Dietary Betaine Reduces the Negative Effects of Cyclic Heat Exposure on Growth Performance, Blood Gas Status and Meat Quality in Broiler Chickens


 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Animals, Experimental Design and Diets
2.3. Slaughter and Objective Meat Quality Measures
2.4. Biochemical and Endocrinologal Analyses
2.5. Branch Chain Amino Acid
2.6. Statistical Analyses
3. Results
3.1. Growth Performance
3.2. Physiological Responses
3.3. Blood and Plasma Parameters and Muscle Betaine
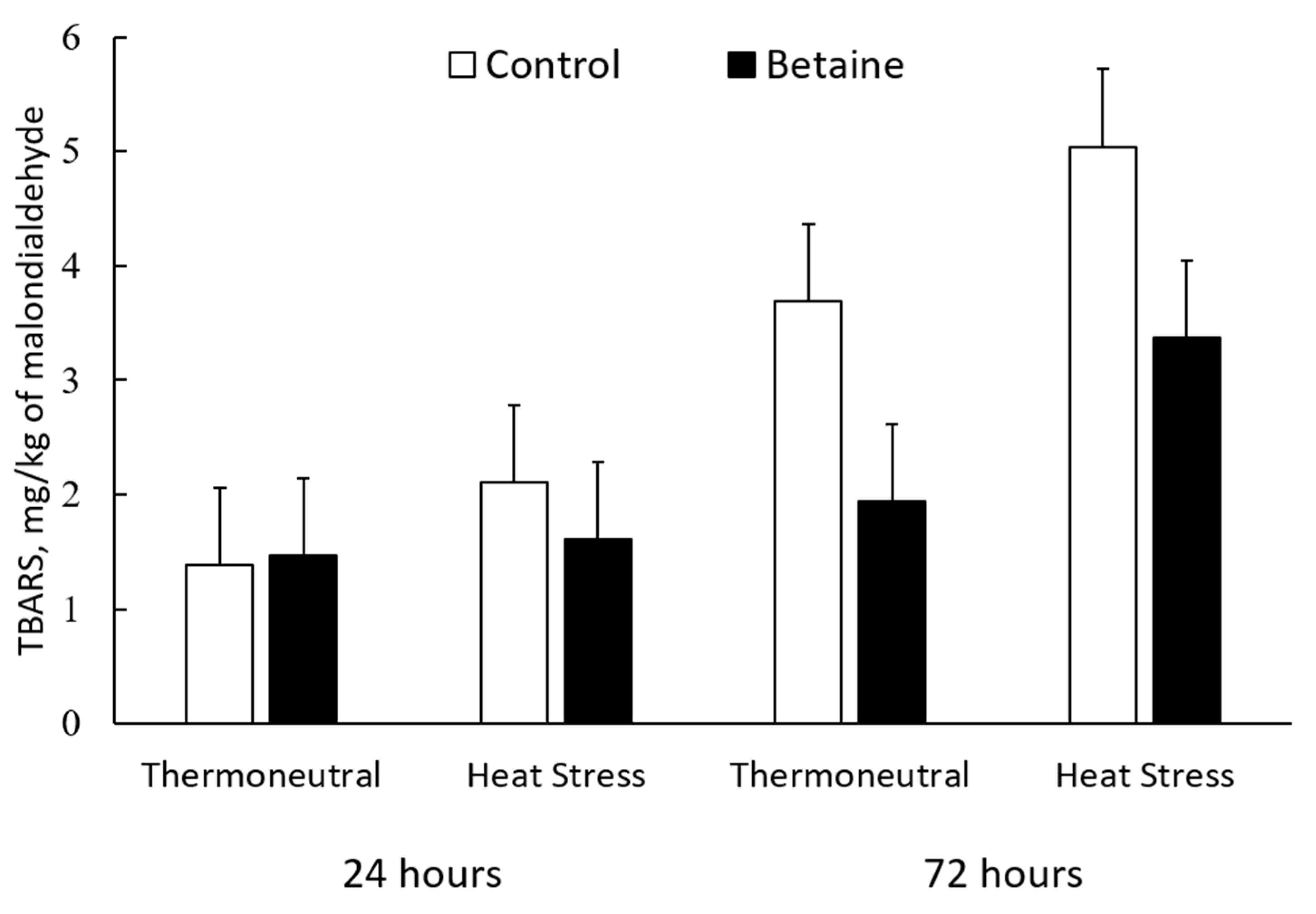
3.4. Objective Meat Quality
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Henry, B.; Charmley, E.d.; Eckard, R.; Gaughan, J.B.; Hegarty, R. Livestock production in a changing climate: Adaptation and mitigation research in Australia. Crop Pasture Sci. 2012, 63, 191–202. [Google Scholar] [CrossRef]
- Temim, S.; Chagneau, A.M.; Peresson, R.; Tesseraud, S. Chronic heat exposure alters protein turnover of three different skeletal muscles in finishing broiler chickens fed 20 or 25% protein diets. J. Nutr. 2000, 130, 813–819. [Google Scholar] [CrossRef] [Green Version]
- Akşit, M.; Yalcin, S.; Özkan, S.; Metin, K.; Özdemir, D. Effects of temperature during rearing and crating on stress parameters and meat quality of broilers. Poult. Sci. 2006, 85, 1867–1874. [Google Scholar] [CrossRef]
- Love, J.D.; Pearson, A.M. Lipid oxidation in meat and meat products—A review. J. Am. Oil Chem. Soc. 1971, 48, 547–549. [Google Scholar] [CrossRef]
- Huang, C.; Jiao, H.; Song, Z.; Zhao, J.; Wang, X.; Lin, H. Heat stress impairs mitochondria functions and induces oxidative injury in broiler chickens. J. Anim. Sci. 2015, 93, 2144–2153. [Google Scholar] [CrossRef] [PubMed]
- Shakeri, M.; Cottrell, J.J.; Wilkinson, S.; Zhao, W.; Le, H.H.; McQuade, R.; Furness, J.B.; Dunshea, F.R. Dietary Betaine Improves Intestinal Barrier Function and Ameliorates the Impact of Heat Stress in Multiple Vital Organs as Measured by Evans Blue Dye in Broiler Chickens. Animals 2020, 10, 38. [Google Scholar] [CrossRef] [Green Version]
- DiGiacomo, K.; Leury, B.J.; Dunshea, F.R. Potential nutritional strategies for the amelioration or prevention of high rigor temperature in cattle–a review. Anim. Prod. Sci. 2014, 54, 430–443. [Google Scholar] [CrossRef] [Green Version]
- Cottrell, J.J.; Ponnampalam, E.N.; Dunshea, F.R.; Warner, R.D. Effects of infusing nitric oxide donors and inhibitors on plasma metabolites, muscle lactate production and meat quality in lambs fed a high quality roughage-based diet. Meat Sci. 2015, 105, 8–15. [Google Scholar] [CrossRef]
