
Serological Humoral Immunity Following Natural Infection of Children with High Burden Gastrointestinal Viruses

 and
and
Abstract
:1. Introduction
2. Rotavirus
2.1. Rotavirus Structure and Antigenic Characteristics
2.2. Rotavirus Pathogenesis
2.3. Mechanisms of Protection against Rotavirus Infection and Disease
2.4. Host Genetics and Health Outcomes
2.5. Correlates of Rotavirus Immunity and Protection in Children
2.6. Impact of Rotavirus Vaccination on Disease Burden in Children
2.7. Conclusions and Next Steps
3. Norovirus
3.1. Norovirus Burden of Disease
3.2. Virus Background
3.3. Correlates of Protection
3.4. Vaccine Development
4. Sapovirus
4.1. Burden of Disease across the Ages and State of Vaccine Development
4.2. Virus Structure and Genetic Diversity
4.3. Health Outcomes/Host Population
4.4. Immune Protection against Sapovirus
4.5. Early Knowledge about Viral Epitopes
4.6. What We Need to Move Forward with Better Understanding of Immunity
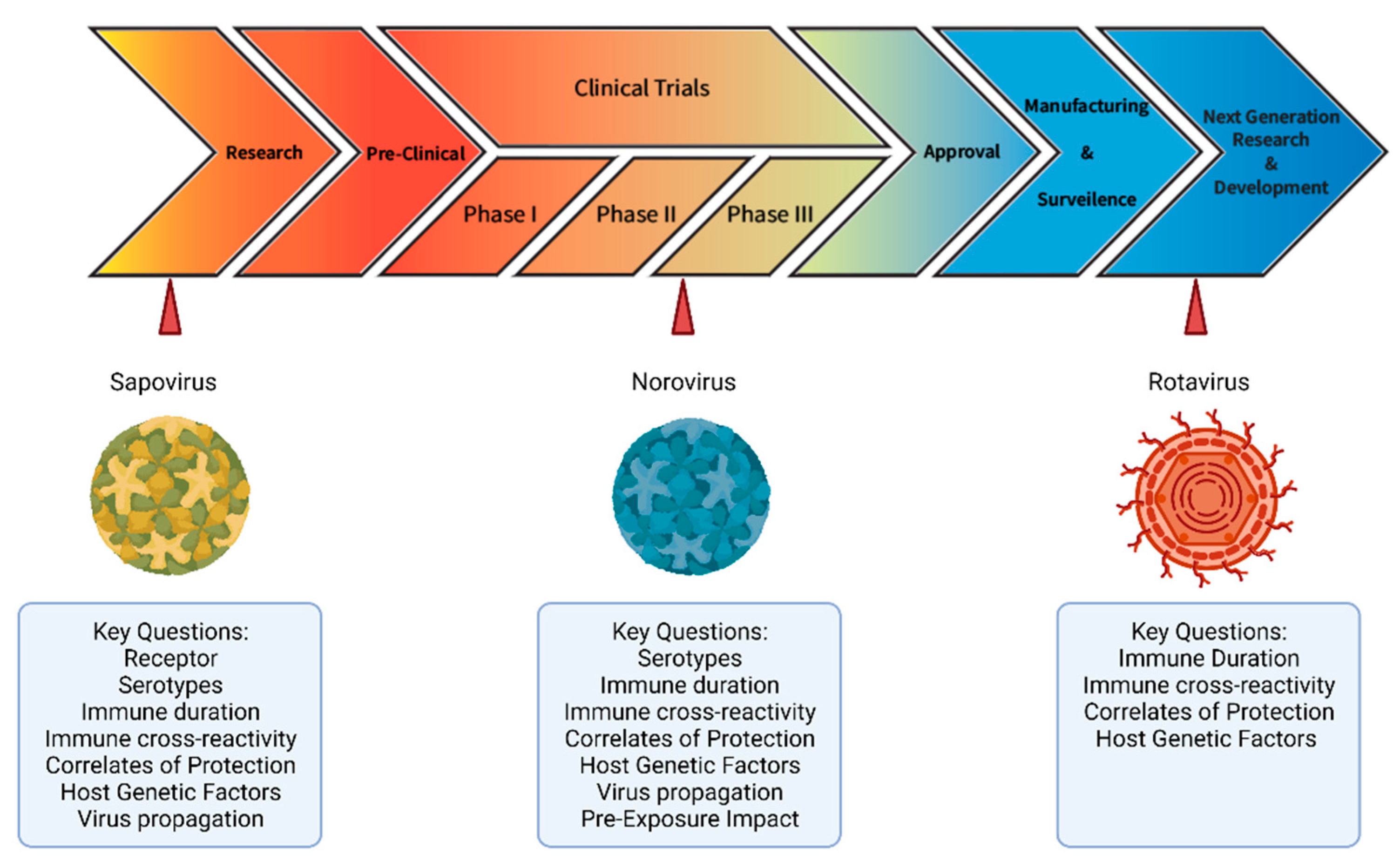
5. Conclusions and Key Questions Moving Forward

{kind=link}
| Virus | Correlate | Infection | Symptoms | Reference |
|---|---|---|---|---|
| Rotavirus | Rotavirus IgA and IgG in serum | X | X | [86,91,247] |
| Genotype specific IgA and IgG in serum | X | X | [85] | |
| Rotavirus IgA in stools | X | X | [248,249] | |
| VP6 serum and fecal IgA | X | [45,46,250] | ||
| NSP4 IgG | X | [42,93,94] | ||
| Human Norovirus | Salivary IgA | X | X | [61,180] |
| Memory IgG cells | X | [39,89] | ||
| Blockade Ab titer | X | X | [173,179,182,190] | |
| Hemagglutination Inhibition titer | X | X | [251] | |
| Serum IgA | X | X | [179] | |
| Sapovirus | Serum Ig | X | [234] |
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Platts-Mills, J.A.; Liu, J.; Rogawski, E.T.; Kabir, F.; Lertsethtakarn, P.; Siguas, M.; Khan, S.S.; Praharaj, I.; Murei, A.; Nshama, R.; et al. Use of quantitative molecular diagnostic methods to assess the aetiology, burden, and clinical characteristics of diarrhoea in children in low-resource settings: A reanalysis of the MAL-ED cohort study. Lancet Glob. Health 2018, 6, e1309–e1318. [Google Scholar] [CrossRef] [Green Version]
- Burnett, E.; Parashar, U.D.; Tate, J.E. Global Impact of Rotavirus Vaccination on Diarrhea Hospitalizations and Deaths Among Children <5 Years Old: 2006–2019. J. Infect. Dis. 2020, 222, 1731–1739. [Google Scholar] [CrossRef]
- Zhang, M.; Fu, M.; Hu, Q. Advances in Human Norovirus Vaccine Research. Vaccines 2021, 9, 732. [Google Scholar] [CrossRef] [PubMed]
- Nordgren, J.; Sharma, S.; Bucardo, F.; Nasir, W.; Gunaydin, G.; Ouermi, D.; Nitiema, L.W.; Becker-Dreps, S.; Simpore, J.; Hammarstrom, L.; et al. Both Lewis and secretor status mediate susceptibility to rotavirus infections in a rotavirus genotype-dependent manner. Clin. Infect. Dis. 2014, 59, 1567–1573. [Google Scholar] [CrossRef] [Green Version]
- Velázquez, F.R.; Matson, D.O.; Calva, J.J.; Guerrero, L.; Morrow, A.L.; Carter-Campbell, S.; Glass, R.I.; Estes, M.K.; Pickering, L.K.; Ruiz-Palacios, G.M. Rotavirus infection in infants as protection against subsequent infections. N. Engl. J. Med. 1996, 335, 1022–1028. [Google Scholar] [CrossRef]
- Ruvoën-Clouet, N.; Belliot, G.; Le Pendu, J. Noroviruses and histo-blood groups: The impact of common host genetic polymorphisms on virus transmission and evolution. Rev. Med. Virol. 2013, 23, 355–366. [Google Scholar] [CrossRef]
- Gozalbo-Rovira, R.; Ciges-Tomas, J.R.; Vila-Vicent, S.; Buesa, J.; Santiso-Bellon, C.; Monedero, V.; Yebra, M.J.; Marina, A.; Rodriguez-Diaz, J. Unraveling the role of the secretor antigen in human rotavirus attachment to histo-blood group antigens. PLoS Pathog. 2019, 15, e1007865. [Google Scholar] [CrossRef]
- Reyes, Y.; González, F.; Gutierrez, L.; Blandon, P.; Centeno, E.; Zepeda, O.; Toval-Ruíz, C.; Lindesmith, L.C.; Baric, R.S.; Vielot, N.; et al. Secretor status strongly influences the incidence of symptomatic norovirus infection in a genotype-dependent manner in a Nicaraguan birth cohort. J. Infect. Dis. 2021, jiab316. [Google Scholar] [CrossRef]
- Bucardo, F.; Reyes, Y.; Ronnelid, Y.; Gonzalez, F.; Sharma, S.; Svensson, L.; Nordgren, J. Histo-blood group antigens and rotavirus vaccine shedding in Nicaraguan infants. Sci. Rep. 2019, 9, 10764. [Google Scholar] [CrossRef] [PubMed]
- Jayaram, H.; Estes, M.K.; Prasad, B.V. Emerging themes in rotavirus cell entry, genome organization, transcription and replication. Virus Res. 2004, 101, 67–81. [Google Scholar] [CrossRef] [PubMed]
- Afchangi, A.; Jalilvand, S.; Mohajel, N.; Marashi, S.M.; Shoja, Z. Rotavirus VP6 as a potential vaccine candidate. Rev. Med. Virol. 2019, 29, e2027. [Google Scholar] [CrossRef]
- Hoshino, Y.; Kapikian, A.Z. Rotavirus serotypes: Classification and importance in epidemiology, immunity, and vaccine development. J. Health Popul. Nutr. 2000, 18, 5–14. [Google Scholar]
- Clarke, E.; Desselberger, U. Correlates of protection against human rotavirus disease and the factors influencing protection in low-income settings. Mucosal. Immunol. 2015, 8, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arias, C.F.; Lopez, S. Rotavirus cell entry: Not so simple after all. Curr. Opin. Virol. 2021, 48, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Matthijnssens, J.; Heylen, E.; Zeller, M.; Rahman, M.; Lemey, P.; Van Ranst, M. Phylodynamic analyses of rotavirus genotypes G9 and G12 underscore their potential for swift global spread. Mol. Biol. Evol. 2010, 27, 2431–2436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jain, S.; Vashistt, J.; Changotra, H. Rotaviruses: Is their surveillance needed? Vaccine 2014, 32, 3367–3378. [Google Scholar] [CrossRef]
- Amit, L.N.; Mori, D.; John, J.L.; Chin, A.Z.; Mosiun, A.K.; Jeffree, M.S.; Ahmed, K. Emergence of equine-like G3 strains as the dominant rotavirus among children under five with diarrhea in Sabah, Malaysia during 2018–2019. PLoS ONE 2021, 16, e0254784. [Google Scholar] [CrossRef] [PubMed]
- Bonura, F.; Banyai, K.; Mangiaracina, L.; Bonura, C.; Martella, V.; Giammanco, G.M.; De Grazia, S. Emergence in 2017-2019 of novel reassortant equine-like G3 rotavirus strains in Palermo, Sicily. Transbound. Emerg. Dis. 2021. [Google Scholar] [CrossRef]
- Akane, Y.; Tsugawa, T.; Fujii, Y.; Honjo, S.; Kondo, K.; Nakata, S.; Fujibayashi, S.; Ohara, T.; Mori, T.; Higashidate, Y.; et al. Molecular and clinical characterization of the equine-like G3 rotavirus that caused the first outbreak in Japan, 2016. J. Gen. Virol. 2021, 102, 001548. [Google Scholar] [CrossRef]
- Athiyyah, A.F.; Utsumi, T.; Wahyuni, R.M.; Dinana, Z.; Yamani, L.N.; Sudarmo, S.M.; Ranuh, R.G.; Darma, A.; Raharjo, D.; Matsui, C.; et al. Molecular Epidemiology and Clinical Features of Rotavirus Infection Among Pediatric Patients in East Java, Indonesia During 2015–2018: Dynamic Changes in Rotavirus Genotypes from Equine-Like G3 to Typical Human G1/G3. Front. Microbiol. 2019, 10, 940. [Google Scholar] [CrossRef] [Green Version]
- Utsumi, T.; Wahyuni, R.M.; Doan, Y.H.; Dinana, Z.; Soegijanto, S.; Fujii, Y.; Yamani, L.N.; Matsui, C.; Deng, L.; Abe, T.; et al. Equine-like G3 rotavirus strains as predominant strains among children in Indonesia in 2015–2016. Infect. Genet. Evol. 2018, 61, 224–228. [Google Scholar] [CrossRef] [Green Version]
- Pietsch, C.; Liebert, U.G. Molecular characterization of different equine-like G3 rotavirus strains from Germany. Infect. Genet. Evol. 2018, 57, 46–50. [Google Scholar] [CrossRef]
- Guerra, S.F.S.; Soares, L.S.; Lobo, P.S.; Penha Junior, E.T.; Sousa Junior, E.C.; Bezerra, D.A.M.; Vaz, L.R.; Linhares, A.C.; Mascarenhas, J.D.P. Detection of a novel equine-like G3 rotavirus associated with acute gastroenteritis in Brazil. J. Gen. Virol. 2016, 97, 3131–3138. [Google Scholar] [CrossRef] [PubMed]
- Doro, R.; Marton, S.; Bartokne, A.H.; Lengyel, G.; Agocs, Z.; Jakab, F.; Banyai, K. Equine-like G3 rotavirus in Hungary, 2015—Is it a novel intergenogroup reassortant pandemic strain? Acta Microbiol. Immunol. Hung. 2016, 63, 243–255. [Google Scholar] [CrossRef] [Green Version]
- Hoa-Tran, T.N.; Nakagomi, T.; Vu, H.M.; Do, L.P.; Gauchan, P.; Agbemabiese, C.A.; Nguyen, T.T.; Nakagomi, O.; Thanh, N.T. Abrupt emergence and predominance in Vietnam of rotavirus A strains possessing a bovine-like G8 on a DS-1-like background. Arch. Virol. 2016, 161, 479–482. [Google Scholar] [CrossRef]
- Jagannath, M.R.; Vethanayagam, R.R.; Reddy, B.S.; Raman, S.; Rao, C.D. Characterization of human symptomatic rotavirus isolates MP409 and MP480 having ‘long’ RNA electropherotype and subgroup I specificity, highly related to the P6[1], G8 type bovine rotavirus A5, from Mysore, India. Arch. Virol. 2000, 145, 1339–1357. [Google Scholar] [CrossRef] [Green Version]
