In-Vitro Plant Growth Promotion of Rhizobium Strains Isolated from Lentil Root Nodules under Abiotic Stresses

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Growth
2.2. Measurement of Phosphate Solubilization Activity
2.2.1. Qualitative Test
2.2.2. Quantitative Test
2.3. Measurement of Phytohormones Production
2.3.1. IAA (Indole Acetic Acid) Detection
Qualitative Test
Quantitative Test
2.3.2. Gibberellic Acid Detection
2.4. Measurement of Siderophore Production
2.5. Assessment of Bacterial Growth, Phosphate Solubilization Activity, and IAA (Indole Acetic Acid) Production under Abiotic Stress
2.6. Statistical Analysis and Graphic Presentation
3. Results
3.1. Qualitative Test
3.2. Plant Growth-Promoting Traits of Rhizobium Strains
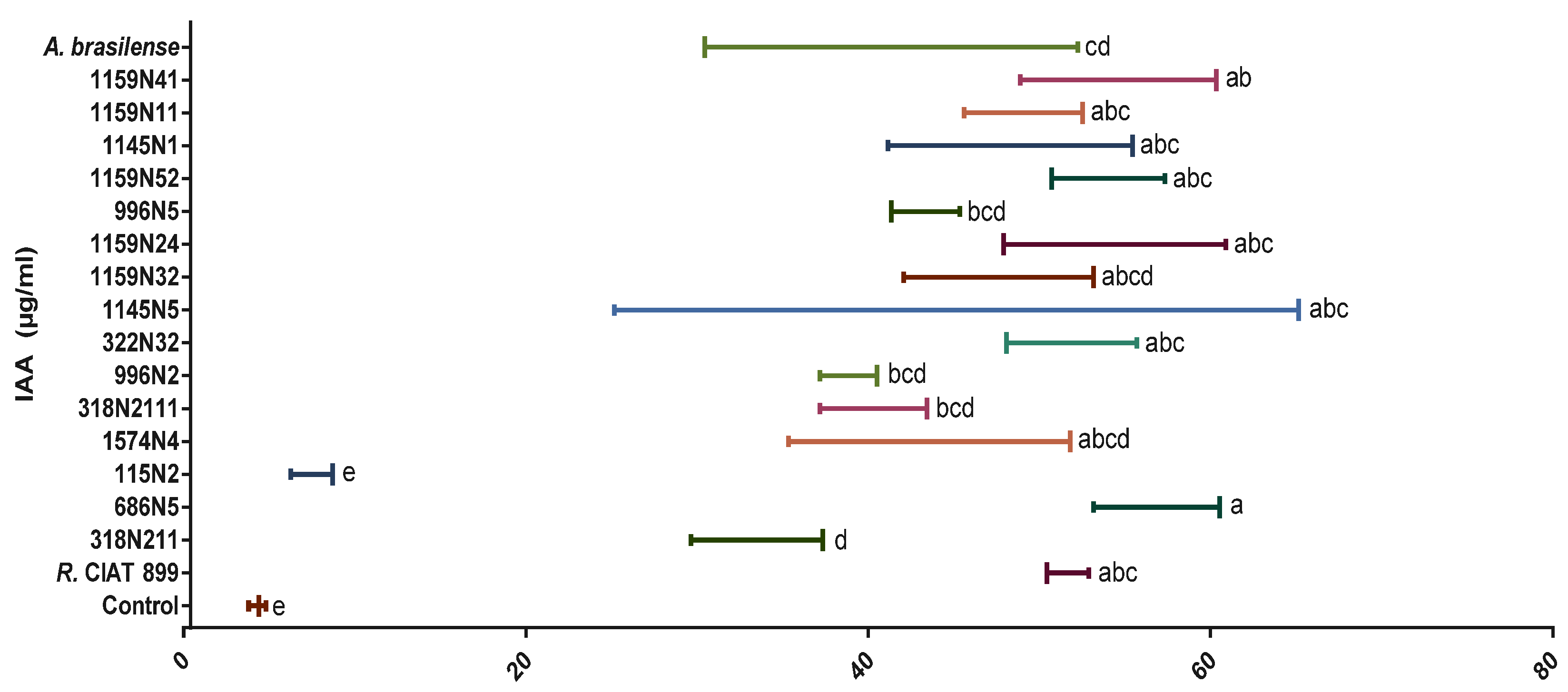
Phosphate Solubilization Activity and Phytohormones Production
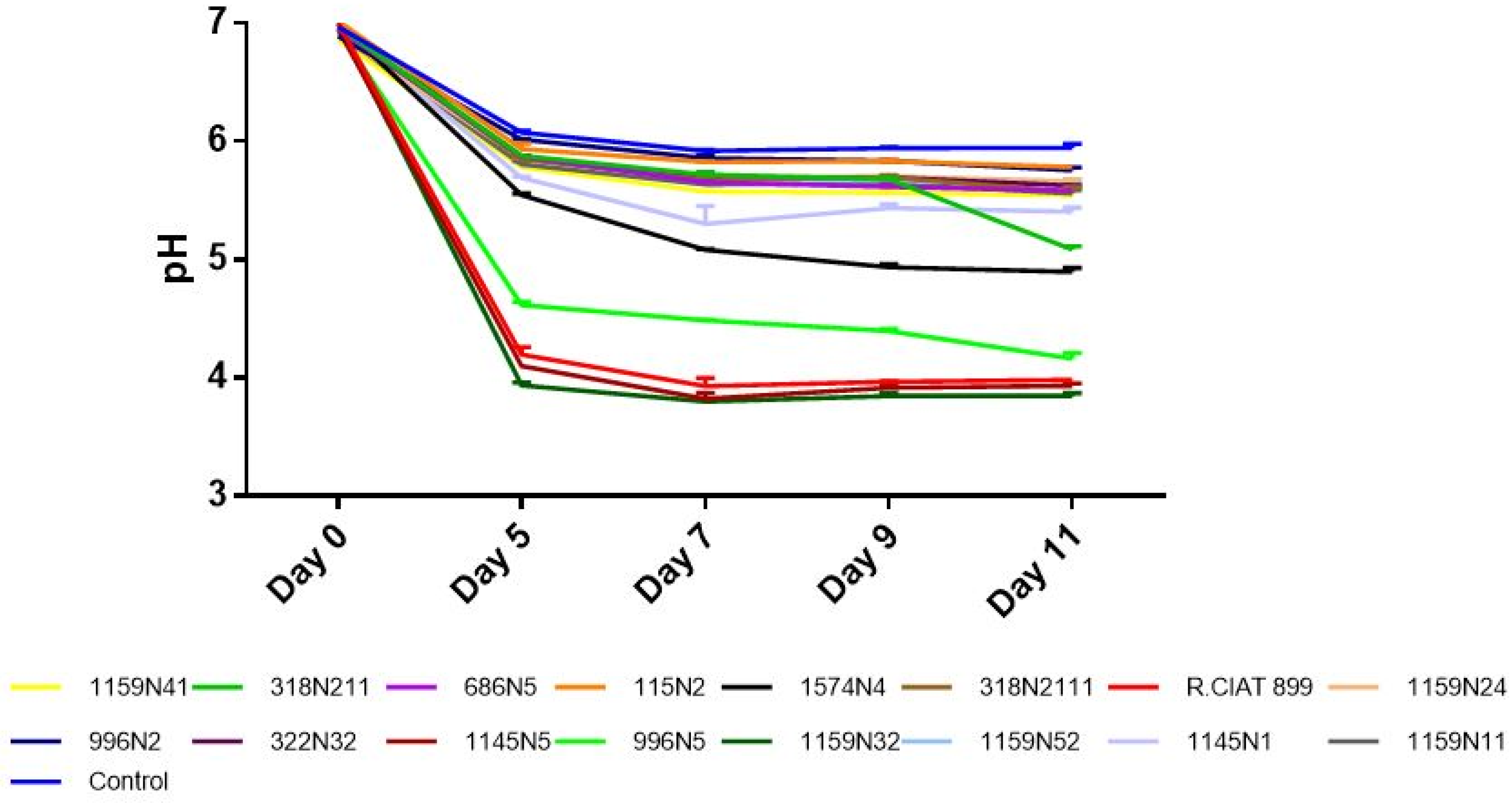
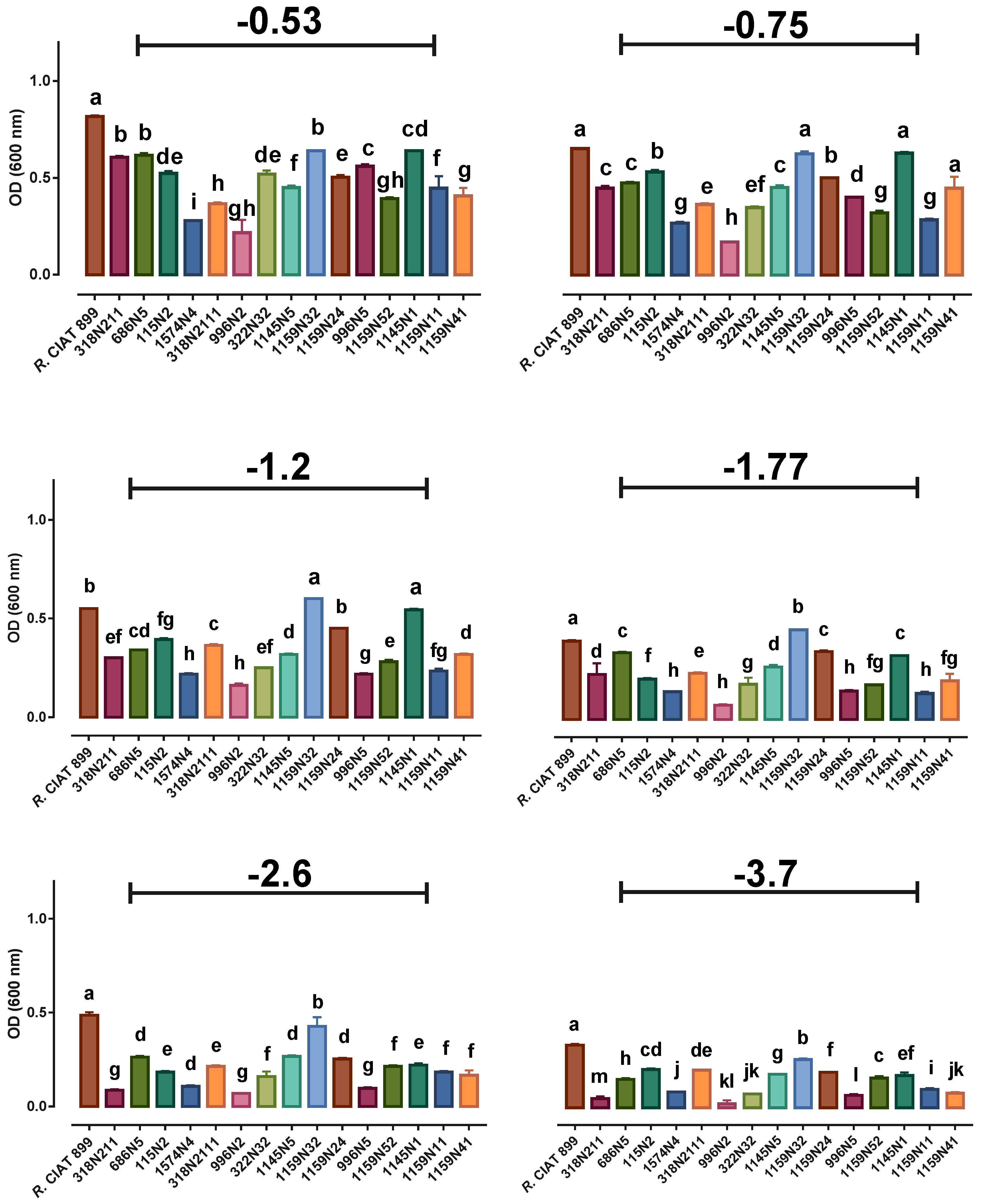
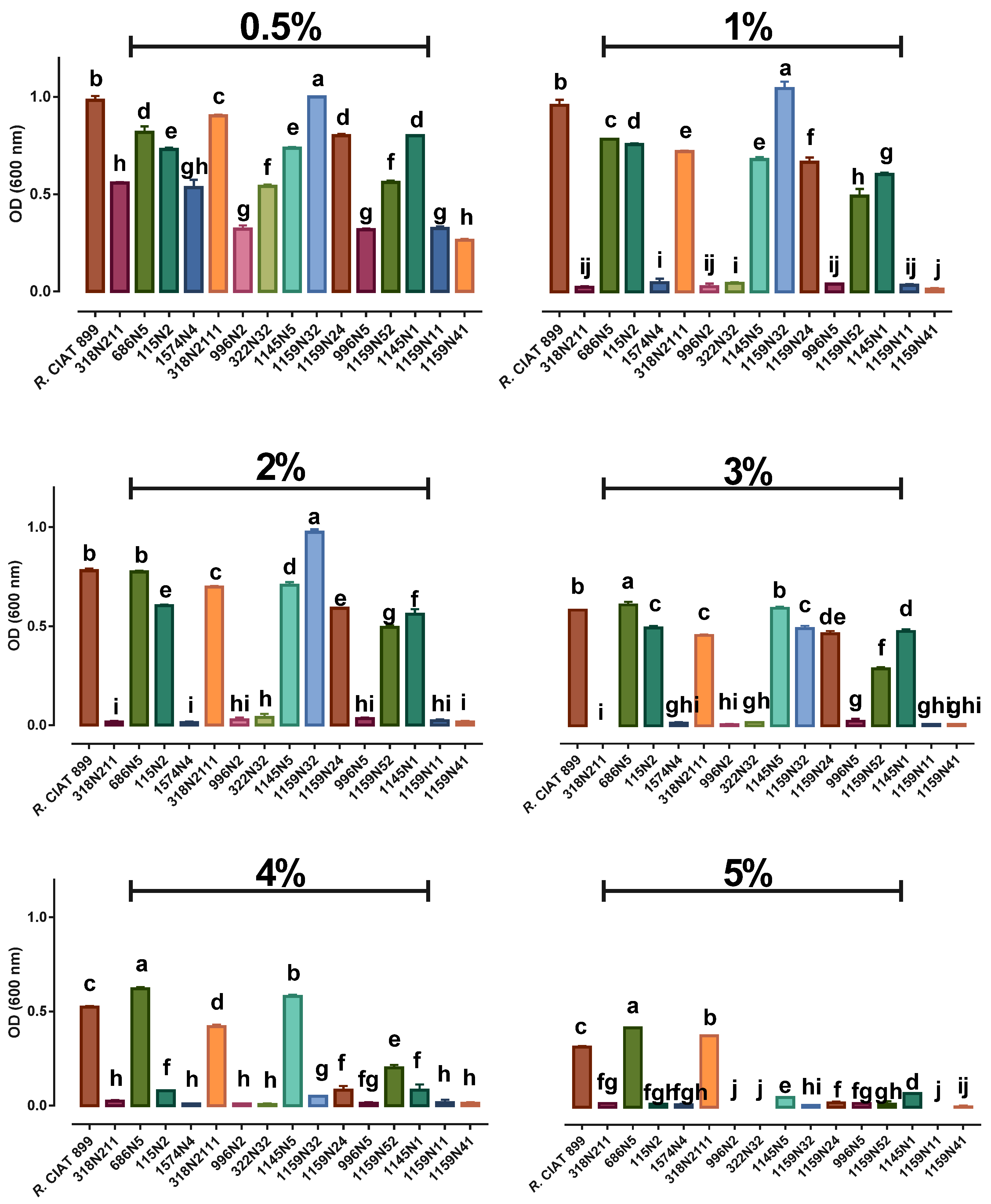
3.3. Rhizobium Growth under Abiotic Stresses
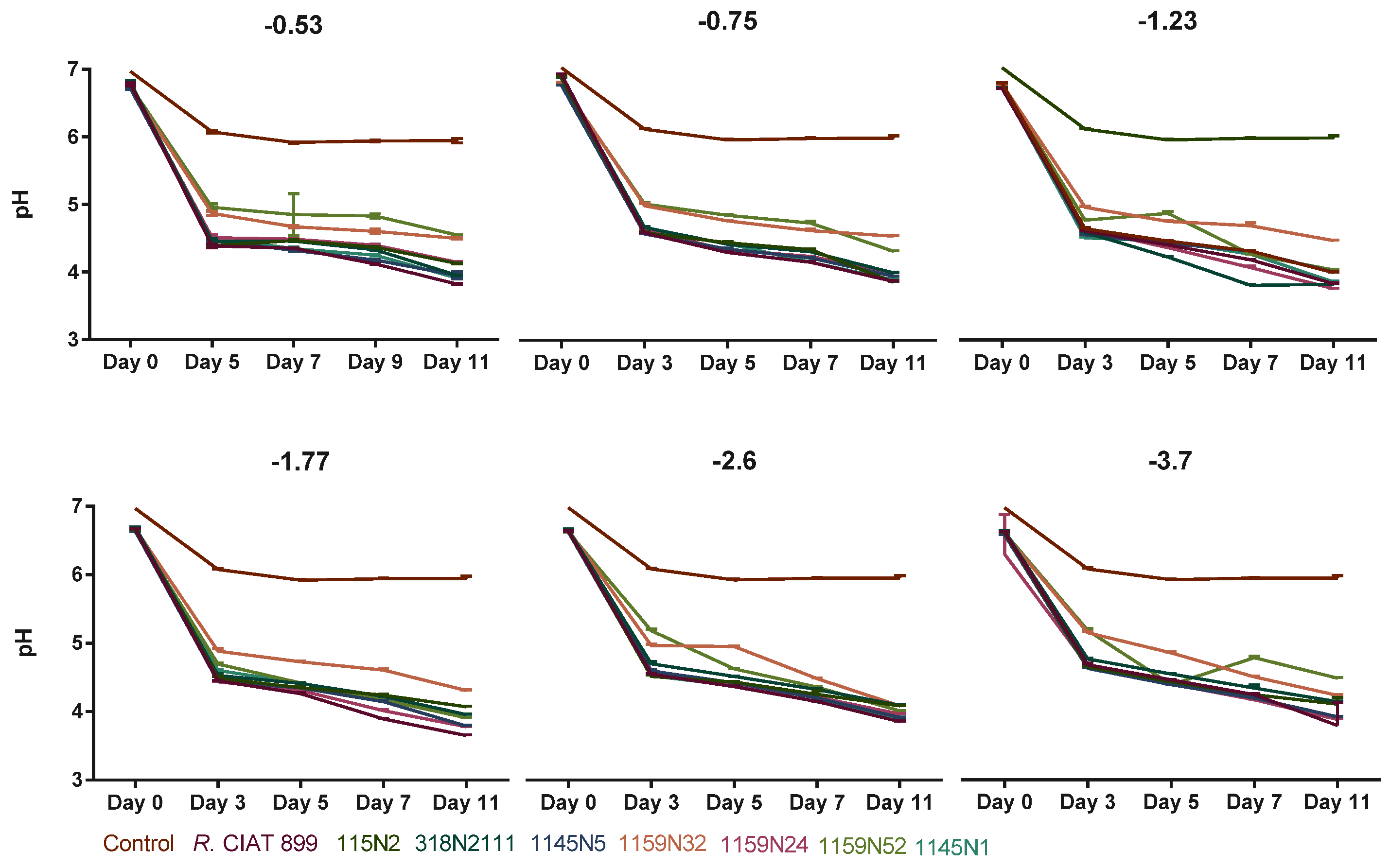
3.3.1. Rhizobium Growth under Drought Stress
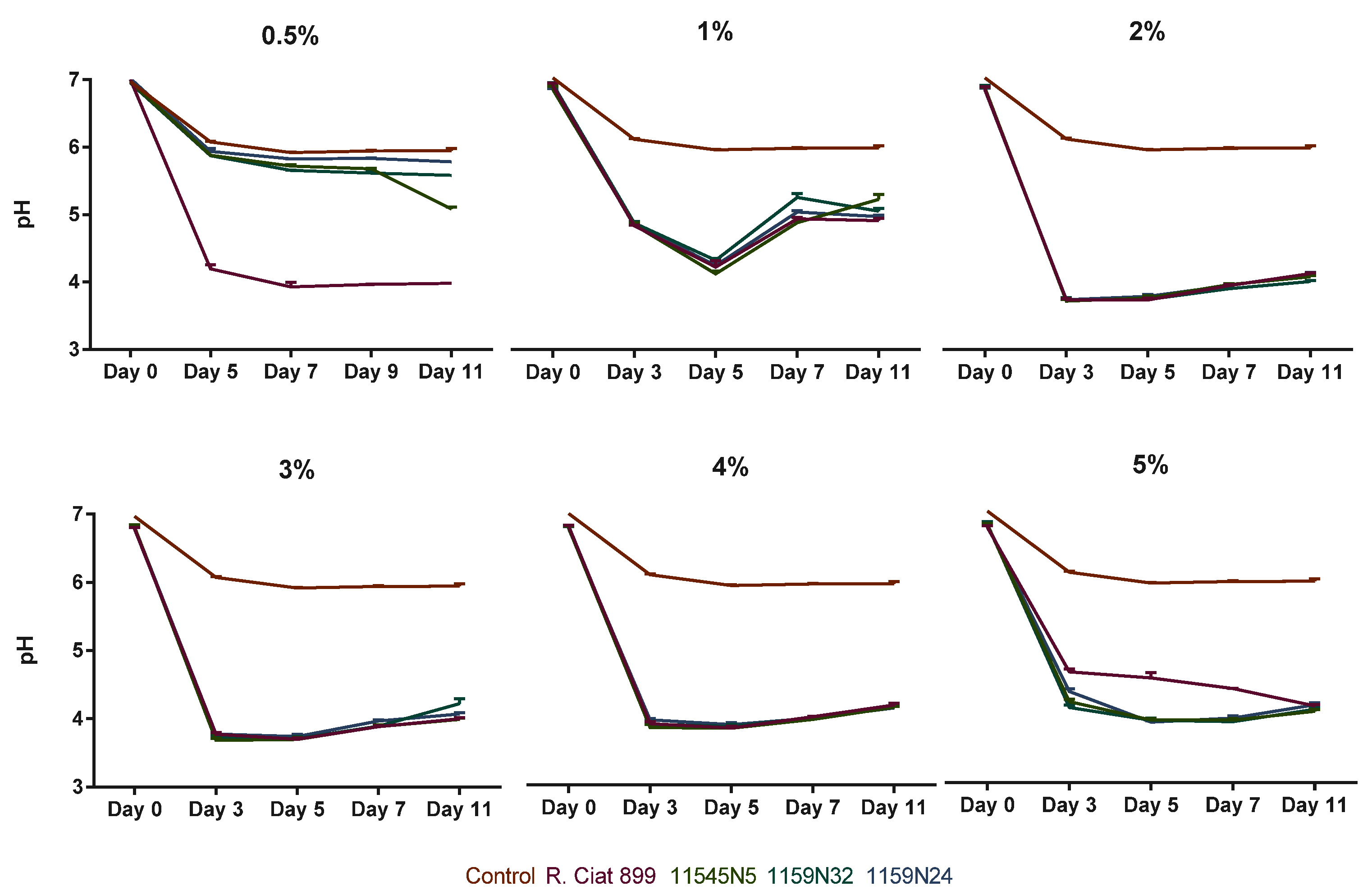
3.3.2. Rhizobium Growth under Salt Stress
3.4. Plant Growth-Promoting Activity under Abiotic Stresses
3.4.1. Phosphate Solubilization Activity under Drought Stress
3.4.2. Phosphate Solubilization Activity under Salt Stress
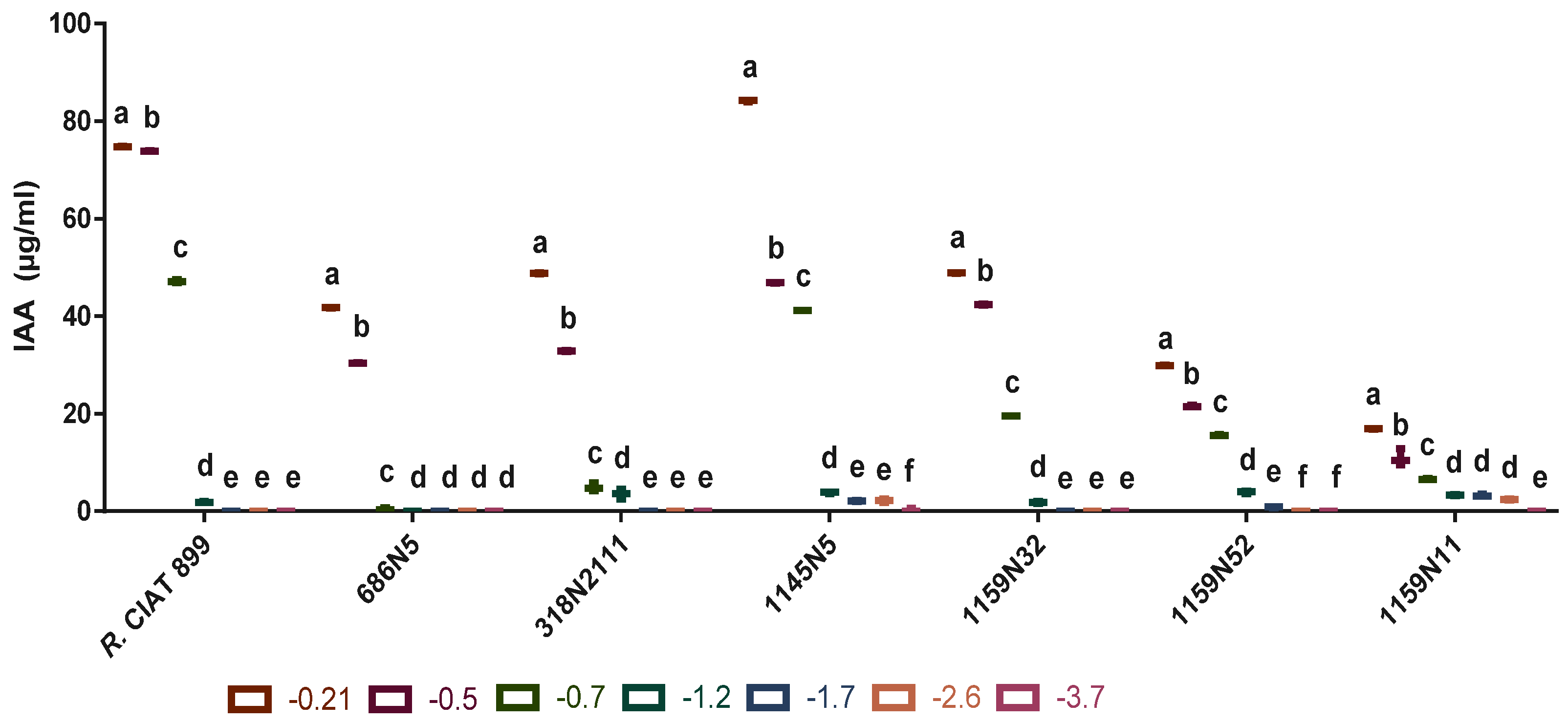
3.4.3. Indole Acetic Acid (IAA) under Drought Stress