- Craig, S.A.S. Betaine in human nutrition. Am. J. Clin. Nutr. 2004, 80, 539–549. [Google Scholar] [CrossRef] [Green Version]
- Alirezaei, M.; Gheisari, H.R.; Ranjbar, V.R.; Hajibemani, A. Betaine: A promising antioxidant agent for enhancement of broiler meat quality. Br. Poult. Sci. 2012, 53, 699–707. [Google Scholar] [CrossRef]
- Shakeri, M.; Cottrell, J.J.; Wilkinson, S.; Ringuet, M.; Furness, J.B.; Dunshea, F.R. Betaine and Antioxidants Improve Growth Performance, Breast Muscle Development and Ameliorate Thermoregulatory Responses to Cyclic Heat Exposure in Broiler Chickens. Animals 2018, 8, 162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alirezaei, M.; Khoshdel, Z.; Dezfoulian, O.; Rashidipour, M.; Taghadosi, V. Beneficial antioxidant properties of betaine against oxidative stress mediated by levodopa/benserazide in the brain of rats. J. Physiol. Sci. 2015, 65, 243–252. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Zhao, S.; Dai, S.; Liu, D.; Bokhari, S.G. Effects of dietary betaine on growth performance, fat deposition and serum lipids in broilers subjected to chronic heat stress. Anim. Sci. J. 2015, 86, 897–903. [Google Scholar] [CrossRef] [PubMed]
- Shakeri, M.; Cottrell, J.J.; Wilkinson, S.; Le, H.H.; Suleria, H.A.R.; Warner, R.D.; Dunshea, F.R. Growth Performance and Characterization of Meat Quality of Broiler Chickens Supplemented with Betaine and Antioxidants under Cyclic Heat Stress. Antioxidants 2019, 8, 336. [Google Scholar] [CrossRef] [Green Version]
- Fagan, J.M.; Sleczka, B.G.; Sohar, I. Quantitation of oxidative damage to tissue proteins. Int. J. Biochem. Cell Biol. 1999, 31, 751–757. [Google Scholar] [CrossRef]
- Nofal, M.E.; Magda, A.G.; Mousa, S.M.M.; Doaa, M.M.Y.; Bealsh, A.M.A. Effect of dietary betaine supplementation on productive, physiological and immunological performance and carcass characteristic of growing developed chicks uinder the condition of heat stress. Egypt Poult. Sci. 2015, 35, 237–259. [Google Scholar]
- Attia, Y.; Hassan, R.; Qota, E. Recovery from adverse effects of heat stress on slow-growing chicks in the tropics 1: Effect of ascorbic acid and different levels of betaine. Trop. Anim. Health Prod. 2009, 41, 807–818. [Google Scholar] [CrossRef]
- Schrama, J.; Heetkamp, M.; Simmins, P.; Gerrits, W. Dietary betaine supplementation affects energy metabolism of pigs. J. Anim. Sci. 2003, 81, 1202–1209. [Google Scholar] [CrossRef]
- Suster, D.; Leury, B.J.; King, R.H.; Mottram, M.; Dunshea, F.R. Interrelationships between porcine somatotropin (pST), betaine, and energy level on body composition and tissue distribution of finisher boars. Aust. J. Agric. Res. 2004, 55, 983–990. [Google Scholar] [CrossRef]
