
Mechanism of Cross-Species Transmission, Adaptive Evolution and Pathogenesis of Hepatitis E Virus


Division of Virology, Department of Infection and Immunity, Jichi Medical University School of Medicine, Tochigi 329-0498, Japan
*
Author to whom correspondence should be addressed.
Viruses 2021, 13(5), 909; https://doi.org/10.3390/v13050909
Submission received: 28 April 2021
/
Revised: 12 May 2021
/
Accepted: 13 May 2021
/
Published: 14 May 2021
(This article belongs to the Special Issue RNA Viruses: Structure, Adaptation, and Evolution)
Abstract
:Hepatitis E virus (HEV) is the leading cause of acute hepatitis worldwide. While the transmission in developing countries is dominated by fecal-oral route via drinking contaminated water, the zoonotic transmission is the major route of HEV infection in industrialized countries. The discovery of new HEV strains in a growing number of animal species poses a risk to zoonotic infection. However, the exact mechanism and the determinant factors of zoonotic infection are not completely understood. This review will discuss the current knowledge on the mechanism of cross-species transmission of HEV infection, including viral determinants, such as the open reading frames (ORFs), codon usage and adaptive evolution, as well as host determinants, such as host cellular factors and the host immune status, which possibly play pivotal roles during this event. The pathogenesis of hepatitis E infection will be briefly discussed, including the special forms of this disease, including extrahepatic manifestations, chronic infection, and fulminant hepatitis in pregnant women.
1. Introduction
Hepatitis E virus (HEV) is a single-stranded positive-sense RNA virus that belongs to the family Hepeviridae [1]. HEV is the leading cause of acute hepatitis around the world. The World Health Organization (WHO) estimated that there are 20 million HEV infections worldwide with approximately 44,000 deaths in 2015 [2]. HEV is known to cause acute hepatitis, rarely leading to fulminant hepatitis. However, chronic hepatitis has increasingly been reported in immunocompromised patients, including organ transplant recipients, as well as patients with hematological malignancy and human immunodeficiency virus (HIV) infection [3,4,5]. Besides manifesting as typical hepatitis, HEV infection can also cause extrahepatic manifestations, including neurological manifestations [6,7], kidney injury, and hematological disorders [8,9].
Among the five known hepatitis viruses, HEV has unique transmission routes ranging from the fecal-oral route, as its classical mode of transmission, and the zoonotic route, to less frequent routes such as via organ transplantation, transfusion of blood or blood products, or vertical transmission from mother to fetus [3,10,11,12,13,14]. Although the overall mortality rate is low, it could reach as high as 30% in infected pregnant women entering their third trimester of pregnancy [14].
HEV infection is globally distributed. In developing countries, HEV has caused multiple outbreaks where the virus is transmitted via the fecal-oral route by drinking contaminated water. Meanwhile, HEV infections in industrialized countries are mainly associated with zoonotic causes [15]. The isolation of the first HEV strain in an animal, in swine [16], opened the door to the discovery of more HEV strains and their variants in animal species. The report by the same research group in the following year showed evidence that some HEV strains recovered from patients with acute hepatitis E are closely related to swine HEV, which can cross the species barrier to infect humans [17,18], proving an HEV zoonosis event.
Isolation of new HEV strains in a growing number of animal species poses a risk to zoonotic infection and has become a public health concern. Despite the inevitable risk of zoonotic infection, many questions surrounding the mechanism of the cross-species transmission in HEV infection remain unanswered and will be discussed in this review, encompassing the viral and host factors involved in the event, such as the viral adaptive evolution and host cellular factors, as some of the determinants of host species tropism. In addition, the pathogenesis of hepatitis E infection will be briefly described in this review, including special forms of the disease, such as chronic infection, extrahepatic manifestations, and the severe cases in pregnant women.
2. Genome and Classification
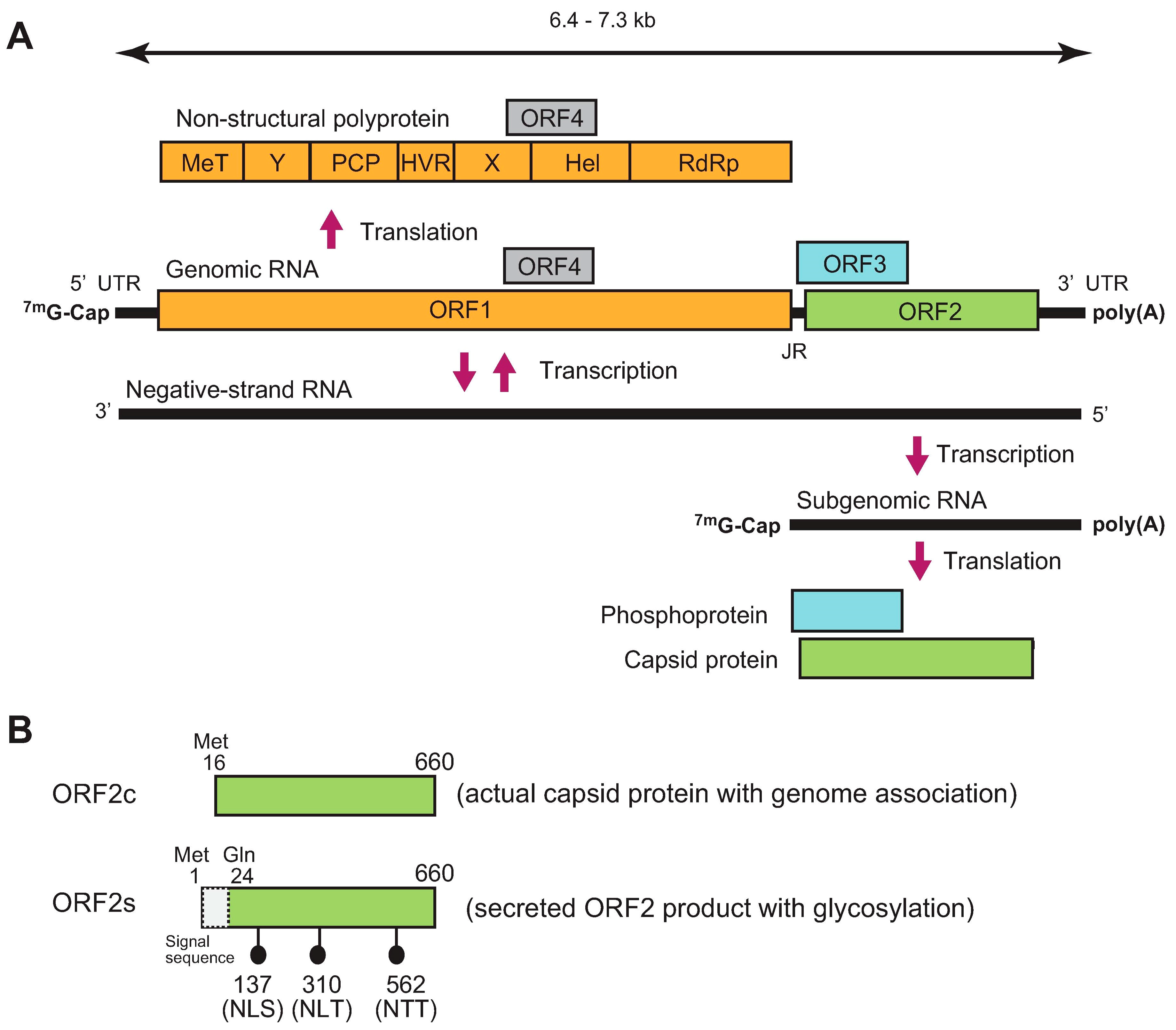
HEV is a single-stranded positive-sense RNA virus. Its genome ranges from 6.4 kilobases (kb) to 7.3 kb with three major open reading frames (ORFs; ORF1–ORF3) (Figure 1A). The genome has a short 5′ untranslated region (5′ UTR) capped at its 5′ end and a short 3′ UTR terminated by the poly(A) tract [19,20]. ORF1 is translated from genomic RNA, while ORF2 and ORF3 are translated from a subgenomic RNA strand [21,22]. ORF1 covers two-thirds of the genome, and encodes a non-structural polyprotein containing multiple functional domains involved in viral replication. However, whether the ORF1 product functions as a single polyprotein or whether it needs to be further processed into smaller units by viral or cellular proteases following translation remains to be elucidated [23,24]. Notably, however, in a recent hemagglutinin (HA)-tagged full-length HEV replicon system, only an uncleaved ~190 kDa ORF1 product was detected [25]. ORF2 encodes capsid protein, which is essential during virion assembly and viral attachment to host cells, and is a major target for neutralizing antibodies [26,27,28]. Recently, it has been reported that HEV produces three forms of ORF2 capsid proteins: the infectious/intracellular ORF2 (ORF2i), which is associated with infectious particles, as well as glycosylated ORF2 (ORF2g) and cleaved ORF2 (ORF2c), which are not associated with infectious particles [29]. Another study determined the initiation codons for the secreted form of the ORF2 product and the actual capsid protein [30]. The secreted form of the ORF2 product (ORF2s) is initiated from the previously presumed start codon (Met 1) and its N-terminal 23 amino acids are cleaved by signal peptidase, while the actual capsid protein (ORF2c) is initiated from an internal AUG (Met 16) located 16 codons downstream of the first AUG (Figure 1B). Between the two studies, functionally, the ORF2s and ORF2c may correspond to ORF2g and ORF2i, respectively, with the absence of the cleaved form ORF2c, presumably due to its lower level [29]. ORF3 encodes a multifunctional phosphoprotein required for virion egress [31,32,33] and has been reported to be a functional ion channel, as viroporin [34].
Exclusively in genotype 1 HEV (HEV-1), ORF4 codes for a novel protein identified in the coding sequence of ORF1. It is synthesized only under the condition of endoplasmic reticulum (ER) stress and is a short-lived protein. ORF4 protein is reported to enhance the replication of HEV-1 by promoting the viral RNA-dependent RNA polymerase (RdRp) activity of ORF1 [35]. A recent report demonstrated that ORF4 provided in trans enhances the viral replication of genotype 3 HEV (HEV-3) although it does not naturally encode ORF4 in its genome [36].
HEV is a member of family Hepeviridae [1], which has been identified in humans as well as a broad range of animal species [15]. This family is divided into genus Orthohepevirus which consists of HEV strains from mammals and birds, and genus Piscihepevirus with its single species, Piscihepevirus A. The genus Orthohepevirus consists of species Orthohepevirus A, Orthohepevirus B, Orthohepevirus C, and Orthohepevirus D. To date, there are eight genotypes within Orthohepevirus A species which infect humans, pigs, wild boars, rabbits, deer, camels, and other animal species. Species Orthohepevirus B is only identified in birds, species Orthohepevirus C is identified in rodents, while species Orthohepevirus D is identified in bats [1].
Orthohepevirus A has been assigned to eight distinct genotypes (HEV-1–8) [1]. While HEV-1 and HEV-2 have only been isolated in humans, HEV-3 and HEV-4 have been identified in humans as well as several animal species, including pigs, wild boars, deer, and rabbits [37,38,39,40,41,42]. HEV-5 and HEV-6 have only been detected from wild boars in Japan [43,44,45,46]. HEV-7 has been isolated in dromedary camels and a human [47,48,49]. Finally, HEV-8 has been detected in Bactrian camels [50,51]. In addition, there are many as-yet unclassified HEV strains identified from moose [52,53], tree shrew (GenBank accession number KR905549), sparrow [54], silkie fowl [55], little egret [56], hamster [57], voles [58,59], and various bats [60].
3. Modes of Transmission
In developing countries, HEV is mainly transmitted via the fecal–oral route and has been associated with multiple outbreaks due to poor sanitary conditions (e.g., contaminated drinking water). Meanwhile, sporadic cases and clusters in industrialized countries are often reported where the routes of transmission are more variable and are dominated by zoonotic infection [6,61,62,63,64,65]. Although less frequent than the two main transmission routes, HEV can also be transmitted through organ transplantation or the transfusion of blood or blood products where it can cause chronic HEV infection or from an infected mother to the fetus [3,10,11,12,13,14].
Zoonotic HEV is mainly transmitted via foodborne routes, such as from the consumption of undercooked meat or milk products of infected animals. Another possible route is associated with occupational risk among workers, including veterinarians, forestry workers, slaughterhouse workers, or animal farm workers [66,67,68] as they are continuously in direct contact with probably infected animals.
HEV is excreted in feces where the viral particles can reach the environment and contaminate water sources and irrigated food. HEV in environmental samples can reach the food chain (e.g., in mussels and contaminated pork products, such as sausages or meat) [69]. Besides being detected in packaged sausages and liver samples from processing sites and supermarkets in Spain, Italy, and Czech Republic, HEV is also detected from environmental samples collected in production farms and processing plants and at the points of sale of items such as knives, flooring, belt surfaces, workers’ hands, and toilets [69,70]. Foods other than pork products have also been implicated, where HEV is isolated from shellfish, fruits, and vegetables [71]. This is likely due to pig slurry contaminating watercourses or being used as fertilizer [72]. Coastal water may be contaminated with HEV, leading to the accumulation of the virus in the digestive tissue of shellfish due to filter-feeding [69]. An outbreak of hepatitis E on a cruise ship was linked to the consumption of shellfish while on board [73]. HEV has also been detected in sewage and run-offs, where the virus is concentrated in waste products generated during sewage and drinking water treatment. The following land application of the waste can cause the contamination of water in aquifers, can contaminate irrigated vegetables, and can therefore be a risk factor for HEV infection [69,74,75,76]. Another transmission route that warrants further exploration is through contact with an environment contaminated with the droppings or body fluid of infected animals.
4. Pathogenesis
The HEV particles in bile and those shed in the feces are non-enveloped. Meanwhile, the particles in circulating blood and culture supernatants are in membrane-associated form, which is different from classical enveloped viruses due to the absence of viral glycoproteins in the surrounding lipid bilayer in which membrane-associated HEV particles seem to be completely covered with a lipid membrane [31,77,78]. These findings have led to the designation of the membrane-associated form of HEV as quasi-enveloped HEV [79,80]. Although a primary site of HEV propagation in humans remains undetermined, it is tempting to speculate that HEV replicates primarily in the liver (not in the gut) and that the non-enveloped virions entering via the gastrointestinal tract will be neutralized by immune sera before reaching the liver via the portal vein. This can explain how humans who were immunized with a recombinant ORF2-based vaccine can be protected against infection with enterically-transmitted HEV [78,81].