- da Silva, M.F.; Tort, L.F.; Gomez, M.M.; Assis, R.M.; de Mendonca, M.C.; Volotao Ede, M.; Leite, J.P. Phylogenetic analysis of VP1, VP2, and VP3 gene segments of genotype G5 group A rotavirus strains circulating in Brazil between 1986 and 2005. Virus Res. 2011, 160, 381–388. [Google Scholar] [CrossRef]
- Lundgren, O.; Svensson, L. Pathogenesis of rotavirus diarrhea. Microbes Infect. 2001, 3, 1145–1156. [Google Scholar] [CrossRef]
- Crawford, S.E.; Ramani, S.; Tate, J.E.; Parashar, U.D.; Svensson, L.; Hagbom, M.; Franco, M.A.; Greenberg, H.B.; O’Ryan, M.; Kang, G.; et al. Rotavirus infection. Nat. Rev. Dis. Primers 2017, 3, 17083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hellysaz, A.; Hagbom, M. Understanding the Central Nervous System Symptoms of Rotavirus: A Qualitative Review. Viruses 2021, 13, 658. [Google Scholar] [CrossRef] [PubMed]
- Hagbom, M.; Hellysaz, A.; Istrate, C.; Nordgren, J.; Sharma, S.; de-Faria, F.M.; Magnusson, K.E.; Svensson, L. The 5-HT3 Receptor Affects Rotavirus-Induced Motility. J. Virol. 2021, 95, e0075121. [Google Scholar] [CrossRef] [PubMed]
- Istrate, C.; Hagbom, M.; Vikstrom, E.; Magnusson, K.E.; Svensson, L. Rotavirus infection increases intestinal motility but not permeability at the onset of diarrhea. J. Virol. 2014, 88, 3161–3169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hemming, M.; Huhti, L.; Rasanen, S.; Salminen, M.; Vesikari, T. Rotavirus antigenemia in children is associated with more severe clinical manifestations of acute gastroenteritis. Pediatr. Infect. Dis. J. 2014, 33, 366–371. [Google Scholar] [CrossRef] [PubMed]
- Moon, S.; Wang, Y.; Dennehy, P.; Simonsen, K.A.; Zhang, J.; Jiang, B. Antigenemia, RNAemia, and innate immunity in children with acute rotavirus diarrhea. FEMS Immunol. Med. Microbiol. 2012, 64, 382–391. [Google Scholar] [CrossRef] [Green Version]
- Yu, T.H.; Tsai, C.N.; Lai, M.W.; Chen, C.C.; Chao, H.C.; Lin, C.W.; Chiu, C.H.; Chen, S.Y. Antigenemia and cytokine expression in rotavirus gastroenteritis in children. J. Microbiol Immunol. Infect. 2012, 45, 265–270. [Google Scholar] [CrossRef] [Green Version]
- Sugata, K.; Taniguchi, K.; Yui, A.; Miyake, F.; Suga, S.; Asano, Y.; Ohashi, M.; Suzuki, K.; Nishimura, N.; Ozaki, T.; et al. Analysis of rotavirus antigenemia and extraintestinal manifestations in children with rotavirus gastroenteritis. Pediatrics 2008, 122, 392–397. [Google Scholar] [CrossRef] [Green Version]
- Justino, M.C.A.; Campos, E.A.; Mascarenhas, J.D.P.; Soares, L.S.; Guerra, S.F.S.; Furlaneto, I.P.; Pavao, M.J.C., Jr.; Maciel, T.S.; Farias, F.P.; Bezerra, O.M.; et al. Rotavirus antigenemia as a common event among children hospitalised for severe, acute gastroenteritis in Belem, northern Brazil. BMC Pediatrics 2019, 19, 193. [Google Scholar] [CrossRef]
- Ward, R. Mechanisms of protection against rotavirus infection and disease. Pediatr. Infect. Dis. J. 2009, 28, S57–S59. [Google Scholar] [CrossRef]
- McNeal, M.M.; Barone, K.S.; Rae, M.N.; Ward, R.L. Effector functions of antibody and CD8+ cells in resolution of rotavirus infection and protection against reinfection in mice. Virology 1995, 214, 387–397. [Google Scholar] [CrossRef] [Green Version]
- Jaimes, M.C.; Feng, N.; Greenberg, H.B. Characterization of homologous and heterologous rotavirus-specific T-cell responses in infant and adult mice. J. Virol. 2005, 79, 4568–4579. [Google Scholar] [CrossRef] [Green Version]
- Davis, S.K.; Selva, K.J.; Kent, S.J.; Chung, A.W. Serum IgA Fc effector functions in infectious disease and cancer. Immunol. Cell Biol. 2020, 98, 276–286. [Google Scholar] [CrossRef] [Green Version]
- Iosef, C.; Chang, K.O.; Azevedo, M.S.; Saif, L.J. Systemic and intestinal antibody responses to NSP4 enterotoxin of Wa human rotavirus in a gnotobiotic pig model of human rotavirus disease. J. Med. Virol. 2002, 68, 119–128. [Google Scholar] [CrossRef]
- Yeom, J.S.; Kim, Y.S.; Jun, J.S.; Do, H.J.; Park, J.S.; Seo, J.H.; Park, E.S.; Lim, J.Y.; Woo, H.O.; Park, C.H.; et al. NSP4 antibody levels in rotavirus gastroenteritis patients with seizures. Eur. J. Paediatr. Neurol. EJPN Off. J. Eur. Paediatr. Neurol. Soc. 2017, 21, 367–373. [Google Scholar] [CrossRef]
- Vizzi, E.; Calvino, E.; Gonzalez, R.; Perez-Schael, I.; Ciarlet, M.; Kang, G.; Estes, M.K.; Liprandi, F.; Ludert, J.E. Evaluation of serum antibody responses against the rotavirus nonstructural protein NSP4 in children after rhesus rotavirus tetravalent vaccination or natural infection. Clin. Diagn. Lab. Immunol. 2005, 12, 1157–1163. [Google Scholar] [CrossRef] [Green Version]
- Caddy, S.L.; Vaysburd, M.; Wing, M.; Foss, S.; Andersen, J.T.; O’Connell, K.; Mayes, K.; Higginson, K.; Iturriza-Gomara, M.; Desselberger, U.; et al. Intracellular neutralisation of rotavirus by VP6-specific IgG. PLoS Pathog. 2020, 16, e1008732. [Google Scholar] [CrossRef]
- Lappalainen, S.; Pastor, A.R.; Tamminen, K.; Lopez-Guerrero, V.; Esquivel-Guadarrama, F.; Palomares, L.A.; Vesikari, T.; Blazevic, V. Immune responses elicited against rotavirus middle layer protein VP6 inhibit viral replication in vitro and in vivo. Hum. Vaccines Immunother. 2014, 10, 2039–2047. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, R.; Nandi, S.; Lo, M.; Gope, A.; Chawla-Sarkar, M. Viperin, an IFN-Stimulated Protein, Delays Rotavirus Release by Inhibiting Non-Structural Protein 4 (NSP4)-Induced Intrinsic Apoptosis. Viruses 2021, 13, 1324. [Google Scholar] [CrossRef] [PubMed]
- Malm, M.; Hyöty, H.; Knip, M.; Vesikari, T.; Blazevic, V. Development of T cell immunity to norovirus and rotavirus in children under five years of age. Sci. Rep. 2019, 9, 3199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makela, M.; Marttila, J.; Simell, O.; Ilonen, J. Rotavirus-specific T-cell responses in young prospectively followed-up children. Clin. Exp. Immunol. 2004, 137, 173–178. [Google Scholar] [CrossRef] [PubMed]
- Offit, P.A.; Hoffenberg, E.J.; Santos, N.; Gouvea, V. Rotavirus-specific humoral and cellular immune response after primary, symptomatic infection. J. Infect. Dis. 1993, 167, 1436–1440. [Google Scholar] [CrossRef] [PubMed]
- Payne, D.C.; Currier, R.L.; Staat, M.A.; Sahni, L.C.; Selvarangan, R.; Halasa, N.B.; Englund, J.A.; Weinberg, G.A.; Boom, J.A.; Szilagyi, P.G.; et al. Epidemiologic Association Between FUT2 Secretor Status and Severe Rotavirus Gastroenteritis in Children in the United States. JAMA Pediatrics 2015, 169, 1040–1045. [Google Scholar] [CrossRef] [Green Version]
- Fix, J.; Chandrashekhar, K.; Perez, J.; Bucardo, F.; Hudgens, M.G.; Yuan, L.; Twitchell, E.; Azcarate-Peril, M.A.; Vilchez, S.; Becker-Dreps, S. Association between Gut Microbiome Composition and Rotavirus Vaccine Response among Nicaraguan Infants. Am. J. Trop. Med. Hyg. 2020, 102, 213–219. [Google Scholar] [CrossRef]
- Mwila, K.; Chilengi, R.; Simuyandi, M.; Permar, S.R.; Becker-Dreps, S. Contribution of Maternal Immunity to Decreased Rotavirus Vaccine Performance in Low- and Middle-Income Countries. Clin. Vaccine Immunol. CVI 2017, 24, e00405-1. [Google Scholar] [CrossRef] [Green Version]
- Monedero, V.; Buesa, J.; Rodriguez-Diaz, J. The Interactions between Host Glycobiology, Bacterial Microbiota, and Viruses in the Gut. Viruses 2018, 10, 96. [Google Scholar] [CrossRef] [Green Version]
- Becker-Dreps, S.; Vilchez, S.; Bucardo, F.; Twitchell, E.; Choi, W.S.; Hudgens, M.G.; Perez, J.; Yuan, L. The Association Between Fecal Biomarkers of Environmental Enteropathy and Rotavirus Vaccine Response in Nicaraguan Infants. Pediatr. Infect. Dis. J. 2017, 36, 412–416. [Google Scholar] [CrossRef]
- Vlasova, A.N.; Takanashi, S.; Miyazaki, A.; Rajashekara, G.; Saif, L.J. How the gut microbiome regulates host immune responses to viral vaccines. Curr. Opin. Virol. 2019, 37, 16–25. [Google Scholar] [CrossRef]
- Desselberger, U. Differences of Rotavirus Vaccine Effectiveness by Country: Likely Causes and Contributing Factors. Pathogens 2017, 6, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, V.C.; Armah, G.; Fuentes, S.; Korpela, K.E.; Parashar, U.; Victor, J.C.; Tate, J.; de Weerth, C.; Giaquinto, C.; Wiersinga, W.J.; et al. Significant Correlation Between the Infant Gut Microbiome and Rotavirus Vaccine Response in Rural Ghana. J. Infect. Dis. 2017, 215, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Harris, V.; Ali, A.; Fuentes, S.; Korpela, K.; Kazi, M.; Tate, J.; Parashar, U.; Wiersinga, W.J.; Giaquinto, C.; de Weerth, C.; et al. Rotavirus vaccine response correlates with the infant gut microbiota composition in Pakistan. Gut Microbes 2018, 9, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Nordgren, J.; Svensson, L. Genetic Susceptibility to Human Norovirus Infection: An Update. Viruses 2019, 11, 226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindesmith, L.; Moe, C.; Marionneau, S.; Ruvoen, N.; Jiang, X.; Lindblad, L.; Stewart, P.; LePendu, J.; Baric, R. Human susceptibility and resistance to Norwalk virus infection. Nat. Med. 2003, 9, 548–553. [Google Scholar] [CrossRef]