- Wolfe, R.R. Branched-chain amino acids and muscle protein synthesis in humans: Myth or reality? J. Int. Soc. Sports Nutr. 2017, 14, 30. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Zhang, Q.; Applegate, T. Impact of dietary branched chain amino acids concentration on broiler chicks during aflatoxicosis. Poult. Sci. 2016, 95, 1281–1289. [Google Scholar] [CrossRef] [PubMed]
- Nie, C.; He, T.; Zhang, W.; Zhang, G.; Ma, X. Branched chain amino acids: Beyond nutrition metabolism. Int. J. Mol. Sci. 2018, 19, 954. [Google Scholar] [CrossRef] [Green Version]
- Cooper, M.; Washburn, K. The relationships of body temperature to weight gain, feed consumption, and feed utilization in broilers under heat stress. Poult. Sci. 1998, 77, 237–242. [Google Scholar] [CrossRef] [PubMed]
- Settar, P.; Yalcin, S.; Turkmut, L.; Ozkan, S.; Cahanar, A. Season by genotype interaction related to broiler growth rate and heat tolerance. Poult. Sci. 1999, 78, 1353–1358. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.; Wen, J.; Zhang, H. Effect of chronic heat exposure on fat deposition and meat quality in two genetic types of chicken. Poult. Sci. 2007, 86, 1059–1064. [Google Scholar] [CrossRef]
- Yalcin, S.; Özkan, S.; Türkmut, L.; Siegel, P. Responses to heat stress in commercial and local broiler stocks. 1. Performance traits. Br. Poult. Sci. 2001, 42, 149–152. [Google Scholar] [CrossRef] [PubMed]
- Deeb, N.; Shlosberg, A.; Cahaner, A. Genotype-by-environment interaction with broiler genotypes differing in growth rate. 4. Association between responses to heat stress and to cold-induced ascites. Poult. Sci. 2002, 81, 1454–1462. [Google Scholar] [CrossRef]
- Collin, A.; Cassy, S.; Buyse, J.; Decuypere, E.; Damon, M. Potential involvement of mammalian and avian uncoupling proteins in the thermogenic effect of thyroid hormones. Domest. Anim. Endocrinol. 2005, 29, 78–87. [Google Scholar] [CrossRef]
- Bowen, S.; Washburn, K. Thyroid and adrenal response to heat stress in chickens and quail differing in heat tolerance. Poult. Sci. 1985, 64, 149–154. [Google Scholar] [CrossRef]
- Chiang, W.; Booren, A.; Strasburg, G. The effect of heat stress on thyroid hormone response and meat quality in turkeys of two genetic lines. Meat Sci. 2008, 80, 615–622. [Google Scholar] [CrossRef]
- Strasburg, G.; Chiang, W. Pale, soft, exudative turkey—The role of ryanodine receptor variation in meat quality. Poult. Sci. 2009, 88, 1497–1505. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Jia, G.; Zuo, J.; Zhang, Y.; Lei, J.; Ren, L.; Feng, D. Effects of constant and cyclic heat stress on muscle metabolism and meat quality of broiler breast fillet and thigh meat. Poult. Sci. 2012, 91, 2931–2937. [Google Scholar] [CrossRef] [PubMed]