Previously, it was hypothesized that lipid-associated HEV is released from both infected cultured cells (in vitro) and infected hepatocytes (in vivo) as lipid-associated virions, accompanied by ORF3 protein, and that the quasi-envelope is dissociated from the virion after shedding the lipid in the bile duct, which contains detergent (deoxycholic acid), and shedding the ORF3 protein in the duodenum, which contains protease (trypsin) secreted from the pancreas [82,83,84]. In support of this hypothesis, Capelli et al. [85] utilized F2 cells (subclone of the human hepatocarcinoma cell line HepG2/C3A, which grew as a polarized monolayer culture and had better HEV production than the mother cell line) in a cell culture due to the polarized nature of hepatocytes, which have exosomal pathways (basolateral, oriented toward the blood; and apical, oriented toward the bile). In this study, they showed that irrespective of the virus form (naked or membrane-associated), and whether HEV-3 or HEV-1 is used as inoculum, the polarized cells released around 95% of the HEV RNA from their apical sides as lipid-associated particles. The ratios of infectious particles to HEV RNA copies on the apical were higher in comparison to the basolateral side, suggesting that the apical pathway is the main release route and that the majority of infectious HEV particles are released from the bile sides of hepatocytes while the small fraction released into the bloodstream could spread HEV throughout the host [85].
Although most HEV infections are self-limiting, infections in immunosuppressed patients may progress into chronic disease or may cause extrahepatic manifestations [6,7,8,9,86]. The host innate immune system is the first line of defense against viral infection. Dysregulation of this system can cause severe pathogenesis. In HEV infection, host innate immunity has shown to involve an active response in experimental models and patients [87,88,89,90,91]. HEV has also developed strategies to counteract the host immune system where recent knowledge mainly outlines the interactions between HEV viral proteins with the host innate immunity, as reported for ORF1 (e.g., the papain-like cysteine protease (PCP) and X domains of ORF1 inhibit the activation of retinoic acid-inducible gene I (RIG-I) and TANK-binding-kinase 1 (TBK-1), as well as the phosphorylation of interferon regulatory factor 3 (IRF3)) [92], ORF2 (e.g., ORF2 suppresses nuclear factor-kappa B (NF-κB) activity by blocking the ubiquitination of inhibitor (IκBα) of NF-κB alpha) [93], and ORF3 (e.g., ORF3 inhibits the phosphorylation of signal transducer and activator of transcription 1 (STAT1) and the expression of interferon-stimulated gene (ISG) upon HEV infection) [94].
Extrahepatic manifestations reported in HEV-infected patients are possibly associated with either direct viral effects due to cytopathic tissue damage caused by extrahepatic replication in affected tissues, or with indirect effects related to the immunological processes induced by a cross-reactive host immune response to HEV [95]. Besides being detected in various tissues in vivo, the ability of HEV to cause extrahepatic manifestations has also been demonstrated in vitro, where HEV can infect a range of cell types (e.g., human lung epithelial cells, human colon epithelial cells, human neuronal-derived cells, and human placental cells) [95,96].
HEV infection is associated with a high mortality rate in pregnant women, particularly those in their third trimester of pregnancy, where the highest incidence is reported for HEV-1 [14,97,98]. The alteration of the immune system during hormonal changes in pregnancy has been reported as a potential pathogenic mechanism of HEV infection, which is related to cellular immunity [14,99]. A recent study by Gong et al. [100] indicated that HEV infection significantly inhibits the signaling pathways of cyclic adenosine 3′, 5′-monophosphate/protein kinase A/response element-binding protein (cAMP/PKA/CREB) and phosphatidylinositol 3-kinase/protein kinase B/mammalian target of the rapamycin (PI3K/AKT/mTOR). The increasing estrogen levels and high activation of estrogen receptor alpha (ER-α) during pregnancy aggravates HEV replication. The exacerbation of HEV replication inhibits the expression of ER-α and suppresses the cAMP/PKA/CREB and PI3K/AKT/mTOR signaling pathways [100]. Another factor that might enhance the virulence of HEV in pregnant women is the presence of the ORF4 protein, which is exclusively found in HEV-1. The ORF4 protein is only synthesized under the condition of ER stress and is essential to stimulate the RdRp of HEV-1, which causes enhanced virus replication [35]. Since pregnancy can induce ER stress and fulminant hepatitis E in pregnant women is mostly caused by HEV-1, it can be hypothesized that ER stress caused by pregnancy can induce the synthesis of ORF4, thereby causing higher viral replication (in comparison to non-pregnant patients) and consequently leading to the development of fulminant HEV infection.
Most chronic HEV infections are associated with HEV-3, with a single case of HEV-1 [101] or HEV-7 [49], which to date, are only anecdotal and unique; and HEV-4, which has been described in small numbers of reports [102]. Viral and host factors are likely the determinants of the chronicity of HEV infection [103]. Viral factors, such as genotype, zoonotic potential, specificity, and adaptability to a host play important roles in the establishment of persistent infection. Meanwhile, host factors, such as the immune state and the nutritional health of the patient can also determine the course of HEV infection. The type of immunosuppressive drugs is reported to affect the progression to chronic infection, with patients treated with calcineurin inhibitors (e.g., tacrolimus, cyclosporin A) or mTOR inhibitors (e.g., everolimus, sirolimus), reported to be more prone to persistent HEV infection [3,103].
5. Zoonotic HEV Infection
HEV infection caused by zoonotic HEV-3 or HEV-4 has increased in industrial countries, including Japan and several European countries such as France, the Netherlands, Spain, and UK [69,104,105,106]. It is also found to be endemic in the US population and circulates abundantly among North American pig herds [37,106]. Zoonotic transmission is responsible for the sporadic cases and clusters of human infection caused by HEV-3 and HEV-4, and chronic infection reported in immunocompromised patients is mostly linked to zoonotic HEV-3 [37].
Swine are the primary reservoir of HEV; in swine, infection is subclinical [107]. Most animal strains of HEV have been isolated from swine. Soon after the report of the first known zoonotic infection related to HEV [17], the number of zoonotic HEV infections continued to increase, particularly in industrialized countries. Within species Orthohepevirus A, HEV strains of five genotypes (HEV-1, -2, -3, -4, and -7) are capable of infecting humans (Table 1). While HEV-1 and HEV-2 are restricted to humans, HEV-3, -4, and -7 are capable of crossing the species barrier to cause zoonotic infection in humans. Besides HEV-3 and HEV-4 strains from pigs, wild boars, and deer, variants of HEV-3 in rabbits were also found to be capable of causing human infection [42,108].
Under experimental conditions, swine HEV-3 and HEV-4 infected rhesus monkeys and, conversely, HEV-3 and HEV-4 of human origin infected pigs [17,109,110]. However, HEV-1 and HEV-2 failed to infect pigs under experimental conditions [18], indicating that HEV-1 and HEV-2 have a limited host range [111].
To date, only a single case of human infection related to HEV-7 has been reported. It is a chronic case in an immunocompromised patient caused by the regular consumption of milk and meat products of dromedary camels [49]. The risk of zoonotic infection was also suggested in HEV-5 (from wild boar) and HEV-8 (from a Bactrian camel) as they were successfully transmitted to cynomolgus macaques in experimental settings [112,113,114]. Besides the members of species Orthohepevirus A, rat HEV, which is a member of species Orthohepevirus C, is also capable of infecting humans, as was recently reported in both immunocompromised and immunocompetent patients [115,116,117]. With growing numbers of isolated rat HEV strains from around the world [118], as well as evidence of human infection by rat HEV [115,116,117]—in limited areas—the number of cases of human infection by rat HEV may increase in the future.

{kind=link}
{kind=link}
{kind=link}
Table 1.
Reported cross-species transmission of genus Orthohepevirus.
| Species | Genotype | Experimental Infection | Zoonotic Potential |
|---|---|---|---|
| Orthohepevirus A | Human HEV-1 | Non-human primate [119,120] | No |
| Human HEV-2 | No | ||
| Human HEV-3 | Rabbit [121] | ||
| Swine HEV-3 | Non-human primate [17] | Yes [16,122,123,124,125,126] | |
| Wild boar HEV-3 | Pig [127,128] Rabbit [129] | Yes [38,130,131,132] Pig [127,133] | |
| Deer HEV-3 | Yes [39] | ||
| Rabbit HEV-3 | Pig [134,135] Non-human primate [135] | Yes [42,108] | |
| Human HEV-4 | Pig [110] | ||
| Swine HEV-4 | Non-human primate [109,136] Rabbit [137,138] Mongolian gerbil [139,140] | Yes [123,141,142,143] | |
| Wild boar HEV-4 | Yes [144,145] | ||
| Wild boar HEV-5 | Non-human primate [112] | Likely | |
| Wild boar HEV-6 | Unknown | ||
| Dromedary camel HEV-7 | Non-human primate [113] | Yes [49] | |
| Bactrian camel HEV-8 | Non-human primate [114] | Likely | |
| Orthohepevirus B | Avian HEV | Turkey [146,147] | Unlikely |
| Orthohepevirus C | Rat HEV | Yes [115,116,117] | |
| Orthohepevirus D | Bat HEV | Unlikely |
6. Mechanism of Cross-Species Transmission in HEV Infection
The key event in zoonosis is when an animal virus starts replicating in the first human subject. Here, the virus will experience the selective environment of the human body, possibly rendering viral adaptation and refinement for humans [148]. The processes responsible for cross-species transmission and the emergence of viruses in natural systems are diverse, involving ecological, evolutionary, and genetic factors. Several evolutionary mechanisms have been reported [149,150,151,152]. Among them, molecular adaptation by natural selection is an important occurrence that requires the generation and spread of beneficial mutations to increase virus fitness in a specific environment [153]. Viruses with frequent cross-species transmission events might exhibit parallel evolution (independent evolution of the same genotype or phenotype from distinct ancestors) as a result of adaptation to new host environments [153].
One of the efforts to demonstrate the broad range of HEV hosts was reported by Shukla et al. [154]. HEV-3 Kernow-C1 strain was semi-purified from the feces of a chronic hepatitis E patient co-infected with HIV [155]. Other than infecting human hepatoma cells (HepG2/C3A) and pig kidney cells (LLC-PK1), the fecal virus was surprisingly able to infect a variety of non-primate cells, including deer, cow, mouse, chicken, cat, dog, and rabbit cells. Meanwhile, the HEV-1 strains (Sar-55 and Akluj) used in this study successfully infected not only HepG2/C3A cells but also LLC-PK1 cells, albeit less efficiently. The HEV-3 Kernow-C1 strain was also serially passaged six times (p6). The p6 virus was shown to infect LLC-PK1 more efficiently than it did HepG2/C3A, suggesting that despite adaptation to grow in human HepG2/C3A cells, it still infected more pig cells than human cells [154].
The precise mechanism of cross-species transmission in HEV infection is not fully understood. The establishment of cross-species transmission requires the interaction of viral determinants and host determinants. Within the past decade, progress has been made regarding the viral and host determinants involved in the cross-species transmission of HEV, among which the HEV ORFs and evolutionary events have become the most studied to date (Table 2).
6.1. The HEV ORFs
When the ORFs were analyzed separately, ORF1 and ORF3 were demonstrated to more efficiently separate the HEV strains into anthropotropic and enzootic genotypes, suggesting their evolutionary role in host species tropism [157,158,162]. Consistent with the broad host range of enzootic HEV genotypes, the HVR of ORF1 in HEV-3 and HEV-4 is around two-fold more heterogenous than that in HEV-1 [162], and the analysis of the relative synonymous codon usage values of ORF3s showed that the HEV strains were grouped into only two clusters consisting of the HEV-1 and HEV-2 strains (anthropotropic genotypes), and a cluster of the remaining strains (HEV-3–8, enzootic genotypes) [158]. On the other hand, ORF2 was previously thought to be the viral determinant of host tropism, since it encodes the capsid (the only structural protein) that possibly binds to unknown host cell receptors [141,163,164,165].
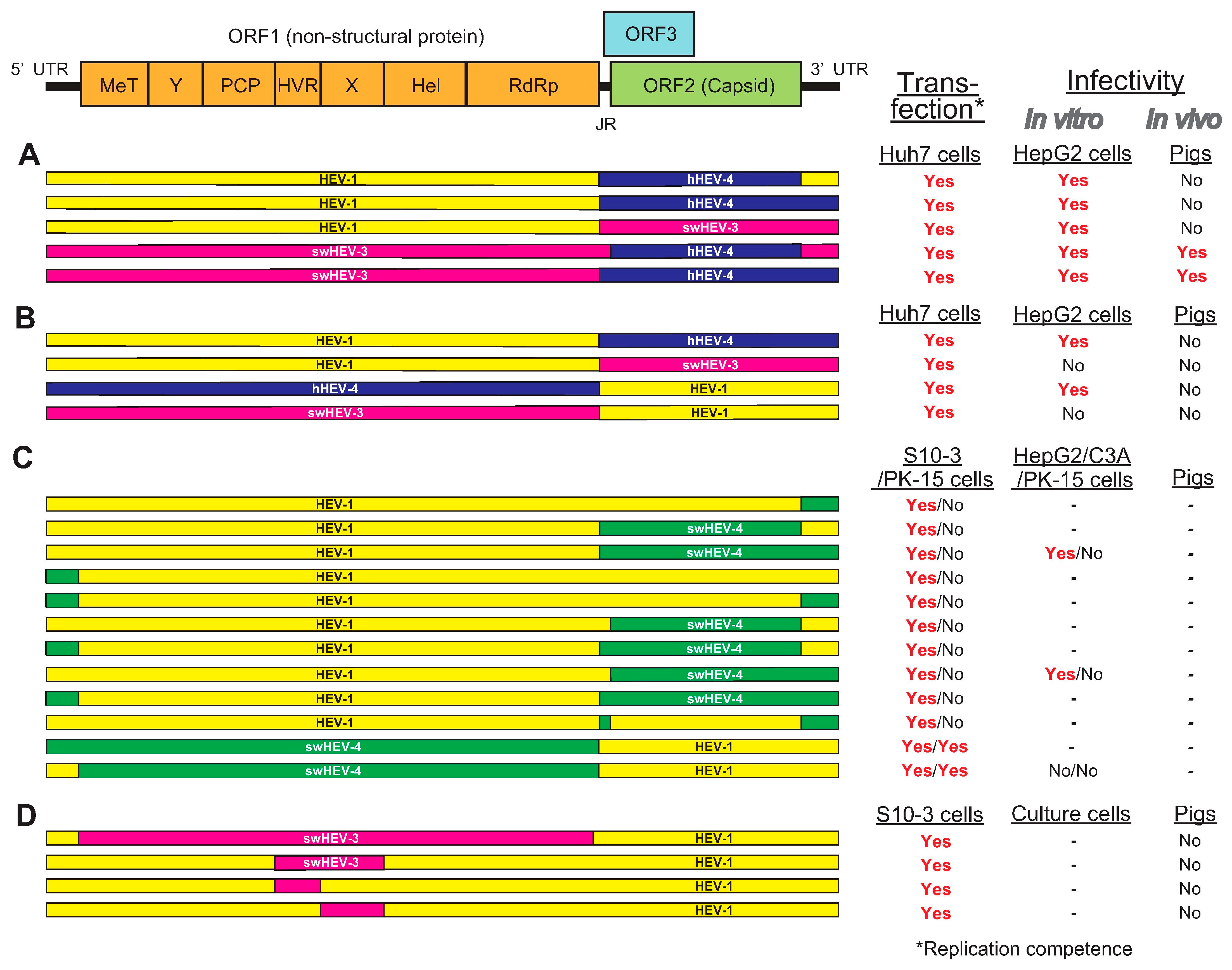
To unveil the viral determinant of species tropism in HEV cross-species transmission, several research groups have been utilizing the intergenotypic chimeric virus strategy. The potential role of ORF2 in HEV cross-species infection was investigated by Feagins et al. [111] and Cordoba et al. [161]. Feagins et al. [111] constructed intergenotypic chimeric viruses by swapping the ORF2 capsid gene, either alone or in combination with its adjacent junction region (JR) and the 3′ UTR, between HEV-1 and human HEV-4, swine HEV-3 and human HEV-4, and HEV-1 and swine HEV-3 (Figure 2A). Their infectivity was tested in vitro and in pigs. Three chimeric viruses containing the JR, ORF2, and 3′ UTR of swine HEV-3 or human HEV-4 in the backbone of HEV-1 failed to infect pigs. However, the other two chimeric viruses containing the ORF2 capsid gene either alone or in combination with its adjacent 5′ JR and the 3′ UTR from human HEV-4 in the backbone of swine HEV-3 were replication-competent in Huh7 cells and infectious in both HepG2/C3A cells and pigs, suggesting that the 5′ UTR and ORF1 may be involved in cross-species transmission [111].
