Toll-Like Receptor 21 of Chicken and Duck Recognize a Broad Array of Immunostimulatory CpG-oligodeoxynucleotide Sequences

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Approval of Animal Work
2.2. Reagents and Antibodies
2.3. Molecular Cloning of Chicken and Duck TLR21s cDNA
2.4. Bioinformatics Analysis
2.5. Expression Vectors for Chicken, Duck, and Grouper TLR21s
2.6. TLR21 Activation Assays
2.7. Tissue Isolation from Chicken and Duck for First-Strand cDNA Preparation
2.8. Preparation and Culture of Chicken and Duck Splenocytes
2.9. RT-qPCR Analysis of Gene Expression
2.10. SDS-PAGE and Immunoblot Analysis
2.11. Statistical Analysis
3. Results
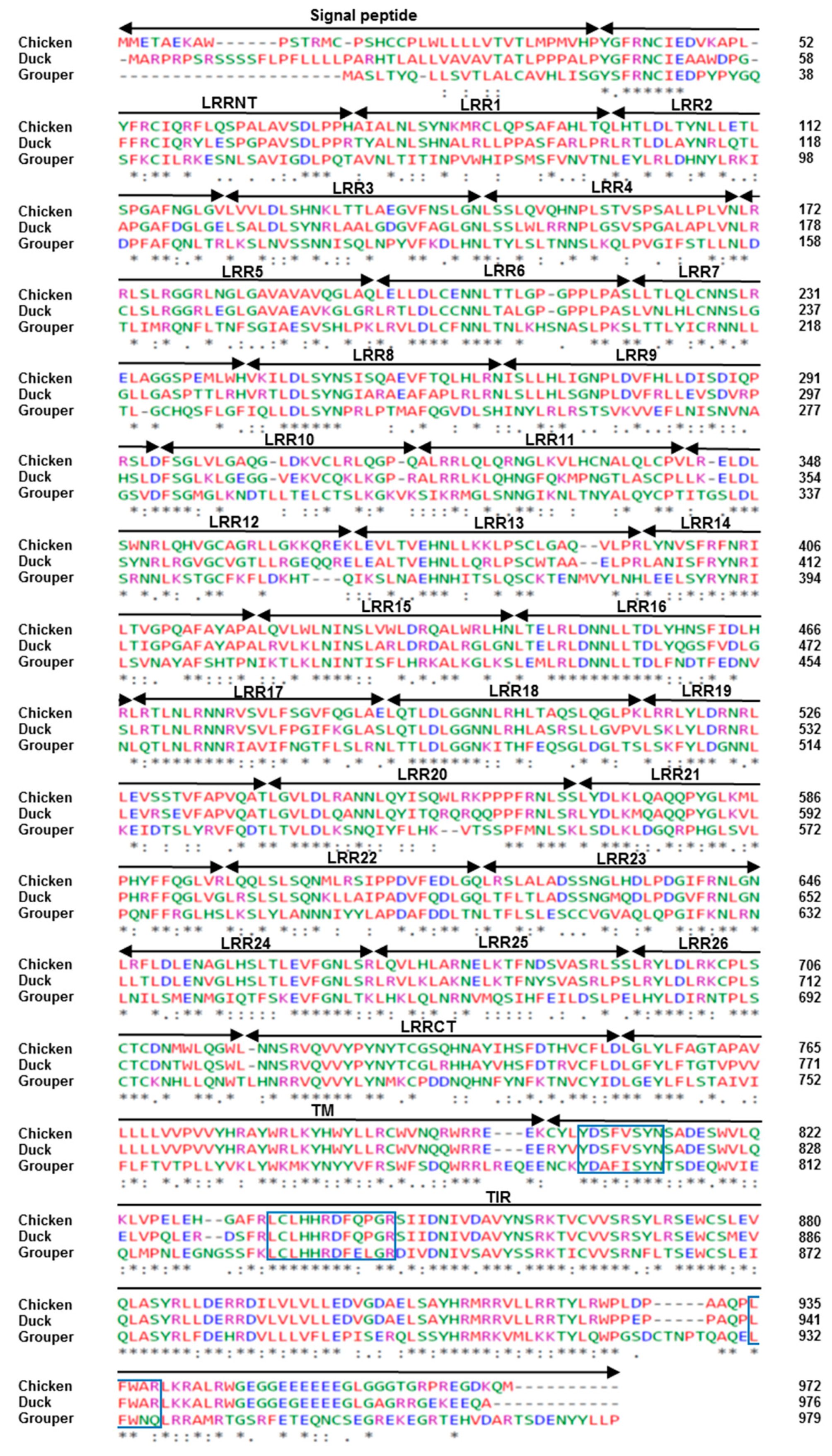
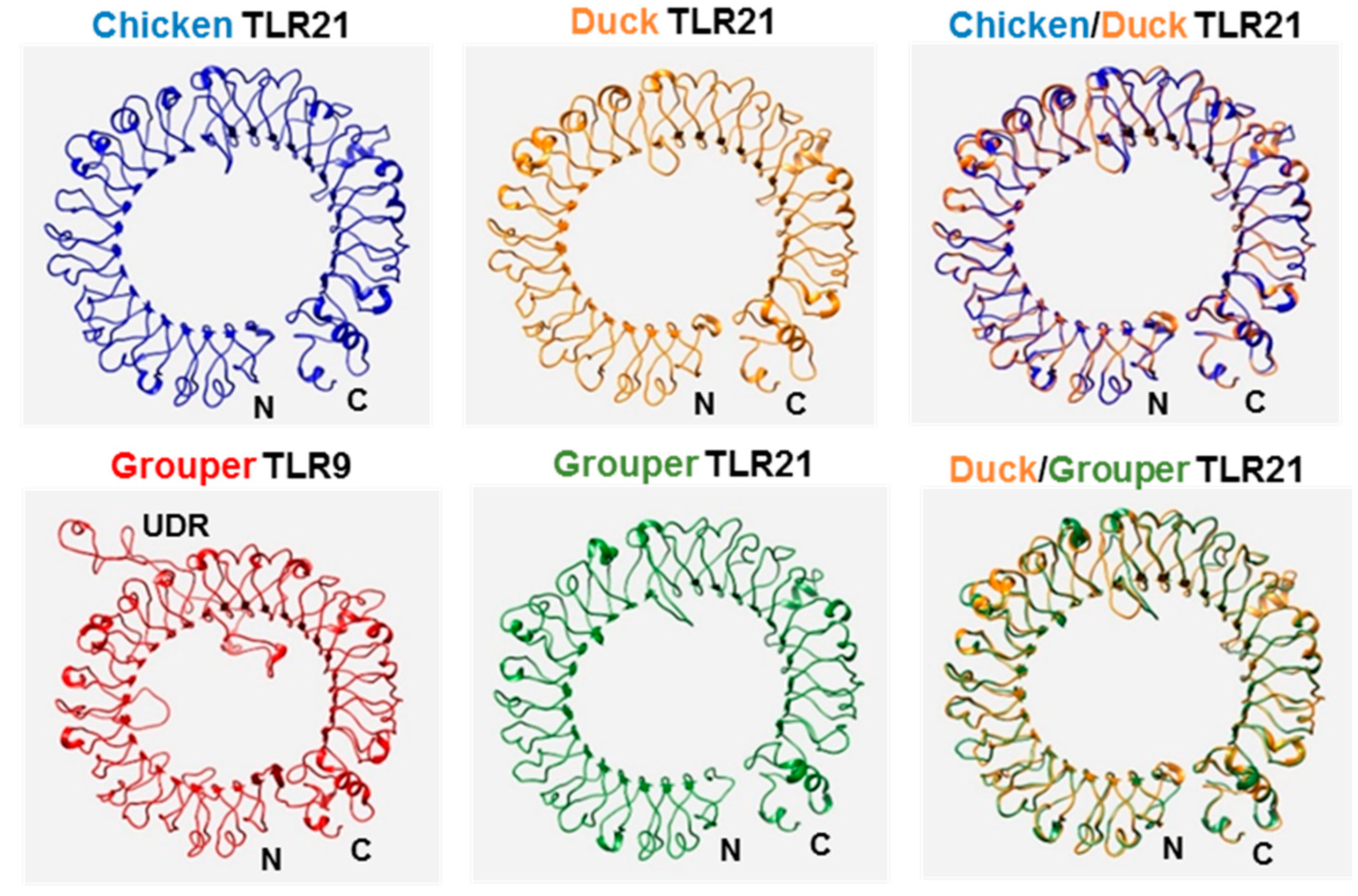
3.1. Characterization of Chicken and Duck TLR21s
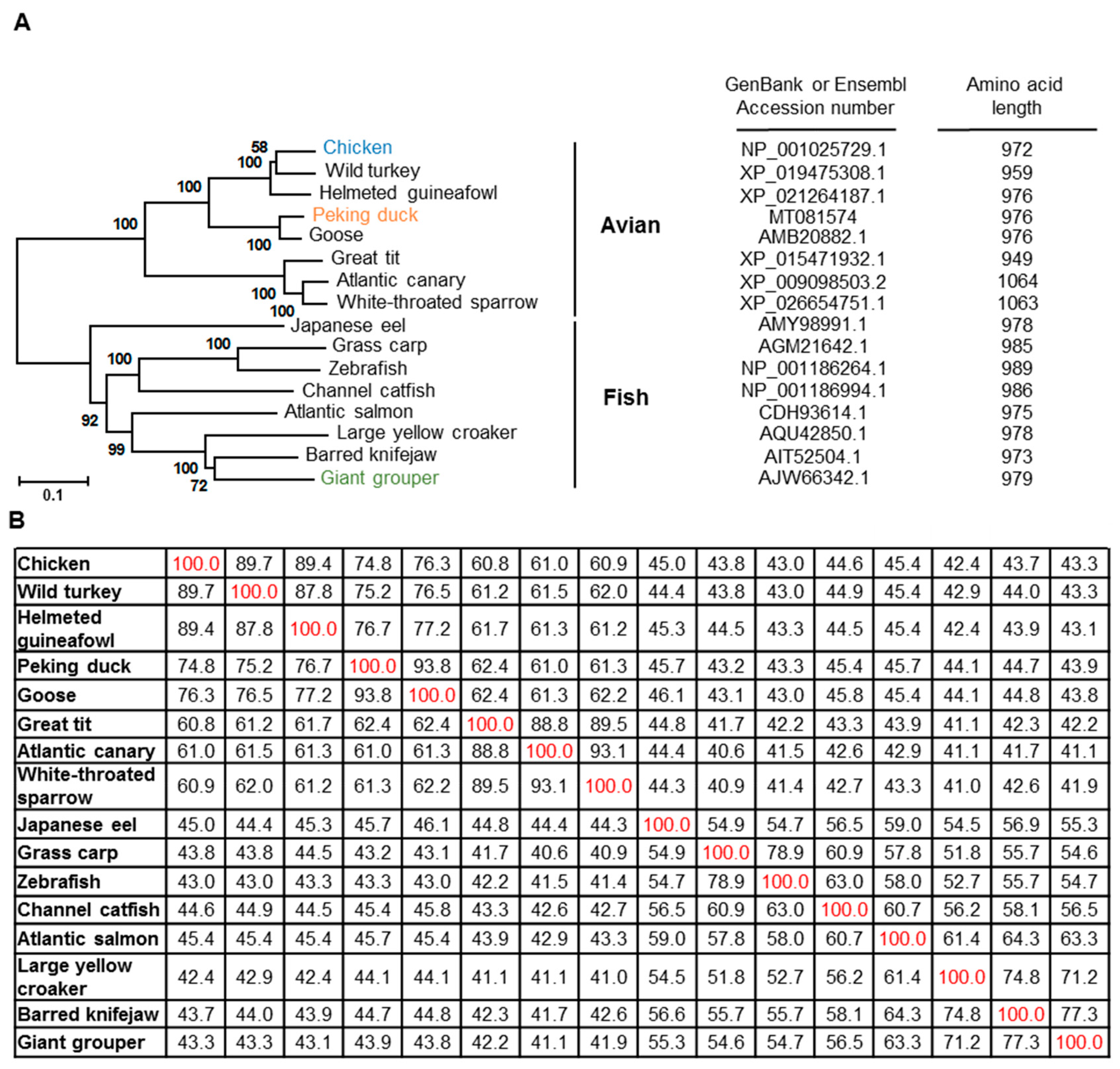
3.2. Phylogenetic Analysis of Chicken and Duck TLR21s
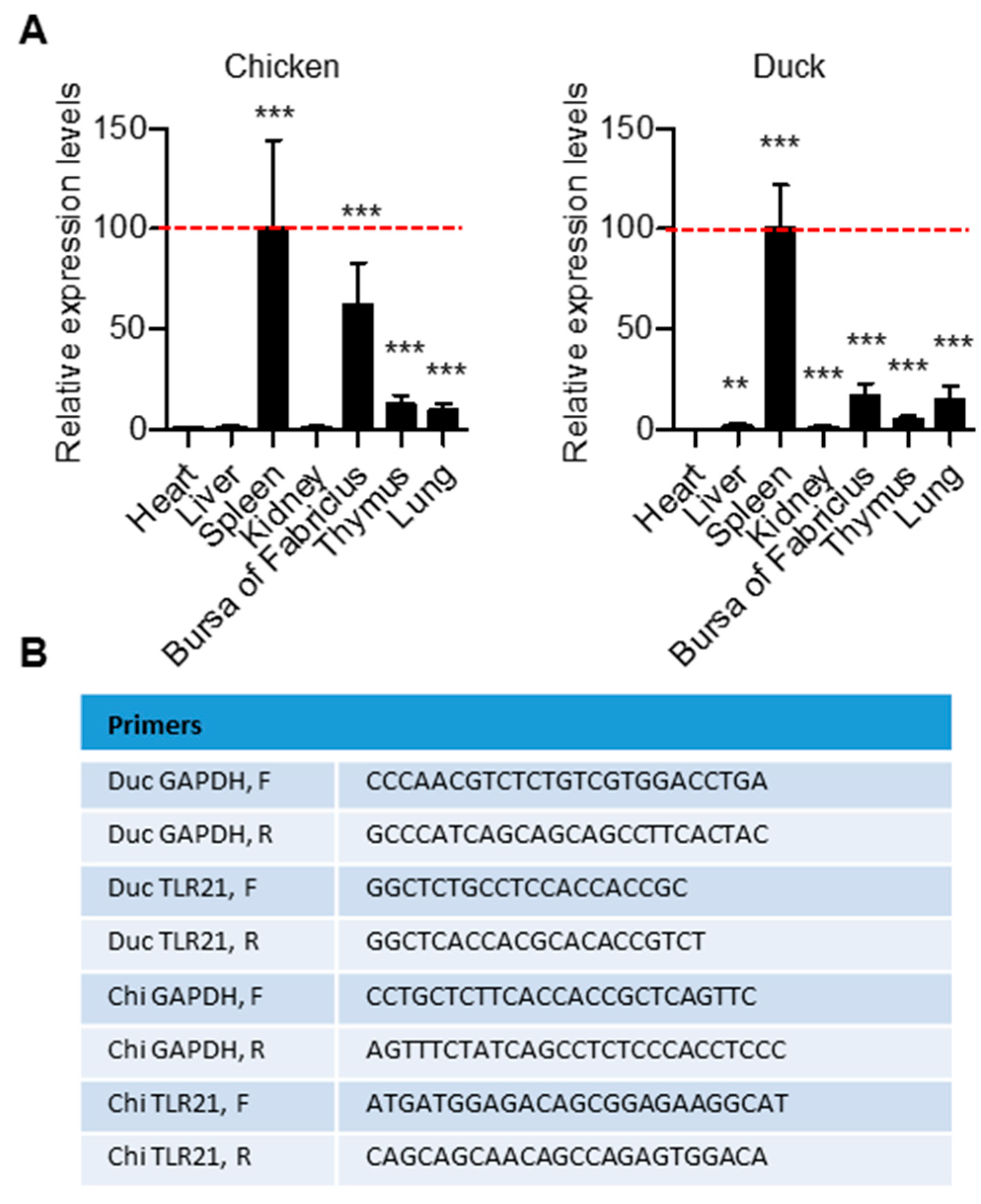
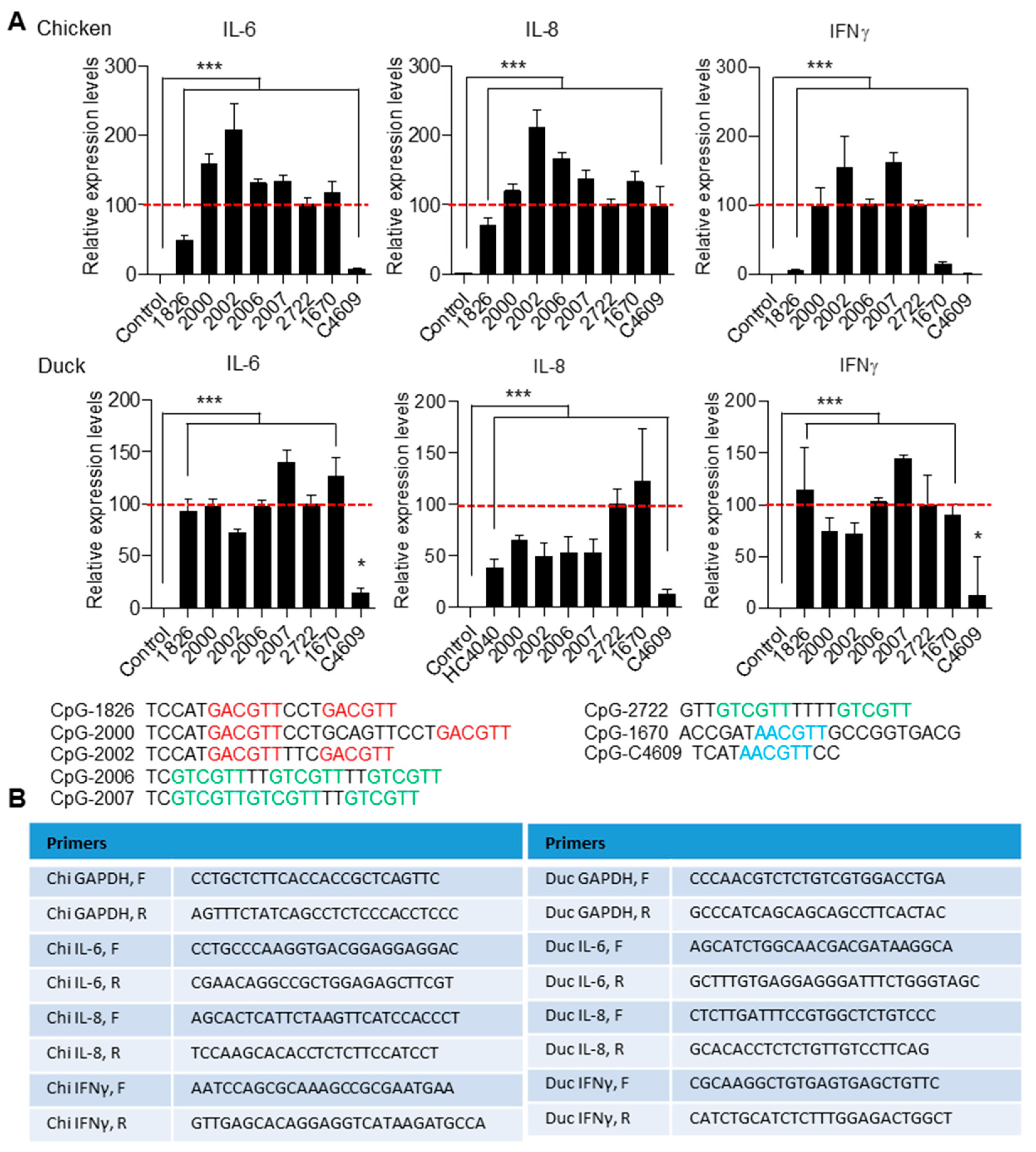
3.3. Tissue Distribution of TLR21 in Chicken and Duck and Activation Their Splenocytes by CpG-ODNs
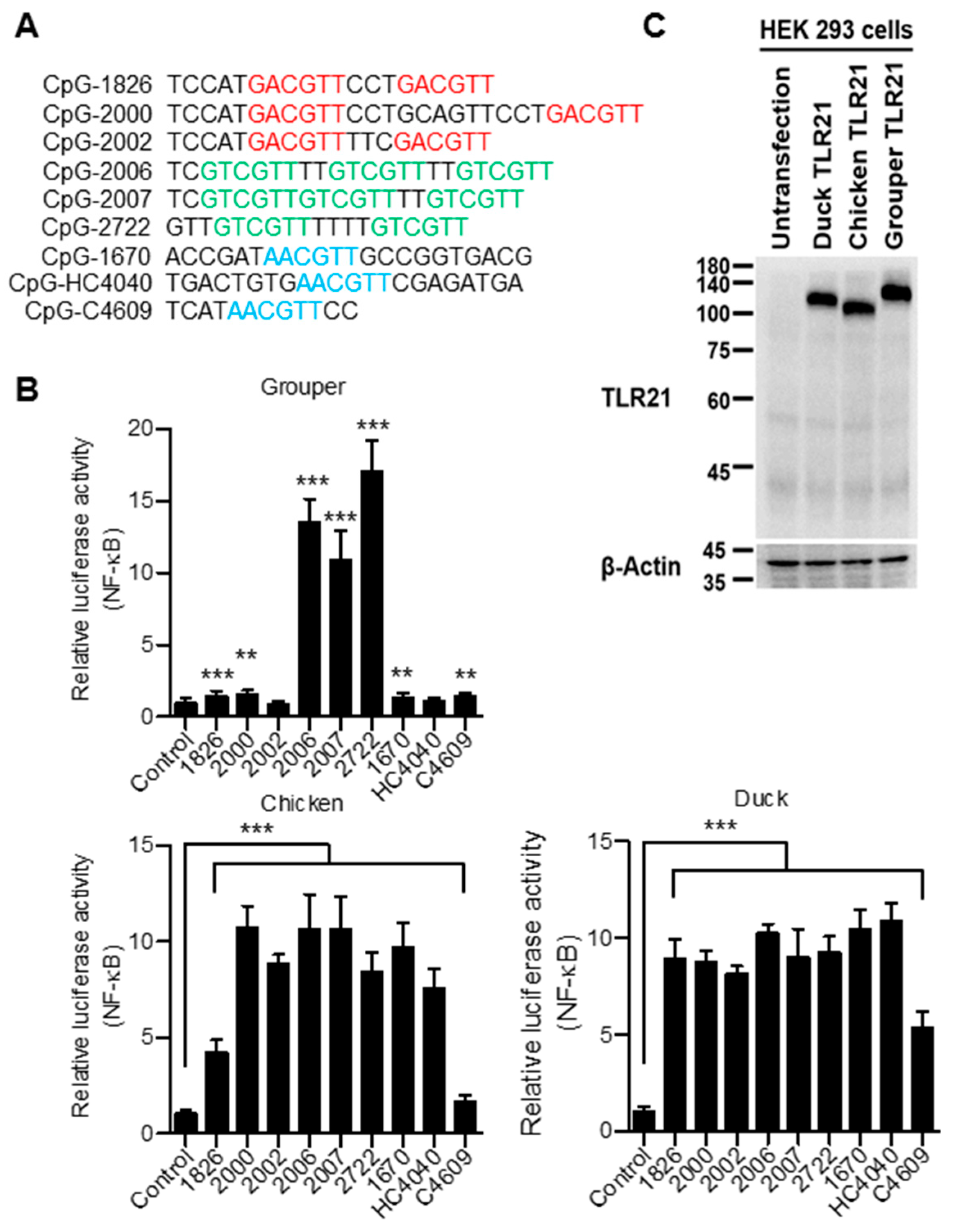
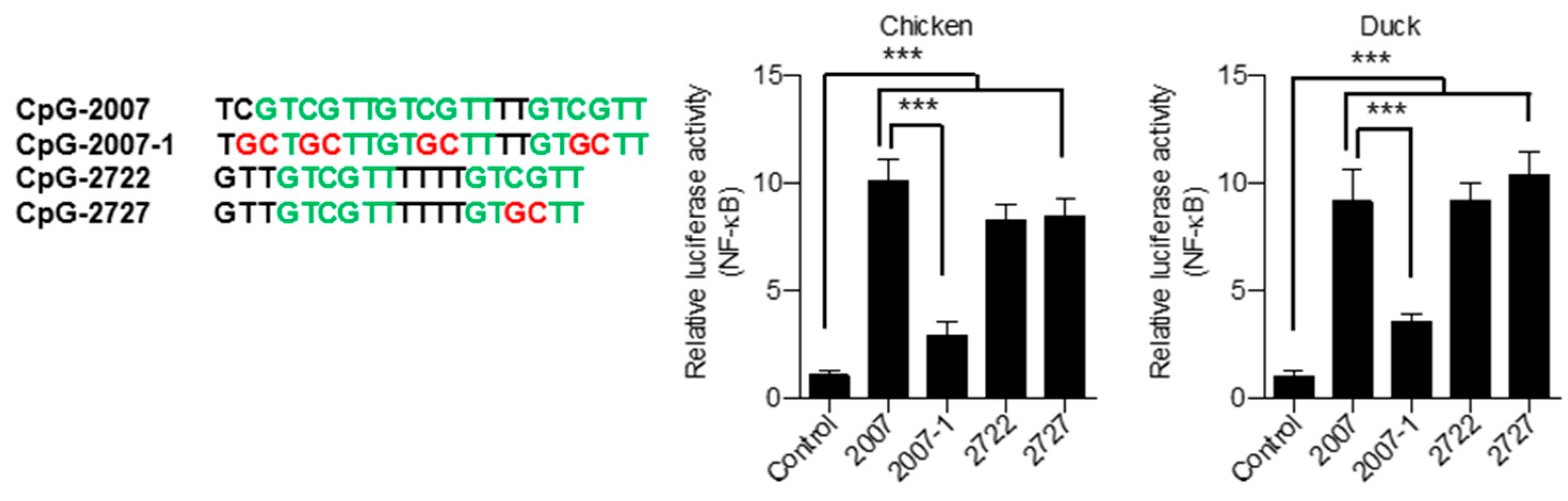
3.4. Broad CpG-ODN Sequence Recognition of Chicken and Duck TLR21s
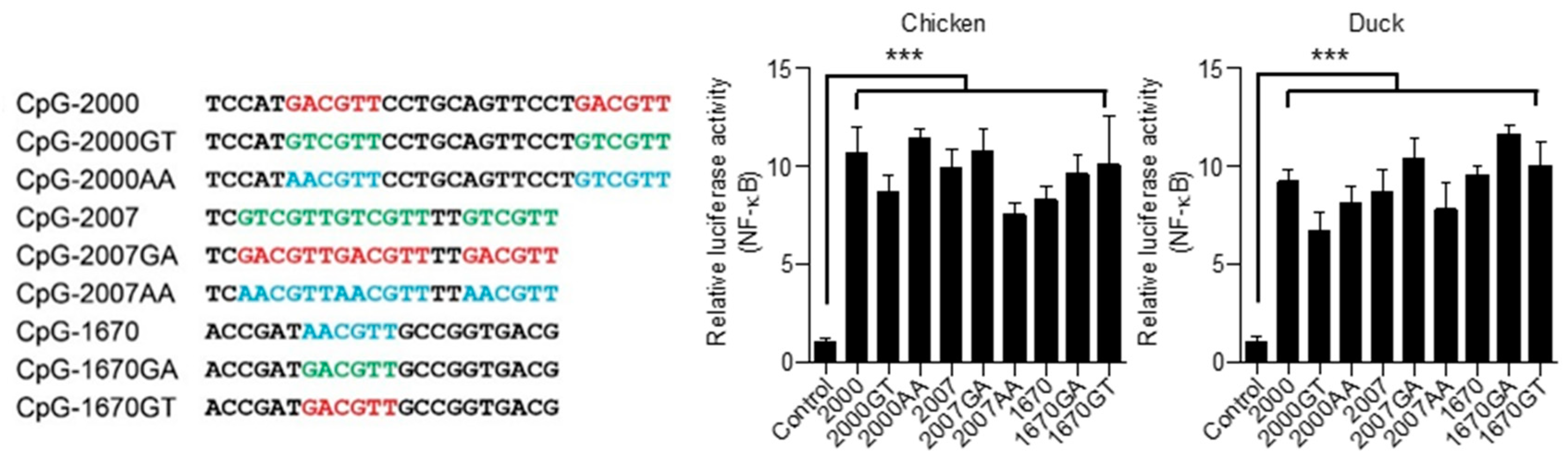
3.5. Chicken and Duck TLR21s Do Not Distinguish Different Types of CpG-hexamer Motifs
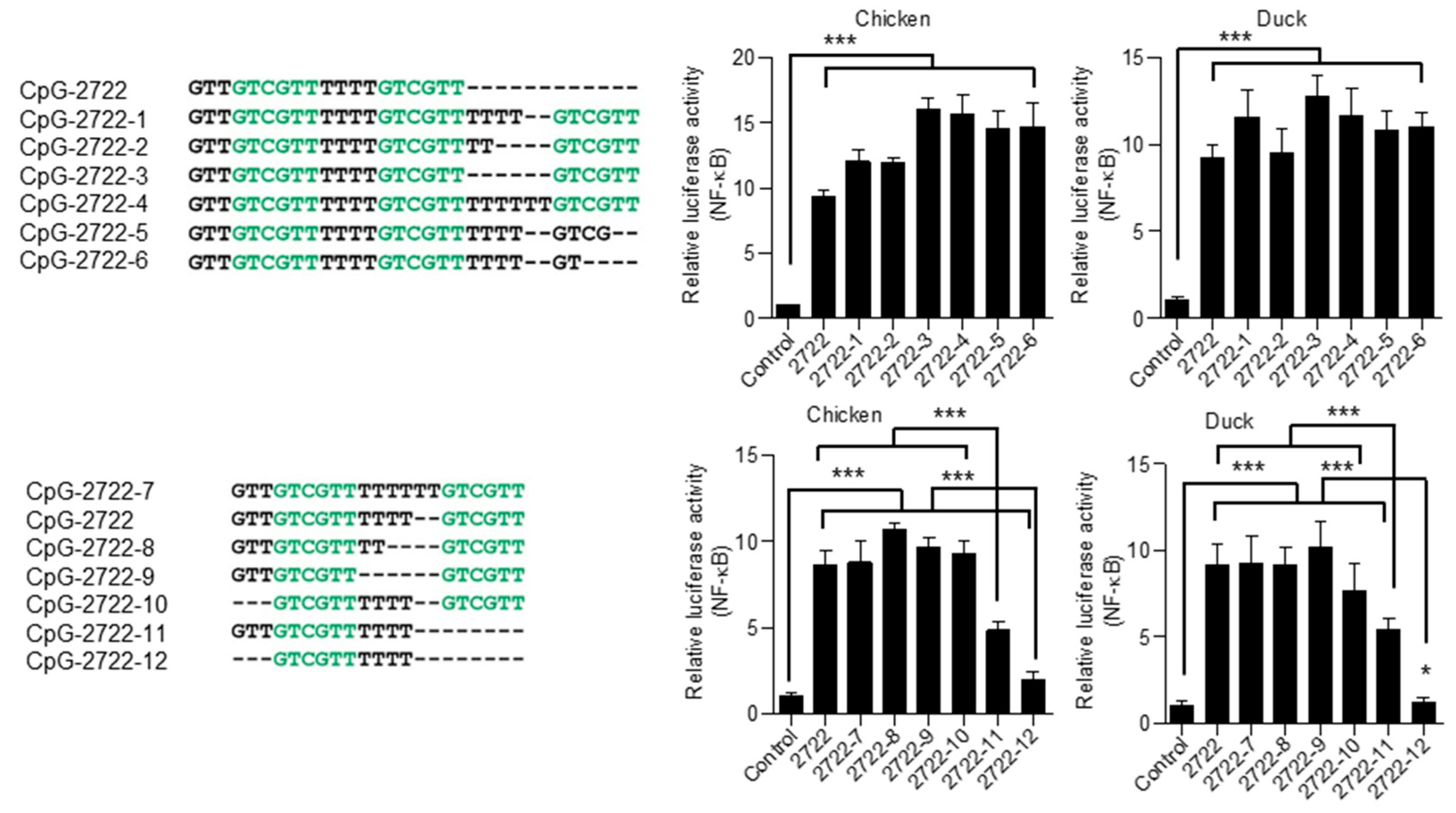
3.6. Responsiveness of Chicken and Duck TLR21s to CpG-ODNs with Different Lengths and Varied Spacing between Their CpG-hexamer Motifs
3.7. Requirement of CpG-dideoxynucleotides for Activation of Chicken and Duck TLR21s
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hafez, H.M.; Attia, Y.A. Challenges to the Poultry Industry: Current Perspectives and Strategic Future After the COVID-19 Outbreak. Front. Vet. Sci. 2020, 2020, 516. [Google Scholar] [CrossRef] [PubMed]
- Marangon, S.; Busani, L. The use of vaccination in poultry production. Rev. Sci. Techol. 2007, 26, 265–274. [Google Scholar] [CrossRef] [Green Version]
- Biggs, P.M. The world of poultry disease. Avian Pathol. 1982, 11, 281–300. [Google Scholar] [CrossRef] [PubMed]
