
Glycyrrhizin Effectively Inhibits SARS-CoV-2 Replication by Inhibiting the Viral Main Protease

 , ,
, ,  , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Licorice Root Extract
2.2. Glycyrrhizin Acid Ammonium-Nitrate
2.3. Cells and Virus
2.4. Antiviral Activity of Aqueous Licorice Root Extract and Glycyrrhizin
2.5. Cell Viability Assay
2.6. Determination of EC50 of Glycyrrhizin
2.7. Quantitative SARS-CoV-2 RT-PCR
2.8. Determination of Main Protease Inhibition
3. Results
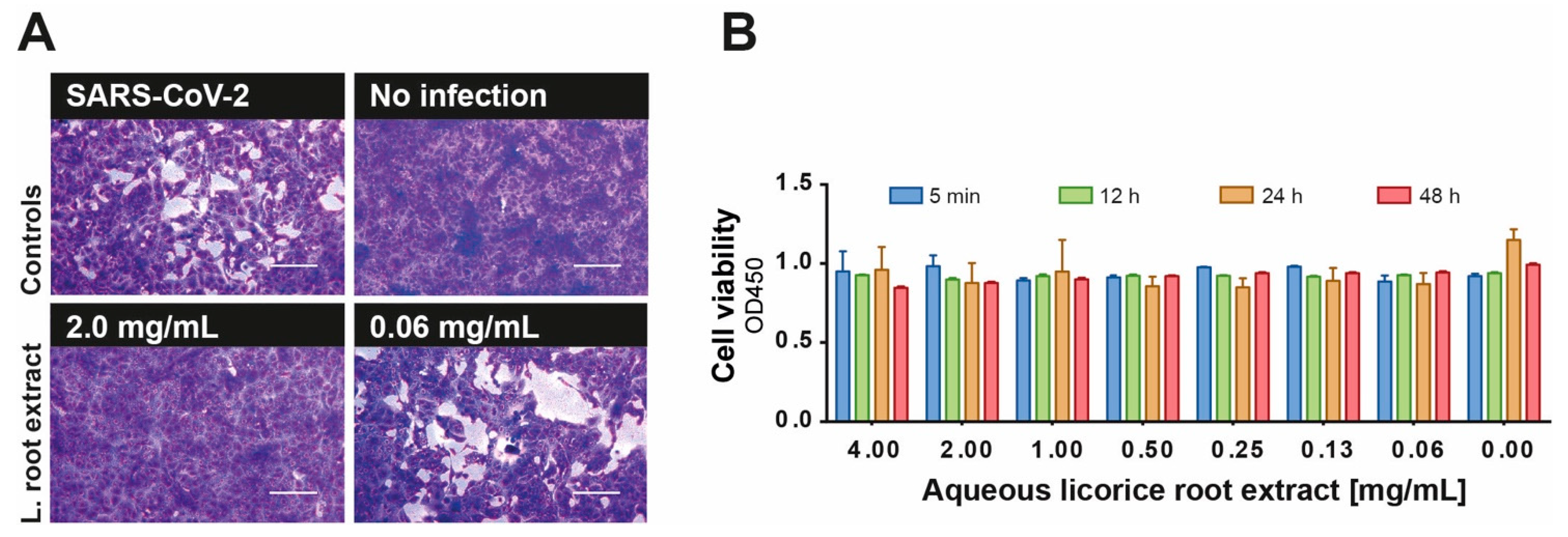
3.1. Antiviral Activity of Licorice Root Extract
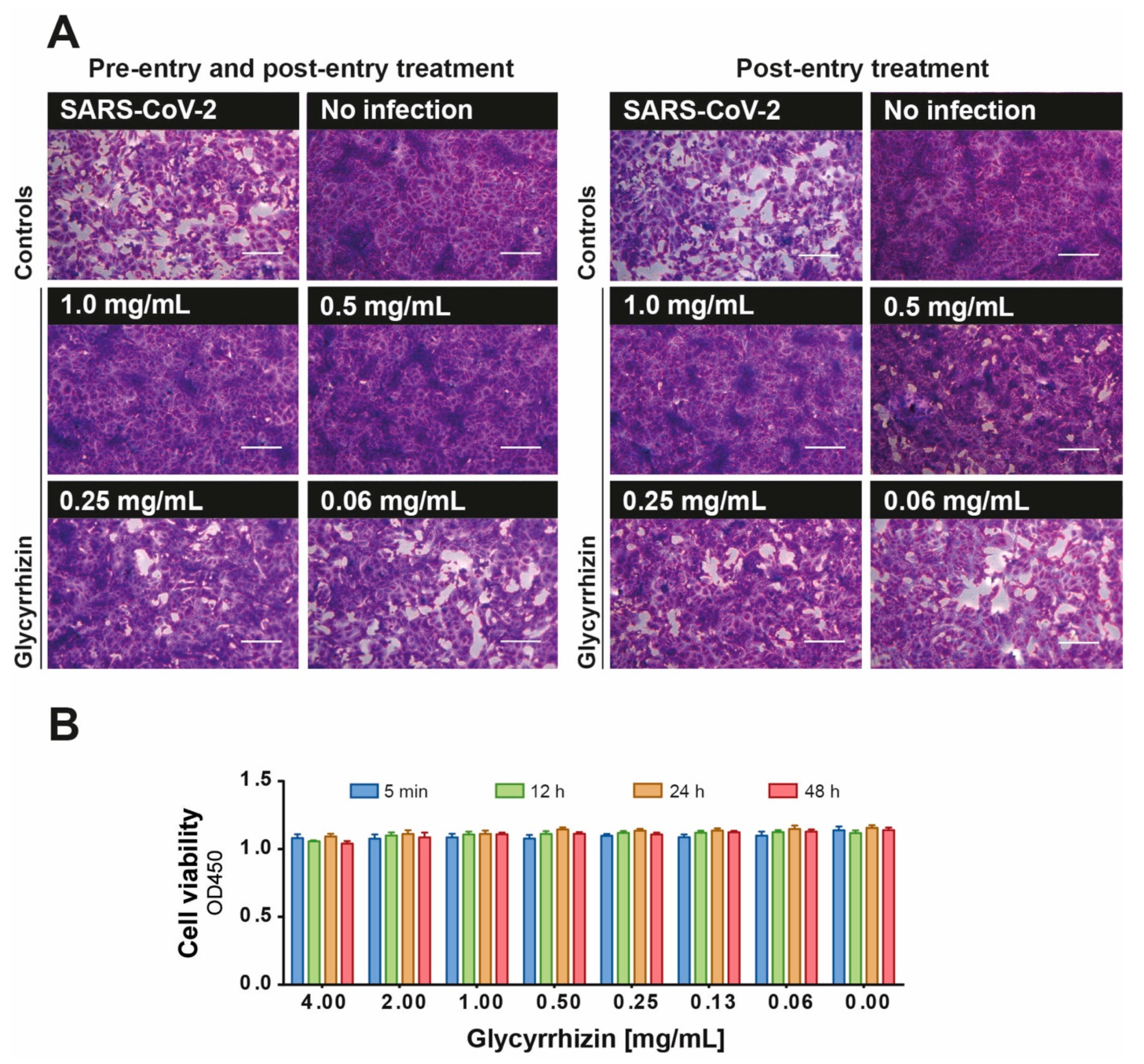
3.2. Effective Inhibition of SARS-Cov-2 Replication by Glycyrrhizin
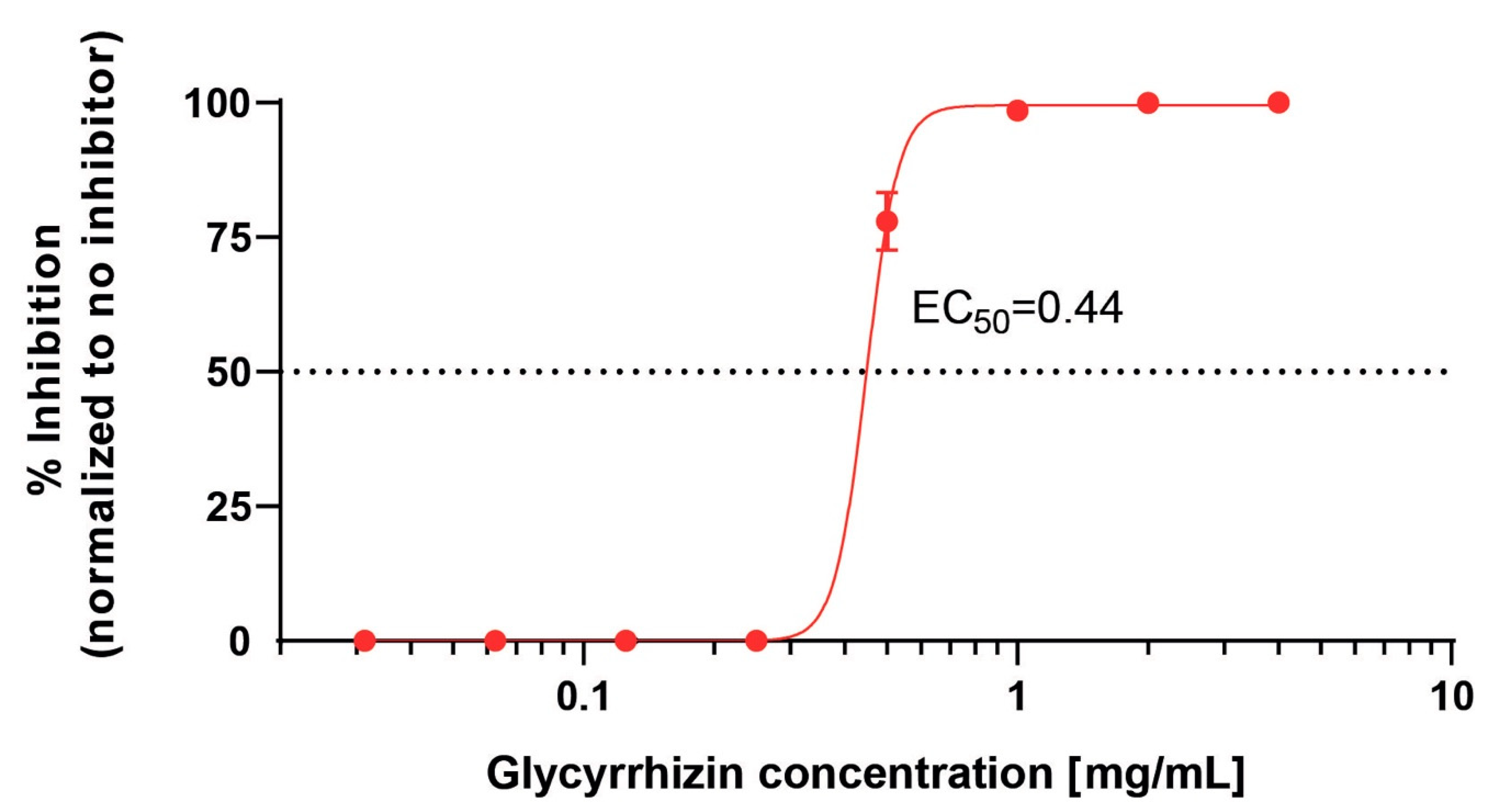
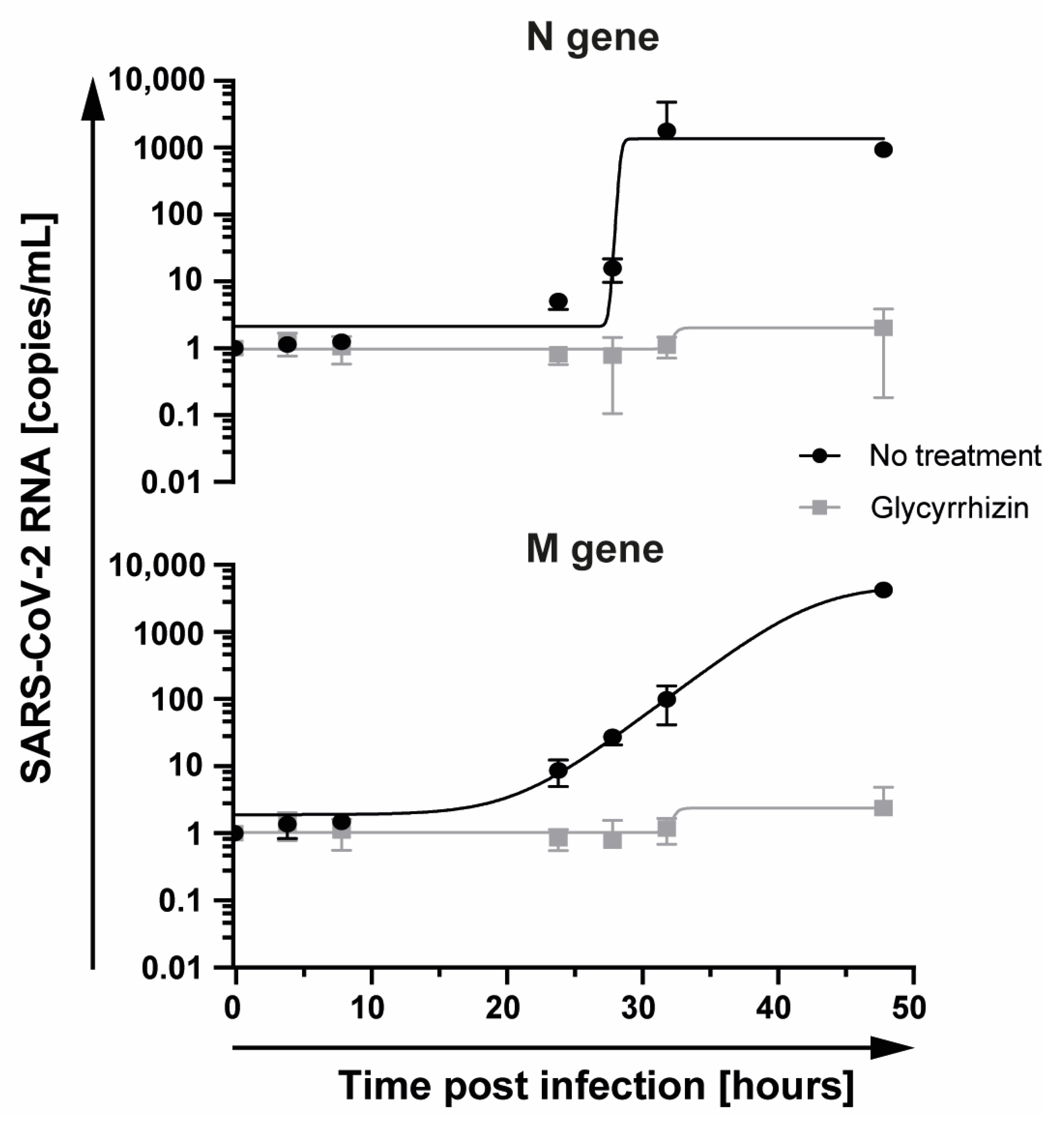
3.3. Quantitative Analysis of the Antiviral Activity of Glycyrrhizin Against SARS-Cov-2
3.4. Inhibition of the Viral Main Protease by Glycyrrhizin
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, M.; Cao, R.; Zhang, L.; Yang, X.; Liu, j.; Xu, M.; Shi, Z.; Hu, Z.; Zhong, W.; Xiao, G. Remdesivir and chloroquine effectively inhibit the recently emerged novel coronavirus (2019-nCoV) in vitro. Cell Res. 2020, 30, 269–271. [Google Scholar] [CrossRef] [PubMed]