Following this, Cordoba et al. [161] constructed four intergenotypic chimeric clones by swapping the ORF2 gene along with its JR, ORF3, and 3′ UTR between HEV-1 and HEV-4 of human origin, and HEV-1 and swine HEV-3 (Figure 2B). Only two chimeras with sequences swapped between HEV-1 and HEV-4 of human origin successfully established infection in HepG2/C3A cells. The inability of chimeras with sequences swapped between HEV-1 and swine HEV-3 to produce infectious viral particles may be due to incompatibility of viral genes from HEV-1 with those from swine HEV-3. The cDNA clone of human HEV-4 used in this study was confirmed to be capable of infecting pigs. However, none of the four chimeras were able to establish robust infection in pigs [161].
It was demonstrated that ORF2 and ORF3 are not involved in HEV cross-species infection, as evidenced by the failure of host range expansion of chimeric viruses in which the ORF2 or ORF3 gene was swapped [111,161], suggesting that other genomic regions (5′ UTR and ORF1) may play a role in determining the HEV host tropism. Chattarjee et al. [156] reported the construction of 12 different HEV-1–HEV-4 chimeric full genome clones with HEV-1 as the backbone, where the structural (ORF2 and ORF3), non-structural (ORF1), or UTRs were replaced by the corresponding segments from swine HEV-4 clone (Figure 2C). Although all chimeric clones were able to replicate in S10-3 of human hepatoma cells, only two chimeric clones (HEV-1 as the backbone where the ORF1 either alone or in combination with its 5′ UTR was swapped with those of swine HEV-4) were able to replicate in PK-15 of pig kidney cells. This shows the crucial role of the ORF1 polyprotein in crossing the species barrier at the cellular level [156].
Although the available evidence suggests that ORF1 may contribute in HEV cross-species transmission, the exact region within ORF1 that is essential in determining the host tropism is still unknown. Insertion and/or deletion in ORF1 have been demonstrated to affect not only virus replication but also host tropism [166,167]. The insertion of the host ribosomal protein sequence (RPS17) within the HVR of ORF1 led to expanded host adaptability, where the RPS17-inserted mutant was able to establish infection in cell lines of multiple animal origins in vitro [154,166], suggesting a potential role of ORF1, particularly HVR, in interspecies infection.
To determine which viral genetic element in the ORF1 is essential in determining host tropism, Tian et al. [168] utilized genomic backbone of HEV-1 which only infects humans to construct a panel of intergenotypic chimeras in which the entire ORF1 gene or its functional domains were swapped with the corresponding regions from swine HEV-3, which infects both humans and pigs (Figure 2D). Although the chimeric HEVs were replication-competent in Huh7 (S10-3) cells (a human liver cell line) and swine HEV-3 parental clone-inoculated pigs developed viremia, the RNA transcripts subsequently inoculated intrahepatically into pigs did not confer the ability to infect pigs in the chimera; this can be partially linked to the limitation of this study, where the HEV-1 backbone used here technically has low virus replication efficiency [168]. The results from this study may also suggest that—in addition to the unidentified viral genetic element within ORF1—determinants (e.g., host factors) may play a role in host tropism, either alone or in association with the viral genetic element itself. In addition, Tian et al. [168] also investigated the role of the human ribosome protein sequence S17, which expanded the host range in cultured cells [154,166], in HEV cross-species infection. The results demonstrated that the insertion of S17 in the HVR of ORF1 did not abolish HEV replication competency in vitro and that it did not expand HEV host tropism in vivo, suggesting that S17 is not a determinant factor for HEV cross-species infection in vivo [168].
Figure 2.
A schematic diagram of the genomic organization of HEV and its intergenotypic chimeric constructs summarized from four relevant papers. (A) Intergenotypic chimeric viruses where the ORF2 capsid gene—either alone or in combination—was swapped with its adjacent JR and 3′ UTR, between HEV-1 and human HEV-4, swine HEV-3 and human HEV-4, and HEV-1 and swine HEV-3 [111]. (B) Intergenotypic chimeric viruses where the ORF2 gene along with its JR, ORF3, and 3′ UTR were swapped between HEV-1 and human HEV-4, and between HEV-1 and swine HEV-3 [161]. (C) Twelve different HEV-1–swine HEV-4 intergenotypic chimeric viruses with HEV-1 as the backbone where ORF2, ORF3, ORF1, or UTRs were replaced by corresponding segments from swine HEV-4 [156]. (D) The genomic backbone of HEV-1 was used to construct a panel of intergenotypic chimeras in which the entire ORF1 gene or its functional domains were swapped with the corresponding regions from swine HEV-3 [168].
Figure 2.
A schematic diagram of the genomic organization of HEV and its intergenotypic chimeric constructs summarized from four relevant papers. (A) Intergenotypic chimeric viruses where the ORF2 capsid gene—either alone or in combination—was swapped with its adjacent JR and 3′ UTR, between HEV-1 and human HEV-4, swine HEV-3 and human HEV-4, and HEV-1 and swine HEV-3 [111]. (B) Intergenotypic chimeric viruses where the ORF2 gene along with its JR, ORF3, and 3′ UTR were swapped between HEV-1 and human HEV-4, and between HEV-1 and swine HEV-3 [161]. (C) Twelve different HEV-1–swine HEV-4 intergenotypic chimeric viruses with HEV-1 as the backbone where ORF2, ORF3, ORF1, or UTRs were replaced by corresponding segments from swine HEV-4 [156]. (D) The genomic backbone of HEV-1 was used to construct a panel of intergenotypic chimeras in which the entire ORF1 gene or its functional domains were swapped with the corresponding regions from swine HEV-3 [168].

6.2. Adaptive Evolution and the Codon Usage of HEV
The common ancestor of the modern HEV strains emerged approximately 6800 years ago, following the domestication of pigs and intensification of agriculture. Domestication caused adaptive changes in the common ancestor leading to the emergence of both human-restricted (HEV-1 and HEV-2) and zoonotic (HEV-3 and HEV-4) strains [158]. An analysis performed before the identification of the wild boar- and camel-derived genotypes (HEV-5 to HEV-8) indicated that the ancestor of all Orthohepevirus A was enzootic [169]. The same study also mentioned that the ancestors of HEV may have adapted to a succession of animal hosts, leading to humans. In contrast, a recent study suggested that the Orthohepevirus A species originated from humans and subsequently evolved into the zoonotic and human-restricted genotypes [170]. However, this study group also mentioned that their inference of an ancestral human host for extant Orthohepevirus A strains did not exclude the possibility that humans might have acquired HEV through cross-species transmission from other animals.
Synonymous codon usage is the non-random selection of frequently used codons. This type of selection is limited by codon bias for different genes. Determinant factors for synonymous codon usage bias include natural selection, mutational pressure, and translational efficiency, as well as compositional constraints of the mammalian genome [160,171]. It is widely known that the synonymous codon usage in viral coding sequences serves as an important mediator in viral adaptation to the host [172]. Both restricted and broad host HEV infection can provide strong natural selection leading to the shaping of the codon usage pattern of different genotypes of HEV. Viral adaptation to the host in codon usage is a long-term evolutional feedback, which is derived from the host-range of HEV infection [159].
Codon adaptation may be the essential factor in determining the viral host tropism [158]. The evolving process embodies coherent patterns of synonymous codon usage that have been established between the virus and the host. This process assists the virus in adapting to susceptible hosts and expanding the scope of zoonosis not only by specifying amino acid usage and translation dynamics but also by controlling gene expression levels [173,174].
Natural selection was identified as a major factor affecting codon usage patterns in HEV ORFs that may explain the wide range of HEV hosts, while still being influenced by mutational pressure as part of the evolution process [157,158]. In a correspondence analysis based on relative synonymous codon usage data, different HEV genotypes appeared to cluster (in particular, HEV-1, and HEV-3 and HEV-4). The analysis showed that—based on ORF1—HEV-1 is clearly separated from the other groups [160]. Considering that ORF1 encodes an essential non-structural polyprotein that controls the life cycle of HEV, these results may partially reflect the fact that HEV-1 is restricted to human hosts, while the HEV-3 and HEV-4 strains were found in various animal species with the capability of cross-species transmission [160]. Factors such as natural selection and mutational pressure enabled significant adaptation changes in the ORF1 of HEV-1 to humans, rendering ORF1 the evolutionary indicator of host speciation [157,158].
The genotype-specific codon usage pattern that results from the balance between mutational and natural selection provides an evolutionary pathway where HEV-1—with neutral evolution caused by synonymous mutations—is able to sustain its stable genetic characterization by strong natural selection. In contrast, the synonymous codon usage patterns of HEV-3 and HEV-4 may not be caused by neutral evolution. Obvious divergence of codon usage bias in a four-fold degenerate codon family was identified in HEV ORFs between HEV-1, and HEV-3 and HEV-4 [159]. The genotype-specific codon usage bias in HEV-1 is generally stronger than that of HEV-3 and HEV-4 [159]. Unlike the unique codon usage pattern of HEV-1, the HEV-3 and HEV-4 strains derived from either humans or swine have more diverse codon usage patterns in the ORFs. These can serve as evolutionary monuments and may explain the transmission from swine to human [159].
In a study by Bouquet et al. [157], although codon preferences of humans and swine are very close, a lower codon usage bias was observed for zoonotic HEV (HEV-3 and HEV-4). The codon adaptation index (CAI) in this study was calculated with the general codon usage table of humans and swine. The index is >0.5, which indicates good adaptation of HEV to its hosts; thus, it can be assumed that the gene expression of HEV of human and zoonotic genotypes is very well adapted to the translational kinetics in humans [157]. In another study, Shukla et al. [154] inoculated Sar-55 of HEV-1 and the Kernow-C1 of HEV-3 onto deer cells to study host-range restriction. Immunofluorescence staining of ORF2 protein showed that in comparison to Kernow-C1, Sar-55 was deficient in the production of ORF2 capsid protein. The introduction of the first 29 nucleotides of Kernow-C1 at the translation initiation site increased the ORF2 production of Sar-55 in deer cells, suggesting that the modulation of translation from closely spaced codons can differ significantly according to host species, and this difference may provide one mechanism for restricting the host range [154].
Synonymous codon usage patterns of HEV that are constantly modified by mutation pressure and natural selection from the host are important for the zoonotic nature of this virus. Various synonymous codon usage patterns of HEV strains may reflect the viral requirement of evolutionary adaptation to the host.
6.3. Interaction between HEV and the Host Cell
One of the major obstacles to interspecies virus infection is the difference between cell-receptor sequences. The primary structure of the receptor protein can affect the host susceptibility to cross-species viral infections. A virus can only adapt to a new host if the similarity between the receptor proteins present in the potential host species is high enough for them to cross the species barrier [175].
Receptor differences—either quantitative or qualitative—offer an alternative explanation for differences in host range. Supporting the hypothesis of receptor-determined host range, Shukla et al. [154] reported that the fecal Kernow-C1 p6 virus maintained a higher titer in pig cells than in human cells even though it was adapted to grow in human cells [154]. They also stated that there are 54 amino acid differences in the capsid protein between Sar-55 of HEV-1 and Kernow-C1 of HEV-3, while there are only five between the fecal and p6 Kernow-C1 strains, suggesting that the adapted virus may have retained the receptor-interacting specificity of the fecal virus [154].
Nguyen et al. [176] compared the ability of HEV-1 and HEV-3 to infect or transfect cultured human HepG2/C3A cells and swine LLC-PK cells. Immunofluorescence microscopy showed that the HEV-1 isolate infected disproportionately fewer swine cells than human cells. This study group also utilized the virus replicons of HEV-1 and HEV-3 containing the Gaussia luciferase gene in place of the viral ORF2 gene and showed that the luciferase expression from the HEV-1 replicon is limited and may reflect the kidney cell origin of the swine LLC-PK cells used in the study. When a kidney cell line of rhesus macaques was used for comparison, the luciferase expression patterns demonstrated that the translation of the ORF2 capsid gene of HEV-1 is inhibited in swine kidney cells in comparison to its translation in rhesus macaque kidney or human liver cells, suggesting that this virus may produce insufficient capsid protein for optimal assembly in swine cells. The severely restricted ability of both HEV-1 and its intergenotypic chimera (consisting of 450 nucleotides encoding the putative receptor-binding region of HEV-3 in place of the corresponding region of HEV-1) to infect swine kidney cells in comparison to their ability to infect human hepatoma cells supports the notion that amino acids 456 to 605 of the virus capsid protein encompass the receptor-binding region and suggests that HEV-1 may be prevented from infecting swine cells due to the absence of a suitable cellular receptor or co-receptor [176].