- Takeda, K.; Kaisho, T.; Akira, S. Toll-like receptors. Annu. Rev. Immunol. 2003, 21, 335–376. [Google Scholar] [CrossRef] [PubMed]
- Hoebe, K.; Jiang, Z.; Georgel, P.; Tabeta, K.; Janssen, E.; Du, X.; Beutler, B. TLR signaling pathways: Opportunities for activation and blockade in pursuit of therapy. Curr. Pharm. Des. 2006, 12, 4123–4134. [Google Scholar] [CrossRef]
- Jin, M.S.; Lee, J.O. Structures of the toll-like receptor family and its ligand complexes. Immunity 2008, 29, 182–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miggin, S.M.; O’Neill, L.A. New insights into the regulation of TLR signaling. J. Leukoc. Biol. 2006, 80, 220–226. [Google Scholar] [CrossRef]
- Brownlie, R.; Allan, B. Avian toll-like receptors. Cell Tissue Res. 2011, 343, 121–130. [Google Scholar] [CrossRef]
- Neerukonda, S.N.; Katneni, U. Avian Pattern Recognition Receptor Sensing and Signaling. Vet. Sci. 2020, 7, 14. [Google Scholar] [CrossRef] [Green Version]
- Mbow, M.L.; Sarisky, R.T. Modulating toll-like receptor signalling as a novel antiinfective approach. Drug News Perspect 2005, 18, 179–184. [Google Scholar] [CrossRef]
- Chaung, H.C. CpGoligodeoxynucleotides as DNA adjuvants in vertebrates and their applications in immunotherapy. Int. Immunopharmacol. 2006, 6, 1586–1596. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.K.; Deb, R.; Dey, S.; Chellappa, M.M. Toll-like receptor-based adjuvants: Enhancing the immune response to vaccines against infectious diseases of chicken. Expert Rev. Vaccines 2014, 13, 909–925. [Google Scholar] [CrossRef]
- Akira, S. Mammalian Toll-like receptors. Curr. Opin. Immunol. 2003, 15, 5–11. [Google Scholar] [CrossRef]
- Rebl, A.; Goldammer, T.; Seyfert, H.M. Toll-like receptor signaling in bony fish. Vet. Immunol. Immunopathol. 2010, 134, 139–150. [Google Scholar] [CrossRef] [PubMed]
- Buchmann, K. Evolution of Innate Immunity: Clues from Invertebrates via Fish to Mammals. Front. Immunol. 2014, 5, 459. [Google Scholar] [CrossRef] [Green Version]
- Temperley, N.D.; Berlin, S.; Paton, I.R.; Griffin, D.K.; Burt, D.W. Evolution of the chicken Toll-like receptor gene family: A story of gene gain and gene loss. BMC Genom. 2008, 9, 62. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Cheng, A.; Wang, M. Innate sensing of viruses by pattern recognition receptors in birds. Vet. Res. 2013, 44, 82. [Google Scholar] [CrossRef] [Green Version]
