
Development and Application of a Loop-Mediated Isothermal Amplification (LAMP) Assay for the Detection of Pseudomonas syringae Pathovars pisi and syringae

Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Isolates
2.2. Bioinformatics Pipeline for LAMP Primer Design
2.3. LAMP Assay Validation
2.3.1. Specificity Analysis
2.3.2. Limit of Detection
2.4. LAMP as Surveillance and Diagnostic Tool
2.4.1. Early Detection of Bacterial Blight
2.4.2. Seed Testing
2.4.3. Field Sample Testing
3. Results
3.1. Primer Design and Development of the LAMP Assays
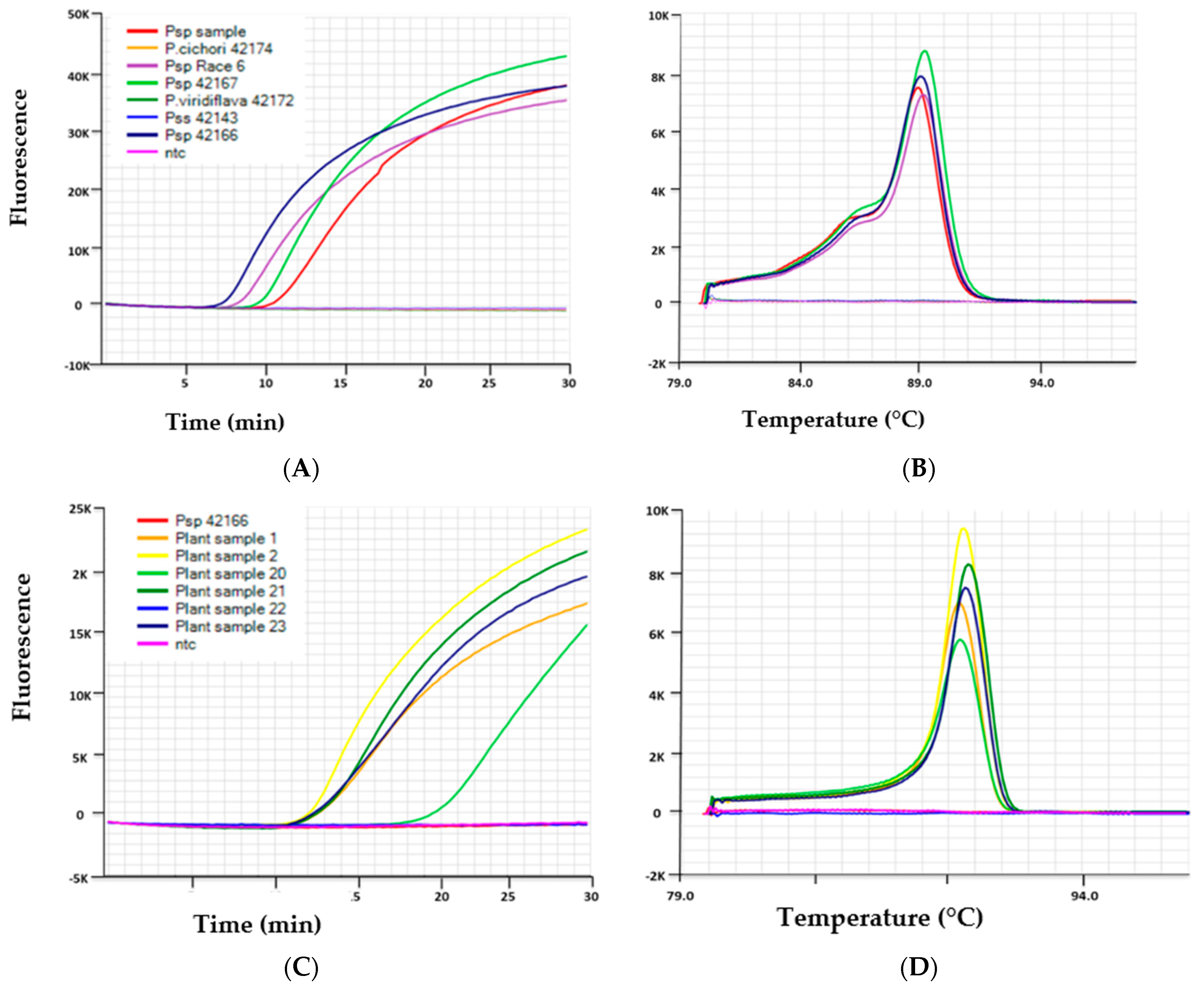
3.2. Specificity of the LAMP Assays
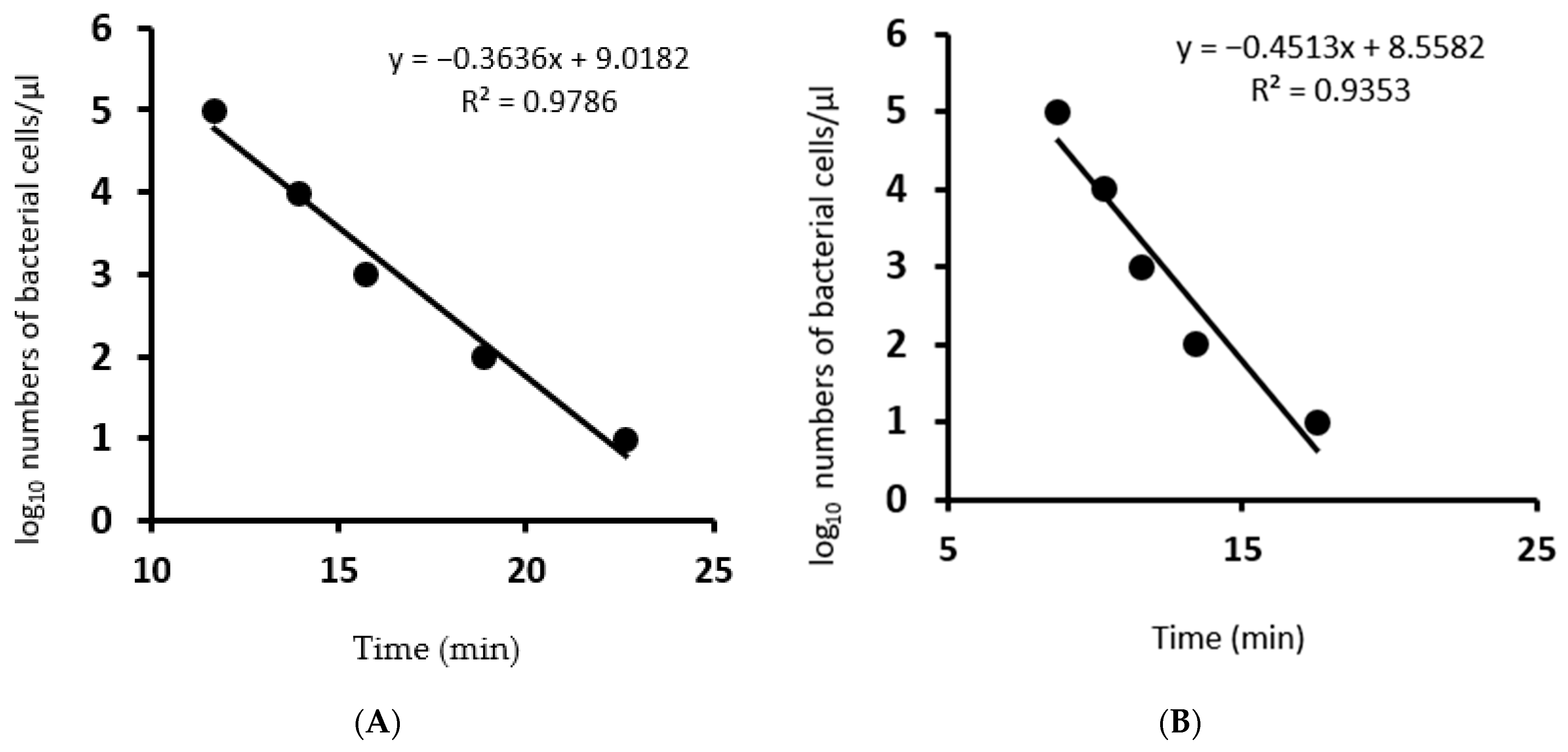
3.3. Limit of Detection
3.4. LAMP as Surveillance and Diagnostic Tool
3.4.1. Early Detection of Bacterial Blight
3.4.2. Detection of Bacteria from Seeds
3.4.3. Field Sample Testing
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Bull, C.T.; De Boer, S.H.; Denny, T.P.; Firrao, G.; Fisher-Le Saux, M.; Saddler, G.S.; Scortichini, M.; Stead, D.E.; Takikawa, Y. Comprehensive list of names of plant pathogenic bacteria, 1980–2007. J. Plant Pathol. 2010, 92, 551–592. [Google Scholar]
- Young, J. Taxonomy of Pseudomonas syringae. J. Plant Pathol. 2010, 92 (Suppl. 1), S5–S14. [Google Scholar]
- Martín-Sanz, A.; Palomo, J.; Pérez de la Vega, M.; Caminero, C. Identification of pathovars and races of Pseudomonas syringae, the main causal agent of bacterial disease in pea in North-Central Spain, and the search for disease resistance. Eur. J. Plant Pathol. 2011, 129, 57–69. [Google Scholar] [CrossRef]
- Richardson, H.J.; Hollaway, G.J. Bacterial blight caused by Pseudomonas syringae pv. syringae shown to be an important disease of field pea in south eastern Australia. Australas. Plant Pathol. 2011, 40, 9. [Google Scholar] [CrossRef]
- Roberts, S.; Phelps, K.; McKeown, B.; Heath, M.; Cockerell, V. Effect of pea bacterial blight (Pseudomonas syringae pv. pisi) on the yield of spring sown combining peas (Pisum sativum). Ann. Appl. Biol. 1995, 126, 61–73. [Google Scholar] [CrossRef]
- Grondeau, C.; Mabiala, A.; Ait-Oumeziane, R.; Samson, R. Epiphytic life is the main characteristic of the life cycle of Pseudomonas syringae pv. pisi, pea bacterial blight agent. Eur. J. Plant Pathol. 1996, 102, 353–363. [Google Scholar] [CrossRef]
- Taylor, J.; Dye, D. A survey of the organisms associated with bacterial blight of peas. N. Z. J. Agric. Res. 1972, 15, 432–440. [Google Scholar] [CrossRef] [Green Version]
- Bevan, J.R.; Taylor, J.D.; Crute, I.R.; Hunter, P.J.; Vivian, A. Genetics of specific resistance in pea (Pisum sativum) cultivars to seven races of Pseudomonas syringae pv. pisi. Plant Pathol. 1995, 44, 98–108. [Google Scholar] [CrossRef]