Although the ORF2 capsid protein was thought to play an important role in HEV attachment and entry [177], earlier studies utilizing HEV-1 as the backbone, where the ORF2 capsid gene was replaced with that of swine HEV-3 or human HEV-4, showed that HEV-1 did not acquire the ability to infect pigs [111,161]. In fact, binding to cells followed by virus entry is only the first step in virus replication. After entering the cell, the virus must interact correctly with host cellular factors to reproduce its genome and package the progeny virions [178,179,180]. Inability of the intergenotypic chimeras to infect pigs in the study by Cordoba et al. [161] showed that other than the role of viral factor itself, swine cells might lack the essential host factors required by HEV-1 to establish successful infection in pigs. Furthermore, the inability of the chimeras to infect pigs in the study by Cordoba et al. [161] may also reflect the functional importance of species-specific protein-protein interactions during HEV replication, considering that the stem-loop within the JR and 3′ UTR of HEV is known to interact with viral RdRp (and possibly with the host cellular factors) required for viral replication. In addition, the viral mechanism to counteract the host immune response can be species-specific and can partially explain the unsuccessful infection in pigs as species-specific amino acid residues that are important for evading the immune response in pigs may be absent in this setting [161]. Supporting this hypothesis, the successful replication of intergenotypic chimeras in pig kidney cells in the study by Chaterjee et al. [156] indicates that besides the importance of the interaction between ORF1 protein domains and host cell-specific factors during replication (as described above), cellular factors might play a critical role in virus establishment as a post-entry barrier.
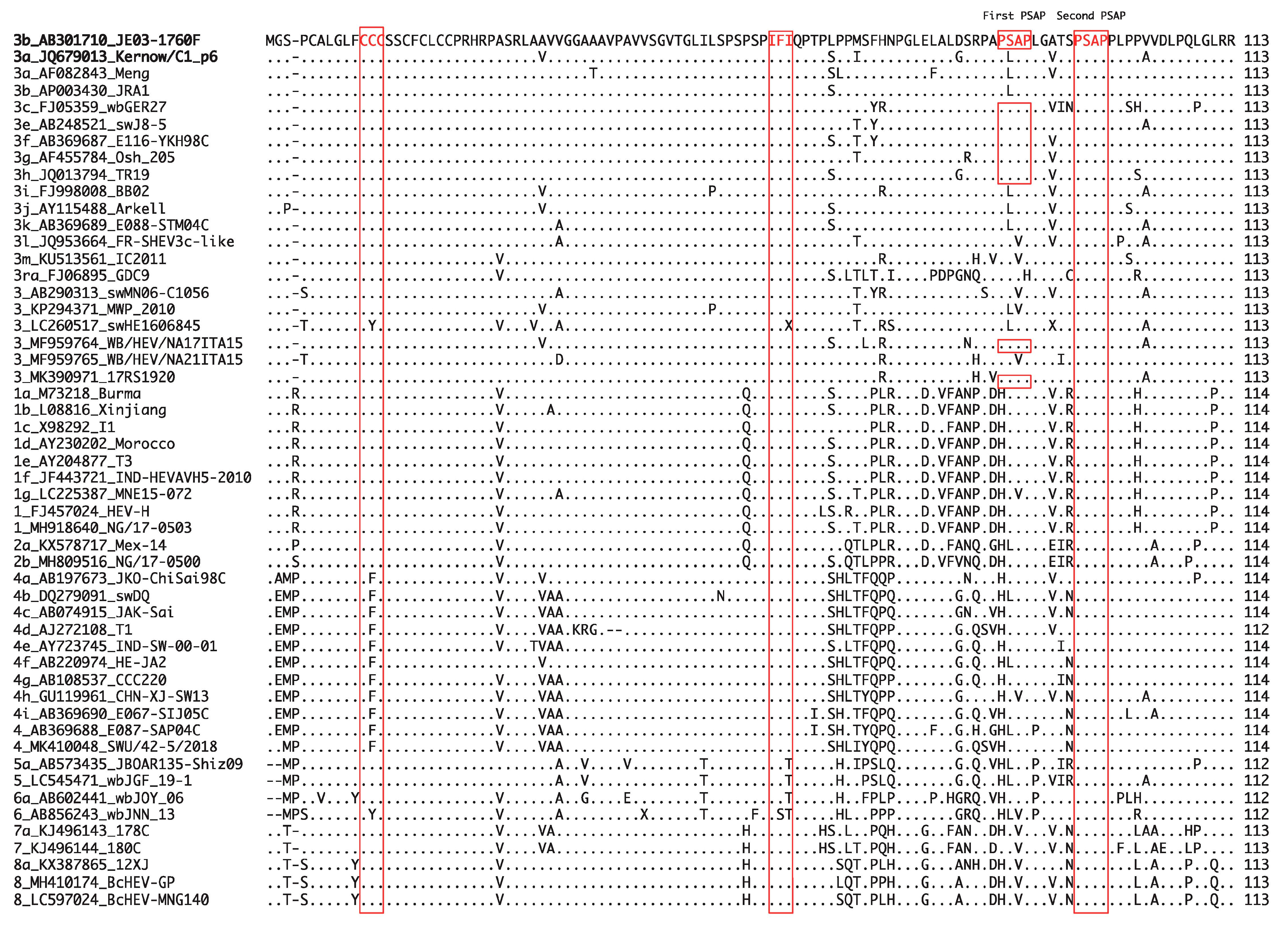
It has been demonstrated that Sar-55 and Akluj, human strains of HEV-1, can infect LLC-PK1 swine cells [154], suggesting that human and swine HEV may share at least one cell receptor. In addition, the species-specific interactions between viral and cellular proteins may be one of the determinants of the successful replication of HEV in the host [161,181]. For instance, ORF3 is known to interact with a host cellular factor, tumor susceptibility gene 101 (Tsg101), through its PSAP motif to promote virion egress [33,182] and, thus, successful viral replication. This motif is well conserved in the HEV-1–8 strains of Orthohepevirus A species (Figure 3, second PSAP). In addition to this motif, JE03-1760F of HEV-3 (used in our lab) and several other isolates of HEV-3 have one additional PSAP motif located at the N-terminal, while in other HEV-3 isolates, excluding HEV-3ra, the PXXP motif is highly conserved (Figure 3, first PSAP). Besides the highly conserved PSAP motif which interacts with the host cellular factor to facilitate virion release, the highly conserved motifs of CCC and IFI of the ORF3 in Orthohepevirus A species (Figure 3) were recently reported to promote HEV release and ion fluxes [34]. Of note, HEV-4 has CFC in place of CCC, while HEV-5 and HEV-6 have IFT or IST in place of IFI (Figure 3). Because these motifs are highly conserved, it is possible that a host cellular factor might be the other determinant of successful viral replication.
The co-evolution of virus and the hosts often leads to species specificity. The presence of specific attachment proteins/receptors and availability of a complex pool of cellular factors required for viral replication can contribute to the host specificity [156].
6.4. The Host Immune Status
The immune state of the host may also be a possible determinant in HEV cross-species transmission. In most cases, humans infected with rat HEV were in an immunocompromised state [115,116]. Host adaptation after cross-species transmission is associated with rapid amino acid sequence changes of viral genes, typically those associated with receptor interactions and the evasion of innate immunity but often pervasive throughout the entire virus genome [183]. As described above, Shukla et al. [154] demonstrated the extraordinary ability of the Kernow-C1 strain to infect cells from a broad spectrum of species, ranging from rodents to primates. This probably reflects a high titer and a complex quasi-species generated during prolonged infection in an immunocompromised host. This possibility and the demonstration that HEV can acquire new information through recombination with host cell sequences leads to the conclusion that chronic HEV infection of a patient has important implications for the evolution of this emerging virus [154].
7. Conclusions
Knowledge on the mechanism of HEV cross-species transmission has improved drastically within the past decade. However, several important questions remain to be clarified. Based on the available data to date, ORF1 has been demonstrated as an essential viral determinant in cross-species transmission of HEV; thus, dissection of the precise domain in ORF1 that confers the ability to cross the species barrier may answer one of these important questions. In addition, as the presence and interaction of viral and host determinants are essential to enable cross-species transmission, the discovery of HEV attachment protein and its interaction with host cellular receptors will greatly help in elucidating the mechanism of HEV cross-species infection.
Author Contributions
P.P.P. drafted the manuscript. S.N. assisted in the preparation of the draft of the manuscript. H.O. reviewed and edited the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported in part by Grant-in-Aid for Research Activity Start-up from Japan Society for the Promotion of Science, JSPS (to P.P.P.: 20K22771) and the Research Program on Hepatitis from Japan Agency for Medical Research and Development, AMED (to H.O.: JP21fk0210075).
Acknowledgments
We thank Kazumoto Murata (Jichi Medical University, Japan) for the suggestions and encouragement.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Smith, D.B.; Izopet, J.; Nicot, F.; Simmonds, P.; Jameel, S.; Meng, X.J.; Norder, H.; Okamoto, H.; van der Poel, W.H.M.; Reuter, G.; et al. Update: Proposed reference sequences for subtypes of hepatitis E virus (species Orthohepevirus A). J. Gen. Virol. 2020, 101, 692–698. [Google Scholar] [CrossRef]
- WHO. Hepatitis E. Available online: https://www.who.int/news-room/fact-sheets/detail/hepatitis-e (accessed on 16 March 2021).
- Kamar, N.; Selves, J.; Mansuy, J.M.; Ouezzani, L.; Peron, J.M.; Guitard, J.; Cointault, O.; Esposito, L.; Abravanel, F.; Danjoux, M.; et al. Hepatitis E virus and chronic hepatitis in organ-transplant recipients. N. Engl. J. Med. 2008, 358, 811–817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shrestha, A.; Adhikari, A.; Bhattarai, M.; Rauniyar, R.; Debes, J.D.; Boonstra, A.; Lama, T.K.; Al Mahtab, M.; Butt, A.S.; Akbar, S.M.F.; et al. Prevalence and risk of hepatitis E virus infection in the HIV population of Nepal. Virol. J. 2017, 14, 228. [Google Scholar] [CrossRef] [Green Version]
- Tavitian, S.; Peron, J.M.; Huguet, F.; Kamar, N.; Abravanel, F.; Beyne-Rauzy, O.; Oberic, L.; Faguer, S.; Alric, L.; Roussel, M.; et al. Ribavirin for chronic hepatitis prevention among patients with hematologic malignancies. Emerg. Infect. Dis. 2015, 21, 1466–1469. [Google Scholar] [CrossRef] [PubMed]
- Kamar, N.; Izopet, J.; Pavio, N.; Aggarwal, R.; Labrique, A.; Wedemeyer, H.; Dalton, H.R. Hepatitis E virus infection. Nat. Rev. Dis. Primers 2017, 3, 17086. [Google Scholar] [CrossRef] [PubMed]
- Dalton, H.R.; Kamar, N.; van Eijk, J.J.; McLean, B.N.; Cintas, P.; Bendall, R.P.; Jacobs, B.C. Hepatitis E virus and neurological injury. Nat. Rev. Neurol. 2016, 12, 77–85. [Google Scholar] [CrossRef]
- Kamar, N.; Marion, O.; Abravanel, F.; Izopet, J.; Dalton, H.R. Extrahepatic manifestations of hepatitis E virus. Liver Int. 2016, 36, 467–472. [Google Scholar] [CrossRef]
- Bazerbachi, F.; Haffar, S.; Garg, S.K.; Lake, J.R. Extra-hepatic manifestations associated with hepatitis E virus infection: A comprehensive review of the literature. Gastroenterol. Rep. 2016, 4, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Matsubayashi, K.; Nagaoka, Y.; Sakata, H.; Sato, S.; Fukai, K.; Kato, T.; Takahashi, K.; Mishiro, S.; Imai, M.; Takeda, N.; et al. Transfusion-transmitted hepatitis E caused by apparently indigenous hepatitis E virus strain in Hokkaido, Japan. Transfusion 2004, 44, 934–940. [Google Scholar] [CrossRef]
- Mitsui, T.; Tsukamoto, Y.; Yamazaki, C.; Masuko, K.; Tsuda, F.; Takahashi, M.; Nishizawa, T.; Okamoto, H. Prevalence of hepatitis E virus infection among hemodialysis patients in Japan: Evidence for infection with a genotype 3 HEV by blood transfusion. J. Med. Virol. 2004, 74, 563–572. [Google Scholar] [CrossRef]
- Boxall, E.; Herborn, A.; Kochethu, G.; Pratt, G.; Adams, D.; Ijaz, S.; Teo, C.G. Transfusion-transmitted hepatitis E in a ‘nonhyperendemic’ country. Transfus. Med. 2006, 16, 79–83. [Google Scholar] [CrossRef]
- Satake, M.; Matsubayashi, K.; Hoshi, Y.; Taira, R.; Furui, Y.; Kokudo, N.; Akamatsu, N.; Yoshizumi, T.; Ohkohchi, N.; Okamoto, H.; et al. Unique clinical courses of transfusion-transmitted hepatitis E in patients with immunosuppression. Transfusion 2017, 57, 280–288. [Google Scholar] [CrossRef] [PubMed]
- Perez-Gracia, M.T.; Suay-Garcia, B.; Mateos-Lindemann, M.L. Hepatitis E and pregnancy: Current state. Rev. Med. Virol. 2017, 27, e1929. [Google Scholar] [CrossRef] [PubMed]
- Primadharsini, P.P.; Nagashima, S.; Okamoto, H. Genetic variability and evolution of hepatitis E virus. Viruses 2019, 11, 456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, X.J.; Purcell, R.H.; Halbur, P.G.; Lehman, J.R.; Webb, D.M.; Tsareva, T.S.; Haynes, J.S.; Thacker, B.J.; Emerson, S.U. A novel virus in swine is closely related to the human hepatitis E virus. Proc. Natl. Acad. Sci. USA 1997, 94, 9860–9865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, X.J.; Halbur, P.G.; Shapiro, M.S.; Govindarajan, S.; Bruna, J.D.; Mushahwar, I.K.; Purcell, R.H.; Emerson, S.U. Genetic and experimental evidence for cross-species infection by swine hepatitis E virus. J. Virol. 1998, 72, 9714–9721. [Google Scholar] [CrossRef] [Green Version]
- Meng, X.J.; Halbur, P.G.; Haynes, J.S.; Tsareva, T.S.; Bruna, J.D.; Royer, R.L.; Purcell, R.H.; Emerson, S.U. Experimental infection of pigs with the newly identified swine hepatitis E virus (swine HEV), but not with human strains of HEV. Arch. Virol. 1998, 143, 1405–1415. [Google Scholar] [CrossRef] [PubMed]
- Tam, A.W.; Smith, M.M.; Guerra, M.E.; Huang, C.C.; Bradley, D.W.; Fry, K.E.; Reyes, G.R. Hepatitis E virus (HEV): Molecular cloning and sequencing of the full-length viral genome. Virology 1991, 185, 120–131. [Google Scholar] [CrossRef]
- Kabrane-Lazizi, Y.; Meng, X.J.; Purcell, R.H.; Emerson, S.U. Evidence that the genomic RNA of hepatitis E virus is capped. J. Virol. 1999, 73, 8848–8850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graff, J.; Torian, U.; Nguyen, H.; Emerson, S.U. A bicistronic subgenomic mRNA encodes both the ORF2 and ORF3 proteins of hepatitis E virus. J. Virol. 2006, 80, 5919–5926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ichiyama, K.; Yamada, K.; Tanaka, T.; Nagashima, S.; Jirintai; Takahashi, M.; Okamoto, H. Determination of the 5′-terminal sequence of subgenomic RNA of hepatitis E virus strains in cultured cells. Arch. Virol. 2009, 154, 1945–1951. [Google Scholar] [CrossRef] [PubMed]