- Sams, A. The effect of seasonal heat stress on rigor development and the incidence of pale, exudative turkey meat. Poult. Sci. 1997, 76, 1616–1620. [Google Scholar]
- Akter, Y.; Libinaki, R.; Hutchison, C.; Hopcroft, R.; Edwards, A.C.; Edwards, M.; O’Shea, C.J. Optimisation and investigations into the effect of a phosphorylated tocopherol mixture on growth performance, meat quality and plasma inflammatory biomarkers in broilers. Anim. Feed Sci. Technol. 2019, 253, 181–190. [Google Scholar] [CrossRef]
- Dai, S.; Gao, F.; Xu, X.; Zhang, W.; Song, S.; Zhou, G. Effects of dietary glutamine and gamma-aminobutyric acid on meat colour, pH, composition, and water-holding characteristic in broilers under cyclic heat stress. Br. Poult. Sci. 2012, 53, 471–481. [Google Scholar] [CrossRef]
- Hamm, R. Functional properties of the myofibrillar system and their measurements. In Muscle as Food, 1st ed.; Bechtel, P.J., Ed.; Academic Press: UniCambridge, MA, USA, 1986; pp. 135–199. [Google Scholar]
- Chauhan, S.; Celi, P.; Leury, B.; Clarke, I.; Dunshea, F. Dietary antioxidants at supranutritional doses improve oxidative status and reduce the negative effects of heat stress in sheep. J. Anim. Sci. 2014, 92, 3364–3374. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.; Cottrell, J.J.; Furness, J.B.; Rivera, L.R.; Kelly, F.W.; Wijesiriwardana, U.; Pustovit, R.V.; Fothergill, L.J.; Bravo, D.M.; Celi, P.; et al. Selenium and vitamin E together improve intestinal epithelial barrier function and alleviate oxidative stress in heat-stressed pigs. Exp. Physiol. 2016, 101, 801–810. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Pan, X.; Peng, Z. Effects of heat exposure on muscle oxidation and protein functionalities of pectoralis majors in broilers. Poult. Sci. 2009, 88, 1078–1084. [Google Scholar] [CrossRef]
- Mujahid, A.; Pumford, N.R.; Bottje, W.; Nakagawa, K.; Miyazawa, T.; Akiba, Y.; Toyomizu, M. Mitochondrial oxidative damage in chicken skeletal muscle induced by acute heat stress. J. Poult. Sci. 2007, 44, 439–445. [Google Scholar] [CrossRef] [Green Version]
- Imik, H.; Atasever, M.A.; Urcar, S.; Ozlu, H.; Gumus, R.; Atasever, M. Meat quality of heat stress exposed broilers and effect of protein and vitamin E. Br. Poult. Sci. 2012, 53, 689–698. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Temperature (T) | TN | HS | p-Value | |||||
|---|---|---|---|---|---|---|---|---|
| Diet (D) | CON | BET | CON | BET | SED 1 | T | D | T × D |
