, 175–180. [Google Scholar] [CrossRef] [Green Version]
- George, J.; Valiant, W.G.; Mattapallil, M.J.; Walker, M.; Huang, Y.S.; Vanlandingham, D.L.; Misamore, J.; Greenhouse, J.; Weiss, D.E.; Verthelyi, D.; et al. Prior Exposure to Zika Virus Significantly Enhances Peak Dengue-2 Viremia in Rhesus Macaques. Sci. Rep. 2017, 7, 10498. [Google Scholar] [CrossRef]
- Kam, Y.W.; Lee, C.Y.; Teo, T.H.; Howland, S.W.; Amrun, S.N.; Lum, F.M.; See, P.; Kng, N.Q.; Huber, R.G.; Xu, M.H.; et al. Cross-reactive dengue human monoclonal antibody prevents severe pathologies and death from Zika virus infections. JCI Insight 2017, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCracken, M.K.; Gromowski, G.D.; Friberg, H.L.; Lin, X.; Abbink, P.; de La Barrera, R.; Eckles, K.H.; Garver, L.S.; Boyd, M.; Jetton, D.; et al. Impact of prior flavivirus immunity on Zika virus infection in rhesus macaques. PLoS Pathog. 2017, 13, e1006487. [Google Scholar] [CrossRef]
- Dejnirattisai, W.; Supasa, P.; Wongwiwat, W.; Rouvinski, A.; Barba-Spaeth, G.; Duangchinda, T.; Sakuntabhai, A.; Cao-Lormeau, V.M.; Malasit, P.; Rey, F.A.; et al. Dengue virus sero-cross-reactivity drives antibody-dependent enhancement of infection with zika virus. Nat. Immunol. 2016, 17, 1102–1108. [Google Scholar] [CrossRef] [PubMed]
- Waggoner, J.J.; Pinsky, B.A. Zika Virus: Diagnostics for an Emerging Pandemic Threat. J. Clin. Microbiol. 2016, 54, 860–867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rabe, I.B.; Staples, J.E.; Villanueva, J.; Hummel, K.B.; Johnson, J.A.; Rose, L.; Rose, L.; Hills, S.; Wasley, A.; Fischer, M.; et al. Interim Guidance for Interpretation of Zika Virus Antibody Test Results. MMWR Morb. Mortal. Wkly. Rep. 2016, 65, 543–546. [Google Scholar] [CrossRef] [Green Version]
- L’Huillier, A.G.; Lombos, E.; Tang, E.; Perusini, S.; Eshaghi, A.; Nagra, S.; Frantz, C.; Olsha, R.; Kristjanson, E.; Dimitrova, K.; et al. Evaluation of Altona Diagnostics RealStar Zika Virus Reverse Transcription-PCR Test Kit for Zika Virus PCR Testing. J. Clin. Microbiol. 2017, 55, 1576–1584. [Google Scholar] [CrossRef] [Green Version]
- Pongsiri, P.; Praianantathavorn, K.; Theamboonlers, A.; Payungporn, S.; Poovorawan, Y. Multiplex real-time RT-PCR for detecting chikungunya virus and dengue virus. Asian Pac. J. Trop. Med. 2012, 5, 342–346. [Google Scholar] [CrossRef] [Green Version]
- Calisher, C.H.; Karabatsos, N.; Dalrymple, J.M.; Shope, R.E.; Porterfield, J.S.; Westaway, E.G.; Brandt, W.E. Antigenic relationships between flaviviruses as determined by cross-neutralization tests with polyclonal antisera. J. Gen. Virol. 1989, 70, 37–43. [Google Scholar] [CrossRef]
- De La Vega, M.A.; Caleo, G.; Audet, J.; Qiu, X.; Kozak, R.A.; Brooks, J.I.; Kern, S.; Wolz, A.; Sprecher, A.; Greig, J.; et al. Ebola viral load at diagnosis associates with patient outcome and outbreak evolution. J. Clin. Investig. 2015, 125, 4421–4428. [Google Scholar] [CrossRef] [Green Version]