- Frenck, R.; Bernstein, D.I.; Xia, M.; Huang, P.; Zhong, W.; Parker, S.; Dickey, M.; McNeal, M.; Jiang, X. Predicting susceptibility to norovirus GII.4 by use of a challenge model involving humans. J. Infect. Dis. 2012, 206, 1386–1393. [Google Scholar] [CrossRef] [Green Version]
- Tan, M.; Jiang, X. Histo-blood group antigens: A common niche for norovirus and rotavirus. Expert Rev. Mol. Med. 2014, 16, e5. [Google Scholar] [CrossRef]
- Hu, L.; Crawford, S.E.; Czako, R.; Cortes-Penfield, N.W.; Smith, D.F.; Le Pendu, J.; Estes, M.K.; Prasad, B.V. Cell attachment protein VP8* of a human rotavirus specifically interacts with A-type histo-blood group antigen. Nature 2012, 485, 256–259. [Google Scholar] [CrossRef]
- Huang, P.; Xia, M.; Tan, M.; Zhong, W.; Wei, C.; Wang, L.; Morrow, A.; Jiang, X. Spike protein VP8* of human rotavirus recognizes histo-blood group antigens in a type-specific manner. J. Virol. 2012, 86, 4833–4843. [Google Scholar] [CrossRef] [Green Version]
- Xu, S.; McGinnis, K.R.; Liu, Y.; Huang, P.; Tan, M.; Stuckert, M.R.; Burnside, R.E.; Jacob, E.G.; Ni, S.; Jiang, X.; et al. Structural basis of P[II] rotavirus evolution and host ranges under selection of histo-blood group antigens. Proc. Natl. Acad. Sci. USA 2021, 118. [Google Scholar] [CrossRef]
- Sharma, S.; Hagbom, M.; Svensson, L.; Nordgren, J. The Impact of Human Genetic Polymorphisms on Rotavirus Susceptibility, Epidemiology, and Vaccine Take. Viruses 2020, 12, 324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farahmand, M.; Jalilvand, S.; Arashkia, A.; Shahmahmoodi, S.; Afchangi, A.; Mollaei-Kandelous, Y.; Shoja, Z. Association between circulating rotavirus genotypes and histo-blood group antigens in the children hospitalized with acute gastroenteritis in Iran. J. Med. Virol. 2021, 93, 4817–4823. [Google Scholar] [CrossRef] [PubMed]
- Perez-Ortin, R.; Vila-Vicent, S.; Carmona-Vicente, N.; Santiso-Bellon, C.; Rodriguez-Diaz, J.; Buesa, J. Histo-Blood Group Antigens in Children with Symptomatic Rotavirus Infection. Viruses 2019, 11, 339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.K.; Oh, S.J.; Choi, S.; Choi, S.H.; Shin, S.H.; Lee, E.J.; Cho, E.J.; Hyun, J.; Kim, H.S. Relationship Between Rotavirus P[6] Infection in Korean Neonates and Histo-Blood Group Antigen: A Single-Center Study. Ann. Lab. Med. 2021, 41, 181–189. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, J.; Groome, M.J.; Mans, J.; Page, N. Fut2 Secretor Status Influences Susceptibility to Vp4 Strain-Specific Rotavirus Infections in South African Children. Pathogens 2020, 9, 795. [Google Scholar] [CrossRef]
- Ouermi, D.; Soubeiga, D.; Nadembega, W.M.C.; Sawadogo, P.M.; Zohoncon, T.M.; Obiri-Yeboah, D.; Djigma, F.W.; Nordgren, J.; Simpore, J. Molecular Epidemiology of Rotavirus in Children under Five in Africa (2006–2016): A Systematic Review. Pak. J. Biol. Sci. 2017, 20, 59–69. [Google Scholar] [CrossRef]
- Hu, L.; Sankaran, B.; Laucirica, D.R.; Patil, K.; Salmen, W.; Ferreon, A.C.M.; Tsoi, P.S.; Lasanajak, Y.; Smith, D.F.; Ramani, S.; et al. Glycan recognition in globally dominant human rotaviruses. Nat. Commun. 2018, 9, 2631. [Google Scholar] [CrossRef] [Green Version]
- Papp, H.; Borzak, R.; Farkas, S.; Kisfali, P.; Lengyel, G.; Molnar, P.; Melegh, B.; Matthijnssens, J.; Jakab, F.; Martella, V.; et al. Zoonotic transmission of reassortant porcine G4P[6] rotaviruses in Hungarian pediatric patients identified sporadically over a 15 year period. Infect. Genet. Evol. 2013, 19, 71–80. [Google Scholar] [CrossRef]
- Zhou, X.; Wang, Y.H.; Ghosh, S.; Tang, W.F.; Pang, B.B.; Liu, M.Q.; Peng, J.S.; Zhou, D.J.; Kobayashi, N. Genomic characterization of G3P[6], G4P[6] and G4P[8] human rotaviruses from Wuhan, China: Evidence for interspecies transmission and reassortment events. Infect. Genet. Evol. 2015, 33, 55–71. [Google Scholar] [CrossRef]
- Malasao, R.; Khamrin, P.; Kumthip, K.; Ushijima, H.; Maneekarn, N. Complete genome sequence analysis of rare G4P[6] rotavirus strains from human and pig reveals the evidence for interspecies transmission. Infect. Genet. Evol. 2018, 65, 357–368. [Google Scholar] [CrossRef]
- Tacharoenmuang, R.; Guntapong, R.; Upachai, S.; Singchai, P.; Fukuda, S.; Ide, T.; Hatazawa, R.; Sutthiwarakom, K.; Kongjorn, S.; Onvimala, N.; et al. Full genome-based characterization of G4P[6] rotavirus strains from diarrheic patients in Thailand: Evidence for independent porcine-to-human interspecies transmission events. Virus Genes 2021, 57, 338–357. [Google Scholar] [CrossRef] [PubMed]
- Mokoena, F.; Esona, M.D.; Seheri, L.M.; Nyaga, M.M.; Magagula, N.B.; Mukaratirwa, A.; Mulindwa, A.; Abebe, A.; Boula, A.; Tsolenyanu, E.; et al. Whole Genome Analysis of African G12P[6] and G12P[8] Rotaviruses Provides Evidence of Porcine-Human Reassortment at NSP2, NSP3, and NSP4. Front. Microbiol. 2020, 11, 604444. [Google Scholar] [CrossRef] [PubMed]
- Bucardo, F.; Gonzalez, F.; Reyes, Y.; Blandon, P.; Saif, L.; Nordgren, J. Seroprevalence in Household Raised Pigs Indicate High Exposure to GII Noroviruses in Rural Nicaragua. Zoonoses Public Health 2016, 63, 600–607. [Google Scholar] [CrossRef]
- Armah, G.E.; Cortese, M.M.; Dennis, F.E.; Yu, Y.; Morrow, A.L.; McNeal, M.M.; Lewis, K.D.C.; Awuni, D.A.; Armachie, J.; Parashar, U.D. Rotavirus Vaccine Take in Infants Is Associated with Secretor Status. J. Infect. Dis. 2019, 219, 746–749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Günaydın, G.; Nordgren, J.; Sharma, S.; Hammarström, L. Association of elevated rotavirus-specific antibody titers with HBGA secretor status in Swedish individuals: The FUT2 gene as a putative susceptibility determinant for infection. Virus Res. 2016, 211, 64–68. [Google Scholar] [CrossRef]
- Kazi, A.M.; Cortese, M.M.; Yu, Y.; Lopman, B.; Morrow, A.L.; Fleming, J.A.; McNeal, M.M.; Steele, A.D.; Parashar, U.D.; Zaidi, A.K.M.; et al. Secretor and Salivary ABO Blood Group Antigen Status Predict Rotavirus Vaccine Take in Infants. J. Infect. Dis. 2017, 215, 786–789. [Google Scholar] [CrossRef] [Green Version]
- Bennett, A.; Pollock, L.; Jere, K.C.; Pitzer, V.E.; Parashar, U.; Tate, J.E.; Heyderman, R.S.; Mwansambo, C.; French, N.; Nakagomi, O.; et al. Direct and possible indirect effects of vaccination on rotavirus hospitalisations among children in Malawi four years after programmatic introduction. Vaccine 2018, 36, 7142–7148. [Google Scholar] [CrossRef] [PubMed]
- Bucardo, F.; Nordgren, J.; Reyes, Y.; Gonzalez, F.; Sharma, S.; Svensson, L. The Lewis A phenotype is a restriction factor for Rotateq and Rotarix vaccine-take in Nicaraguan children. Sci. Rep. 2018, 9, 1502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Ryan, M.L.; Matson, D.O.; Estes, M.K.; Pickering, L.K. Anti-rotavirus G type-specific and isotype-specific antibodies in children with natural rotavirus infections. J. Infect. Dis. 1994, 169, 504–511. [Google Scholar] [CrossRef]
- Velazquez, F.R.; Matson, D.O.; Guerrero, M.L.; Shults, J.; Calva, J.J.; Morrow, A.L.; Glass, R.I.; Pickering, L.K.; Ruiz-Palacios, G.M. Serum antibody as a marker of protection against natural rotavirus infection and disease. J. Infect. Dis. 2000, 182, 1602–1609. [Google Scholar] [CrossRef] [Green Version]
- Angel, J.; Steele, A.D.; Franco, M.A. Correlates of protection for rotavirus vaccines: Possible alternative trial endpoints, opportunities, and challenges. Hum. Vaccines Immunother. 2014, 10, 3659–3671. [Google Scholar] [CrossRef] [Green Version]
- Patton, J.T. Rotavirus diversity and evolution in the post-vaccine world. Discov. Med. 2012, 13, 85–97. [Google Scholar] [PubMed]
- Yuan, L.; Saif, L.J. Induction of mucosal immune responses and protection against enteric viruses: Rotavirus infection of gnotobiotic pigs as a model. Vet. Immunol. Immunopathol. 2002, 87, 147–160. [Google Scholar] [CrossRef]
- Lewnard, J.A.; Lopman, B.A.; Parashar, U.D.; Bar-Zeev, N.; Samuel, P.; Guerrero, M.L.; Ruiz-Palacios, G.M.; Kang, G.; Pitzer, V.E. Naturally Acquired Immunity Against Rotavirus Infection and Gastroenteritis in Children: Paired Reanalyses of Birth Cohort Studies. J. Infect. Dis. 2017, 216, 317–326. [Google Scholar] [CrossRef] [Green Version]
- Patel, M.; Glass, R.I.; Jiang, B.; Santosham, M.; Lopman, B.; Parashar, U. A systematic review of anti-rotavirus serum IgA antibody titer as a potential correlate of rotavirus vaccine efficacy. J. Infect. Dis. 2013, 208, 284–294. [Google Scholar] [CrossRef] [Green Version]
- Hou, Z.; Huang, Y.; Huan, Y.; Pang, W.; Meng, M.; Wang, P.; Yang, M.; Jiang, L.; Cao, X.; Wu, K.K. Anti-Nsp4 Antibody Can Block Rotavirus-Induced Diarrhea in Mice. J. Pediatr. Gastroenterol. Nutr. 2008, 46, 376–385. [Google Scholar] [CrossRef]
- Rodríguez-Díaz, J.; Montava, R.; García-Díaz, A.; Buesa, J. Humoral immune response to rotavirus NSP4 enterotoxin in Spanish children. J. Med. Virol. 2005, 77, 317–322. [Google Scholar] [CrossRef]