- Hollaway, G.J.; Bretag, T.W.; Price, T.V. The epidemiology and management of bacterial blight (Pseudomonas syringae pv. pisi) of field pea (Pisum sativum) in Australia: A review. Aust. J. Agric. Res. 2007, 58, 1086–1099. [Google Scholar] [CrossRef]
- Martín-Sanz, A.; Pérez de la Vega, M.; Caminero, C. Resistance to Pseudomonas syringae in a collection of pea germplasm under field and controlled conditions. Plant Pathol. 2011, 61, 375–387. [Google Scholar] [CrossRef]
- Bradbury, J. Guide to Plant Pathogenic Bacteria; CAB International Mycological Institute: London, UK, 1986; 334p. [Google Scholar]
- Martín-Sanz, A.; de la Vega, M.P.; Murillo, J.; Caminero, C. Strains of Pseudomonas syringae pv. syringae from pea are phylogenetically and pathogenically diverse. Phytopathology 2013, 103, 673–681. [Google Scholar] [CrossRef] [Green Version]
- Schaad, N.W.; Jones, J.B.; Chun, W. Laboratory Guide for the Identification of Plant Pathogenic Bacteria; APS Press: St. Paul, MN, USA, 2001. [Google Scholar]
- Lelliott, R.; Billing, E.; Hayward, A. A determinative scheme for the fluorescent plant pathogenic Pseudomonads. J. Appl. Bacteriol. 1966, 29, 470–489. [Google Scholar] [CrossRef]
- Arnold, D.L.; Athey-Pollard, A.; Gibbon, M.J.; Taylor, J.D.; Vivian, A. Specific oligonucleotide primers for the identification of Pseudomonas syringae pv. pisi yield one of two possible DNA fragments by PCR amplification: Evidence for phylogenetic divergence. Physiol. Mol. Plant Pathol. 1996, 49, 233–245. [Google Scholar] [CrossRef]
- Sorensen, K.N.; Kim, K.-H.; Takemoto, J.Y. PCR Detection of Cyclic Lipodepsinonapeptide-Producing Pseudomonas syringae pv. syringae and Similarity of Strains. Appl. Environ. Microbiol. 1998, 64, 226–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bultreys, A.; Gheysen, I. Biological and molecular detection of toxic lipodepsipeptide-producing Pseudomonas syringae strains and PCR identification in plants. Appl. Environ. Microbiol. 1999, 65, 1904–1909. [Google Scholar] [CrossRef] [Green Version]
- Best, N.; Rawlin, G.; Suter, R.; Rodoni, B.; Beddoe, T. Optimization of a Loop Mediated Isothermal Amplification (LAMP) Assay for In-Field Detection of Dichelobacter nodosus with aprV2 (VDN LAMP) in Victorian Sheep Flocks. Front. Vet. Sci. 2019, 6. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Nie, J.; Ward, L.J.; Nickerson, J.; De Boer, S.H. Development and evaluation of a loop-mediated isothermal amplification assay for rapid detection and identification of Pectobacterium atrosepticum. Can. J. Plant Pathol. 2011, 33, 447–457. [Google Scholar] [CrossRef]
- Rodda, M.; Kant, P.; Lindbeck, K.; Gnanasambandam, A.; Hollaway, G. A high-throughput glasshouse based screening method to evaluate bacterial blight resistance in field pea (Pisum sativum). Australas. Plant Pathol. 2015, 44, 515–526. [Google Scholar] [CrossRef]