| Average daily weight gain, g/day | ||||||||
| 0 to 21 days | 33.6 | 34.0 | 33.4 | 31.8 | 1.52 | 0.27 | 0.57 | 0.32 |
| 21 to 35 days | 84.4 | 96.0 | 77.7 | 82.7 | 4.58 | 0.005 | 0.023 | 0.14 |
| 0 to 35 days | 53.9 | 58.8 | 51.1 | 52.1 | 2.42 | 0.009 | 0.094 | 0.26 |
| Average feed consumption, g/day | ||||||||
| 0 to 21 days | 48.7 | 50.4 | 45.4 | 45.1 | 5.93 | 0.33 | 0.86 | 0.81 |
| 21 to 35 days | 147 | 150 | 128 | 148 | 11.0 | 0.22 | 0.17 | 0.31 |
| 0 to 35 days | 87.9 | 90.2 | 78.5 | 86.2 | 7.48 | 0.23 | 0.36 | 0.62 |
| Feed conversion ratio | ||||||||
| 0 to 21 days | 1.46 | 1.48 | 1.45 | 1.43 | 0.162 | 0.78 | 0.98 | 0.85 |
| 21 to 35 days | 1.76 | 1.58 | 1.60 | 1.80 | 0.159 | 0.78 | 0.96 | 0.12 |
| 0 to 35 days | 1.65 | 1.53 | 1.54 | 1.66 | 0.131 | 0.93 | 0.95 | 0.23 |
| Slaughter weight, g | 1925 | 2097 | 1827 | 1862 | 85.1 | 0.009 | 0.092 | 0.26 |
| Temperature (T) | TN | HS | p-Value | |||||
|---|---|---|---|---|---|---|---|---|
| Diet (D) | CON | BET | CON | BET | SED 1 | T | D | T × D |
| Rectal temperature, °C | 41.6 | 41.3 | 42.7 | 42.0 | 0.11 | <0.001 | <0.001 | 0.005 |
| Respiration, bpm | 66.0 | 62.0 | 177 | 142 | 8.30 | <0.001 | 0.004 | 0.017 |
| pH | 7.27 | 7.41 | 7.45 | 7.38 | 0.034 | 0.017 | 0.09 | 0.003 |
| pCO2, mm Hg | 55.6 | 47.6 | 40.1 | 46.1 | 4.47 | 0.023 | 0.53 | 0.050 |
| Total CO2, mM | 27.4 | 31.5 | 29.8 | 28.5 | 1.76 | 0.79 | 0.17 | 0.054 |
| pO2, mm Hg | 30.6 | 35.4 | 32.8 | 34.2 | 4.38 | 0.87 | 0.29 | 0.59 |
| O2 saturation, % | 48.6 | 66.6 | 64.8 | 63.7 | 9.11 | 0.32 | 0.15 | 0.17 |
| HCO3, mM | 25.7 | 30.1 | 28.5 | 27.1 | 1.68 | 0.95 | 0.13 | 0.033 |
| Anion gap, mM | 20.8 | 14.8 | 15.0 | 18.3 | 1.28 | 0.26 | 0.050 | <0.001 |
| Base excess, mM | −1.18 | 4.95 | 4.23 | 1.73 | 1.78 | 0.40 | 0.080 | 0.006 |
| Temperature (T) | TN | HS | p-Value | |||||
|---|---|---|---|---|---|---|---|---|
| Diet (D) | CON | BET | CON | BET | SED 1 | T | D | T × D |
| Haematocrit, % | 19.5 | 17.3 | 19.0 | 18.7 | 0.917 | 0.50 | 0.050 | 0.17 |
| Hgb 2, g/dL | 6.65 | 5.88 | 6.40 | 6.40 | 0.296 | 0.53 | 0.058 | 0.09 |
| Potassium, mM | 6.78 | 6.13 | 5.20 | 6.07 | 0.453 | 0.029 | 1.00 | 0.040 |
| Sodium, mM | 151 | 149 | 148 | 148 | 1.1 | 0.056 | 0.18 | 0.13 |
| Chloride, mM | 112 | 110 | 110 | 109 | 1.7 | 0.35 | 0.43 | 0.82 |
| Calcium, mM | 1.51 | 1.50 | 1.48 | 1.47 | 0.042 | 0.25 | 0.78 | 0.88 |
| Lactate, mM | 10.8 | 5.19 | 3.76 | 6.70 | 1.348 | 0.017 | 0.069 | 0.001 |
| Glucose, mM | 14.2 | 14.0 | 15.1 | 14.4 | 0.53 | 0.09 | 0.33 | 0.58 |
| T3, pg/mL | 4.11 | 5.27 | 5.62 | 5.96 | 0.547 | 0.009 | 0.069 | 0.30 |
| T4, pg/mL | 6.95 | 6.29 | 9.56 | 9.75 | 1.64 | 0.016 | 0.85 | 0.72 |