- Fourcade, C.; Mansuy, J.M.; Dutertre, M.; Delpech, M.; Marchou, B.; Delobel, P.; Izopet, J.; Martin-Blondel, G. Viral load kinetics of Zika virus in plasma, urine and saliva in a couple returning from Martinique, French West Indies. J. Clin. Virol. 2016, 82, 1–4. [Google Scholar] [CrossRef]
- Read, J.S.; Torres-Velasquez, B.; Lorenzi, O.; Rivera Sanchez, A.; Torres-Torres, S.; Rivera, L.V.; Capre-Franceschi, S.M.; Garcia-Gubern, C.; Munoz-Jordan, J.; Santiago, G.A.; et al. Symptomatic Zika Virus Infection in Infants, Children, and Adolescents Living in Puerto Rico. JAMA Pediatr. 2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colombo, T.E.; Estofolete, C.F.; Reis, A.F.N.; da Silva, N.S.; Aguiar, M.L.; Cabrera, E.M.S.; Dos Santos, I.N.P.; Costa, F.R.; Cruz, L.; Rombola, P.L.; et al. Clinical, laboratory and virological data from suspected ZIKV patients in an endemic arbovirus area. J. Clin. Virol. 2017, 96, 20–25. [Google Scholar] [CrossRef] [PubMed]
- Dick, G.W. Zika virus. II. Pathogenicity and physical properties. Trans. R. Soc. Trop. Med. Hyg. 1952, 46, 521–534. [Google Scholar] [CrossRef]
- Dick, G.W.; Kitchen, S.F.; Haddow, A.J. Zika virus. I. Isolations and serological specificity. Trans. R. Society Trop. Med. Hyg. 1952, 46, 509–520. [Google Scholar] [CrossRef]
- Terzian, A.C.B.; Schanoski, A.S.; Mota, M.T.O.; da Silva, R.A.; Estofolete, C.F.; Colombo, T.E.; Rahal, P.; Hanley, K.A.; Vasilakis, N.; Kalil, J.; et al. Viral Load and Cytokine Response Profile Does Not Support Antibody-Dependent Enhancement in Dengue-Primed Zika Virus-Infected Patients. Clin. Infect. Dis. 2017, 65, 1260–1265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santiago, G.A.; Sharp, T.M.; Rosenberg, E.; Sosa Cardona, I.I.; Alvarado, L.; Paz-Bailey, G.; Munoz-Jordan, J.L. Prior Dengue Virus Infection Is Associated With Increased Viral Load in Patients Infected With Dengue but Not Zika Virus. Open Forum Infect. Dis. 2019, 6. [Google Scholar] [CrossRef]
- Petersen, L.R.; Jamieson, D.J.; Powers, A.M.; Honein, M.A. El virus del Zika. N. Engl. J. Med. 2016, 374, 1552–1563. [Google Scholar] [CrossRef]
- Halai, U.A.; Nielsen-Saines, K.; Moreira, M.L.; de Sequeira, P.C.; Junior, J.P.P.; de Araujo Zin, A.; Cherry, J.; Gabaglia, C.R.; Gaw, S.L.; Adachi, K.; et al. Maternal Zika Virus Disease Severity, Virus Load, Prior Dengue Antibodies, and Their Relationship to Birth Outcomes. Clin. Infect. Dis. 2017, 65, 877–883. [Google Scholar] [CrossRef] [Green Version]
- Pedroso, C.; Fischer, C.; Feldmann, M.; Sarno, M.; Luz, E.; Moreira-Soto, A.; Cabral, R.; Netto, E.M.; Brites, C.; Kummerer, B.M.; et al. Cross-Protection of Dengue Virus Infection against Congenital Zika Syndrome, Northeastern Brazil. Emerg. Infect. Dis. 2019, 25, 1485–1493. [Google Scholar] [CrossRef] [Green Version]