- Mazarei, M.; Kerr, A. Distinguishing pathovars of Pseudomonas syringae on peas: Nutritional, pathogenicity and serological tests. Plant Pathol. 1990, 39, 278–285. [Google Scholar] [CrossRef]
- Jakob, K.; Kniskern, J.M.; Bergelson, J. The role of pectate lyase and the jasmonic acid defense response in Pseudomonas viridiflava virulence. Mol. Plant-Microbe Interact. 2007, 20, 146–158. [Google Scholar] [CrossRef] [Green Version]
- King, E.O.; Ward, M.K.; Raney, D.E. Two simple media for the demonstration of pyocyanin and fluorescin. J. Lab. Clin. Med. 1954, 44, 301–307. [Google Scholar] [PubMed]
- Harris, R.S. Improved Pairwise Alignmnet of Genomic DNA; The Pennsylvania State University: University Park, PA, USA, 2007. [Google Scholar]
- Chen, Y.; Ye, W.; Zhang, Y.; Xu, Y. High speed BLASTN: An accelerated MegaBLAST search tool. Nucleic Acids Res. 2015, 43, 7762–7768. [Google Scholar] [CrossRef] [Green Version]
- Busot, G.; Arif, M.; Stack, J. Genome analysis of nonpathogenic Pseudomonas syringae: Secretion systems, effectors and toxins. In Proceedings of the APS Annual Meeting, Pasadena, CA, USA, 1–5 August 2015; p. 758. [Google Scholar]
- Lu, S.-E.; Scholz-Schroeder, B.K.; Gross, D.C. Characterization of the salA, syrF, and syrG regulatory genes located at the right border of the syringomycin gene cluster of Pseudomonas syringae pv. syringae. Mol. Plant-Microbe Interact. 2002, 15, 43–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hollaway, G.J.; Bretag, T.W. The occurrence of Pseudomonas syringae pv. pisi in field pea (Pisum sativum) crops in the Wimmera region of Victoria, Australia. Australas. Plant Pathol. 1995, 24, 133–136. [Google Scholar] [CrossRef]
- Baltrus, D.A.; Dougherty, K.; Beckstrom-Sternberg, S.M.; Beckstrom-Sternberg, J.S.; Foster, J.T. Incongruence between multi-locus sequence analysis (MLSA) and whole-genome-based phylogenies: Pseudomonas syringae pathovar pisi as a cautionary tale. Mol. Plant Pathol. 2014, 15, 461–465. [Google Scholar] [CrossRef]
- Silby, M.W.; Winstanley, C.; Godfrey, S.A.; Levy, S.B.; Jackson, R.W. Pseudomonas genomes: Diverse and adaptable. FEMS Microbiol. Rev. 2011, 35, 652–680. [Google Scholar] [CrossRef] [Green Version]
- Lu, S.-E.; Wang, N.; Wang, J.; Chen, Z.J.; Gross, D.C. Oligonucleotide microarray analysis of the salA regulon controlling phytotoxin production by Pseudomonas syringae pv. syringae. Mol. Plant-Microbe Interact. 2005, 18, 324–333. [Google Scholar] [CrossRef]
- Wilkie, J.P.; Dye, D.W.; Watson, D.R.W. Further hosts of Pseudomonas viridiflava. N. Z. J. Agric. Res. 1973, 16, 315–323. [Google Scholar] [CrossRef]
- Paula Wilkie, J.; Dye, D.W. Pseudomonas cichorii causing tomato and celery diseases in New Zealand. N. Z. J. Agric. Res. 1974, 17, 123–130. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Identity | Collection | Numbers | Year of Collection | Host Plant | Pathovar-Specific PCR | Time to Positive for Psp | Anneal Derivative | Time to Positive for Pss | Anneal Derivative | Reference |