- Johansen, K.; Hinkula, J.; Espinoza, F.; Levi, M.; Zeng, C.; Rudén, U.; Vesikari, T.; Estes, M.; Svensson, L. Humoral and cell-mediated immune responses in humans to the NSP4 enterotoxin of rotavirus. J. Med. Virol. 1999, 59, 369–377. [Google Scholar] [CrossRef]
- Ray, P.; Malik, J.; Singh, R.K.; Bhatnagar, S.; Bahl, R.; Kumar, R.; Bhan, M.K. Rotavirus nonstructural protein NSP4 induces heterotypic antibody responses during natural infection in children. J. Infect. Dis. 2003, 187, 1786–1793. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Fu, Y.; Sun, J.; Zhang, J.; Feng, M.; Cheng, X.; Xu, J. Analysis of rotavirus NSP4 genotypes and age-dependent antibody response against NSP4 in Shanghai, China. Jpn. J. Infect. Dis. 2010, 63, 280–282. [Google Scholar] [PubMed]
- Nair, N.; Feng, N.; Blum, L.K.; Sanyal, M.; Ding, S.; Jiang, B.; Sen, A.; Morton, J.M.; He, X.S.; Robinson, W.H.; et al. VP4- and VP7-specific antibodies mediate heterotypic immunity to rotavirus in humans. Sci. Transl Med. 2017, 9, eaam5434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parashar, U.D.; Burton, A.; Lanata, C.; Boschi-Pinto, C.; Shibuya, K.; Steele, D.; Birmingham, M.; Glass, R.I. Global mortality associated with rotavirus disease among children in 2004. J. Infect. Dis. 2009, 200 (Suppl. 1), S9–S15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Troeger, C.; Khalil, I.A.; Rao, P.C.; Cao, S.; Blacker, B.F.; Ahmed, T.; Armah, G.; Bines, J.E.; Brewer, T.G.; Colombara, D.V.; et al. Rotavirus Vaccination and the Global Burden of Rotavirus Diarrhea Among Children Younger Than 5 Years. JAMA Pediatrics 2018, 172, 958–965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aliabadi, N.; Antoni, S.; Mwenda, J.M.; Weldegebriel, G.; Biey, J.N.M.; Cheikh, D.; Fahmy, K.; Teleb, N.; Ashmony, H.A.; Ahmed, H.; et al. Global impact of rotavirus vaccine introduction on rotavirus hospitalisations among children under 5 years of age, 2008–2016: Findings from the Global Rotavirus Surveillance Network. Lancet Glob. Health 2019, 7, e893–e903. [Google Scholar] [CrossRef] [Green Version]
- Soares-Weiser, K.; Bergman, H.; Henschke, N.; Pitan, F.; Cunliffe, N. Vaccines for preventing rotavirus diarrhoea: Vaccines in use. Cochrane Database Syst. Rev. 2019, 3, Cd008521. [Google Scholar] [CrossRef]
- Cárcamo-Calvo, R.; Muñoz, C.; Buesa, J.; Rodríguez-Díaz, J.; Gozalbo-Rovira, R. The Rotavirus Vaccine Landscape, an Update. Pathogens 2021, 10, 520. [Google Scholar] [CrossRef]
- Patel, M.M.; Hall, A.J.; Vinje, J.; Parashar, U.D. Noroviruses: A comprehensive review. J. Clin. Virol. Off. Publ. Pan. Am. Soc. Clin. Virol. 2009, 44, 1–8. [Google Scholar] [CrossRef]
- Blazevic, V.; Malm, M.; Honkanen, H.; Knip, M.; Hyöty, H.; Vesikari, T. Development and maturation of norovirus antibodies in childhood. Microbes Infect. 2016, 18, 263–269. [Google Scholar] [CrossRef]
- Bartsch, S.M.; O’Shea, K.J.; Lee, B.Y. The Clinical and Economic Burden of Norovirus Gastroenteritis in the United States. J. Infect. Dis. 2020, 222, 1910–1919. [Google Scholar] [CrossRef]
- Burke, R.M.; Mattison, C.; Marsh, Z.; Shioda, K.; Donald, J.; Salas, S.B.; Naleway, A.L.; Biggs, C.; Schmidt, M.A.; Hall, A.J. Norovirus and Other Viral Causes of Medically Attended Acute Gastroenteritis Across the Age Spectrum: Results from the MAAGE Study in the United States. Clin. Infect. Dis. 2021. [Google Scholar] [CrossRef]
- Safadi, M.A.; Riera-Montes, M.; Bravo, L.; Tangsathapornpong, A.; Lagos, R.; Thisyakorn, U.; Linhares, A.C.; Capeding, R.; Prommalikit, O.; Verstraeten, T.; et al. The burden of norovirus disease in children: A multi-country study in Chile, Brazil, Thailand and the Philippines. Int. J. Infect. Dis. IJID Off. Publ. Int. Soc. Infect. Dis. 2021, 109, 77–84. [Google Scholar] [CrossRef]
- Tarr, G.A.M.; Pang, X.L.; Zhuo, R.; Lee, B.E.; Chui, L.; Ali, S.; Vanderkooi, O.G.; Michaels-Igbokwe, C.; Tarr, P.I.; MacDonald, S.E.; et al. Attribution of Pediatric Acute Gastroenteritis Episodes and Emergency Department Visits to Norovirus Genogroups I and II. J. Infect. Dis. 2021, 223, 452–461. [Google Scholar] [CrossRef] [PubMed]
- Bányai, K.; Estes, M.K.; Martella, V.; Parashar, U.D. Viral gastroenteritis. Lancet 2018, 392, 175–186. [Google Scholar] [CrossRef]
- Gurwith, M.; Wenman, W.; Hinde, D.; Feltham, S.; Greenberg, H. A prospective study of rotavirus infection in infants and young children. J. Infect. Dis. 1981, 144, 218–224. [Google Scholar] [CrossRef] [PubMed]
- Bhavanam, S.; Freedman, S.B.; Lee, B.E.; Zhuo, R.; Qiu, Y.; Chui, L.; Xie, J.; Ali, S.; Vanderkooi, O.G.; Pang, X.L.; et al. Differences in Illness Severity among Circulating Norovirus Genotypes in a Large Pediatric Cohort with Acute Gastroenteritis. Microorganisms 2020, 8, 1873. [Google Scholar] [CrossRef] [PubMed]
- Haddadin, Z.; Batarseh, E.; Hamdan, L.; Stewart, L.S.; Piya, B.; Rahman, H.; Spieker, A.J.; Chappell, J.; Wikswo, M.E.; Dunn, J.R.; et al. Characteristics of GII.4 Norovirus versus other Genotypes in Sporadic Pediatric Infections in Davidson County, Tennessee, USA. Clin. Infect. Dis. 2020, 73, e1525–e1531. [Google Scholar] [CrossRef] [PubMed]
- Duan, L.; Yang, X.; Xie, J.; Zhan, W.; Zhang, C.; Liu, H.; Wei, M.; Tang, Y.; Zhao, H.; Luo, M. Prevalence of GII.4 Sydney Norovirus Strains and Associated Factors of Acute Gastroenteritis in Children: 2019/2020 Season in Guangzhou, China. Food Environ. Virol. 2021, 13, 357–367. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.Y.; Lai, H.H.; Elaine Chen, Y.F.; Chao, H.C.; Tsai, C.N.; Chang, Y.J.; Chen, S.Y. Clinical significance of the fucosyltransferase 2 (FUT2) secretor status in children hospitalized with acute gastroenteritis in Taiwan. J. Med. Assoc. 2021, 120, 212–216. [Google Scholar] [CrossRef]
- Freedman, S.B.; Xie, J.; Lee, B.E.; Ali, S.; Pang, X.L.; Chui, L.; Zhuo, R.; Vanderkooi, O.G.; Tellier, R.; Funk, A.L.; et al. Microbial Etiologies and Clinical Characteristics of Children Seeking Emergency Department Care due to Vomiting in the Absence of Diarrhea. Clin. Infect. Dis. 2021, ciab451. [Google Scholar] [CrossRef]
- Prasad, B.V.; Hardy, M.E.; Dokland, T.; Bella, J.; Rossmann, M.G.; Estes, M.K. X-ray crystallographic structure of the Norwalk virus capsid. Science 1999, 286, 287–290. [Google Scholar] [CrossRef] [Green Version]
- Cao, S.; Lou, Z.; Tan, M.; Chen, Y.; Liu, Y.; Zhang, Z.; Zhang, X.C.; Jiang, X.; Li, X.; Rao, Z. Structural basis for the recognition of blood group trisaccharides by norovirus. J. Virol. 2007, 81, 5949–5957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, H.Q.; Smith, T.J. The Dynamic Capsid Structures of the Noroviruses. Viruses 2019, 11, 235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mallory, M.L.; Lindesmith, L.C.; Graham, R.L.; Baric, R.S. GII.4 Human Norovirus: Surveying the Antigenic Landscape. Viruses 2019, 11, 177. [Google Scholar] [CrossRef] [Green Version]
- Hoa Tran, T.N.; Trainor, E.; Nakagomi, T.; Cunliffe, N.A.; Nakagomi, O. Molecular epidemiology of noroviruses associated with acute sporadic gastroenteritis in children: Global distribution of genogroups, genotypes and GII.4 variants. J. Clin. Virol. Off. Publ. Pan. Am. Soc. Clin. Virol. 2013, 56, 185–193. [Google Scholar] [CrossRef] [Green Version]
- Chhabra, P.; de Graaf, M.; Parra, G.I.; Chan, M.C.; Green, K.; Martella, V.; Wang, Q.; White, P.A.; Katayama, K.; Vennema, H.; et al. Updated classification of norovirus genogroups and genotypes. J. Gen. Virol. 2019, 100, 1393–1406. [Google Scholar] [CrossRef]
- Farahmand, M.; Moghoofei, M.; Dorost, A.; Shoja, Z.; Ghorbani, S.; Kiani, S.J.; Khales, P.; Esteghamati, A.; Sayyahfar, S.; Jafarzadeh, M.; et al. Global prevalence and genotype distribution of norovirus infection in children with gastroenteritis: A meta-analysis on 6 years of research from 2015 to 2020. Rev. Med. Virol. 2021, e2237. [Google Scholar] [CrossRef]
- Cannon, J.L.; Bonifacio, J.; Bucardo, F.; Buesa, J.; Bruggink, L.; Chan, M.C.; Fumian, T.M.; Giri, S.; Gonzalez, M.D.; Hewitt, J.; et al. Global Trends in Norovirus Genotype Distribution among Children with Acute Gastroenteritis. Emerg. Infect. Dis. 2021, 27, 1438–1445. [Google Scholar] [CrossRef] [PubMed]
- Teunis, P.F.M.; Moe, C.L.; Liu, P.; Miller, S.E.; Lindesmith, L.; Baric, R.S.; Le Pendu, J.; Calderon, R.L. Norwalk virus: How infectious is it? J. Med. Virol. 2008, 80, 1468–1476. [Google Scholar] [CrossRef]
- Ramesh, A.K.; Parreño, V.; Schmidt, P.J.; Lei, S.; Zhong, W.; Jiang, X.; Emelko, M.B.; Yuan, L. Evaluation of the 50% Infectious Dose of Human Norovirus Cin-2 in Gnotobiotic Pigs: A Comparison of Classical and Contemporary Methods for Endpoint Estimation. Viruses 2020, 12, 955. [Google Scholar] [CrossRef] [PubMed]