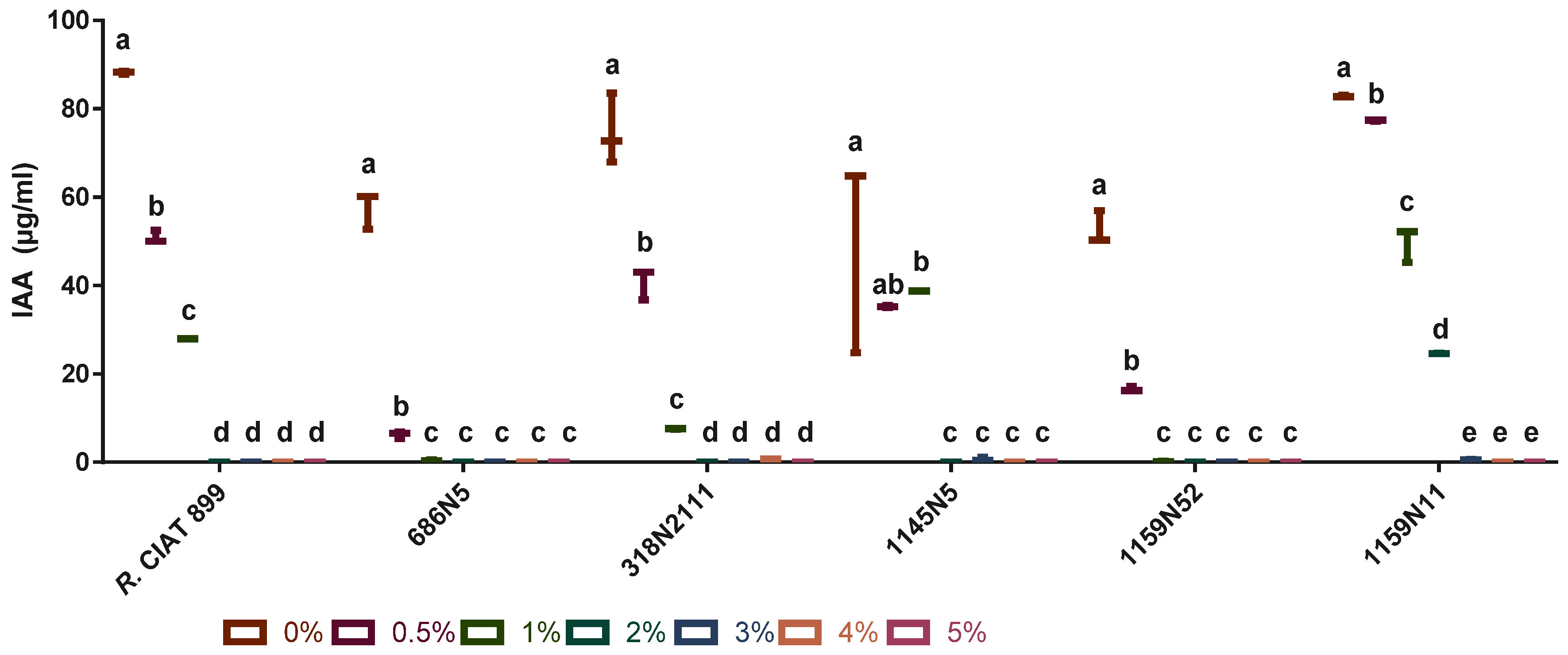
3.4.4. Indole Acetic Acid (IAA) under Salt Stress
4. Discussions
4.1. Potential of Rhizobium as Plant Growth Promoters
4.2. Effect of Abiotic Stresses on Rhizobium Growth
4.3. Effect of Abiotic Stresses on Rhizobium PGP Activities
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kavamura, V.; Santos, S.N.; da Silva, L.; Parma, M.; Ávila, A.; Visconti, A.; de Melo, I. Screening of Brazilian cacti rhizobacteria for plant growth promotion under drought. Microbiol. Res. 2013, 168, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Egamberdieva, D. The Role of Phytohormone Producing Bacteria in Alleviating Salt Stress in Crop Plants. Biotechnological Techniques of Stress Tolerance in Plants 2013; Stadium Press LLC: Houston, TX, USA, 2013; pp. 21–39. [Google Scholar]
- Vurukonda, S.; Vardharajula, S.; Shrivastava, M.; SkZ, A. Enhancement of drought stress tolerance in crops by plant growth promoting rhizobacteria. Microbiol. Res. 2016, 184, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Kloepper, J.; Ryu, C. Rhizosphere bacteria help plants tolerate abiotic stress. Trends Plant Sci. 2009, 14, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Meena, K.; Sorty, A.; Bitla, U.; Choudhary, K.; Gupta, P.; Pareek, A.; Singh, H. Abiotic stress responses and microbe-mediated mitigation in plants: The omics strategies. Front. Plant Sci. 2017, 8, 172. [Google Scholar] [CrossRef]