|---|---|---|---|---|---|---|---|---|---|---|
| P. cichorii | VPRI | 42,174 | 1986 | Lactuca sativa (Lettuce) | ND | UD | UD | UD | UD | |
| P. viridiflava | VPRI | 42,172 | 2003 | Pisum sativum (Field Pea) | PEL | UD | UD | UD | UD | |
| P. viridiflava | VPRI | 42,173 | 1987 | Ranunculus sp. (Buttercup) | PEL | UD | UD | UD | UD | [22] |
| Psp Race2 | VPRI | 42,161 | 1944 | Pisum sativum (Field Pea) | Psp | 5.6 | 90.1 | UD | UD | [22] |
| Psp Race2 | VPRI | 42,165 | 1992 | Pisum sativum (Field Pea) | Psp | 7.7 | 90.1 | UD | UD | [23] |
| Psp Race3 | VPRI | 42,162 | 1975 | Pisum sativum (Field Pea) | Psp | 8.0 | 89.9 | UD | UD | [22] |
| Psp Race3 | VPRI | 42,166 | 1992 | Pisum sativum (Field Pea) | Psp | 5.4 | 89.6 | UD | UD | [20] |
| Psp Race4 | VPRI | 42,163 | 1975 | Pisum sativum (Field Pea) | Psp | 4.6 | 89.6 | UD | UD | [22] |
| Psp Race6 | VPRI | 42,167 | 2003 | Pisum sativum (Field Pea) | Psp | 4.9 | 89.9 | UD | UD | [20] |
| Psp Race6 | VPRI | 42,168 | 2001 | Pisum sativum (Field Pea) | Psp | 9.6 | 89.7 | UD | UD | |
| Psp Race6 | VPRI | 42,170 | 1993 | Pisum sativum (Field Pea) | Psp | 8.0 | 89.8 | UD | UD | |
| Psp Race6 | VPRI | 42,171 | 1993 | Pisum sativum (Field Pea | Psp | 9.6 | 89.7 | UD | UD | [23] |
| Psp Race7 | VPRI | 42,164 | NA | Pisum sativum (Field Pea) | Psp | 9.2 | 89.7 | UD | UD | [3] |
| Pss | VPRI | 42,146 | 1981 | Phaseolus vulgaris (French Bean) | Pss | UD | UD | 5.5 | 90.7 | |
| Pss | VPRI | 42,147 | 1978 | Prunus domestica (Plum) | Pss | UD | UD | 3.7 | 90.3 | |
| Pss | VPRI | 22,505 | NA | Prunus domestica (Rosaceae) | Pss | UD | UD | 4.6 | 90.1 | |
| Pss | VPRI | 32,448 | NA | Impatiens sp. [Balsaminaceae] | Pss | UD | UD | 4.8 | 91.7 | |
| Pss | VPRI | 42,148 | 1983 | Pisum sativum (Field Pea) | Pss | UD | UD | 4.8 | 90.5 | [21] |
| Pss | VPRI | 42,149 | 1984 | Pisum sativum (Field Pea) | Pss | UD | UD | 5.7 | 90.7 | [21] |
| Pss | VPRI | 42,150 | 2003 | Pisum sativum (Field Pea) | Pss | UD | UD | 3.8 | 90.7 | |
| Pss | VPRI | 42,143 | 2012 | Phaseolus vulgaris (French Bean) | Pss | UD | UD | 6.7 | 90.7 | [20] |
| Pss | VPRI | 42,151 | 2003 | Pisum sativum (Field Pea) | Pss | UD | UD | 5.6 | 90.7 | [20] |
| Pss | VPRI | 42,607 | 2014 | Pisum sativum (Field Pea) | Pss | UD | UD | 5.6 | 90.6 | |
| Pss | VPRI | 21,225 | 1996 | Prunus persica [Rosaceae] | Pss | UD | UD | 5.7 | 90.5 | |
| Pss | VPRI | 42,827 | 2016 | Vicia faba [Fabaceae] | Pss | UD | UD | 4.6 | 90.6 | |
| Pss | VPRI | 41,526 | 1999 | Brassica oleracea [Brassicaceae] | Pss | UD | UD | 5.6 | 90.7 | |
| Pss | VPRI | 32,662 | 2006 | Grevillea sp. [Proteaceae] | Pss | UD | UD | 8.5 | 90.7 | |
| Pseudomonas marginalis | VPRI | 32,782 | 2006 | Pastinaca sativa [Apiaceae] | ND | UD | UD | UD | UD | |