- Teunis, P.F.M.; Le Guyader, F.S.; Liu, P.; Ollivier, J.; Moe, C.L. Noroviruses are highly infectious but there is strong variation in host susceptibility and virus pathogenicity. Epidemics 2020, 32, 100401. [Google Scholar] [CrossRef] [PubMed]
- Guix, S.; Fuentes, C.; Pintó, R.M.; Blanco, A.; Sabrià, A.; Anfruns-Estrada, E.; Garrido, V.R.; Alonso, M.; Bartolomé, R.; Cornejo, T.; et al. Infectivity of Norovirus GI and GII from Bottled Mineral Water during a Waterborne Outbreak, Spain. Emerg. Infect. Dis. 2020, 26, 134–137. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.Y.; Lee, C.C.; Chang, Y.C.; Tsai, C.N.; Chao, H.C.; Tsai, Y.T.; Hsieh, C.H.; Su, S.S.; Chen, S.Y. Viral shedding in gastroenteritis in children caused by variants and novel recombinant norovirus infections. Medicine 2021, 100, e25123. [Google Scholar] [CrossRef] [PubMed]
- Patel, M.M.; Widdowson, M.A.; Glass, R.I.; Akazawa, K.; Vinjé, J.; Parashar, U.D. Systematic literature review of role of noroviruses in sporadic gastroenteritis. Emerg. Infect. Dis. 2008, 14, 1224–1231. [Google Scholar] [CrossRef]
- Trivedi, T.K.; Desai, R.; Hall, A.J.; Patel, M.; Parashar, U.D.; Lopman, B.A. Clinical characteristics of norovirus-associated deaths: A systematic literature review. Am. J. Infect. Control. 2013, 41, 654–657. [Google Scholar] [CrossRef]
- Hall, A.J.; Lopman, B.A.; Payne, D.C.; Patel, M.M.; Gastañaduy, P.A.; Vinjé, J.; Parashar, U.D. Norovirus disease in the United States. Emerg. Infect. Dis. 2013, 19, 1198–1205. [Google Scholar] [CrossRef]
- Tenge, V.R.; Murakami, K.; Salmen, W.; Lin, S.-C.; Crawford, S.E.; Neill, F.H.; Prasad, B.V.V.; Atmar, R.L.; Estes, M.K. Bile Goes Viral. Viruses 2021, 13, 998. [Google Scholar] [CrossRef]
- Lindesmith, L.C.; Donaldson, E.F.; Lobue, A.D.; Cannon, J.L.; Zheng, D.P.; Vinje, J.; Baric, R.S. Mechanisms of GII.4 norovirus persistence in human populations. PLoS Med. 2008, 5, e31. [Google Scholar] [CrossRef]
- de Rougemont, A.; Ruvoen-Clouet, N.; Simon, B.; Estienney, M.; Elie-Caille, C.; Aho, S.; Pothier, P.; Le Pendu, J.; Boireau, W.; Belliot, G. Qualitative and quantitative analysis of the binding of GII.4 norovirus variants onto human blood group antigens. J. Virol. 2011, 85, 4057–4070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anfruns-Estrada, E.; Sabrià, A.; Fuentes, C.; Sabaté, S.; Razquin, E.; Cornejo, T.; Bartolomé, R.; Torner, N.; Izquierdo, C.; Soldevila, N.; et al. Detection of Norovirus in Saliva Samples from Acute Gastroenteritis Cases and Asymptomatic Subjects: Association with Age and Higher Shedding in Stool. Viruses 2020, 12, 1369. [Google Scholar] [CrossRef]
- Lindesmith, L.C.; Brewer-Jensen, P.D.; Mallory, M.L.; Jensen, K.; Yount, B.L.; Costantini, V.; Collins, M.H.; Edwards, C.E.; Sheahan, T.P.; Vinjé, J.; et al. Virus-Host Interactions between Nonsecretors and Human Norovirus. Cell. Mol. Gastroenterol. Hepatol. 2020, 10, 245–267. [Google Scholar] [CrossRef]
- Cao, R.R.; Ma, X.Z.; Li, W.Y.; Wang, B.N.; Yang, Y.; Wang, H.R.; Kuang, Y.; You, J.Z.; Zhao, Z.Y.; Ren, M.; et al. Epidemiology of norovirus gastroenteritis in hospitalized children under five years old in western China, 2015–2019. J. Microbiol. Immunol. Infect. 2021, S1684. [Google Scholar] [CrossRef]
- Manouana, G.P.; Nguema-Moure, P.A.; Mbong Ngwese, M.; Bock, C.; Kremsner, P.G.; Borrmann, S.; Eibach, D.; Mordmüller, B.; Velavan, T.P.; Niendorf, S.; et al. Genetic Diversity of Enteric Viruses in Children under Five Years Old in Gabon. Viruses 2021, 13, 545. [Google Scholar] [CrossRef]
- Bucardo, F.; Reyes, Y.; Becker-Dreps, S.; Bowman, N.; Gruber, J.F.; Vinjé, J.; Espinoza, F.; Paniagua, M.; Balmaseda, A.; Svensson, L.; et al. Pediatric norovirus GII.4 infections in Nicaragua, 1999–2015. Infect. Genet. Evol. 2017, 55, 305–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Utsumi, T.; Lusida, M.I.; Dinana, Z.; Wahyuni, R.M.; Soegijanto, S.; Athiyyah, A.F.; Sudarmo, S.M.; Ranuh, R.G.; Darma, A.; Yamani, L.N.; et al. Molecular epidemiology and genetic diversity of norovirus infection in children hospitalized with acute gastroenteritis in East Java, Indonesia in 2015–2019. Infect. Genet. Evol. 2021, 88, 104703. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Dong, Z.; Liu, Y.; Wang, W.; Hou, M.; Wu, J.; Wang, L.; Zhao, Y. Molecular epidemiology and genetic diversity of norovirus among hospitalized children with acute gastroenteritis in Tianjin, China, 2018–2020. BMC Infect. Dis. 2021, 21, 682. [Google Scholar] [CrossRef]
- Rossouw, E.; Brauer, M.; Meyer, P.; du Plessis, N.M.; Avenant, T.; Mans, J. Virus Etiology, Diversity and Clinical Characteristics in South African Children Hospitalised with Gastroenteritis. Viruses 2021, 13, 215. [Google Scholar] [CrossRef]
- Saito, M.; Tsukagoshi, H.; Ishigaki, H.; Aso, J.; Ishii, H.; Okayama, K.; Ryo, A.; Ishioka, T.; Kuroda, M.; Saruki, N.; et al. Molecular evolution of the capsid (VP1) region in human norovirus genogroup II genotype 3. Heliyon 2020, 6, e03835. [Google Scholar] [CrossRef]
- Ludwig-Begall, L.F.; Mauroy, A.; Thiry, E. Norovirus recombinants: Recurrent in the field, recalcitrant in the lab—A scoping review of recombination and recombinant types of noroviruses. J. Gen. Virol. 2018, 99, 970–988. [Google Scholar] [CrossRef]
- Ruis, C.; Roy, S.; Brown, J.R.; Allen, D.J.; Goldstein, R.A.; Breuer, J. The emerging GII.P16-GII.4 Sydney 2012 norovirus lineage is circulating worldwide, arose by late-2014 and contains polymerase changes that may increase virus transmission. PLoS ONE 2017, 12, e0179572. [Google Scholar] [CrossRef]
- Lindesmith, L.C.; Brewer-Jensen, P.D.; Mallory, M.L.; Debbink, K.; Swann, E.W.; Vinje, J.; Baric, R.S. Antigenic Characterization of a Novel Recombinant GII.P16-GII.4 Sydney Norovirus Strain with Minor Sequence Variation Leading to Antibody Escape. J. Infect. Dis. 2018, 217, 1145–1152. [Google Scholar] [CrossRef]
- Barclay, L.; Cannon, J.L.; Wikswo, M.E.; Phillips, A.R.; Browne, H.; Montmayeur, A.M.; Tatusov, R.L.; Burke, R.M.; Hall, A.J.; Vinje, J. Emerging Novel GII.P16 Noroviruses Associated with Multiple Capsid Genotypes. Viruses 2019, 11, 535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tohma, K.; Lepore, C.J.; Martinez, M.; Degiuseppe, J.I.; Khamrin, P.; Saito, M.; Mayta, H.; Nwaba, A.U.A.; Ford-Siltz, L.A.; Green, K.Y.; et al. Genome-wide analyses of human noroviruses provide insights on evolutionary dynamics and evidence of coexisting viral populations evolving under recombination constraints. PLoS Pathog. 2021, 17, e1009744. [Google Scholar] [CrossRef] [PubMed]
- Alsuwaidi, A.R.; Al Dhaheri, K.; Al Hamad, S.; George, J.; Ibrahim, J.; Ghatasheh, G.; Issa, M.; Al-Hammadi, S.; Narchi, H. Etiology of diarrhea by multiplex polymerase chain reaction among young children in the United Arab Emirates: A case-control study. BMC Infect. Dis. 2021, 21, 7. [Google Scholar] [CrossRef] [PubMed]
- Dey, S.K.; Sharif, N.; Billah, B.; Siddique, T.T.; Islam, T.; Parvez, A.K.; Talukder, A.A.; Phan, T.; Ushijima, H. Molecular epidemiology and genetic diversity of norovirus infection in children with acute gastroenteritis in Bangladesh, 2014–2019. J. Med. Virol. 2021, 93, 3564–3571. [Google Scholar] [CrossRef] [PubMed]
- Vilchez, S.; Reyes, D.; Paniagua, M.; Bucardo, F.; Möllby, R.; Weintraub, A. Prevalence of diarrhoeagenic Escherichia coli in children from León, Nicaragua. J. Med. Microbiol. 2009, 58, 630–637. [Google Scholar] [CrossRef] [Green Version]
- Hesse, S.; Neill, F.H.; Estes, M.K.; Shanker, S.; Prasad, B.V.; Ferreira, J.; Atmar, R.L. Serological Responses to a Norovirus Nonstructural Fusion Protein after Vaccination and Infection. Clin. Vaccine Immunol. CVI 2016, 23, 181–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kulkarni, R.; Lole, K.; Chitambar, S.D. Seroprevalence of antibodies against GII.4 norovirus among children in Pune, India. J. Med. Virol. 2016, 88, 1636–1640. [Google Scholar] [CrossRef]
- Parker, S.P.; Cubitt, W.D.; Jiang, X. Enzyme immunoassay using baculovirus-expressed human calicivirus (Mexico) for the measurement of IgG responses and determining its seroprevalence in London, UK. J. Med. Virol. 1995, 46, 194–200. [Google Scholar] [CrossRef]
- Taylor, M.B.; Parker, S.; Grabow, W.O.; Cubitt, W.D. An epidemiological investigation of Norwalk virus infection in South Africa. Epidemiol. Infect. 1996, 116, 203–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parker, S.P.; Cubitt, W.D.; Jiang, X.J.; Estes, M.K. Seroprevalence studies using a recombinant Norwalk virus protein enzyme immunoassay. J. Med. Virol. 1994, 42, 146–150. [Google Scholar] [CrossRef] [PubMed]
- Arnold, B.F.; Martin, D.L.; Juma, J.; Mkocha, H.; Ochieng, J.B.; Cooley, G.M.; Omore, R.; Goodhew, E.B.; Morris, J.F.; Costantini, V.; et al. Enteropathogen antibody dynamics and force of infection among children in low-resource settings. eLife 2019, 8, e45594. [Google Scholar] [CrossRef] [PubMed]