- Nawab, A.; An, L.; Wu, J.; Li, G.; Liu, W.; Zhao, Y.; Wu, Q.; Xiao, M. Chicken toll-like receptors and their significance in immune response and disease resistance. Int. Rev. Immunol. 2019, 38, 284–306. [Google Scholar] [CrossRef]
- Brownlie, R.; Zhu, J.; Allan, B.; Mutwiri, G.K.; Babiuk, L.A.; Potter, A.; Griebel, P. Chicken TLR21 acts as a functional homologue to mammalian TLR9 in the recognition of CpGoligodeoxynucleotides. Mol. Immunol. 2009, 46, 3163–3170. [Google Scholar] [CrossRef]
- Keestra, A.M.; de Zoete, M.R.; Bouwman, L.I.; van Putten, J.P. Chicken TLR21 is an innate CpG DNA receptor distinct from mammalian TLR9. J. Immunol. 2010, 185, 460–467. [Google Scholar] [CrossRef] [Green Version]
- Yeh, D.W.; Liu, Y.L.; Lo, Y.C.; Yuh, C.H.; Yu, G.Y.; Lo, J.F.; Luo, Y.; Xiang, R.; Chuang, T.H. Toll-like receptor 9 and 21 have different ligand recognition profiles and cooperatively mediate activity of CpG-oligodeoxynucleotides in zebrafish. Proc. Natl. Acad. Sci. USA 2013, 110, 20711–20716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeh, D.W.; Lai, C.Y.; Liu, Y.L.; Lu, C.H.; Tseng, P.H.; Yuh, C.H.; Yu, G.Y.; Liu, S.J.; Leng, C.H.; Chuang, T.H. CpG-oligodeoxynucleotides developed for grouper toll-like receptor (TLR) 21s effectively activate mouse and human TLR9s mediated immune responses. Sci. Rep. 2017, 7, 17297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, C.Y.; Yu, G.Y.; Luo, Y.; Xiang, R.; Chuang, T.H. Immunostimulatory Activities of CpG-Oligodeoxynucleotides in Teleosts: Toll-Like Receptors 9 and 21. Front. Immunol. 2019, 10, 179. [Google Scholar] [CrossRef] [PubMed]
- Tokunaga, T.; Yamamoto, H.; Shimada, S.; Abe, H.; Fukuda, T.; Fujisawa, Y.; Furutani, Y.; Yano, O.; Kataoka, T.; Sudo, T.; et al. Antitumor activity of deoxyribonucleic acid fraction from Mycobacterium bovis BCG. I. Isolation, physicochemical characterization, and antitumor activity. J. Natl. Cancer Inst. 1984, 72, 955–962. [Google Scholar] [PubMed]