- Nan, Y.; Zhang, Y.J. Molecular biology and infection of hepatitis E virus. Front. Microbiol. 2016, 7, 1419. [Google Scholar] [CrossRef]
- Sridhar, S.; Teng, J.L.L.; Chiu, T.H.; Lau, S.K.P.; Woo, P.C.Y. Hepatitis E virus genotypes and evolution: Emergence of camel hepatitis E variants. Int. J. Mol. Sci. 2017, 18, 869. [Google Scholar] [CrossRef] [Green Version]
- Szkolnicka, D.; Pollan, A.; Da Silva, N.; Oechslin, N.; Gouttenoire, J.; Moradpour, D. Recombinant hepatitis E viruses harboring tags in the ORF1 protein. J. Virol. 2019, 93, e00459-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reyes, G.R.; Huang, C.C.; Tam, A.W.; Purdy, M.A. Molecular organization and replication of hepatitis E virus (HEV). Arch. Virol. Suppl. 1993, 7, 15–25. [Google Scholar]
- Kalia, M.; Chandra, V.; Rahman, S.A.; Sehgal, D.; Jameel, S. Heparan sulfate proteoglycans are required for cellular binding of the hepatitis E virus ORF2 capsid protein and for viral infection. J. Virol. 2009, 83, 12714–12724. [Google Scholar] [CrossRef] [Green Version]
- Xing, L.; Wang, J.C.; Li, T.C.; Yasutomi, Y.; Lara, J.; Khudyakov, Y.; Schofield, D.; Emerson, S.U.; Purcell, R.H.; Takeda, N.; et al. Spatial configuration of hepatitis E virus antigenic domain. J. Virol. 2011, 85, 1117–1124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montpellier, C.; Wychowski, C.; Sayed, I.M.; Meunier, J.C.; Saliou, J.M.; Ankavay, M.; Bull, A.; Pillez, A.; Abravanel, F.; Helle, F.; et al. Hepatitis E virus lifecycle and identification of 3 forms of the ORF2 capsid protein. Gastroenterology 2018, 154, 211–223.e218. [Google Scholar] [CrossRef]
- Yin, X.; Ying, D.; Lhomme, S.; Tang, Z.; Walker, C.M.; Xia, N.; Zheng, Z.; Feng, Z. Origin, antigenicity, and function of a secreted form of ORF2 in hepatitis E virus infection. Proc. Natl. Acad. Sci. USA 2018, 115, 4773–4778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamada, K.; Takahashi, M.; Hoshino, Y.; Takahashi, H.; Ichiyama, K.; Nagashima, S.; Tanaka, T.; Okamoto, H. ORF3 protein of hepatitis E virus is essential for virion release from infected cells. J. Gen. Virol. 2009, 90 Pt 8, 1880–1891. [Google Scholar] [CrossRef]
- Emerson, S.U.; Nguyen, H.T.; Torian, U.; Burke, D.; Engle, R.; Purcell, R.H. Release of genotype 1 hepatitis E virus from cultured hepatoma and polarized intestinal cells depends on open reading frame 3 protein and requires an intact PXXP motif. J. Virol. 2010, 84, 9059–9069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagashima, S.; Takahashi, M.; Jirintai; Tanaka, T.; Yamada, K.; Nishizawa, T.; Okamoto, H. A PSAP motif in the ORF3 protein of hepatitis E virus is necessary for virion release from infected cells. J. Gen. Virol. 2011, 92 Pt2, 269–278. [Google Scholar] [CrossRef]
- Ding, Q.; Heller, B.; Capuccino, J.M.; Song, B.; Nimgaonkar, I.; Hrebikova, G.; Contreras, J.E.; Ploss, A. Hepatitis E virus ORF3 is a functional ion channel required for release of infectious particles. Proc. Natl. Acad. Sci. USA 2017, 114, 1147–1152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nair, V.P.; Anang, S.; Subramani, C.; Madhvi, A.; Bakshi, K.; Srivastava, A.; Shalimar; Nayak, B.; Ranjith Kumar, C.T.; Surjit, M. Endoplasmic reticulum stress induced synthesis of a novel viral factor mediates efficient replication of genotype-1 hepatitis E virus. PLoS Pathog. 2016, 12, e1005521. [Google Scholar] [CrossRef]
- Yadav, K.K.; Boley, P.A.; Fritts, Z.; Kenney, S.P. Ectopic expression of genotype 1 hepatitis E virus ORF4 increases genotype 3 HEV viral replication in cell culture. Viruses 2021, 13, 75. [Google Scholar] [CrossRef]
- Meng, X.J. Zoonotic and foodborne transmission of hepatitis E virus. Semin. Liver Dis. 2013, 33, 41–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, T.C.; Chijiwa, K.; Sera, N.; Ishibashi, T.; Etoh, Y.; Shinohara, Y.; Kurata, Y.; Ishida, M.; Sakamoto, S.; Takeda, N.; et al. Hepatitis E virus transmission from wild boar meat. Emerg. Infect. Dis. 2005, 11, 1958–1960. [Google Scholar] [CrossRef] [PubMed]
- Tei, S.; Kitajima, N.; Takahashi, K.; Mishiro, S. Zoonotic transmission of hepatitis E virus from deer to human beings. Lancet 2003, 362, 371–373. [Google Scholar] [CrossRef]
- Takahashi, K.; Kitajima, N.; Abe, N.; Mishiro, S. Complete or near-complete nucleotide sequences of hepatitis E virus genome recovered from a wild boar, a deer, and four patients who ate the deer. Virology 2004, 330, 501–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cossaboom, C.M.; Cordoba, L.; Dryman, B.A.; Meng, X.J. Hepatitis E virus in rabbits, Virginia, USA. Emerg. Infect. Dis. 2011, 17, 2047–2049. [Google Scholar] [CrossRef]
- Abravanel, F.; Lhomme, S.; El Costa, H.; Schvartz, B.; Peron, J.M.; Kamar, N.; Izopet, J. Rabbit hepatitis E virus infections in humans, France. Emerg. Infect. Dis. 2017, 23, 1191–1193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, K.; Terada, S.; Kokuryu, H.; Arai, M.; Mishiro, S. A wild boar-derived hepatitis E virus isolate presumably representing so far unidentified “genotype 5”. Kanzo 2010, 51, 536–538. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, M.; Nishizawa, T.; Sato, H.; Sato, Y.; Jirintai; Nagashima, S.; Okamoto, H. Analysis of the full-length genome of a hepatitis E virus isolate obtained from a wild boar in Japan that is classifiable into a novel genotype. J. Gen. Virol. 2011, 92 Pt 4, 902–908. [Google Scholar] [CrossRef]
- Takahashi, M.; Nishizawa, T.; Nagashima, S.; Jirintai, S.; Kawakami, M.; Sonoda, Y.; Suzuki, T.; Yamamoto, S.; Shigemoto, K.; Ashida, K.; et al. Molecular characterization of a novel hepatitis E virus (HEV) strain obtained from a wild boar in Japan that is highly divergent from the previously recognized HEV strains. Virus Res. 2014, 180, 59–69. [Google Scholar] [CrossRef]
- Takahashi, M.; Nishizawa, T.; Sato, Y.; Miyazaki, S.; Aikawa, T.; Ashida, K.; Tamaru, T.; Oguro, K.; Hayakawa, F.; Matsuoka, H.; et al. Prevalence and genotype/subtype distribution of hepatitis E virus (HEV) among wild boars in Japan: Identification of a genotype 5 HEV strain. Virus Res. 2020, 287, 198106. [Google Scholar] [CrossRef] [PubMed]
- Woo, P.C.; Lau, S.K.; Teng, J.L.; Tsang, A.K.; Joseph, M.; Wong, E.Y.; Tang, Y.; Sivakumar, S.; Xie, J.; Bai, R.; et al. New hepatitis E virus genotype in camels, the Middle East. Emerg. Infect. Dis. 2014, 20, 1044–1048. [Google Scholar] [CrossRef] [PubMed]
- Rasche, A.; Saqib, M.; Liljander, A.M.; Bornstein, S.; Zohaib, A.; Renneker, S.; Steinhagen, K.; Wernery, R.; Younan, M.; Gluecks, I.; et al. Hepatitis E virus infection in dromedaries, North and East Africa, United Arab Emirates, and Pakistan, 1983–2015. Emerg. Infect. Dis. 2016, 22, 1249–1252. [Google Scholar] [CrossRef]
- Lee, G.H.; Tan, B.H.; Teo, E.C.; Lim, S.G.; Dan, Y.Y.; Wee, A.; Aw, P.P.; Zhu, Y.; Hibberd, M.L.; Tan, C.K.; et al. Chronic infection with camelid hepatitis E virus in a liver transplant recipient who regularly consumes camel meat and milk. Gastroenterology 2016, 150, 355–357.e353. [Google Scholar] [CrossRef] [Green Version]
- Woo, P.C.; Lau, S.K.; Teng, J.L.; Cao, K.Y.; Wernery, U.; Schountz, T.; Chiu, T.H.; Tsang, A.K.; Wong, P.C.; Wong, E.Y.; et al. New hepatitis E virus genotype in Bactrian camels, Xinjiang, China, 2013. Emerg. Infect. Dis. 2016, 22, 2219–2221. [Google Scholar] [CrossRef]
- Nishizawa, T.; Takahashi, M.; Tsatsralt-Od, B.; Nyamdavaa, K.; Dulmaa, N.; Osorjin, B.; Tseren-Ochir, E.O.; Sharav, T.; Bayasgalan, C.; Sukhbaatar, B.; et al. Identification and a full genome analysis of novel camel hepatitis E virus strains obtained from Bactrian camels in Mongolia. Virus Res. 2021, 299, 198355. [Google Scholar] [CrossRef]
- Lin, J.; Norder, H.; Uhlhorn, H.; Belak, S.; Widen, F. Novel hepatitis E like virus found in Swedish moose. J. Gen. Virol. 2014, 95 Pt 3, 557–570. [Google Scholar] [CrossRef] [Green Version]
- Lin, J.; Karlsson, M.; Olofson, A.S.; Belak, S.; Malmsten, J.; Dalin, A.M.; Widen, F.; Norder, H. High prevalence of hepatitis E virus in Swedish moose-a phylogenetic characterization and comparison of the virus from different regions. PLoS ONE 2015, 10, e0122102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.; Wang, L.; Shen, H.; Zheng, Y.; Gauger, P.C.; Chen, Q.; Zhang, J.; Yoon, K.J.; Harmon, K.M.; Main, R.G.; et al. Detection and genomic characterization of new avian-like hepatitis E virus in a sparrow in the United States. Arch. Virol. 2018, 163, 2861–2864. [Google Scholar] [CrossRef]
- Liu, B.; Chen, Y.; Zhao, L.; Zhang, M.; Ren, X.; Zhang, Y.; Zhang, B.; Fan, M.; Zhao, Q.; Zhou, E.M. Identification and pathogenicity of a novel genotype avian hepatitis E virus from silkie fowl (Gallus gallus). Vet. Microbiol. 2020, 245, 108688. [Google Scholar] [CrossRef] [PubMed]
- Reuter, G.; Boros, A.; Matics, R.; Kapusinszky, B.; Delwart, E.; Pankovics, P. A novel avian-like hepatitis E virus in wild aquatic bird, little egret (Egretta garzetta), in Hungary. Infect. Genet. Evol. 2016, 46, 74–77. [Google Scholar] [CrossRef]
- Wang, B.; Harms, D.; Yang, X.L.; Bock, C.T. Orthohepevirus C: An expanding species of emerging hepatitis E virus variants. Pathogens 2020, 9, 154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, B.; Li, W.; Zhou, J.H.; Li, B.; Zhang, W.; Yang, W.H.; Pan, H.; Wang, L.X.; Bock, C.T.; Shi, Z.L.; et al. Chevrier’s field mouse (Apodemus chevrieri) and Pere David’s vole (Eothenomys melanogaster) in China carry orthohepeviruses that form two putative novel genotypes within the species Orthohepevirus C. Virol. Sin. 2018, 33, 44–58. [Google Scholar] [CrossRef] [PubMed]
- Ryll, R.; Heckel, G.; Corman, V.M.; Drexler, J.F.; Ulrich, R.G. Genomic and spatial variability of a European common vole hepevirus. Arch. Virol. 2019, 164, 2671–2682. [Google Scholar] [CrossRef]
- Wu, Z.; Yang, L.; Ren, X.; He, G.; Zhang, J.; Yang, J.; Qian, Z.; Dong, J.; Sun, L.; Zhu, Y.; et al. Deciphering the bat virome catalog to better understand the ecological diversity of bat viruses and the bat origin of emerging infectious diseases. ISME J. 2016, 10, 609–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nimgaonkar, I.; Ding, Q.; Schwartz, R.E.; Ploss, A. Hepatitis E virus: Advances and challenges. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 96–110. [Google Scholar] [CrossRef]
- Hoofnagle, J.H.; Nelson, K.E.; Purcell, R.H. Hepatitis E. N. Engl. J. Med. 2012, 367, 1237–1244. [Google Scholar] [CrossRef]
- Boccia, D.; Guthmann, J.P.; Klovstad, H.; Hamid, N.; Tatay, M.; Ciglenecki, I.; Nizou, J.Y.; Nicand, E.; Guerin, P.J. High mortality associated with an outbreak of hepatitis E among displaced persons in Darfur, Sudan. Clin. Infect. Dis. 2006, 42, 1679–1684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tripathy, A.S.; Sharma, M.; Deoshatwar, A.R.; Babar, P.; Bharadwaj, R.; Bharti, O.K. Study of a hepatitis E virus outbreak involving drinking water and sewage contamination in Shimla, India, 2015-2016. Trans. R. Soc. Trop. Med. Hyg. 2019, 113, 789–796. [Google Scholar] [CrossRef] [PubMed]
- Tricou, V.; Bouscaillou, J.; Laghoe-Nguembe, G.L.; Bere, A.; Konamna, X.; Selekon, B.; Nakoune, E.; Kazanji, M.; Komas, N.P. Hepatitis E virus outbreak associated with rainfall in the Central African Republic in 2008-2009. BMC Infect. Dis. 2020, 20, 260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galiana, C.; Fernandez-Barredo, S.; Garcia, A.; Gomez, M.T.; Perez-Gracia, M.T. Occupational exposure to hepatitis E virus (HEV) in swine workers. Am. J. Trop. Med. Hyg. 2008, 78, 1012–1015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teixeira, J.; Mesquita, J.R.; Pereira, S.S.; Oliveira, R.M.; Abreu-Silva, J.; Rodrigues, A.; Myrmel, M.; Stene-Johansen, K.; Overbo, J.; Goncalves, G.; et al. Prevalence of hepatitis E virus antibodies in workers occupationally exposed to swine in Portugal. Med. Microbiol. Immunol. 2017, 206, 77–81. [Google Scholar] [CrossRef] [Green Version]