- Thorne, L.; Nalwoga, A.; Mentzer, A.J.; de Rougemont, A.; Hosmillo, M.; Webb, E.; Nampiija, M.; Muhwezi, A.; Carstensen, T.; Gurdasani, D.; et al. The First Norovirus Longitudinal Seroepidemiological Study from Sub-Saharan Africa Reveals High Seroprevalence of Diverse Genotypes Associated with Host Susceptibility Factors. J. Infect. Dis. 2018, 218, 716–725. [Google Scholar] [CrossRef] [Green Version]
- Menon, V.K.; George, S.; Shanti, A.A.; Saravanabavan, A.; Samuel, P.; Ramani, S.; Estes, M.K.; Kang, G. Exposure to human and bovine noroviruses in a birth cohort in southern India from 2002 to 2006. J. Clin. Microbiol. 2013, 51, 2391–2395. [Google Scholar] [CrossRef] [Green Version]
- van Beek, J.; de Graaf, M.; Xia, M.; Jiang, X.; Vinje, J.; Beersma, M.; de Bruin, E.; van de Vijver, D.; Holwerda, M.; van Houten, M.; et al. Comparison of norovirus genogroup I, II and IV seroprevalence among children in the Netherlands, 1963, 1983 and 2006. J. Gen. Virol. 2016, 97, 2255–2264. [Google Scholar] [CrossRef]
- Grytdal, S.P.; DeBess, E.; Lee, L.E.; Blythe, D.; Ryan, P.; Biggs, C.; Cameron, M.; Schmidt, M.; Parashar, U.D.; Hall, A.J. Incidence of Norovirus and Other Viral Pathogens That Cause Acute Gastroenteritis (AGE) among Kaiser Permanente Member Populations in the United States, 2012–2013. PLoS ONE 2016, 11, e0148395. [Google Scholar] [CrossRef] [Green Version]
- Poulain, C.; Galeno, H.; Loayza, S.; Vergara, N.; Valdivieso, F.; Coria, P.; Sotomayor, V.; Simian, M.E.; Acevedo, J.; Farfán, M.J. Molecular detection of gastrointestinal pathogens among children under 5 years old with diarrhea in a hospital center for rotavirus sentinel surveillance in Chile. Rev. Chil. Infectol. 2021, 38, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Lindesmith, L.; Moe, C.; Lependu, J.; Frelinger, J.A.; Treanor, J.; Baric, R.S. Cellular and humoral immunity following Snow Mountain virus challenge. J. Virol. 2005, 79, 2900–2909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindesmith, L.C.; Beltramello, M.; Donaldson, E.F.; Corti, D.; Swanstrom, J.; Debbink, K.; Lanzavecchia, A.; Baric, R.S. Immunogenetic mechanisms driving norovirus GII.4 antigenic variation. PLoS Pathog. 2012, 8, e1002705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saito, M.; Goel-Apaza, S.; Espetia, S.; Velasquez, D.; Cabrera, L.; Loli, S.; Crabtree, J.E.; Black, R.E.; Kosek, M.; Checkley, W.; et al. Multiple norovirus infections in a birth cohort in a Peruvian Periurban community. Clin. Infect. Dis. 2014, 58, 483–491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cannon, J.L.; Lopman, B.A.; Payne, D.C.; Vinjé, J. Birth Cohort Studies Assessing Norovirus Infection and Immunity in Young Children: A Review. Clin. Infect. Dis. 2019, 69, 357–365. [Google Scholar] [CrossRef] [Green Version]
- Parra, G.I.; Green, K.Y. Sequential gastroenteritis episodes caused by 2 norovirus genotypes. Emerg. Infect. Dis. 2014, 20, 1016–1018. [Google Scholar] [CrossRef] [PubMed]
- Chhabra, P.; Rouhani, S.; Browne, H.; Peñataro Yori, P.; Siguas Salas, M.; Paredes Olortegui, M.; Moulton, L.H.; Kosek, M.N.; Vinjé, J. Homotypic and Heterotypic Protection and Risk of Reinfection Following Natural Norovirus Infection in a Highly Endemic Setting. Clin. Infect. Dis. 2021, 72, 222–229. [Google Scholar] [CrossRef] [Green Version]
- Blazevic, V.; Malm, M.; Salminen, M.; Oikarinen, S.; Hyöty, H.; Veijola, R.; Vesikari, T. Multiple consecutive norovirus infections in the first 2 years of life. Eur. J. Pediatr. 2015, 174, 1679–1683. [Google Scholar] [CrossRef] [Green Version]
- Mateo, R.; Lindesmith, L.C.; Garg, S.J.; Gottlieb, K.; Lin, K.; Said, S.; Leon, J.S.; Sims, A.C.; Weber, D.J.; Baric, R.S.; et al. Production and Clinical Evaluation of Norwalk GI.1 Virus Lot 001-09NV in Norovirus Vaccine Development. J. Infect. Dis. 2020, 221, 919–926. [Google Scholar] [CrossRef] [Green Version]
- Lindesmith, L.C.; McDaniel, J.R.; Changela, A.; Verardi, R.; Kerr, S.A.; Costantini, V.; Brewer-Jensen, P.D.; Mallory, M.L.; Voss, W.N.; Boutz, D.R.; et al. Sera Antibody Repertoire Analyses Reveal Mechanisms of Broad and Pandemic Strain Neutralizing Responses after Human Norovirus Vaccination. Immunity 2019, 50, 1530–1541.e1538. [Google Scholar] [CrossRef] [Green Version]
- Alvarado, G.; Salmen, W.; Ettayebi, K.; Hu, L.; Sankaran, B.; Estes, M.K.; Venkataram Prasad, B.V.; Crowe, J.E., Jr. Broadly cross-reactive human antibodies that inhibit genogroup I and II noroviruses. Nat. Commun. 2021, 12, 4320. [Google Scholar] [CrossRef]
- Malm, M.; Uusi-Kerttula, H.; Vesikari, T.; Blazevic, V. High serum levels of norovirus genotype-specific blocking antibodies correlate with protection from infection in children. J. Infect. Dis. 2014, 210, 1755–1762. [Google Scholar] [CrossRef] [Green Version]
- Alexander, E.; Hommeida, S.; Stephens, M.C.; Manini, M.L.; Absah, I. The Role of Oral Administration of Immunoglobulin in Managing Diarrheal Illness in Immunocompromised Children. Paediatr. Drugs 2020, 22, 331–334. [Google Scholar] [CrossRef]
- Nussbaum, E.Z.; Azar, M.M.; Cohen, E.; McManus, D.; Topal, J.E.; Malinis, M. Orally Administered Human Immunoglobulin Therapy for Norovirus Enteritis in Solid Organ Transplant Recipients: A Case Series at a Single Academic Transplant Center. Clin. Infect. Dis. 2020, 71, e206–e209. [Google Scholar] [CrossRef] [PubMed]
- Shillitoe, B.M.J.; Ponsford, M.; Slatter, M.A.; Evans, J.; Struik, S.; Cosgrove, M.; Doull, I.; Jolles, S.; Gennery, A.R. Haematopoietic Stem Cell Transplant for Norovirus-Induced Intestinal Failure in X-linked Agammaglobulinemia. J. Clin. Immunol. 2021, 41, 1574–1581. [Google Scholar] [CrossRef]
- Jain, P.; Mishra, A.; Gupta, D.; Kulkarni, S. Chronic enteropathy-related malabsorption syndrome in an adult with common variable immunodeficiency and symptomatic norovirus infection of the gut. BMJ Case Rep. 2021, 14, e241752. [Google Scholar] [CrossRef] [PubMed]
- Lindesmith, L.C.; Brewer-Jensen, P.D.; Mallory, M.L.; Yount, B.; Collins, M.H.; Debbink, K.; Graham, R.L.; Baric, R.S. Human Norovirus Epitope D Plasticity Allows Escape from Antibody Immunity without Loss of Capacity for Binding Cellular Ligands. J. Virol. 2019, 93, e01813-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atmar, R.L.; Bernstein, D.I.; Lyon, G.M.; Treanor, J.J.; Al-Ibrahim, M.S.; Graham, D.Y.; Vinje, J.; Jiang, X.; Gregoricus, N.; Frenck, R.W.; et al. Serological Correlates of Protection against a GII.4 Norovirus. Clin. Vaccine Immunol. CVI 2015, 22, 923–929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramani, S.; Neill, F.H.; Opekun, A.R.; Gilger, M.A.; Graham, D.Y.; Estes, M.K.; Atmar, R.L. Mucosal and Cellular Immune Responses to Norwalk Virus. J. Infect. Dis. 2015, 212, 397–405. [Google Scholar] [CrossRef] [Green Version]
- Lindesmith, L.C.; Ferris, M.T.; Mullan, C.W.; Ferreira, J.; Debbink, K.; Swanstrom, J.; Richardson, C.; Goodwin, R.R.; Baehner, F.; Mendelman, P.M.; et al. Broad blockade antibody responses in human volunteers after immunization with a multivalent norovirus VLP candidate vaccine: Immunological analyses from a phase I clinical trial. PLoS Med. 2015, 12, e1001807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reeck, A.; Kavanagh, O.; Estes, M.K.; Opekun, A.R.; Gilger, M.A.; Graham, D.Y.; Atmar, R.L. Serological correlate of protection against norovirus-induced gastroenteritis. J. Infect. Dis. 2010, 202, 1212–1218. [Google Scholar] [CrossRef] [Green Version]
- Kim, L.; Liebowitz, D.; Lin, K.; Kasparek, K.; Pasetti, M.F.; Garg, S.J.; Gottlieb, K.; Trager, G.; Tucker, S.N. Safety and immunogenicity of an oral tablet norovirus vaccine, a phase I randomized, placebo-controlled trial. JCI Insight 2018, 3, e121077. [Google Scholar] [CrossRef]
- Treanor, J.; Dolin, R.; Madore, H.P. Production of a monoclonal antibody against the Snow Mountain agent of gastroenteritis by in vitro immunization of murine spleen cells. Proc. Natl. Acad. Sci. USA 1988, 85, 3613–3617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harrington, P.R.; Lindesmith, L.; Yount, B.; Moe, C.L.; Baric, R.S. Binding of Norwalk virus-like particles to ABH histo-blood group antigens is blocked by antisera from infected human volunteers or experimentally vaccinated mice. J. Virol. 2002, 76, 12335–12343. [Google Scholar] [CrossRef] [Green Version]
- Atmar, R.L.; Ettayebi, K.; Ayyar, B.V.; Neill, F.H.; Braun, R.P.; Ramani, S.; Estes, M.K. Comparison of Microneutralization and Histo-Blood Group Antigen-Blocking Assays for Functional Norovirus Antibody Detection. J. Infect. Dis. 2020, 221, 739–743. [Google Scholar] [CrossRef]
- Alvarado, G.; Ettayebi, K.; Atmar, R.L.; Bombardi, R.G.; Kose, N.; Estes, M.K.; Crowe, J.E., Jr. Human Monoclonal Antibodies That Neutralize Pandemic GII.4 Noroviruses. Gastroenterology 2018, 155, 1898–1907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tohma, K.; Lepore, C.J.; Gao, Y.; Ford-Siltz, L.A.; Parra, G.I. Population Genomics of GII.4 Noroviruses Reveal Complex Diversification and New Antigenic Sites Involved in the Emergence of Pandemic Strains. mBio 2019, 10, e02202-19. [Google Scholar] [CrossRef] [Green Version]