- Chia, P.Y.; Yew, H.S.; Ho, H.; Chow, A.; Sadarangani, S.P.; Chan, M.; Kam, Y.W.; Chong, C.Y.; Thoon, K.C.; Yung, C.F.; et al. Clinical features of patients with Zika and dengue virus co-infection in Singapore. J. Infect. 2017, 74, 611–615. [Google Scholar] [CrossRef] [Green Version]
- Ward, M.J.; Alger, J.; Berrueta, M.; Bock, H.; Buekens, P.; Cafferata, M.L.; Ciganda, A.; Garcia, J.; Garcia, K.; Lopez, W.; et al. Zika Virus and the World Health Organization Criteria for Determining Recent Infection Using Plaque Reduction Neutralization Testing. Am. J. Trop. Med. Hyg. 2018, 99, 780–782. [Google Scholar] [CrossRef] [Green Version]
- Heukelbach, J.; Alencar, C.H.; Kelvin, A.A.; de Oliveira, W.K.; Pamplona de Goes Cavalcanti, L. Zika virus outbreak in Brazil. J. Infect. Dev. Ctries. 2016, 10, 116–120. [Google Scholar] [CrossRef] [PubMed]
- Pantoja, P.; Perez-Guzman, E.X.; Rodriguez, I.V.; White, L.J.; Gonzalez, O.; Serrano, C.; Giavedoni, L.; Hodara, V.; Cruz, L.; Arana, T.; et al. Zika virus pathogenesis in rhesus macaques is unaffected by pre-existing immunity to dengue virus. Nat. Commun. 2017, 8, 15674. [Google Scholar] [CrossRef] [PubMed]
- Montoya, M.; Gresh, L.; Mercado, J.C.; Williams, K.L.; Vargas, M.J.; Gutierrez, G.; Kuan, G.; Gordon, A.; Balmaseda, A.; Harris, E. Symptomatic versus inapparent outcome in repeat dengue virus infections is influenced by the time interval between infections and study year. PLoS Negl. Trop. Dis. 2013, 7, e2357. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| DENV PRNT Reactive (n = 16) | DENV PRNT Non-Reactive (n = 44) | |
|---|---|---|
| Mean age (yrs) | 51.9 | 43 |
| Median age (yrs) | 52.5 | 44 |
| Age range (yrs) | 33–88 | 18–75 |
| % Female | 62.5% (n = 10) | 50% (n = 22) |
| Mean number of days of travel (days) | 15 (range 7–33) | 11 (range 0–37) |
| Mean time from last day of travel to symptom onset (days) | 3 (range 0–9) | 1 (range −8–9) |
| Mean period from symptom onset to specimen collection (days) | 5 (range 0–12) | 5 (range 1–13) |
| ZIKV IgM reactive | 87.5% (n = 14) | 97% (n = 43) |
| DENV PRNT positive | 100% (n = 16) | 0% Δ |
| % tested for acute DENV * | 68% (n = 11) | 57% (n = 25) |
| % tested for acute CHKV * | 63% (n = 10) | 52% (n = 23) |
| Patient | ZIKA PRNT | DENV PRNT | Fold Difference | Interpretation Δ |
|---|---|---|---|---|
| 1 | 0 | 1:20 | N/A | Previous dengue virus exposure |
| 2 | 1:160 | 1:40 | 0.25 | Flavivirus infection, specific virus cannot be identified |
| 3 | 1:20 | 1:40 | 2 | Flavivirus infection, specific virus cannot be identified |
| 4 | 1:20 | 1:80 | 4 | Flavivirus infection, specific virus cannot be identified |