- Chrzanowski, J.; Chrzanowska, A.; Graboń, W. Glycyrrhizin: An old weapon against a novel coronavirus. Phytother Res. 2021, 35, 629–636. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Chen, X.; Li, Q.; Li, P.; Zhao, G.; Xu, M.; Xie, P. Inhibition of intercellular adhesion in herpex simplex virus infection by glycyrrhizin. Cell Biochem. Biophys. 2012, 62, 137–140. [Google Scholar] [CrossRef] [PubMed]
- Cinatl, J.; Morgenstern, B.; Bauer, G.; Chandra, P.; Rabenau, H.; Doerr, H.W. Glycyrrhizin, an active component of liquorice roots, and replication of SARS-associated coronavirus. Lancet 2003, 361, 2045–2046. [Google Scholar] [CrossRef] [Green Version]
- Heilingloh, C.S.; Aufderhorst, U.W.; Schipper, L.; Dittmer, U.; Witzke, O.; Yang, D.; Zheng, X.; Sutter, K.; Trilling, M.; MSc, M.A.; et al. Susceptibility of SARS-CoV-2 to UV irradiation. Am. J. Infect Control 2020, 48, 1273–1275. [Google Scholar] [CrossRef]
- Lutter, A.-H.; Scholka, J.; Richter, H.; Anderer, U. Applying XTT, WST-1, and WST-8 to human chondrocytes: A comparison of membrane-impermeable tetrazolium salts in 2D and 3D cultures. Clin. Hemorheol. Microcirc. 2017, 67, 327–342. [Google Scholar] [CrossRef]