- Meester, M.; Tobias, T.J.; Bouwknegt, M.; Kusters, N.E.; Stegeman, J.A.; van der Poel, W.H.M. Infection dynamics and persistence of hepatitis E virus on pig farms—A review. Porc. Health Manag. 2021, 7, 16. [Google Scholar] [CrossRef] [PubMed]
- Clemente-Casares, P.; Ramos-Romero, C.; Ramirez-Gonzalez, E.; Mas, A. Hepatitis E virus in industrialized countries: The silent threat. Biomed. Res. Int. 2016, 2016, 9838041. [Google Scholar] [CrossRef]
- Di Bartolo, I.; Diez-Valcarce, M.; Vasickova, P.; Kralik, P.; Hernandez, M.; Angeloni, G.; Ostanello, F.; Bouwknegt, M.; Rodriguez-Lazaro, D.; Pavlik, I.; et al. Hepatitis E virus in pork production chain in Czech Republic, Italy, and Spain, 2010. Emerg. Infect. Dis. 2012, 18, 1282–1289. [Google Scholar] [CrossRef]
- Webb, G.W.; Dalton, H.R. Hepatitis E: An underestimated emerging threat. Ther. Adv. Infect. Dis. 2019, 6, 2049936119837162. [Google Scholar] [CrossRef] [Green Version]
- Webb, G.W.; Kelly, S.; Dalton, H.R. Hepatitis A and Hepatitis E: Clinical and epidemiological features, diagnosis, treatment, and prevention. Clin. Microbiol. Newsl. 2020, 42, 171–179. [Google Scholar] [CrossRef]
- Said, B.; Ijaz, S.; Kafatos, G.; Booth, L.; Thomas, H.L.; Walsh, A.; Ramsay, M.; Morgan, D. Hepatitis E Incident Investigation Team. Hepatitis E outbreak on cruise ship. Emerg. Infect. Dis. 2009, 15, 1738–1744. [Google Scholar] [CrossRef]
- Bofill-Mas, S.; Albinana-Gimenez, N.; Clemente-Casares, P.; Hundesa, A.; Rodriguez-Manzano, J.; Allard, A.; Calvo, M.; Girones, R. Quantification and stability of human adenoviruses and polyomavirus JCPyV in wastewater matrices. Appl. Environ. Microbiol. 2006, 72, 7894–7896. [Google Scholar] [CrossRef] [Green Version]
- Albinana-Gimenez, N.; Clemente-Casares, P.; Bofill-Mas, S.; Hundesa, A.; Ribas, F.; Girones, R. Distribution of human polyomaviruses, adenoviruses, and hepatitis E virus in the environment and in a drinking-water treatment plant. Environ. Sci. Technol. 2006, 40, 7416–7422. [Google Scholar] [CrossRef] [PubMed]
- Clemente-Casares, P.; Rodriguez-Manzano, J.; Girones, R. Hepatitis E virus genotype 3 and sporadically also genotype 1 circulate in the population of Catalonia, Spain. J. Water Health 2009, 7, 664–673. [Google Scholar] [CrossRef]
- Takahashi, M.; Yamada, K.; Hoshino, Y.; Takahashi, H.; Ichiyama, K.; Tanaka, T.; Okamoto, H. Monoclonal antibodies raised against the ORF3 protein of hepatitis E virus (HEV) can capture HEV particles in culture supernatant and serum but not those in feces. Arch. Virol. 2008, 153, 1703–1713. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, M.; Tanaka, T.; Takahashi, H.; Hoshino, Y.; Nagashima, S.; Jirintai; Mizuo, H.; Yazaki, Y.; Takagi, T.; Azuma, M.; et al. Hepatitis E virus (HEV) strains in serum samples can replicate efficiently in cultured cells despite the coexistence of HEV antibodies: Characterization of HEV virions in blood circulation. J. Clin. Microbiol. 2010, 48, 1112–1125. [Google Scholar] [CrossRef] [Green Version]
- Yin, X.; Ambardekar, C.; Lu, Y.; Feng, Z. Distinct entry mechanisms for nonenveloped and quasi-enveloped hepatitis E viruses. J. Virol. 2016, 90, 4232–4242. [Google Scholar] [CrossRef] [Green Version]
- Nagashima, S.; Takahashi, M.; Kobayashi, T.; Tanggis; Nishizawa, T.; Nishiyama, T.; Primadharsini, P.P.; Okamoto, H. Characterization of the quasi-enveloped hepatitis E virus particles released by the cellular exosomal pathway. J. Virol. 2017, 91, e00822-17. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Wei, M.; Pan, H.; Lin, Z.; Wang, K.; Weng, Z.; Zhu, Y.; Xin, L.; Zhang, J.; Li, S.; et al. Robust manufacturing and comprehensive characterization of recombinant hepatitis E virus-like particles in Hecolin(®). Vaccine 2014, 32, 4039–4050. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, H. Hepatitis E virus cell culture models. Virus Res. 2011, 161, 65–77. [Google Scholar] [CrossRef]
- Okamoto, H. Culture systems for hepatitis E virus. J. Gastroenterol. 2013, 48, 147–158. [Google Scholar] [CrossRef] [Green Version]
- Ju, X.; Ding, Q. Hepatitis E virus assembly and release. Viruses 2019, 11, 539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capelli, N.; Marion, O.; Dubois, M.; Allart, S.; Bertrand-Michel, J.; Lhomme, S.; Abravanel, F.; Izopet, J.; Chapuy-Regaud, S. Vectorial release of hepatitis E virus in polarized human hepatocytes. J. Virol. 2019, 93, e01207–e01218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fousekis, F.S.; Mitselos, I.V.; Christodoulou, D.K. Extrahepatic manifestations of hepatitis E virus: An overview. Clin. Mol. Hepatol. 2020, 26, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Devhare, P.B.; Chatterjee, S.N.; Arankalle, V.A.; Lole, K.S. Analysis of antiviral response in human epithelial cells infected with hepatitis E virus. PLoS ONE 2013, 8, e63793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahn, H.S.; Han, S.H.; Kim, Y.H.; Park, B.J.; Kim, D.H.; Lee, J.B.; Park, S.Y.; Song, C.S.; Lee, S.W.; Choi, C.; et al. Adverse fetal outcomes in pregnant rabbits experimentally infected with rabbit hepatitis E virus. Virology 2017, 512, 187–193. [Google Scholar] [CrossRef]
- Cao, D.; Cao, Q.M.; Subramaniam, S.; Yugo, D.M.; Heffron, C.L.; Rogers, A.J.; Kenney, S.P.; Tian, D.; Matzinger, S.R.; Overend, C.; et al. Pig model mimicking chronic hepatitis E virus infection in immunocompromised patients to assess immune correlates during chronicity. Proc. Natl. Acad. Sci. USA 2017, 114, 6914–6923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Majumdar, M.; Ratho, R.K.; Chawla, Y.; Singh, M.P. Role of TLR gene expression and cytokine profiling in the immunopathogenesis of viral hepatitis E. J. Clin. Virol. 2015, 73, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Devi, S.G.; Kar, P.; Agarwal, S.; Husain, S.A.; Gupta, R.K.; Sharma, S. Association of cytokines in hepatitis E with pregnancy outcome. Cytokine 2014, 65, 95–104. [Google Scholar] [CrossRef]
- Nan, Y.; Yu, Y.; Ma, Z.; Khattar, S.K.; Fredericksen, B.; Zhang, Y.J. Hepatitis E virus inhibits type I interferon induction by ORF1 products. J. Virol. 2014, 88, 11924–11932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Surjit, M.; Varshney, B.; Lal, S.K. The ORF2 glycoprotein of hepatitis E virus inhibits cellular NF-kappaB activity by blocking ubiquitination mediated proteasomal degradation of IkappaBalpha in human hepatoma cells. BMC Biochem. 2012, 13, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, C.; Zafrullah, M.; Mixson-Hayden, T.; Dai, X.; Liang, J.; Meng, J.; Kamili, S. Suppression of interferon-alpha signaling by hepatitis E virus. Hepatology 2012, 55, 1324–1332. [Google Scholar] [CrossRef]
- Pischke, S.; Hartl, J.; Pas, S.D.; Lohse, A.W.; Jacobs, B.C.; Van der Eijk, A.A. Hepatitis E virus: Infection beyond the liver? J. Hepatol. 2017, 66, 1082–1095. [Google Scholar] [CrossRef] [Green Version]
- Yin, X.; Feng, Z. Hepatitis E virus entry. Viruses 2019, 11, 883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, H.; Zhao, Y.; Zhang, X.; Wang, B.; Liu, P. Case-fatality risk of pregnant women with acute viral hepatitis type E: A systematic review and meta-analysis. Epidemiol. Infect. 2016, 144, 2098–2106. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Wu, X.; Xia, J. Hepatitis E virus infection during pregnancy. Virol. J. 2020, 17, 73. [Google Scholar] [CrossRef] [PubMed]
- Navaneethan, U.; Al Mohajer, M.; Shata, M.T. Hepatitis E and pregnancy: Understanding the pathogenesis. Liver Int. 2008, 28, 1190–1199. [Google Scholar] [CrossRef] [Green Version]
- Gong, S.; Hao, X.; Bi, Y.; Yang, C.; Wang, W.; Mickael, H.K.; Zhang, Y.; Chen, S.; Qian, Z.; Huang, F.; et al. Hepatitis E viral infection regulates estrogen signaling pathways: Inhibition of the cAMPK-PKA-CREB and PI3K-AKT-mTOR signaling pathways. J. Med. Virol. 2021, 93, 3769–3778. [Google Scholar] [CrossRef]
- Robins, A.E.M.; Bowden, D.J.; Gelson, W.T.H. Chronic genotype 1 hepatitis E infection from immunosuppression for ileo-colonic Crohn’s disease. Oxf. Med. Case Rep. 2018, 2018, omy059. [Google Scholar] [CrossRef] [PubMed]
- Geng, Y.; Zhang, H.; Huang, W.; Harrison, T.J.; Geng, K.; Li, Z.; Wang, Y. Persistent hepatitis E virus genotype 4 infection in a child with acute lymphoblastic leukemia. Hepat. Mon. 2014, 14, e15618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thakur, V.; Ratho, R.K.; Kumar, S.; Saxena, S.K.; Bora, I.; Thakur, P. Viral hepatitis E and chronicity: A growing public health concern. Front. Microbiol. 2020, 11, 577339. [Google Scholar] [CrossRef] [PubMed]
- Miyamura, T. Hepatitis E virus infection in developed countries. Virus Res. 2011, 161, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, M.; Okamoto, H. Features of hepatitis E virus infection in humans and animals in Japan. Hepatol. Res. 2014, 44, 43–58. [Google Scholar] [CrossRef] [PubMed]
- Izopet, J.; Tremeaux, P.; Marion, O.; Migueres, M.; Capelli, N.; Chapuy-Regaud, S.; Mansuy, J.M.; Abravanel, F.; Kamar, N.; Lhomme, S. Hepatitis E virus infections in Europe. J. Clin. Virol. 2019, 120, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Pavio, N.; Meng, X.J.; Renou, C. Zoonotic hepatitis E: Animal reservoirs and emerging risks. Vet. Res. 2010, 41, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Izopet, J.; Dubois, M.; Bertagnoli, S.; Lhomme, S.; Marchandeau, S.; Boucher, S.; Kamar, N.; Abravanel, F.; Guerin, J.L. Hepatitis E virus strains in rabbits and evidence of a closely related strain in humans, France. Emerg. Infect. Dis. 2012, 18, 1274–1281. [Google Scholar] [CrossRef] [PubMed]
- Arankalle, V.A.; Chobe, L.P.; Chadha, M.S. Type-IV Indian swine HEV infects rhesus monkeys. J. Viral Hepat. 2006, 13, 742–745. [Google Scholar] [CrossRef]
- Feagins, A.R.; Opriessnig, T.; Huang, Y.W.; Halbur, P.G.; Meng, X.J. Cross-species infection of specific-pathogen-free pigs by a genotype 4 strain of human hepatitis E virus. J. Med. Virol. 2008, 80, 1379–1386. [Google Scholar] [CrossRef] [Green Version]
- Feagins, A.R.; Cordoba, L.; Sanford, B.J.; Dryman, B.A.; Huang, Y.W.; LeRoith, T.; Emerson, S.U.; Meng, X.J. Intergenotypic chimeric hepatitis E viruses (HEVs) with the genotype 4 human HEV capsid gene in the backbone of genotype 3 swine HEV are infectious in pigs. Virus Res. 2011, 156, 141–146. [Google Scholar] [CrossRef] [Green Version]
- Li, T.C.; Bai, H.; Yoshizaki, S.; Ami, Y.; Suzaki, Y.; Doan, Y.H.; Takahashi, K.; Mishiro, S.; Takeda, N.; Wakita, T. Genotype 5 hepatitis E virus produced by a reverse genetics system has the potential for zoonotic infection. Hepatol. Commun. 2019, 3, 160–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, T.C.; Zhou, X.; Yoshizaki, S.; Ami, Y.; Suzaki, Y.; Nakamura, T.; Takeda, N.; Wakita, T. Production of infectious dromedary camel hepatitis E virus by a reverse genetic system: Potential for zoonotic infection. J. Hepatol. 2016, 65, 1104–1111. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Teng, J.L.L.; Lau, S.K.P.; Sridhar, S.; Fu, H.; Gong, W.; Li, M.; Xu, Q.; He, Y.; Zhuang, H.; et al. Transmission of a novel genotype of hepatitis E virus from Bactrian camels to cynomolgus macaques. J. Virol. 2019, 93, e02014–e02018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sridhar, S.; Yip, C.C.Y.; Wu, S.; Cai, J.; Zhang, A.J.; Leung, K.H.; Chung, T.W.H.; Chan, J.F.W.; Chan, W.M.; Teng, J.L.L.; et al. Rat hepatitis E virus as cause of persistent hepatitis after liver transplant. Emerg. Infect. Dis. 2018, 24, 2241–2250. [Google Scholar] [CrossRef] [Green Version]