- Kendra, J.A.; Tohma, K.; Ford-Siltz, L.A.; Lepore, C.J.; Parra, G.I. Antigenic cartography reveals complexities of genetic determinants that lead to antigenic differences among pandemic GII.4 noroviruses. Proc. Natl. Acad. Sci. USA 2021, 118, e2015874118. [Google Scholar] [CrossRef]
- Atmar, R.L.; Bernstein, D.I.; Harro, C.D.; Al-Ibrahim, M.S.; Chen, W.H.; Ferreira, J.; Estes, M.K.; Graham, D.Y.; Opekun, A.R.; Richardson, C.; et al. Norovirus vaccine against experimental human Norwalk Virus illness. N. Engl. J. Med. 2011, 365, 2178–2187. [Google Scholar] [CrossRef] [Green Version]
- Bernstein, D.I.; Atmar, R.L.; Lyon, G.M.; Treanor, J.J.; Chen, W.H.; Jiang, X.; Vinjé, J.; Gregoricus, N.; Frenck, R.W., Jr.; Moe, C.L.; et al. Norovirus vaccine against experimental human GII.4 virus illness: A challenge study in healthy adults. J. Infect. Dis. 2015, 211, 870–878. [Google Scholar] [CrossRef] [PubMed]
- Knoll, B.M.; Lindesmith, L.C.; Yount, B.L.; Baric, R.S.; Marty, F.M. Resolution of diarrhea in an immunocompromised patient with chronic norovirus gastroenteritis correlates with constitution of specific antibody blockade titer. Infection 2016, 44, 551–554. [Google Scholar] [CrossRef] [PubMed]
- Lindesmith, L.C.; Beltramello, M.; Swanstrom, J.; Jones, T.A.; Corti, D.; Lanzavecchia, A.; Baric, R.S. Serum Immunoglobulin a Cross-Strain Blockade of Human Noroviruses. Open Forum Infect. Dis. 2015, 2, ofv084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sapparapu, G.; Czakó, R.; Alvarado, G.; Shanker, S.; Prasad, B.V.; Atmar, R.L.; Estes, M.K.; Crowe, J.E., Jr. Frequent Use of the IgA Isotype in Human B Cells Encoding Potent Norovirus-Specific Monoclonal Antibodies That Block HBGA Binding. PLoS Pathog. 2016, 12, e1005719. [Google Scholar] [CrossRef]
- Tamminen, K.; Malm, M.; Vesikari, T.; Blazevic, V. Norovirus-specific mucosal antibodies correlate to systemic antibodies and block norovirus virus-like particles binding to histo-blood group antigens. Clin. Immunol. 2018, 197, 110–117. [Google Scholar] [CrossRef] [Green Version]
- Costantini, V.P.; Cooper, E.M.; Hardaker, H.L.; Lee, L.E.; DeBess, E.E.; Cieslak, P.R.; Hall, A.J.; Vinjé, J. Humoral and Mucosal Immune Responses to Human Norovirus in the Elderly. J. Infect. Dis. 2020, 221, 1864–1874. [Google Scholar] [CrossRef]
- Blazevic, V.; Malm, M.; Vesikari, T. Induction of homologous and cross-reactive GII.4-specific blocking antibodies in children after GII.4 New Orleans norovirus infection. J. Med. Virol. 2015, 87, 1656–1661. [Google Scholar] [CrossRef]
- Debbink, K.; Lindesmith, L.C.; Donaldson, E.F.; Costantini, V.; Beltramello, M.; Corti, D.; Swanstrom, J.; Lanzavecchia, A.; Vinje, J.; Baric, R.S. Emergence of new pandemic GII.4 Sydney norovirus strain correlates with escape from herd immunity. J. Infect. Dis. 2013, 208, 1877–1887. [Google Scholar] [CrossRef] [Green Version]
- Lindesmith, L.C.; Costantini, V.; Swanstrom, J.; Debbink, K.; Donaldson, E.F.; Vinje, J.; Baric, R.S. Emergence of a Norovirus GII.4 Strain Correlates with Changes in Evolving Blockade Epitopes. J. Virol. 2013, 87, 2803–2813. [Google Scholar] [CrossRef] [Green Version]
- Lindesmith, L.C.; Donaldson, E.; Leon, J.; Moe, C.L.; Frelinger, J.A.; Johnston, R.E.; Weber, D.J.; Baric, R.S. Heterotypic humoral and cellular immune responses following Norwalk virus infection. J. Virol. 2010, 84, 1800–1815. [Google Scholar] [CrossRef] [Green Version]
- Sherwood, J.; Mendelman, P.M.; Lloyd, E.; Liu, M.; Boslego, J.; Borkowski, A.; Jackson, A.; Faix, D. Efficacy of an intramuscular bivalent norovirus GI.1/GII.4 virus-like particle vaccine candidate in healthy US adults. Vaccine 2020, 38, 6442–6449. [Google Scholar] [CrossRef] [PubMed]
- Atmar, R.L.; Baehner, F.; Cramer, J.P.; Song, E.; Borkowski, A.; Mendelman, P.M. Rapid Responses to 2 Virus-Like Particle Norovirus Vaccine Candidate Formulations in Healthy Adults: A Randomized Controlled Trial. J. Infect. Dis. 2016, 214, 845–853. [Google Scholar] [CrossRef] [Green Version]
- Atmar, R.L.; Baehner, F.; Cramer, J.P.; Lloyd, E.; Sherwood, J.; Borkowski, A.; Mendelman, P.M. Persistence of Antibodies to 2 Virus-Like Particle Norovirus Vaccine Candidate Formulations in Healthy Adults: 1-Year Follow-up with Memory Probe Vaccination. J. Infect. Dis. 2019, 220, 603–614. [Google Scholar] [CrossRef] [PubMed]
- Ford-Siltz, L.A.; Wales, S.; Tohma, K.; Gao, Y.; Parra, G.I. Genotype-Specific Neutralization of Norovirus is Mediated by Antibodies Against the Protruding Domain of the Major Capsid Protein. J. Infect. Dis. 2020, jiaa116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Debbink, K.; Lindesmith, L.C.; Baric, R.S. The state of norovirus vaccines. Clin. Infect. Dis. 2014, 58, 1746–1752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- LoBue, A.D.; Lindesmith, L.; Yount, B.; Harrington, P.R.; Thompson, J.M.; Johnston, R.E.; Moe, C.L.; Baric, R.S. Multivalent norovirus vaccines induce strong mucosal and systemic blocking antibodies against multiple strains. Vaccine 2006, 24, 5220–5234. [Google Scholar] [CrossRef]
- Lindesmith, L.C.; Mallory, M.L.; Jones, T.A.; Richardson, C.; Goodwin, R.R.; Baehner, F.; Mendelman, P.M.; Bargatze, R.F.; Baric, R.S. Impact of Pre-exposure History and Host Genetics on Antibody Avidity Following Norovirus Vaccination. J. Infect. Dis. 2017, 215, 984–991. [Google Scholar] [CrossRef]
- Ramani, S.; Neill, F.H.; Ferreira, J.; Treanor, J.J.; Frey, S.E.; Topham, D.J.; Goodwin, R.R.; Borkowski, A.; Baehner, F.; Mendelman, P.M.; et al. B-Cell Responses to Intramuscular Administration of a Bivalent Virus-Like Particle Human Norovirus Vaccine. Clin. Vaccine Immunol. CVI 2017, 24, e00571-16. [Google Scholar] [CrossRef] [Green Version]
- Bartsch, S.M.; O’Shea, K.J.; Wedlock, P.T.; Ferguson, M.C.; Siegmund, S.S.; Lee, B.Y. Potential Clinical and Economic Value of Norovirus Vaccination in the Community Setting. Am. J. Prev. Med. 2021, 60, 360–368. [Google Scholar] [CrossRef]
- Vielot, N.A.; González, F.; Reyes, Y.; Zepeda, O.; Blette, B.; Paniagua, M.; Toval-Ruíz, C.; Diez-Valcarce, M.; Hudgens, M.G.; Gutiérrez, L.; et al. Risk Factors and Clinical Profile of Sapovirus-associated Acute Gastroenteritis in Early Childhood: A Nicaraguan Birth Cohort Study. J. Pediatr. Infect. Dis. 2021, 40, 220–226. [Google Scholar] [CrossRef]
- Farkas, T.; Deng, X.; Ruiz-Palacios, G.; Morrow, A.; Jiang, X. Development of an enzyme immunoassay for detection of sapovirus-specific antibodies and its application in a study of seroprevalence in children. J. Clin. Microbiol. 2006, 44, 3674–3679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donowitz, J.R.; Drew, J.; Taniuchi, M.; Platts-Mills, J.A.; Alam, M.; Ferdous, T.; Shama, T.; Islam, M.O.; Kabir, M.; Nayak, U.; et al. Diarrheal Pathogens Associated with Growth and Neurodevelopment. Clin. Infect. Dis. 2021, 73, e683–e691. [Google Scholar] [CrossRef] [PubMed]
- Halasa, N.; Piya, B.; Stewart, L.S.; Rahman, H.; Payne, D.C.; Woron, A.; Thomas, L.; Constantine-Renna, L.; Garman, K.; McHenry, R.; et al. The Changing Landscape of Pediatric Viral Enteropathogens in the Post-Rotavirus Vaccine Era. Clin. Infect. Dis. 2021, 72, 576–585. [Google Scholar] [CrossRef] [PubMed]
- Inns, T.; Wilson, D.; Manley, P.; Harris, J.P.; O’Brien, S.J.; Vivancos, R. What proportion of care home outbreaks are caused by norovirus? An analysis of viral causes of gastroenteritis outbreaks in care homes, North East England, 2016–2018. BMC Infect. Dis. 2019, 20, 2. [Google Scholar] [CrossRef] [Green Version]
- Daniel-Wayman, S.; Fahle, G.; Palmore, T.; Green, K.Y.; Prevots, D.R. Norovirus, astrovirus, and sapovirus among immunocompromised patients at a tertiary care research hospital. Diagn. Microbiol. Infect. Dis. 2018, 92, 143–146. [Google Scholar] [CrossRef]
- Pietsch, C.; Liebert, U.G. Intrahost viral evolution during chronic sapovirus infections. J. Clin. Virol. Off. Publ. Pan. Am. Soc. Clin. Virol. 2019, 113, 1–7. [Google Scholar] [CrossRef]
- Diez Valcarce, M.; Kambhampati, A.K.; Calderwood, L.E.; Hall, A.J.; Mirza, S.A.; Vinjé, J. Global distribution of sporadic sapovirus infections: A systematic review and meta-analysis. PLoS ONE 2021, 16, e0255436. [Google Scholar] [CrossRef]
- Chang, K.O.; Sosnovtsev, S.V.; Belliot, G.; Wang, Q.; Saif, L.J.; Green, K.Y. Reverse genetics system for porcine enteric calicivirus, a prototype sapovirus in the Caliciviridae. J. Virol. 2005, 79, 1409–1416. [Google Scholar] [CrossRef] [Green Version]
- Human Calicivirus Typing Tool. Available online: https://norovirus.ng.philab.cdc.gov/ (accessed on 14 June 2021).