- Shimada, S.; Yano, O.; Inoue, H.; Kuramoto, E.; Fukuda, T.; Yamamoto, H.; Kataoka, T.; Tokunaga, T. Antitumor activity of the DNA fraction from Mycobacterium bovis BCG. II. Effects on various syngeneic mouse tumors. J. Natl. Cancer Inst. 1985, 74, 681–688. [Google Scholar]
- Krieg, A.M.; Yi, A.K.; Matson, S.; Waldschmidt, T.J.; Bishop, G.A.; Teasdale, R.; Koretzky, G.A.; Klinman, D.M. CpG motifs in bacterial DNA trigger direct B-cell activation. Nature 1995, 374, 546–549. [Google Scholar] [CrossRef]
- Roman, M.; Martin-Orozco, E.; Goodman, J.S.; Nguyen, M.D.; Sato, Y.; Ronaghy, A.; Kornbluth, R.S.; Richman, D.D.; Carson, D.A.; Raz, E. Immunostimulatory DNA sequences function as T helper-1-promoting adjuvants. Nat. Med. 1997, 3, 849–854. [Google Scholar] [CrossRef]
- Krieg, A.M. CpG motifs in bacterial DNA and their immune effects. Annu. Rev. Immunol. 2002, 20, 709–760. [Google Scholar] [CrossRef]
- Pisetsky, D.S. Mechanisms of immune stimulation by bacterial DNA. Springer Semin. Immunopathol. 2000, 22, 21–33. [Google Scholar] [CrossRef]
- Yamamoto, S.; Yamamoto, T.; Tokunaga, T. The discovery of immunostimulatory DNA sequence. Springer Semin. Immunopathol. 2000, 22, 11–19. [Google Scholar] [CrossRef]
- Rankin, R.; Pontarollo, R.; Ioannou, X.; Krieg, A.M.; Hecker, R.; Babiuk, L.A.; -van den Hurk, S.v.d.L. CpG motif identification for veterinary and laboratory species demonstrates that sequence recognition is highly conserved. Antisense Nucleic Acid Drug Dev. 2001, 11, 333–340. [Google Scholar] [CrossRef]
- Klinman, D.M.; Yi, A.K.; Beaucage, S.L.; Conover, J.; Krieg, A.M. CpG motifs present in bacteria DNA rapidly induce lymphocytes to secrete interleukin 6, interleukin 12, and interferon gamma. Proc. Natl. Acad. Sci. USA 1996, 93, 2879–2883. [Google Scholar] [CrossRef] [Green Version]
- Ballas, Z.K.; Rasmussen, W.L.; Krieg, A.M. Induction of NK activity in murine and human cells by CpG motifs in oligodeoxynucleotides and bacterial DNA. J. Immunol. 1996, 157, 1840–1845. [Google Scholar] [PubMed]