- Ullah, A.; Sun, H.; Yang, X.; Zhang, X. Drought coping strategies in cotton: Increased crop per drop. Plant Biotechnol. J. 2017, 15, 271–284. [Google Scholar] [CrossRef] [PubMed]
- Michalak. A. Phenolic compounds and their antioxidant activity in plants growing under heavy metal stress. Pol. J. Environ. Stud. 2006, 15, 4. [Google Scholar]
- Jouyban, Z. The effects of salt stress on plant growth. Tech. J. Eng. Appl. Sci. 2012, 2, 7–10. [Google Scholar]
- Egamberdieva, D.; Wirth, S.; Abd_Allah, E. Plant hormones as key regulators in plant-microbe interactions under salt stress. In Plant Microbiome: Stress Response; Springer: Singapore, 2018; pp. 165–182. [Google Scholar]
- Kamilova, F.; Kravchenko, L.V.; Shaposhnikov, A.I.; Azarova, T.; Makarova, N.; Lugtenberg, B. Organic acids, sugars, and L-tryptophane in exudates of vegetables growing on stonewool and their effects on activities of rhizosphere bacteria. Mol. Plant Microbe Interact 2006, 19, 250–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, S.B.; Sayyed, R.Z.; Trivedi, M.H.; Gobi, A.T. Phosphate solubilizing microbes: Sustainable approach for managing phosphorus deficiency in agricultural soils. Springerplus 2013, 2, 587. [Google Scholar] [CrossRef] [Green Version]
- Deshwal, V.K.; Dubey, R.C.; Maheshwari, D.K. Isolation of plant growth promoting strains of Bradyrhizobium (Arachis) sp. with biocontrol potential against Macrophomina phaseolina causing charcoal rot of peanut. Curr. Sci. 2003, 84, 443–444. [Google Scholar]