| Pseudomonas fluorescens | VPRI | 41,564 | 1996 | Lactuca sativa [Asteraceae] | ND | UD | UD | UD | UD | |
| Pseudomonas fluorescens | VPRI | 41,213 | 2007 | Beta vulgaris [Chenopodiaceae] | ND | UD | UD | UD | UD | |
| Pseudomonas corrugata | VPRI | 41,570 | 1997 | Solanum lycopersicum [Solanaceae] | ND | UD | UD | UD | UD | |
| Pseudomonas marginalis | VPRI | 32,683 | 2006 | Hesperis matronalis [Brassicaceae] | ND | UD | UD | UD | UD | |
| Pseudomonas syringae pv. coriandricola | VPRI | 41,569 | 2003 | Coriandrum sativum [Apiaceae] | ND | UD | UD | UD | UD | |
| Pseudomonas putida | VPRI | 32,399 | 2002 | Solanum tuberosum [Solanaceae] | ND | UD | UD | UD | UD | |
| Pseudomonas agarici | DAR | 41,321 | 1982 | Agaricus bisporus (Lange) Imbach | ND | UD | UD | UD | UD | |
| Pseudomonas pickettii | DAR | 65,900 | NA | Clavicipitaceae | ND | UD | UD | UD | UD | |
| Pseudomonas aeruginosa | DAR | 41,354 | 1983 | Clavicipitaceae | ND | UD | UD | UD | UD | |
| Pseudomonas tolaasii | DAR | 35,636 | NA | Clavicipitaceae | ND | UD | UD | UD | UD | |
| Pseudomonas chlororaphis | DAR | 76,122 | 2001 | Brassica napus L. var napus | ND | UD | UD | UD | UD | |
| Xanthomonas campestris | VPRI | 32,436 | NA | NA | ND | UD | UD | UD | UD | |
| Xanthomonas campestris | VPRI | 41,521 | 1997 | Eriostemon sp. | ND | UD | UD | UD | UD | |
| Erwinia chrysanthemi | VPRI | 42,393 | 1998 | Zingiber officinale Roscoe | ND | UD | UD | UD | UD | |
| Erwinia chrysanthemi | VPRI | 42,392 | 1978 | Solanum tuberosum L | ND | UD | UD | UD | UD | |
| Ralstonia solanacearum | VPRI | 42,679 | 2015 | Solanum tuberosum L. | ND | UD | UD | UD | UD | |
| Ralstonia solanacearum | VPRI | 42,677 | 2015 | Solanum tuberosum L | ND | UD | UD | UD | UD | |
| Pss | VPRI | 43,444 | 2007 | Pisum sativum (Field Pea) | Pss | UD | UD | 7.4 | 90.2 | |
| Pss | VPRI | 43,445 | 2007 | Pisum sativum (Field Pea) | Pss | UD | UD | 7.4 | 90.6 | |
| Pss | VPRI | 43,446 | 2007 | Pisum sativum (Field Pea) | Pss | UD | UD | 7.4 | 90.5 | |
| Pss | VPRI | 43,449 | 1990 | Pisum sativum (Field Pea) | Pss | UD | UD | 8.2 | 90.7 | |
| Pss | VPRI | 43,450 | 1983 | Pisum sativum (Field Pea) | Pss | UD | UD | 17.4 | 90.3 | |
| Pss | VPRI | 43,451 | 2007 | Pisum sativum (Field Pea) | Pss | UD | UD | 10.3 | 90.5 | |
| Pss | VPRI | 43,452 | 1989 | Pisum sativum (Field Pea) | Pss | UD | UD | 8.4 | 90.5 | |
| Pss | VPRI | 43,453 | 1989 | Pisum sativum (Field Pea) | Pss | UD | UD | 10.4 | 90.5 | |
| Pss | VPRI | 43,454 | 2007 | Pisum sativum (Field Pea) | Pss | UD | UD | 8.1 | 90.7 | |
| Pss | VPRI | 43,455 | 1989 | Pisum sativum (Field Pea) | Pss | UD | UD | 7.9 | 90.7 | |
| Pss | VPRI | 43,456 | 1989 | Pisum sativum (Field Pea) | Pss | UD | UD | 8.3 | 90.7 | |
| Pss | VPRI | 43,457 | 1989 | Pisum sativum (Field Pea) | Pss | UD | UD | 10.3 | 90.6 | |