- Sridhar, S.; Yip, C.C.; Wu, S.; Chew, N.F.; Leung, K.H.; Chan, J.F.; Zhao, P.S.; Chan, W.M.; Poon, R.W.; Tsoi, H.W.; et al. Transmission of rat hepatitis E virus infection to humans in Hong Kong: A clinical and epidemiological analysis. Hepatology 2021, 73, 10–22. [Google Scholar] [CrossRef] [PubMed]
- Andonov, A.; Robbins, M.; Borlang, J.; Cao, J.; Hatchette, T.; Stueck, A.; Deschambault, Y.; Murnaghan, K.; Varga, J.; Johnston, L. Rat hepatitis E virus linked to severe acute hepatitis in an immunocompetent patient. J. Infect. Dis. 2019, 220, 951–955. [Google Scholar] [CrossRef]
- Reuter, G.; Boros, A.; Pankovics, P. Review of hepatitis E virus in rats: Evident risk of species Orthohepevirus C to human zoonotic infection and disease. Viruses 2020, 12, 1148. [Google Scholar] [CrossRef] [PubMed]
- Tsarev, S.A.; Emerson, S.U.; Reyes, G.R.; Tsareva, T.S.; Legters, L.J.; Malik, I.A.; Iqbal, M.; Purcell, R.H. Characterization of a prototype strain of hepatitis E virus. Proc. Natl. Acad. Sci. USA 1992, 89, 559–563. [Google Scholar] [CrossRef] [Green Version]
- Emerson, S.U.; Zhang, M.; Meng, X.J.; Nguyen, H.; St Claire, M.; Govindarajan, S.; Huang, Y.K.; Purcell, R.H. Recombinant hepatitis E virus genomes infectious for primates: Importance of capping and discovery of a cis-reactive element. Proc. Natl. Acad. Sci. USA 2001, 98, 15270–15275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Gong, W.; Song, W.T.; Fu, H.; Wang, L.; Li, M.; Wang, L.; Zhuang, H. Different susceptibility and pathogenesis of rabbit genotype 3 hepatitis E virus (HEV-3) and human HEV-3 (JRC-HE3) in SPF rabbits. Vet. Microbiol. 2017, 207, 1–6. [Google Scholar] [CrossRef]
- Takahashi, M.; Nishizawa, T.; Miyajima, H.; Gotanda, Y.; Iita, T.; Tsuda, F.; Okamoto, H. Swine hepatitis E virus strains in Japan form four phylogenetic clusters comparable with those of Japanese isolates of human hepatitis E virus. J. Gen. Virol. 2003, 84 Pt 4, 851–862. [Google Scholar] [CrossRef]
- Yazaki, Y.; Mizuo, H.; Takahashi, M.; Nishizawa, T.; Sasaki, N.; Gotanda, Y.; Okamoto, H. Sporadic acute or fulminant hepatitis E in Hokkaido, Japan, may be food-borne, as suggested by the presence of hepatitis E virus in pig liver as food. J. Gen. Virol. 2003, 84 Pt 9, 2351–2357. [Google Scholar] [CrossRef]
- Inoue, J.; Takahashi, M.; Ito, K.; Shimosegawa, T.; Okamoto, H. Analysis of human and swine hepatitis E virus (HEV) isolates of genotype 3 in Japan that are only 81-83% similar to reported HEV isolates of the same genotype over the entire genome. J. Gen. Virol. 2006, 87 Pt 8, 2363–2369. [Google Scholar] [CrossRef]
- Colson, P.; Borentain, P.; Queyriaux, B.; Kaba, M.; Moal, V.; Gallian, P.; Heyries, L.; Raoult, D.; Gerolami, R. Pig liver sausage as a source of hepatitis E virus transmission to humans. J. Infect. Dis. 2010, 202, 825–834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guillois, Y.; Abravanel, F.; Miura, T.; Pavio, N.; Vaillant, V.; Lhomme, S.; Le Guyader, F.S.; Rose, N.; Le Saux, J.C.; King, L.A.; et al. High proportion of asymptomatic infections in an outbreak of hepatitis E associated with a spit-roasted piglet, France, 2013. Clin. Infect. Dis. 2016, 62, 351–357. [Google Scholar] [CrossRef]
- Schlosser, J.; Eiden, M.; Vina-Rodriguez, A.; Fast, C.; Dremsek, P.; Lange, E.; Ulrich, R.G.; Groschup, M.H. Natural and experimental hepatitis E virus genotype 3-infection in European wild boar is transmissible to domestic pigs. Vet. Res. 2014, 45, 121. [Google Scholar] [CrossRef] [PubMed]
- Thiry, D.; Rose, N.; Mauroy, A.; Paboeuf, F.; Dams, L.; Roels, S.; Pavio, N.; Thiry, E. Susceptibility of pigs to zoonotic hepatitis E virus genotype 3 isolated from a wild boar. Transbound. Emerg. Dis. 2017, 64, 1589–1597. [Google Scholar] [CrossRef] [PubMed]
- Schlosser, J.; Dahnert, L.; Dremsek, P.; Tauscher, K.; Fast, C.; Ziegler, U.; Groner, A.; Ulrich, R.G.; Groschup, M.H.; Eiden, M. Different outcomes of experimental hepatitis E virus infection in diverse mouse strains, Wistar rats, and rabbits. Viruses 2018, 11, 1. [Google Scholar] [CrossRef] [Green Version]
- Okano, H.; Nakano, T.; Sugimoto, K.; Takahashi, K.; Nagashima, S.; Takahashi, M.; Arai, M.; Okamoto, H. High genomic similarity between European type hepatitis E virus subgenotype 3e strains isolated from an acute hepatitis patient and a wild boar in Mie, Japan. Hepatol. Res. 2014, 44, 694–699. [Google Scholar] [CrossRef]
- Rivero-Juarez, A.; Frias, M.; Martinez-Peinado, A.; Risalde, M.A.; Rodriguez-Cano, D.; Camacho, A.; Garcia-Bocanegra, I.; Cuenca-Lopez, F.; Gomez-Villamandos, J.C.; Rivero, A. Familial hepatitis E outbreak linked to wild boar meat consumption. Zoonoses Public Health 2017, 64, 561–565. [Google Scholar] [CrossRef]
- Wang, H.; Castillo-Contreras, R.; Saguti, F.; Lopez-Olvera, J.R.; Karlsson, M.; Mentaberre, G.; Lindh, M.; Serra-Cobo, J.; Norder, H. Genetically similar hepatitis E virus strains infect both humans and wild boars in the Barcelona area, Spain, and Sweden. Transbound. Emerg. Dis. 2019, 66, 978–985. [Google Scholar] [CrossRef]
- Schlosser, J.; Vina-Rodriguez, A.; Fast, C.; Groschup, M.H.; Eiden, M. Chronically infected wild boar can transmit genotype 3 hepatitis E virus to domestic pigs. Vet. Microbiol. 2015, 180, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Cossaboom, C.M.; Cordoba, L.; Sanford, B.J.; Pineyro, P.; Kenney, S.P.; Dryman, B.A.; Wang, Y.; Meng, X.J. Cross-species infection of pigs with a novel rabbit, but not rat, strain of hepatitis E virus isolated in the United States. J. Gen. Virol. 2012, 93 Pt 8, 1687–1695. [Google Scholar] [CrossRef]
- Wang, L.; Liu, L.; Wang, L. An overview: Rabbit hepatitis E virus (HEV) and rabbit providing an animal model for HEV study. Rev. Med. Virol. 2018, 28, 1. [Google Scholar] [CrossRef]
- Yu, W.H.; Hao, X.H.; Li, Y.; Yang, C.C.; Li, Y.L.; He, Z.L.; Huang, F. Vertical transmission of hepatitis E virus in pregnant rhesus macaques. Sci. Rep. UK 2020, 10, 1. [Google Scholar] [CrossRef]
- Wu, Q.X.; An, J.Q.; She, R.P.; Shi, R.H.; Hao, W.Z.; Soomro, M.; Yuan, X.R.; Yang, J.L.; Wang, J.Y. Detection of genotype 4 swine hepatitis E virus in systemic tissues in cross-species infected rabbits. PLoS ONE 2017, 12, e0171277. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.; Sun, Y.; Du, T.; Chen, Y.; Wang, X.; Huang, B.; Li, H.; Nan, Y.; Xiao, S.; Zhang, G.; et al. Rabbit hepatitis E virus is an opportunistic pathogen in specific-pathogen-free rabbits with the capability of cross-species transmission. Vet. Microbiol. 2017, 201, 72–77. [Google Scholar] [CrossRef] [PubMed]
- Li, W.G.; Sun, Q.; She, R.P.; Wang, D.C.; Duan, X.H.; Yin, J.; Ding, Y. Experimental infection of Mongolian gerbils by a genotype 4 strain of swine hepatitis E virus. J. Med. Virol. 2009, 81, 1591–1596. [Google Scholar] [CrossRef] [PubMed]
- Shi, R.; Soomro, M.H.; She, R.; Yang, Y.; Wang, T.; Wu, Q.; Li, H.; Hao, W. Evidence of hepatitis E virus breaking through the blood-brain barrier and replicating in the central nervous system. J. Viral Hepat. 2016, 23, 930–939. [Google Scholar] [CrossRef]
- Meng, X.J. Expanding host range and cross-species infection of hepatitis E virus. PLoS Pathog. 2016, 12, e1005695. [Google Scholar] [CrossRef]
- Dalton, H.R.; Izopet, J. Transmission and epidemiology of hepatitis E virus genotype 3 and 4 infections. Cold Spring Harb. Perspect. Med. 2018, 8, a032144. [Google Scholar] [CrossRef]
- Jeong, S.H.; Park, B.J.; Kim, Y.H.; Choi, Y.S.; Ahn, H.S.; Han, S.H.; Choi, I.S. Isolation of hepatitis E virus genotype 4 from patients with acute cryptogenic hepatitis in Korea. J. Clin. Virol. 2017, 89, 10–13. [Google Scholar] [CrossRef]
- Shimizu, Y.; Yamada, M.; Tatematsu, H.; Ishihara, M.; Morita, K.; Ishiguro, Y.; Katano, Y.; Goto, H.; Takahashi, M.; Okamoto, H. Four cases of hepatitis E after eating wild boar meats in Aichi, Japan. Kanzo 2006, 47, 465–473. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.M.; Jeong, S.H.; Kim, J.Y.; Song, J.C.; Lee, J.H.; Kim, J.W.; Yun, H.; Kim, J.S. The first case of genotype 4 hepatitis E related to wild boar in South Korea. J. Clin. Virol. 2011, 50, 253–256. [Google Scholar] [CrossRef]
- Sun, Z.F.; Larsen, C.T.; Huang, F.F.; Billam, P.; Pierson, F.W.; Toth, T.E.; Meng, X.J. Generation and infectivity titration of an infectious stock of avian hepatitis E virus (HEV) in chickens and cross-species infection of turkeys with avian HEV. J. Clin. Microbiol. 2004, 42, 2658–2662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yugo, D.M.; Hauck, R.; Shivaprasad, H.L.; Meng, X.J. Hepatitis virus infections in poultry. Avian Dis. 2016, 60, 576–588. [Google Scholar] [CrossRef] [PubMed]
- Warren, C.J.; Sawyer, S.L. How host genetics dictates successful viral zoonosis. PLoS Biol. 2019, 17, e3000217. [Google Scholar] [CrossRef] [PubMed]
- Manrubia, S.C.; Lazaro, E. Viral evolution. Phys. Life Rev. 2006, 3, 65–92. [Google Scholar] [CrossRef]
- Vijaykrishna, D.; Mukerji, R.; Smith, G.J. RNA virus reassortment: An evolutionary mechanism for host jumps and immune evasion. PLoS Pathog. 2015, 11, e1004902. [Google Scholar] [CrossRef] [Green Version]
- Stern, A.; Yeh, M.T.; Zinger, T.; Smith, M.; Wright, C.; Ling, G.; Nielsen, R.; Macadam, A.; Andino, R. The evolutionary pathway to virulence of an RNA virus. Cell 2017, 169, 35–46.e19. [Google Scholar] [CrossRef] [Green Version]
- Dolan, P.T.; Whitfield, Z.J.; Andino, R. Mechanisms and concepts in RNA virus population dynamics and evolution. Annu. Rev. Virol. 2018, 5, 69–92. [Google Scholar] [CrossRef]
- Gutierrez, B.; Escalera-Zamudio, M.; Pybus, O.G. Parallel molecular evolution and adaptation in viruses. Curr. Opin. Virol. 2019, 34, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Shukla, P.; Nguyen, H.T.; Torian, U.; Engle, R.E.; Faulk, K.; Dalton, H.R.; Bendall, R.P.; Keane, F.E.; Purcell, R.H.; Emerson, S.U. Cross-species infections of cultured cells by hepatitis E virus and discovery of an infectious virus-host recombinant. Proc. Natl. Acad. Sci. USA 2011, 108, 2438–2443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalton, H.R.; Bendall, R.P.; Keane, F.E.; Tedder, R.S.; Ijaz, S. Persistent carriage of hepatitis E virus in patients with HIV infection. N. Engl. J. Med. 2009, 361, 1025–1027. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, S.N.; Devhare, P.B.; Pingle, S.Y.; Paingankar, M.S.; Arankalle, V.A.; Lole, K.S. Hepatitis E virus (HEV)-1 harbouring HEV-4 non-structural protein (ORF1) replicates in transfected porcine kidney cells. J. Gen. Virol. 2016, 97, 1829–1840. [Google Scholar] [CrossRef] [Green Version]
- Bouquet, J.; Cherel, P.; Pavio, N. Genetic characterization and codon usage bias of full-length Hepatitis E virus sequences shed new lights on genotypic distribution, host restriction and genome evolution. Infect. Genet. Evol. 2012, 12, 1842–1853. [Google Scholar] [CrossRef] [PubMed]
- Baha, S.; Behloul, N.; Liu, Z.; Wei, W.; Shi, R.; Meng, J. Comprehensive analysis of genetic and evolutionary features of the hepatitis E virus. BMC Genom. 2019, 20, 790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.H.; Li, X.R.; Lan, X.; Han, S.H.; Wang, Y.N.; Hu, Y.H. The genetic divergences of codon usage shed new lights on transmission of hepatitis E virus from swine to human. Infect. Genet. Evol. 2019, 68, 23–29. [Google Scholar] [CrossRef]
- Sun, J.; Ren, C.; Huang, Y.; Chao, W.; Xie, F. The effects of synonymous codon usages on genotypic formation of open reading frames in hepatitis E virus. Infect. Genet. Evol. 2020, 85, 104450. [Google Scholar] [CrossRef] [PubMed]