- Ahemad, M.; Khan, M.S. Comparative toxicity of selected insecticides to pea plants and growth promotion in response to insecticide-tolerant and plant growth promoting Rhizobium leguminosarum. Crop. Prot. 2010, 29, 325–329. [Google Scholar] [CrossRef]
- Das, K.; Prasanna, R.; Saxena, A.K. Rhizobia: A potential biocontrol agent for soilborne fungal pathogens. Folia Microbiol. 2017, 62, 425–435. [Google Scholar] [CrossRef] [PubMed]
- Kloepper, J.; Ryu, C.; Zhang, S. Induced systemic resistance and promotion of plant growth by Bacillus spp. Phytopathology 2004, 94, 1259–1266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swaine, E.; Swaine, M.; Killham, K. Effects of drought on isolates of Bradyrhizobium elkanii cultured from Albizia adianthifolia seedlings of different provenances. Agrofor. Syst. 2007, 69, 135–145. [Google Scholar] [CrossRef]
- Murray, R.; Holt, J. The History of Bergey’s Manual. In Bergey’s Manual® of Systematic Bacteriology; Springer: Boston, MA, USA, 2005; pp. 1–14. [Google Scholar]
- Serraj, R.; Sinclair, T. Osmolyte accumulation: Can it really help increase crop yield under drought conditions? Plant Cell Environ. 2002, 25, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Ngumbi, E.; Kloepper, J. Bacterial-mediated drought tolerance: Current and future prospects. Appl. Soil Ecol. 2016, 105, 109–125. [Google Scholar] [CrossRef]