| Pss | VPRI | 43,458 | 1989 | Pisum sativum (Field Pea) | Pss | UD | UD | 7.5 | 90.5 | |
| Pss | VPRI | 43460 | 1989 | Phaseolus vulgaris (French Bean) | Pss | UD | UD | 7.5 | 90.4 | |
| Pss | VPRI | 43,461 | 1991 | NA | Pss | UD | UD | 8.0 | 90.5 | |
| Pss | VPRI | 43,462 | 1989 | NA | Pss | UD | UD | 5.7 | 90.4 | |
| Pss | VPRI | 43,463 | 1991 | NA | Pss | UD | UD | 8.8 | 90.5 | |
| Pss | VPRI | 43,464 | 1991 | Pisum sativum (Field Pea) | Pss | UD | UD | 8.1 | 90.3 | |
| Pss | VPRI | 43,465 | 1991 | Pisum sativum (Field Pea) | Pss | UD | UD | 8.6 | 90.5 | |
| Pss | VPRI | 43,466 | 2007 | Pisum sativum (Field Pea) | Pss | UD | UD | 10.4 | 90.5 | |
| Pss | VPRI | 43,467 | 1991 | Pisum sativum (Field Pea) | Pss | UD | UD | 8.6 | 90.4 | |
| Pss | VPRI | 43,498 | 1991 | Pisum sativum (Field Pea) | Pss | UD | UD | 8.4 | 90.5 | |
| Pss | VPRI | 43,482 | 1991 | Pisum sativum (Field Pea) | Pss | UD | UD | 6.8 | 90.4 | |
| Psp | VPRI | 43,468 | 1991 | Pisum sativum (Field Pea) | Psp | 11.4 | 89.8 | UD | UD | |
| Psp | VPRI | 43,475 | 1991 | Pisum sativum (Field Pea) | Psp | 5.6 | 89.7 | UD | UD | |
| Psp | VPRI | 43,476 | 1991 | Pisum sativum (Field Pea) | Psp | 6.5 | 89.7 | UD | UD | |
| Psp | VPRI | 43,477 | 2007 | Pisum sativum (Field Pea) | Psp | 5.8 | 89.9 | UD | UD | |
| Psp | VPRI | 43,478 | 1991 | Pisum sativum (Field Pea) | Psp | 6.0 | 89.9 | UD | UD | |
| Psp | VPRI | 43,479 | 1991 | Pisum sativum (Field Pea) | Psp | 6.5 | 89.6 | UD | UD | |
| Psp | VPRI | 43,480 | 1991 | Pisum sativum (Field Pea) | Psp | 6.2 | 90.1 | UD | UD | |
| Psp | VPRI | 43,481 | 1991 | Pisum sativum (Field Pea) | Psp | 6.2 | 90.1 | UD | UD | |
| Psp | VPRI | 43,483 | 1991 | Pisum sativum (Field Pea) | Psp | 11.3 | 90.1 | UD | UD | |
| Psp | VPRI | 43,484 | 1991 | Pisum sativum (Field Pea) | Psp | 6.1 | 90.1 | UD | UD | |
| Psp | VPRI | 43,485 | 1991 | Pisum sativum (Field Pea) | Psp | 6.3 | 89.9 | UD | UD | |
| Psp | VPRI | 43,486 | 1991 | Pisum sativum (Field Pea) | Psp | 6.3 | 89.6 | UD | UD | |
| Psp | VPRI | 43,487 | 1991 | Pisum sativum (Field Pea) | Psp | 6.4 | 89.6 | UD | UD | |
| Psp | VPRI | 43,488 | 1991 | Pisum sativum (Field Pea) | Psp | 7.5 | 89.9 | UD | UD | |
| Psp | VPRI | 43,489 | 1991 | Pisum sativum (Field Pea) | Psp | 6.1 | 89.7 | UD | UD | |
| Psp | VPRI | 43,490 | 1991 | Pisum sativum (Field Pea) | Psp | 5.7 | 89.8 | UD | UD | |
| Psp | VPRI | 43,447 | 2007 | Pisum sativum (Field Pea) | Psp | 5.7 | 89.7 | UD | UD | |
| Psp | VPRI | 43,448 | 1983 | Pisum sativum (Field Pea) | Psp | 5.7 | 89.8 | UD | UD | |
| Psp | VPRI | 43,491 | 1991 | Pisum sativum (Field Pea) | Psp | 7.9 | 89.7 | UD | UD | |
| Psp | VPRI | 43,492 | 1991 | Pisum sativum (Field Pea) | Psp | 7.4 | 90.1 | UD | UD | |
| Psp | VPRI | 43,493 | 1991 | Pisum sativum (Field Pea) | Psp | 7.3 | 90.1 | UD | UD | |