- Toptan, T.; Hoehl, S.; Westhaus, S.; Bojkova, D.; Berger, A.; Rotter, B.; Hoffmeier, K.; Cinatl, J., Jr.; Ciesek, S.; Widera, M. Optimized qRT-PCR Approach for the Detection of Intra- and Extra-Cellular SARS-CoV-2 RNAs. Int. J. Mol. Sci. 2020, 21, 4396. [Google Scholar] [CrossRef]
- Srivastava, V.; Yadav, A.; Sarkar, P. Molecular Docking and ADMET Study of Bioactive Compounds of Glycyrrhiza glabra Against Main Protease of SARS-CoV2. Mater. Today Energy 2020, in press. [Google Scholar] [CrossRef]
- Zhang, L.; Lin, D.; Sun, X.; Curth, U.; Drosten, C.; Sauerhering, L.; Becker, S.; Rox, K.; Hilgenfeld, R. Crystal structure of SARS-CoV-2 main protease provides a basis for design of improved α-ketoamide inhibitors. Science 2020, 368, 409–412. [Google Scholar] [CrossRef] [Green Version]
- Morse, J.S.; Lalonde, T.; Xu, S.; Liu, W.R. Learning from the Past: Possible Urgent Prevention and Treatment Options for Severe Acute Respiratory Infections Caused by 2019-nCoV. Chembiochemistry 2020, 21, 730–738. [Google Scholar] [CrossRef]