- Mohammadipanah, F.; Zamanzadeh, M. Bacterial Mechanisms Promoting the Tolerance to Drought Stress in Plants. In Secondary Metabolites of Plant Growth Promoting Rhizomicroorganisms; Springer: Singapore, 2019; pp. 185–224. [Google Scholar]
- Figueiredo, M.V.; Burity, H.A.; Martínez, C.R.; Chanway, C.P. Alleviation of drought stress in the common bean (Phaseolus vulgaris L.) by co-inoculation with Paenibacillus polymyxa and Rhizobium tropici. Appl. Soil Ecol. 2008, 40, 182–188. [Google Scholar] [CrossRef]
- Fukami, J.; Cerezini, P.; Hungria, M. Azospirillum: Benefits that go far beyond biological nitrogen fixation. AMB Express 2018, 8, 73. [Google Scholar] [CrossRef] [PubMed]
- Harley, J.P.; Prescott, L.M. Laboratory Exercises in Microbiology; WCB/McGraw-Hill: Boston, MA, USA, 1999. [Google Scholar]
- Vincent, J.M. A Manual for the Practical Study of the Root-Nodule Bacteria; Blackwell Scientific: Oxford, UK, 1970. [Google Scholar]
- Leonard, W.H. Ten years of research on investigative laboratory instruction strategies. J. Coll. Sci. Teach. 1989, 18, 304–306. [Google Scholar]
- Paul, M.; Rao, W. Phosphate-dissolving bacteria in the rhizosphere of some cultivated legumes. Plant Soil 1971, 35, 127–132. [Google Scholar] [CrossRef]
- Kothamasi, D.; Kothamasi, S.; Bhattacharyya, A.; Kuhad, R.; Babu, C. Arbuscular mycorrhizae and phosphate solubilising bacteria of the rhizosphere of the mangrove ecosystem of Great Nicobar island, India. Biol. Fertil. Soils 2006, 42, 358–361. [Google Scholar] [CrossRef]
- Barton, C.J. Photometric analysis of phosphate rock. Anal. Chem. 1948, 20, 1068–1073. [Google Scholar] [CrossRef]