| Psp | VPRI | 43,494 | 1991 | Pisum sativum (Field Pea) | Psp | 12.9 | 89.8 | UD | UD | |
| Psp | VPRI | 43,495 | 2007 | Pisum sativum (Field Pea) | Psp | 6.7 | 90.1 | UD | UD | |
| Psp | VPRI | 43,496 | 1991 | Pisum sativum (Field Pea) | Psp | 8.5 | 90.0 | UD | UD | |
| Psp | VPRI | 43,497 | 1991 | Pisum sativum (Field Pea) | Psp | 7.5 | 89.9 | UD | UD | |
| Psp | VPRI | 43,499 | 1991 | Pisum sativum (Field Pea) | Psp | 8.5 | 89.9 | UD | UD | |
| Psp | VPRI | 43,500 | 1991 | Pisum sativum (Field Pea) | Psp | 7.3 | 89.8 | UD | UD |
| Primers | Sequence (5′ to 3′) | Nucleotide Start Position | Nucleotide End Position |
|---|---|---|---|
| Pseudomonas syringae pathovar pisi sequence submitted to GenBank accession no. MZ043759 | |||
| Psp_F3 | CTCGTGCACGATTCACCAT | 198 | 219 |
| Psp_B3 | CGCCTTACTCATTAGTACGCTAA | 426 | 488 |
| Psp_FIP * | (F1c) CGTCTACAGCTAATCATGCCTCT | 278 | 300 |
| (F2) AGTCTCCTACCTCTATCCGTCAT | 227 | 248 | |
| Psp_BIP * | (B1c) TAGCGCTTTCGCACAGCCT | 316 | 334 |
| (B2) AGGCATCTTGCTCCACGCAT | 393 | 422 | |
| Psp_LF | TGCTCCACGTGGGTTCACAG | 253 | 272 |
| Psp_LB | GAGGCCATGAGTGGGCTGAGA | 363 | 383 |
| Pseudomonas syringae pathovar syringae to GenBank accession no. MZ043758 | |||
| Pss_F3 | CATCCTTGACCTCCGAAGCG | 148 | 167 |
| Pss_B3 | CAAGCCATCGAGGCCAATGG | 401 | 420 |
| Pss_FIP * | (F1c) CACCGCGCCGTAGTTGAACCT | 262 | 282 |
| (F2) ATGCGCAACGGTTGATGGGTA | 173 | 193 | |
| Pss_BIP * | (B1c) TACCTTGCCGATGTGCTGCGCA | 310 | 331 |
| (B2) ATTGCAACGGTAGTCGCCA | 381 | 399 | |
| Pss_LF | ATCAGCCCTGTACGACTCGCTC | 210 | 231 |
| Pss_LB | GCATCGGCAAATTTGAATGTCC | 359 | 380 |
| Inoculated | Controls | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| DAI 2 | DAI 3 | DAI 4 | DAI 5 | DAI 6 | DAI 7 | DAI 8 | DAI 9 | DAI 10 | Surfactant | Un-Inoculated | |
| Pseudomonas syringae pathovar pisi | |||||||||||
| Tp (min) | *UD | 27.32 ± 3.1 | 21.61 ± 2.2 | 19.84 ± 3.7 | 17.00 ± 2.5 | 14.54 ± 3.9 | 13.97 ± 2.0 | 12.07 ± 1.5 | 9.55 ± 2.1 | UD | UD |
| AD (°C) | - | 90.0 | 90.2 | 89.5 | 90.1 | 90.1 | 90.0 | 90.0 | 90.0 | - | - |
| Pseudomonas syringae pathovar syringae | |||||||||||
| Tp (min) | 21.75 ± 5.1 | 19.79 ± 3.2 | 19.22 ± 2.7 | 17.28 ± 2.1 | 15.60 ± 3.1 | 15.19 ± 2.2 | 12.57 ± 3.8 | 12.19 ± 2.8 | 7.035 ± 1.1 | UD | UD |
| AD (°C) | 90.1 | 90.5 | 90.0 | 90.3 | 90.2 | 90.0 | 90.3 | 90.0 | 90.1 | - | - |
| Year of Release | Field Pea Variety | Psp LAMP | Pss LAMP Tp (min) | Bacteria Isolated | Pathovar Specific PCR |
|---|---|---|---|---|---|
| 1985 | Wirrega | UD | 7.5 | Yes | Pss |
| 1996 | Bohatyar | UD | 11.4 | Yes | Pss |
| 1993 | Jupiter | UD | 8.5 | Yes | Pss |
| 2000 | Mukta | UD | 9.4 | Yes | Pss |
| 2010 | PBA Twilight | UD | 8.6 | Yes | Pss |
| 2003 | Moonlight | UD | 12.5 | Yes | Pss |
| 1998 | Magnet | UD | 15.6 | Yes | Pss |