- Stasi, C.; Fallani, S.; Voller, F.; Silvestri, C. Treatment for COVID-19: An overview. Eur. J. Pharmacol. 2020, 889, 173644. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Wang, B. Randomized clinical trial with two doses (100 and 40 mL) of Stronger Neo-Minophagen C in Chinese patients with chronic hepatitis B. Hepatol Res. 2002, 24, 220. [Google Scholar] [CrossRef]
- van Rossum, T.G.; Vulto, A.G.; Hop, W.C.; Schalm, S.W. Pharmacokinetics of intravenous glycyrrhizin after single and multiple doses in patients with chronic hepatitis C infection. Clin. Ther. 1999, 21, 2080–2090. [Google Scholar] [CrossRef] [Green Version]
- van Gelderen, C.E.; Bijlsma, J.A.; van Dokkum, W.; Savelkoul, T.J. Glycyrrhizic acid: The assessment of a no effect level. Hum. Exp. Toxicol. 2000, 19, 434–439. [Google Scholar] [CrossRef] [PubMed]
- Kwon, Y.J.; Son, D.H.; Chung, T.H.; Lee, Y.J. A Review of the Pharmacological Efficacy and Safety of Licorice Root from Corroborative Clinical Trial Findings. J. Med. Food 2020, 23, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Edelman, E.R.; Butala, N.M.; Avery, L.L.; Lundquist, A.L.; Dighe, A.S. Case 30-2020: A 54-Year-Old Man with Sudden Cardiac Arrest. N. Engl. J. Med. 2020, 383, 1263–1275. [Google Scholar] [CrossRef]