- Chu, R.S.; Targoni, O.S.; Krieg, A.M.; Lehmann, P.V.; Harding, C.V. CpGoligodeoxynucleotides act as adjuvants that switch on T helper 1 (Th1) immunity. J. Exp. Med. 1997, 186, 1623–1631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klinman, D.M.; Klaschik, S.; Sato, T.; Tross, D. CpG oligonucleotides as adjuvants for vaccines targeting infectious diseases. Adv. Drug Deliv. Rev. 2009, 61, 248–255. [Google Scholar] [CrossRef]
- Bode, C.; Zhao, G.; Steinhagen, F.; Kinjo, T.; Klinman, D.M. CpG DNA as a vaccine adjuvant. Expert Rev. Vaccines 2011, 10, 499–511. [Google Scholar] [CrossRef] [Green Version]
- Carrington, A.C.; Secombes, C.J. A review of CpGs and their relevance to aquaculture. Vet. Immunol. Immunopathol. 2006, 112, 87–101. [Google Scholar] [CrossRef]
- Mutwiri, G. TLR9 agonists: Immune mechanisms and therapeutic potential in domestic animals. Vet. Immunol. Immunopathol. 2012, 148, 85–89. [Google Scholar] [CrossRef]
- Wu, H.; Wang, H.; Jiang, W.; Lian, Z. The evolutionary characteristics and structural biology of Gallus toll-like receptor 21. J. Mol. Recognit 2018, 31, e2696. [Google Scholar] [CrossRef] [Green Version]
- Velova, H.; Gutowska-Ding, M.W.; Burt, D.W.; Vinkler, M. Toll-Like Receptor Evolution in Birds: Gene Duplication, Pseudogenization, and Diversifying Selection. Mol. Biol. Evol. 2018, 35, 2170–2184. [Google Scholar] [CrossRef]
- Chuang, T.H.; Lai, C.Y.; Tseng, P.H.; Yuan, C.J.; Hsu, L.C. Development of CpG-oligodeoxynucleotides for effective activation of rabbit TLR9 mediated immune responses. PLoS ONE 2014, 9, e108808. [Google Scholar] [CrossRef] [PubMed]
- Chuang, T.H.; Ulevitch, R.J. Cloning and characterization of a sub-family of human toll-like receptors: hTLR7, hTLR8 and hTLR9. Eur. Cytokine Netw. 2000, 11, 372–378. [Google Scholar] [PubMed]
- Botos, I.; Segal, D.M.; Davies, D.R. The structural biology of Toll-like receptors. Structure 2011, 19, 447–459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, D.; Li, W. Structures and recognition modes of toll-like receptors. Proteins 2017, 85, 3–9. [Google Scholar] [CrossRef]