| Plasma betaine, µmol/L | 133 | 164 | 102 | 140 | 10.9 | <0.001 | <0.001 | 0.66 |
| Muscle betaine, µmol/g | 246 | 174 | 131 | 194 | 30.2 | 0.047 | 0.83 | 0.008 |
| BCAA, nmol/uL | 3.25 | 2.09 | 1.98 | 1.82 | 0.204 | <0.001 | <0.001 | 0.002 |
| Temperature (T) | TN | HS | p-Value | |||||
|---|---|---|---|---|---|---|---|---|
| Diet (D) | CON | BET | CON | BET | SED 1 | T | D | T × D |
| Water content, % | 76.2 | 79.1 | 74.7 | 77.5 | 1.42 | 0.089 | 0.008 | 0.96 |
| Drip loss, % | 2.95 | 1.62 | 2.13 | 1.52 | 0.393 | 0.082 | 0.001 | 0.22 |
| Cooking loss, % | 22.8 | 21.2 | 21.5 | 20.3 | 1.69 | 0.36 | 0.26 | 0.88 |
| Peak shear force, N | 24.1 | 18.6 | 24.8 | 19.1 | 1.87 | 0.49 | <0.001 | 0.92 |
| Carbonyl, nmol/mg protein | 3.22 | 2.02 | 3.77 | 2.89 | 0.486 | 0.053 | 0.008 | 0.64 |
| MFI 2 | 1.88 | 2.04 | 1.72 | 1.93 | 0.105 | 0.081 | 0.017 | 0.74 |
| (75.2) | (109) | (52.0) | (85.1) | |||||
| Temperature (T) | TN | HS | p-Value | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Diet (D) | Time (TM) | CON | BET | CON | BET | SED 1 | T | D | TM |
| L* | 24 h | 55.5 | 58.6 | 55.7 | 55.2 | 2.22 | 0.48 | 0.20 | <0.001 |
| 72 h | 50.1 | 52.5 | 50.6 | 51.0 | |||||
| a* | 24 h | 1.99 | 2.22 | 1.54 | 2.40 | 0.605 | 0.27 | 0.10 | 0.022 |
| 72 h | 1.55 | 1.47 | 0.69 | 1.63 | |||||
| b* | 24 h | 6.07 | 6.09 | 5.11 | 5.69 | 0.781 | 0.34 | 0.21 | 0.015 |
| 72 h | 4.46 | 5.18 | 4.50 | 5.11 | |||||
| pH 2 | 10 min | 6.77 | 6.78 | 6.83 | 6.82 | 0.144 | 0.36 | 0.020 | <0.001 |
| 1 h | 6.44 | 6.57 | 6.60 | 6.50 | |||||
| 24 h | 5.72 | 6.30 | 5.60 | 5.82 | |||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shakeri, M.; Cottrell, J.J.; Wilkinson, S.; Le, H.H.; Suleria, H.A.R.; Warner, R.D.; Dunshea, F.R. Dietary Betaine Reduces the Negative Effects of Cyclic Heat Exposure on Growth Performance, Blood Gas Status and Meat Quality in Broiler Chickens. Agriculture 2020, 10, 176. https://doi.org/10.3390/agriculture10050176
Shakeri M, Cottrell JJ, Wilkinson S, Le HH, Suleria HAR, Warner RD, Dunshea FR. Dietary Betaine Reduces the Negative Effects of Cyclic Heat Exposure on Growth Performance, Blood Gas Status and Meat Quality in Broiler Chickens. Agriculture. 2020; 10(5):176. https://doi.org/10.3390/agriculture10050176
Chicago/Turabian StyleShakeri, Majid, Jeremy J. Cottrell, Stuart Wilkinson, Hieu H. Le, Hafiz A. R. Suleria, Robyn D. Warner, and Frank R. Dunshea. 2020. "Dietary Betaine Reduces the Negative Effects of Cyclic Heat Exposure on Growth Performance, Blood Gas Status and Meat Quality in Broiler Chickens" Agriculture 10, no. 5: 176. https://doi.org/10.3390/agriculture10050176