| 5 | 1:40 | 1:160 | 4 | Flavivirus infection, specific virus cannot be identified |
| 6 | 1:20 | 1:160 | 4 | Flavivirus infection, specific virus cannot be identified |
| 7 | 1:40 | 1:160 | 4 | Flavivirus infection, specific virus cannot be identified |
| 8 | 1:320 | 1:320 | 1 | Flavivirus infection, specific virus cannot be identified |
| 9 | 1:40 | 1:1280 | 32 | Flavivirus infection, specific virus cannot be identified |
| 10 | 0 | >1:40 | 4 | Previous dengue virus exposure |
| 11 | 0 | >1:40 | 4 | Previous dengue virus exposure |
| 12 | 0 | >1:40 | 4 | Previous dengue virus exposure |
| 13 | 0 | >1:40 | 4 | Previous dengue virus exposure |
| 14 | 0 | >1:40 | 4 | Previous dengue virus exposure |
| 15 | 0 | >1:40 | 4 | Previous dengue virus exposure |
| 16 | 1:40 | >1:640 | 16 | Flavivirus infection, specific virus cannot be identified |
| DENV or Flavivirus Exposure (n = 16) | No Exposure (n = 44) | |
|---|---|---|
| ACHES | 18.7% (n = 3) | 2.3% (n = 1) |
| ANOREXIA | 0% (n = 0) | 2.3% (n = 1) |
| ARTHRALGIA | 56.3% (n = 9) | 43.2% (n = 19) |
| CHILLS | 12.5% (n = 2) | 4.5% (n = 2) |
| CUNJUNCTIVITIS | 31.3% (n = 5) | 20.5% (n = 9) |
| FATIGUE | 25% (n = 4) | 15.9% (n = 7) |
| FEVER | 43.8% (n = 7) | 61.4% (n = 27) |
| GASTROENTERITIS | 6.3% (n = 1) | 6.8% (n = 3) |
| HEADACHE | 37.5% (n = 6) | 31.8% (n = 14) |
| LYMPHADENITIS | 6.3% (n = 1) | 9% (n = 4) |
| MALAISE | 6.3% (n = 1) | 2.3% (n = 1) |
| MYALGIA | 25% (n = 4) | 25% (n = 11) |
| NAUSEA/VOMITING | 6.3% (n = 1) | 4.5% (n = 2) |
| NEUROLOGICAL | 0% | 6.8% (n = 3) |
| WEAKNESS | 0% | 2.3% (n = 1) |
| RESP. SYMPTOMS | 31.3% (n = 5) | 9.1% (n = 4) |
| RASH | 93.7% (n = 15) | 93.7% (n = 41) |
| OTHER | 12.5% (n = 2) | 2.3% (n = 1) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kozak, R.A.; Goneau, L.W.; DeLima, C.; Varsaneux, O.; Eshaghi, A.; Kristjanson, E.; Olsha, R.; Safronetz, D.; Perusini, S.; Frantz, C.; et al. Presence of Flavivirus Antibodies Does Not Lead to a Greater Number of Symptoms in a Small Cohort of Canadian Travelers Infected with Zika Virus. Viruses 2020, 12, 140. https://doi.org/10.3390/v12020140
Kozak RA, Goneau LW, DeLima C, Varsaneux O, Eshaghi A, Kristjanson E, Olsha R, Safronetz D, Perusini S, Frantz C, et al. Presence of Flavivirus Antibodies Does Not Lead to a Greater Number of Symptoms in a Small Cohort of Canadian Travelers Infected with Zika Virus. Viruses. 2020; 12(2):140. https://doi.org/10.3390/v12020140
Chicago/Turabian StyleKozak, Robert A., Lee W. Goneau, Cedric DeLima, Olivia Varsaneux, AliReza Eshaghi, Erik Kristjanson, Romy Olsha, David Safronetz, Stephen Perusini, Christine Frantz, and et al. 2020. "Presence of Flavivirus Antibodies Does Not Lead to a Greater Number of Symptoms in a Small Cohort of Canadian Travelers Infected with Zika Virus" Viruses 12, no. 2: 140. https://doi.org/10.3390/v12020140