- Liu, J.; Xu, C.; Hsu, L.C.; Luo, Y.; Xiang, R.; Chuang, T.H. A five-amino-acid motif in the undefined region of the TLR8 ectodomain is required for species-specific ligand recognition. Mol. Immunol. 2010, 47, 1083–1090. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Xu, C.; Liu, Y.L.; Matsuo, H.; Hsieh, R.P.; Lo, J.F.; Tseng, P.H.; Yuan, C.J.; Luo, Y.; Xiang, R.; et al. Activation of rabbit TLR9 by different CpG-ODN optimized for mouse and human TLR9. Comp. Immunol. Microbiol. Infect. Dis 2012, 35, 443–451. [Google Scholar] [CrossRef]
- Slany, M.; Ulmann, V.; Slana, I. Avian Mycobacteriosis: Still Existing Threat to Humans. BioMed. Res. Int. 2016, 2016, 4387461. [Google Scholar] [CrossRef] [Green Version]
- Naguib, M.M.; Verhagen, J.H.; Mostafa, A.; Wille, M.; Li, R.; Graaf, A.; Jarhult, J.D.; Ellstrom, P.; Zohari, S.; Lundkvist, A.; et al. Global patterns of avian influenza A (H7): Virus evolution and zoonotic threats. FEMS Microbiol. Rev. 2019, 43, 608–621. [Google Scholar] [CrossRef] [Green Version]
- Wanke, R.; Schmidt, P.; Erhard, M.H.; Sprick-Sanjose Messing, A.; Stangassinger, M.; Schmahl, W.; Hermanns, W. Freund’s complete adjuvant in the chicken: Efficient immunostimulation with severe local inflammatory reaction. Zentralbl. Veterinarmed. Reihe A 1996, 43, 243–253. [Google Scholar]
- Gupta, R.K. Aluminum compounds as vaccine adjuvants. Adv. Drug Deliv. Rev. 1998, 32, 155–172. [Google Scholar] [CrossRef]
- Marano, N.; Rupprecht, C.; Regnery, R. Vaccines for emerging infections. Rev. Sci. Techol. 2007, 26, 203–215. [Google Scholar] [CrossRef]
- Devlin, J.M.; Vaz, P.K.; Coppo, M.J.; Browning, G.F. Impacts of poultry vaccination on viruses of wild bird. Curr. Opin. Virol. 2016, 19, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Campbell, J.D. Development of the CpG Adjuvant 1018: A Case Study. Methods Mol. Biol. 2017, 1494, 15–27. [Google Scholar]
- Hyer, R.N.; Janssen, R.S. Immunogenicity and safety of a 2-dose hepatitis B vaccine, HBsAg/CpG 1018, in persons with diabetes mellitus aged 60-70years. Vaccine 2019, 37, 5854–5861. [Google Scholar] [CrossRef] [PubMed]