- Gordon, S.; Weber, R. Colorimetric estimation of indoleacetic acid. Plant Physiol. 1951, 26, 192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berríos, J.; Illanes, A.; Aroca, G. Spectrophotometric method for determining gibberellic acid in fermentation broths. Biotechnol. Lett. 2004, 26, 67–70. [Google Scholar] [CrossRef] [PubMed]
- Pandya, N.D.; Desai, P.V. Screening and characterization of GA3 producing Pseudomonas monteilii and its impact on plant growth promotion. Int. J. Curr. Microbiol. App. Sci. 2014, 3, 110–115. [Google Scholar]
- Patel, K.; Goswami, D.; Dhandhukia, P.; Thakker, J. Techniques to study microbial phytohormones. In Bacterial Metabolites in Sustainable Agroecosystem; Springer: Cham, Switzerland, 2015; pp. 1–27. [Google Scholar]
- Vikram, A.; Hamzehzarghani, H.; Alagawadi, A.R.; Krishnaraj, P.U.; Chandrashekar, B.S. Production of plant growth promoting substances by phosphate solubilizing bacteria isolated from vertisols. J. Plant Sci. 2007, 2, 326–333. [Google Scholar]
- Schwyn, B.; Neilands, J.B. Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef]
- Michel, B.E.; Kaufmann, M.R. The osmotic potential of polyethylene glycol 6000. Plant Physiol. 1973, 51, 914–916. [Google Scholar] [CrossRef]
- Guggenheim, E.A. The theoretical basis of Raoult’s law. Trans. Faraday Soc. 1937, 33, 151–156. [Google Scholar] [CrossRef]