| 2003 | Sturt | UD | 10.5 | Yes | Pss |
| 1985 | Alma | UD | 8.9 | Yes | Pss |
| 1992 | Bonzer | UD | 11.5 | Yes | Pss |
| 2000 | Cooke | UD | 12.8 | Yes | Pss |
| 1979 | Whero | UD | 17.8 | Yes | Pss |
| 2000 | Snowpeak | UD | 19.5 | Yes | Pss |
| 1995 | Laura | UD | 14.7 | Yes | Pss |
| 2002 | Dunwa | UD | 18.9 | Yes | Pss |
| 2000 | Excel | UD | 16.9 | Yes | Pss |
| 1964 | Derrimut | UD | 19.0 | Yes | Pss |
| 1920 | White Brunswick | UD | 20.1 | Yes | Pss |
| 2000 | Soupa | UD | 13.8 | Yes | Pss |
| 1998 | Parafield | UD | 17.4 | Yes | Pss |
| 1985 | Glenroy | UD | 8.0 | Yes | Pss |
| 2002 | Kaspa | UD | 5.8 | Yes | Pss |
| 2010 | PBA Oura | UD | 19.3 | Yes | Pss |
| 1893 | Dun | UD | 11.3 | Yes | Pss |
| 2014 | PBA Pearl | UD | 22.6 | Yes | Pss |
| 2011 | PBA Percy | UD | 12.5 | Yes | Pss |
| 1989 | Diakum | UD | 14.2 | Yes | Pss |
| 1999 | Helena | UD | 17.8 | Yes | Pss |
| 2013 | PBA Wharton | UD | 10.9 | Yes | Pss |
| 2000 | Paravic | UD | 11.8 | Yes | Pss |
| 1998 | King | UD | 8.9 | Yes | Pss |
| 2000 | Santi | UD | 5.4 | Yes | Pss |
| 1970 | Dundale | UD | 6.7 | Yes | Pss |
| 1992 | Bluey | UD | 5.7 | Yes | Pss |
| 2000 | Yarrum | UD | 12.8 | Yes | Pss |
| 1939 | Colligian | UD | 17.8 | Yes | Pss |
| 2002 | Kiley | UD | 12.5 | Yes | Pss |
| 1986 | Maitland | UD | 14.6 | Yes | Pss |
| 2011 | Maki | UD | 12.6 | Yes | Pss |
| 1998 | Morgan | UD | 13.5 | Yes | Pss |
| 2002 | Kaspa | UD | 6.5 | Yes | Pss |
| 2014 | PBA Pearl | UD | 19.8 | Yes | Pss |
| 2017 | PBA Butler | UD | 18.0 | Yes | Pss |
| 2010 | PBA Ganyah | UD | 5.6 | Yes | Pss |
| 2011 | PBA Percy | UD | 16.7 | Yes | Pss |
| 2002 | Kaspa | UD | 7.8 | Yes | Pss |
| 1939 | Collegian | UD | 10.3 | Yes | Pss |
| 1976 | Cressy Blue | UD | 10.7 | Yes | Pss |
| 2010 | PBA Oura | UD | 15.6 | yes | Pss |
| 2017 | PBA Butler | UD | 17.1 | Yes | Pss |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kant, P.; Fruzangohar, M.; Mann, R.; Rodoni, B.; Hollaway, G.; Rosewarne, G. Development and Application of a Loop-Mediated Isothermal Amplification (LAMP) Assay for the Detection of Pseudomonas syringae Pathovars pisi and syringae. Agriculture 2021, 11, 875. https://doi.org/10.3390/agriculture11090875
Kant P, Fruzangohar M, Mann R, Rodoni B, Hollaway G, Rosewarne G. Development and Application of a Loop-Mediated Isothermal Amplification (LAMP) Assay for the Detection of Pseudomonas syringae Pathovars pisi and syringae. Agriculture. 2021; 11(9):875. https://doi.org/10.3390/agriculture11090875
Chicago/Turabian StyleKant, Pragya, Mario Fruzangohar, Rachel Mann, Brendan Rodoni, Grant Hollaway, and Garry Rosewarne. 2021. "Development and Application of a Loop-Mediated Isothermal Amplification (LAMP) Assay for the Detection of Pseudomonas syringae Pathovars pisi and syringae" Agriculture 11, no. 9: 875. https://doi.org/10.3390/agriculture11090875