- Dar, A.; Tikoo, S.; Potter, A.; Babiuk, L.A.; Townsend, H.; Gerdts, V.; Mutwiri, G. CpG-ODNs induced changes in cytokine/chemokines genes expression associated with suppression of infectious bronchitis virus replication in chicken lungs. Vet. Immunol. Immunopathol. 2014, 160, 209–217. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.M.; Alkie, T.N.; Hodgins, D.C.; Nagy, E.; Shojadoost, B.; Sharif, S. Systemic immune responses to an inactivated, whole H9N2 avian influenza virus vaccine using class B CpG oligonucleotides in chickens. Vaccine 2015, 33, 3947–3952. [Google Scholar] [CrossRef] [PubMed]
- Taha-Abdelaziz, K.; Hodgins, D.C.; Alkie, T.N.; Quinteiro-Filho, W.; Yitbarek, A.; Astill, J.; Sharif, S. Oral administration of PLGA-encapsulated CpG ODN and Campylobacter jejuni lysate reduces cecal colonization by Campylobacter jejuni in chickens. Vaccine 2018, 36, 388–394. [Google Scholar] [CrossRef]
- Goonewardene, K.B.; Popowich, S.; Gunawardana, T.; Gupta, A.; Kurukulasuriya, S.; Karunarathna, R.; Chow-Lockerbie, B.; Ahmed, K.A.; Tikoo, S.K.; Foldvari, M.; et al. Intrapulmonary Delivery of CpG-ODN Microdroplets Provides Protection Against Escherichia coli Septicemia in Neonatal Broiler Chickens. Avian Dis. 2017, 61, 503–511. [Google Scholar] [CrossRef]
- De Silva Senapathi, U.; Abdul-Cader, M.S.; Amarasinghe, A.; van Marle, G.; Czub, M.; Gomis, S.; Abdul-Careem, M.F. The In Ovo Delivery of CpG Oligonucleotides Protects against Infectious Bronchitis with the Recruitment of Immune Cells into the Respiratory Tract of Chickens. Viruses 2018, 10, 635. [Google Scholar] [CrossRef] [Green Version]
- Sachan, S.; Dhama, K.; Latheef, S.K.; Samad, H.A.; Mariappan, A.K.; Munuswamy, P.; Singh, R.; Singh, K.P.; Malik, Y.S.; Singh, R.K. Immunomodulatory Potential of Tinosporacordifolia and CpG ODN (TLR21 Agonist) against the Very Virulent, Infectious Bursal Disease Virus in SPF Chicks. Vaccines 2019, 7, 106. [Google Scholar] [CrossRef] [Green Version]
- Talebi, A.; Arky-Rezai, S. Efficacy of CpG-ODN Administration Routes on Humoral Responses against Newcastle disease in Broilers. Arch. Razi Inst. 2019, 74, 357–364. [Google Scholar] [PubMed]