- Gopalakrishnan, S.; Sathya, A.; Vijayabharathi, R.; Varshney, R.; Gowda, C.; Krishnamurthy, L. Plant growth promoting rhizobia: Challenges and opportunities. 3 Biotech 2015, 5, 355–377. [Google Scholar] [CrossRef] [Green Version]
- Zou, X.; Binkley, D.; Doxtader, K. A new method for estimating gross phosphorus mineralization and immobilization rates in soils. Plant Soil 1992, 147, 243–250. [Google Scholar] [CrossRef]
- Afzal, A.; Bano, A. Rhizobium and phosphate solubilizing bacteria improve the yield and phosphorus uptake in wheat (Triticum aestivum). Int. J. Agric. Biol. 2008, 10, 85–88. [Google Scholar]
- Sharma, P.; Padh, H.; Shrivastava, N. Hairy root cultures: A suitable biological system for studying secondary metabolic pathways in plants. Eng. Life Sci. 2013, 13, 62–75. [Google Scholar] [CrossRef]
- Saghafi, D.; Ghorbanpour, M.; Lajayer, B. Efficiency of Rhizobium strains as plant growth promoting rhizobacteria on morpho-physiological properties of Brassica napus L. under salinity stress. J. Soil Sci. Plant Nutr. 2018, 18, 253–268. [Google Scholar] [CrossRef] [Green Version]
- Alikhani, H.A.; Saleh-Rastin, N.; Antoun, H. Phosphate solubilization activity of rhizobia native to Iranian soils. In First International Meeting on Microbial Phosphate Solubilization; Springer: Dordrecht, The Netherlands, 2007; pp. 35–41. [Google Scholar]