- Becker-Dreps, S.; González, F.; Bucardo, F. Sapovirus: An emerging cause of childhood diarrhea. Curr. Opin. Infect. Dis. 2020, 33, 388–397. [Google Scholar] [CrossRef]
- Bucardo, F.; Carlsson, B.; Nordgren, J.; Larson, G.; Blandon, P.; Vilchez, S.; Svensson, L. Susceptibility of children to sapovirus infections, Nicaragua, 2005-2006. Emerg. Infect. Dis. 2012, 18, 1875–1878. [Google Scholar] [CrossRef]
- Takagi, H.; Oka, T.; Shimoike, T.; Saito, H.; Kobayashi, T.; Takahashi, T.; Tatsumi, C.; Kataoka, M.; Wang, Q.; Saif, L.J.; et al. Human sapovirus propagation in human cell lines supplemented with bile acids. Proc. Natl. Acad. Sci. USA 2020, 117, 32078–32085. [Google Scholar] [CrossRef]
- Iritani, N.; Yamamoto, S.P.; Abe, N.; Kubo, H.; Oka, T.; Kaida, A. Epidemics of GI.2 sapovirus in gastroenteritis outbreaks during 2012-2013 in Osaka City, Japan. J. Med. Virol. 2016, 88, 1187–1193. [Google Scholar] [CrossRef] [PubMed]
- Lyman, W.H.; Walsh, J.F.; Kotch, J.B.; Weber, D.J.; Gunn, E.; Vinjé, J. Prospective study of etiologic agents of acute gastroenteritis outbreaks in child care centers. J. Pediatr. 2009, 154, 253–257. [Google Scholar] [CrossRef] [PubMed]
- Hassan-Ríos, E.; Torres, P.; Muñoz, E.; Matos, C.; Hall, A.; Gregoricus, N.; Vinjé, J. Sapovirus Gastroenteritis in Preschool Center, Puerto Rico, 2011. Emerg. Infect. Dis. 2013, 19, 174. [Google Scholar] [CrossRef]
- Wang, J.; Li, Y.; Kong, X.; Li, H.; Zhang, Q.; Jin, M.; Wang, Y.; Duan, Z.-J. Two gastroenteritis outbreaks caused by sapovirus in Shenzhen, China. J. Med. Virol. 2018, 90, 1695–1702. [Google Scholar] [CrossRef] [PubMed]
- Johansson, P.J.; Bergentoft, K.; Larsson, P.A.; Magnusson, G.; Widell, A.; Thorhagen, M.; Hedlund, K.O. A nosocomial sapovirus-associated outbreak of gastroenteritis in adults. Scand. J. Infect. Dis. 2005, 37, 200–204. [Google Scholar] [CrossRef] [PubMed]
- Pang, X.L.; Lee, B.E.; Tyrrell, G.J.; Preiksaitis, J.K. Epidemiology and genotype analysis of sapovirus associated with gastroenteritis outbreaks in Alberta, Canada: 2004–2007. J. Infect. Dis. 2009, 199, 547–551. [Google Scholar] [CrossRef] [Green Version]
- Lee, L.E.; Cebelinski, E.A.; Fuller, C.; Keene, W.E.; Smith, K.; Vinjé, J.; Besser, J.M. Sapovirus outbreaks in long-term care facilities, Oregon and Minnesota, USA, 2002–2009. Emerg. Infect. Dis. 2012, 18, 873–876. [Google Scholar] [CrossRef]
- Nidaira, M.; Taira, K.; Kato, T.; Arakaki, E.; Kyan, H.; Takara, T.; Okano, S.; Kuba, Y.; Kudaka, J.; Noda, M. Phylogenetic Analysis of Sapovirus Detected from an Outbreak of Acute Gastroenteritis on Ishigaki Island (Okinawa Prefecture, Japan) in 2012. Jpn. J. Infect. Dis. 2014, 67, 141–143. [Google Scholar] [CrossRef]
- Rogawski McQuade, E.T.; Liu, J.; Kang, G.; Kosek, M.N.; Lima, A.A.M.; Bessong, P.O.; Samie, A.; Haque, R.; Mduma, E.R.; Shrestha, S.; et al. Protection from Natural Immunity Against Enteric Infections and Etiology-Specific Diarrhea in a Longitudinal Birth Cohort. J. Infect. Dis. 2020, 222, 1858–1868. [Google Scholar] [CrossRef]
- Chiba, S.; Sakuma, Y.; Kogasaka, R.; Akihara, M.; Horino, K.; Nakao, T.; Fukui, S. An outbreak of gastroenteritis associated with calicivirus in an infant home. J. Med. Virol. 1979, 4, 249–254. [Google Scholar] [CrossRef] [PubMed]
- Cubitt, W.D.; McSwiggan, D.A. Seroepidemiological survey of the prevalence of antibodies to a strain of human calicivirus. J. Med. Virol. 1987, 21, 361–368. [Google Scholar] [CrossRef] [PubMed]
- Nakata, S.; Chiba, S.; Terashima, H.; Yokoyama, T.; Nakao, T. Humoral immunity in infants with gastroenteritis caused by human calicivirus. J. Infect. Dis. 1985, 152, 274–279. [Google Scholar] [CrossRef] [PubMed]
- Peñaflor-Téllez, Y.; Trujillo-Uscanga, A.; Escobar-Almazán, J.A.; Gutiérrez-Escolano, A.L. Immune Response Modulation by Caliciviruses. Front. Immunol. 2019, 10, 2334. [Google Scholar] [CrossRef]
- Roos-Weil, D.; Ambert-Balay, K.; Lanternier, F.; Mamzer-Bruneel, M.F.; Nochy, D.; Pothier, P.; Avettand-Fenoel, V.; Anglicheau, D.; Snanoudj, R.; Bererhi, L.; et al. Impact of norovirus/sapovirus-related diarrhea in renal transplant recipients hospitalized for diarrhea. Transplantation 2011, 92, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Cubitt, W.D.; Berke, T.; Zhong, W.; Dai, X.; Nakata, S.; Pickering, L.K.; Matson, D.O. Sapporo-like human caliciviruses are genetically and antigenically diverse. Arch. Virol. 1997, 142, 1813–1827. [Google Scholar] [CrossRef] [PubMed]
- Hansman, G.S.; Oka, T.; Sakon, N.; Takeda, N. Antigenic diversity of human sapoviruses. Emerg. Infect. Dis. 2007, 13, 1519–1525. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, G.J.; Mayta, H.; Pajuelo, M.J.; Neira, K.; Xiaofang, L.; Cabrera, L.; Ballard, S.B.; Crabtree, J.E.; Kelleher, D.; Cama, V.; et al. Epidemiology of Sapovirus Infections in a Birth Cohort in Peru. Clin. Infect. Dis. 2018, 66, 1858–1863. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Jahuira, H.; Gilman, R.H.; Alva, A.; Cabrera, L.; Okamoto, M.; Xu, H.; Windle, H.J.; Kelleher, D.; Varela, M.; et al. Etiological Role and Repeated Infections of Sapovirus among Children Aged Less than 2 Years in a Cohort Study in a Peri-urban Community of Peru. J. Clin. Microbiol. 2016, 54, 1598–1604. [Google Scholar] [CrossRef] [Green Version]
- Miyazaki, N.; Taylor, D.W.; Hansman, G.S.; Murata, K. Antigenic and Cryo-Electron Microscopy Structure Analysis of a Chimeric Sapovirus Capsid. J. Virol. 2015, 90, 2664–2675. [Google Scholar] [CrossRef] [Green Version]
- Chen, R.; Neill, J.D.; Noel, J.S.; Hutson, A.M.; Glass, R.I.; Estes, M.K.; Prasad, B.V. Inter- and intragenus structural variations in caliciviruses and their functional implications. J. Virol. 2004, 78, 6469–6479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amin, M.R.; Siddiqui, M.S.; Ahmed, D.; Ahmed, F.; Hossain, A. B- and T-cell epitope mapping of human sapovirus capsid protein: An immunomics approach. Int. J. Bioinform. Res. Appl. 2011, 7, 287–298. [Google Scholar] [CrossRef] [PubMed]
- Diez-Valcarce, M.; Castro, C.J.; Marine, R.L.; Halasa, N.; Mayta, H.; Saito, M.; Tsaknaridis, L.; Pan, C.Y.; Bucardo, F.; Becker-Dreps, S.; et al. Genetic diversity of human sapovirus across the Americas. J. Clin. Virol. Off. Publ. Pan. Am. Soc. Clin. Virol. 2018, 104, 65–72. [Google Scholar] [CrossRef]
- Varela, M.F.; Rivadulla, E.; Lema, A.; Romalde, J.L. Human Sapovirus among Outpatients with Acute Gastroenteritis in Spain: A One-Year Study. Viruses 2019, 11, 144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mann, P.; Pietsch, C.; Liebert, U.G. Genetic Diversity of Sapoviruses among Inpatients in Germany, 2008–2018. Viruses 2019, 11, 726. [Google Scholar] [CrossRef] [Green Version]
- Baker, J.M.; Tate, J.E.; Leon, J.; Haber, M.J.; Pitzer, V.E.; Lopman, B.A. Postvaccination Serum Antirotavirus Immunoglobulin A as a Correlate of Protection Against Rotavirus Gastroenteritis Across Settings. J. Infect. Dis. 2020, 222, 309–318. [Google Scholar] [CrossRef]
- Bernstein, D.I.; Ziegler, J.M.; Ward, R.L. Rotavirus fecal IgA antibody response in adults challenged with human rotavirus. J. Med. Virol. 1986, 20, 297–304. [Google Scholar] [CrossRef]
- Coulson, B.S.; Grimwood, K.; Masendycz, P.J.; Lund, J.S.; Mermelstein, N.; Bishop, R.F.; Barnes, G.L. Comparison of rotavirus immunoglobulin A coproconversion with other indices of rotavirus infection in a longitudinal study in childhood. J. Clin. Microbiol. 1990, 28, 1367–1374. [Google Scholar] [CrossRef] [Green Version]
- Lappalainen, S.; Blazevic, V.; Malm, M.; Vesikari, T. Rotavirus vaccination and infection induce VP6-specific IgA responses. J. Med. Virol. 2017, 89, 239–245. [Google Scholar] [CrossRef]
- Czakó, R.; Atmar, R.L.; Opekun, A.R.; Gilger, M.A.; Graham, D.Y.; Estes, M.K. Serum hemagglutination inhibition activity correlates with protection from gastroenteritis in persons infected with Norwalk virus. Clin. Vaccine Immunol. CVI 2012, 19, 284–287. [Google Scholar] [CrossRef] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zweigart, M.R.; Becker-Dreps, S.; Bucardo, F.; González, F.; Baric, R.S.; Lindesmith, L.C. Serological Humoral Immunity Following Natural Infection of Children with High Burden Gastrointestinal Viruses. Viruses 2021, 13, 2033. https://doi.org/10.3390/v13102033
Zweigart MR, Becker-Dreps S, Bucardo F, González F, Baric RS, Lindesmith LC. Serological Humoral Immunity Following Natural Infection of Children with High Burden Gastrointestinal Viruses. Viruses. 2021; 13(10):2033. https://doi.org/10.3390/v13102033
Chicago/Turabian StyleZweigart, Mark R., Sylvia Becker-Dreps, Filemón Bucardo, Fredman González, Ralph S. Baric, and Lisa C. Lindesmith. 2021. "Serological Humoral Immunity Following Natural Infection of Children with High Burden Gastrointestinal Viruses" Viruses 13, no. 10: 2033. https://doi.org/10.3390/v13102033