- Linghua, Z.; Xingshan, T.; Fengzhen, Z. Vaccination with Newcastle disease vaccine and CpGoligodeoxynucleotides induces specific immunity and protection against Newcastle disease virus in SPF chicken. Vet. Immunol. Immunopathol. 2007, 115, 216–222. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Shan, C.; Ming, S.; Liu, Y.; Du, Y.; Jiang, G. Immunoadjuvant effects of bacterial genomic DNA and CpGoligodeoxynucleotides on avian influenza virus subtype H5N1 inactivated oil emulsion vaccine in chicken. Res. Vet. Sci. 2009, 86, 399–405. [Google Scholar] [CrossRef]
- Yuk, S.S.; Lee, D.H.; Park, J.K.; To, E.O.; Kwon, J.H.; Noh, J.Y.; Gomis, S.; Song, C.S. Immune response in domestic ducks following intradermal delivery of inactivated vaccine against H5N1 highly pathogenic avian influenza virus adjuvanted with oligodeoxynucleotides containing CpG motifs. Poult. Sci. 2015, 94, 1836–1842. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.W.; Lin, Y.M.; Yen, T.Y.; Yang, W.J.; Chu, C.Y. CpGoligodeoxynucleotides containing GACGTT motifs enhance the immune responses elicited by a goose parvovirus vaccine in ducks. Vaccine 2010, 28, 7956–7962. [Google Scholar] [CrossRef]
- Chu, C.Y.; Liu, C.H.; Liou, J.J.; Lee, J.W.; Cheng, L.T. Development of a subunit vaccine containing recombinant Riemerellaanatipestifer outer membrane protein A and CpG ODN adjuvant. Vaccine 2015, 33, 92–99. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chuang, Y.-C.; Tseng, J.-C.; Yang, J.-X.; Liu, Y.-L.; Yeh, D.-W.; Lai, C.-Y.; Yu, G.-Y.; Hsu, L.-C.; Huang, C.-M.; Chuang, T.-H. Toll-Like Receptor 21 of Chicken and Duck Recognize a Broad Array of Immunostimulatory CpG-oligodeoxynucleotide Sequences. Vaccines 2020, 8, 639. https://doi.org/10.3390/vaccines8040639
Chuang Y-C, Tseng J-C, Yang J-X, Liu Y-L, Yeh D-W, Lai C-Y, Yu G-Y, Hsu L-C, Huang C-M, Chuang T-H. Toll-Like Receptor 21 of Chicken and Duck Recognize a Broad Array of Immunostimulatory CpG-oligodeoxynucleotide Sequences. Vaccines. 2020; 8(4):639. https://doi.org/10.3390/vaccines8040639
Chicago/Turabian StyleChuang, Yu-Chen, Jen-Chih Tseng, Jing-Xing Yang, Yi-Ling Liu, Da-Wei Yeh, Chao-Yang Lai, Guann-Yi Yu, Li-Chung Hsu, Chun-Ming Huang, and Tsung-Hsien Chuang. 2020. "Toll-Like Receptor 21 of Chicken and Duck Recognize a Broad Array of Immunostimulatory CpG-oligodeoxynucleotide Sequences" Vaccines 8, no. 4: 639. https://doi.org/10.3390/vaccines8040639