- Luyckx, V.A. Nephrotoxicity of alternative medicine practice. Adv. Chronic Kidney Dis. 2012, 19, 129–141. [Google Scholar] [CrossRef]
- Tsai, T.H.; Liao, J.F.; Shum, A.Y.; Chen, C.F. Pharmacokinetics of glycyrrhizin after intravenous administration to rats. J. Pharm. Sci. 1992, 81, 961–963. [Google Scholar] [CrossRef]
- Yamamura, Y.; Kawakami, J.; Santa, T.; Kotaki, H.; Uchino, K.; Sawada, Y.; Tanaka, N.; Iga, T. Pharmacokinetic profile of glycyrrhizin in healthy volunteers by a new high-performance liquid chromatographic method. J. Pharm. Sci. 1992, 81, 1042–1046. [Google Scholar] [CrossRef]
- Ding, H.; Deng, W.; Ding, L.; Ye, X.; Yin, S.; Huang, W. Glycyrrhetinic acid and its derivatives as potential alternative medicine to relieve symptoms in nonhospitalized COVID-19 patients. J. Med. Virol. 2020, 92, 2200–2204. [Google Scholar] [CrossRef]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Krüger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.-H.; Nitsche, A.; et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020, 181, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Narkhede, R.R.; Pise, A.V.; Cheke, R.S.; Shinde, S.D. Recognition of Natural Products as Potential Inhibitors of COVID-19 Main Protease (Mpro): In-Silico Evidences. Nat. Prod. Bioprospect. 2020, 10, 297–306. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
van de Sand, L.; Bormann, M.; Alt, M.; Schipper, L.; Heilingloh, C.S.; Steinmann, E.; Todt, D.; Dittmer, U.; Elsner, C.; Witzke, O.; et al. Glycyrrhizin Effectively Inhibits SARS-CoV-2 Replication by Inhibiting the Viral Main Protease. Viruses 2021, 13, 609. https://doi.org/10.3390/v13040609
van de Sand L, Bormann M, Alt M, Schipper L, Heilingloh CS, Steinmann E, Todt D, Dittmer U, Elsner C, Witzke O, et al. Glycyrrhizin Effectively Inhibits SARS-CoV-2 Replication by Inhibiting the Viral Main Protease. Viruses. 2021; 13(4):609. https://doi.org/10.3390/v13040609
Chicago/Turabian Stylevan de Sand, Lukas, Maren Bormann, Mira Alt, Leonie Schipper, Christiane Silke Heilingloh, Eike Steinmann, Daniel Todt, Ulf Dittmer, Carina Elsner, Oliver Witzke, and et al. 2021. "Glycyrrhizin Effectively Inhibits SARS-CoV-2 Replication by Inhibiting the Viral Main Protease" Viruses 13, no. 4: 609. https://doi.org/10.3390/v13040609