- Cordoba, L.; Feagins, A.R.; Opriessnig, T.; Cossaboom, C.M.; Dryman, B.A.; Huang, Y.W.; Meng, X.J. Rescue of a genotype 4 human hepatitis E virus from cloned cDNA and characterization of intergenotypic chimeric viruses in cultured human liver cells and in pigs. J. Gen. Virol. 2012, 93 Pt 10, 2183–2194. [Google Scholar] [CrossRef] [Green Version]
- Purdy, M.A.; Lara, J.; Khudyakov, Y.E. The hepatitis E virus polyproline region is involved in viral adaptation. PLoS ONE 2012, 7, e35974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.H.; Purcell, R.H.; Emerson, S.U. A truncated ORF2 protein contains the most immunogenic site on ORF2: Antibody responses to non-vaccine sequences following challenge of vaccinated and non-vaccinated macaques with hepatitis E virus. Vaccine 2005, 23, 3157–3165. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.Z.; Miao, J.; Zhao, M.; Tang, M.; Yeo, A.E.; Yu, H.; Zhang, J.; Xia, N.S. Role of heat-shock protein 90 in hepatitis E virus capsid trafficking. J. Gen. Virol. 2010, 91 Pt 7, 1728–1736. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Chen, T.; Luo, Y.; Fan, J.; Zhang, M.; Zhao, Q.; Nan, Y.; Liu, B.; Zhou, E.M. Synthetic peptides containing three neutralizing epitopes of genotype 4 swine hepatitis E virus ORF2 induced protection against swine HEV infection in rabbit. Vaccines 2020, 8, 178. [Google Scholar] [CrossRef] [PubMed]
- Shukla, P.; Nguyen, H.T.; Faulk, K.; Mather, K.; Torian, U.; Engle, R.E.; Emerson, S.U. Adaptation of a genotype 3 hepatitis E virus to efficient growth in cell culture depends on an inserted human gene segment acquired by recombination. J. Virol. 2012, 86, 5697–5707. [Google Scholar] [CrossRef] [Green Version]
- Sooryanarain, H.; Meng, X.J. Swine hepatitis E virus: Cross-species infection, pork safety and chronic infection. Virus Res. 2020, 284, 197985. [Google Scholar] [CrossRef]
- Tian, D.; Yugo, D.M.; Kenney, S.P.; Lynn Heffron, C.; Opriessnig, T.; Karuppannan, A.K.; Bayne, J.; Halbur, P.G.; Meng, X.J. Dissecting the potential role of hepatitis E virus ORF1 nonstructural gene in cross-species infection by using intergenotypic chimeric viruses. J. Med. Virol. 2020, 92, 3563–3571. [Google Scholar] [CrossRef]
- Purdy, M.A.; Khudyakov, Y.E. Evolutionary history and population dynamics of hepatitis E virus. PLoS ONE 2010, 5, e14376. [Google Scholar] [CrossRef]
- Forni, D.; Cagliani, R.; Clerici, M.; Sironi, M. Origin and dispersal of Hepatitis E virus. Emerg. Microbes Infect. 2018, 7, 11. [Google Scholar] [CrossRef]
- Kumar, C.S.; Kumar, S. Species based synonymous codon usage in fusion protein gene of Newcastle disease virus. PLoS ONE 2014, 9, e114754. [Google Scholar] [CrossRef]
- Bahir, I.; Fromer, M.; Prat, Y.; Linial, M. Viral adaptation to host: A proteome-based analysis of codon usage and amino acid preferences. Mol. Syst. Biol. 2009, 5, 311. [Google Scholar] [CrossRef]
- Zhou, Z.; Dang, Y.; Zhou, M.; Li, L.; Yu, C.H.; Fu, J.; Chen, S.; Liu, Y. Codon usage is an important determinant of gene expression levels largely through its effects on transcription. Proc. Natl. Acad. Sci. USA 2016, 113, E6117–E6125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cannarozzi, G.; Schraudolph, N.N.; Faty, M.; von Rohr, P.; Friberg, M.T.; Roth, A.C.; Gonnet, P.; Gonnet, G.; Barral, Y. A role for codon order in translation dynamics. Cell 2010, 141, 355–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, M.; Son, H.S. Prediction of cross-species infection propensities of viruses with receptor similarity. Infect. Genet. Evol. 2019, 73, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.T.; Shukla, P.; Torian, U.; Faulk, K.; Emerson, S.U. Hepatitis E virus genotype 1 infection of swine kidney cells in vitro is inhibited at multiple levels. J. Virol. 2014, 88, 868–877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, S.; Miao, J.; Zheng, Z.; Wu, T.; Xie, M.; Tang, M.; Zhang, J.; Ng, M.H.; Xia, N. Putative receptor-binding sites of hepatitis E virus. J. Gen. Virol. 2008, 89 Pt 1, 245–249. [Google Scholar] [CrossRef]
- de Chassey, B.; Navratil, V.; Tafforeau, L.; Hiet, M.S.; Aublin-Gex, A.; Agaugue, S.; Meiffren, G.; Pradezynski, F.; Faria, B.F.; Chantier, T.; et al. Hepatitis C virus infection protein network. Mol. Syst. Biol. 2008, 4, 230. [Google Scholar] [CrossRef]
- Fernandez-Garcia, M.D.; Mazzon, M.; Jacobs, M.; Amara, A. Pathogenesis of flavivirus infections: Using and abusing the host cell. Cell Host Microbe 2009, 5, 318–328. [Google Scholar] [CrossRef] [PubMed]
- Panavas, T.; Serviene, E.; Brasher, J.; Nagy, P.D. Yeast genome-wide screen reveals dissimilar sets of host genes affecting replication of RNA viruses. Proc. Natl. Acad. Sci. USA 2005, 102, 7326–7331. [Google Scholar] [CrossRef] [Green Version]
- Wissing, M.H.; Bruggemann, Y.; Steinmann, E.; Todt, D. Virus-host cell interplay during hepatitis E virus infection. Trends Microbiol. 2021, 29, 309–319. [Google Scholar] [CrossRef]
- Nagashima, S.; Takahashi, M.; Jirintai, S.; Tanaka, T.; Nishizawa, T.; Yasuda, J.; Okamoto, H. Tumour susceptibility gene 101 and the vacuolar protein sorting pathway are required for the release of hepatitis E virions. J. Gen. Virol. 2011, 92 Pt 12, 2838–2848. [Google Scholar] [CrossRef]
- Simmonds, P.; Aiewsakun, P.; Katzourakis, A. Prisoners of war—Host adaptation and its constraints on virus evolution. Nat. Rev. Microbiol. 2019, 17, 321–328. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The genomic organization and translation of HEV. (A) Genome map of HEV. MeT, Methyltransferase; Y, Y domain; PCP, Papain-like cysteine protease; HVR, Hypervariable region; X, macro domain; Hel, Helicase; RdRp, RNA-dependent RNA polymerase; JR, junction region having the stem-loop structure and playing a critical role in HEV replication [19,20,21,26]. (B) A schematic representation of two major forms of ORF2 protein. The actual capsid protein (ORF2c) is initiated from an internal AUG (Met 16) located 16 codons downstream of the first AUG, while the secreted form of the ORF2 product (ORF2s) is initiated from the previously presumed start codon (Met 1) and its N-terminal 23 amino acids are cleaved by signal peptidase [30].
Figure 1.
The genomic organization and translation of HEV. (A) Genome map of HEV. MeT, Methyltransferase; Y, Y domain; PCP, Papain-like cysteine protease; HVR, Hypervariable region; X, macro domain; Hel, Helicase; RdRp, RNA-dependent RNA polymerase; JR, junction region having the stem-loop structure and playing a critical role in HEV replication [19,20,21,26]. (B) A schematic representation of two major forms of ORF2 protein. The actual capsid protein (ORF2c) is initiated from an internal AUG (Met 16) located 16 codons downstream of the first AUG, while the secreted form of the ORF2 product (ORF2s) is initiated from the previously presumed start codon (Met 1) and its N-terminal 23 amino acids are cleaved by signal peptidase [30].

Figure 3.
Alignment of the ORF3 amino acid sequences of Orthohepevirus A species showing the conserved PSAP motifs and CCC and IFI motifs. The PSAP motifs are known to be essential to the promotion of virion release [33], and the CCC and IFI motifs are reported to facilitate the virus release and ion fluxes [34].
Figure 3.
Alignment of the ORF3 amino acid sequences of Orthohepevirus A species showing the conserved PSAP motifs and CCC and IFI motifs. The PSAP motifs are known to be essential to the promotion of virion release [33], and the CCC and IFI motifs are reported to facilitate the virus release and ion fluxes [34].

Table 2.
Possible viral and host factors involved in cross-species transmission of HEV.
| Determinants | Remarks | References | |
|---|---|---|---|
| Viral factors | ORF1 (unknown domain) | Chimeric viruses where 5′ UTR, ORF1 and/or JR in the backbone of human HEV-4 were swapped with the corresponding regions of swine HEV-3 successfully infected pigs, suggesting that the 5′ UTR and ORF1 may be involved in cross-species transmission. | [111] |
| Only chimeras with the swine HEV-4 ORF1 region either alone or in combination with the 5′ UTR were able to infect pig kidney cells in vitro, supporting the possible role of ORF1 in cross-species transmission. | [156] | ||
| Adaptive evolution and codon usage | Observed bias against Sar-55 of HEV-1 ORF2 production in deer cells and its amelioration following the introduction of a short 5′ RNA sequence from the Kernow-C1 strain of HEV-3 suggests that the modulation of translation from closely spaced codons can differ significantly according to host species, and this difference may provide one mechanism for restricting the host range. | [154] | |
| A lower codon usage bias was observed for zoonotic HEV (HEV-3 and HEV-4). The codon adaptation index calculated with the general codon usage table for humans and swine indicates the good adaptation of HEV to its hosts. Thus, it can be assumed that the gene expression of human and zoonotic genotypes is very well adapted to the translational kinetics in humans. | [157] | ||
| Codon adaptation may be the essential factor in determining the viral host tropism. | [158] | ||
| The genotype-specific codon usage bias in HEV-1 is generally stronger than that of HEV-3 and HEV-4. Unlike the unique codon usage pattern of HEV-1, HEV-3 and HEV-4 strains derived from either humans or swine have more diverse codon usage patterns in ORFs. | [159] | ||
| In a correspondence analysis based on the relative synonymous codon usage data, the different HEV genotypes appeared to cluster (in particular, HEV-1, and HEV-3 and HEV-4), and based on ORF1, HEV-1 is clearly separated from the other groups, partially reflecting that HEV-1 is restricted to human hosts, while HEV-3 and HEV-4 strains were found in various animal species and were capable of cross-species transmission. | [160] | ||
| Host factors | Host cellular factors | The inability of several intergenotypic chimeras (with HEV-1 as the genomic backbone where various genomic regions were replaced with the corresponding regions of HEV-3 or HEV-4 to infect swine either in vitro or in vivo, excluding chimeras with swapped ORF1) showed that—other than the role of the viral factor itself—swine cells might lack the essential host factors required by HEV-1 to establish successful infection in pigs. In addition, it may also reflect the functional importance of species-specific protein-protein interactions during HEV replication. | [111,156,161] |
| The ability of human HEV-1 to infect pig kidney cells suggests that human and swine HEV might share at least one cell receptor. | [154] | ||
| Host immune status | The Kernow-C1 strain of HEV-3 isolated from an immunocompromised host (HIV-infected patient) demonstrated an extraordinary ability to infect cells from a broad spectrum of species ranging from rodents to primates. | [154] | |
| Most cases of human infection with rat HEV involved immunocompromised individuals. | [115,116] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Primadharsini, P.P.; Nagashima, S.; Okamoto, H. Mechanism of Cross-Species Transmission, Adaptive Evolution and Pathogenesis of Hepatitis E Virus. Viruses 2021, 13, 909. https://doi.org/10.3390/v13050909
AMA Style
Primadharsini PP, Nagashima S, Okamoto H. Mechanism of Cross-Species Transmission, Adaptive Evolution and Pathogenesis of Hepatitis E Virus. Viruses. 2021; 13(5):909. https://doi.org/10.3390/v13050909
Chicago/Turabian StylePrimadharsini, Putu Prathiwi, Shigeo Nagashima, and Hiroaki Okamoto. 2021. "Mechanism of Cross-Species Transmission, Adaptive Evolution and Pathogenesis of Hepatitis E Virus" Viruses 13, no. 5: 909. https://doi.org/10.3390/v13050909
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.