- Sridevi, M.; Mallaiah, K. Phosphate solubilization by Rhizobium strains. Indian J. Microbiol. 2009, 49, 98–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leyval, C.; Barthelin, J. Interactions between Laccaria laccata, Agrobacterium radiobacter and beech roots: Influence on P, K, Mg and Fe mobilization from mineral and plant growth. Plant Soil 1989, 17, 103–110. [Google Scholar] [CrossRef]
- Louw, H.A.; Webley, D.M. A study of soil bacteria dissolving certain phosphate fertilizers and related compounds. J. Appl. Bacteriol. 1959, 22, 227–233. [Google Scholar] [CrossRef]
- Gupta, R.; Singal, R.; Shankar, A.; Kuhad, R.C.; Saxena, R.K. A modified plate assay for screening phosphate solubilizing microorganisms. J. Gen. Appl. Microbiol. 1994, 40, 255–260. [Google Scholar] [CrossRef]
- Nautiyal, C.S. An efficient microbiological growth medium for screening phosphate solubilizing microorganisms. FEMS Microbiol. Lett. 1999, 170, 265–270. [Google Scholar] [CrossRef]
- Baig, K.S.; Arshad, M.; Zahir, Z.A.; Cheema, M.A. Comparative efficacy of qualitative and quantitative methods for rock phosphate solubilization with phosphate solubilizing rhizobacteria. Soil Environ. 2010, 29, 82–86. [Google Scholar]
- Saghafi, D.; Ghorbanpour, M.; Ajirloo, H.; Lajayer, A. Enhancement of growth and salt tolerance in Brassica napus L. seedlings by halotolerant Rhizobium strains containing ACC-deaminase activity. Plant Physiol. Rep. 2019, 24, 1–11. [Google Scholar] [CrossRef]
- Kadmiri, I.; Chaouqui, L.; Azaroual, S.; Sijilmassi, B.; Yaakoubi, K.; Wahby, I. Phosphate-solubilizing and auxin-producing rhizobacteria promote plant growth under saline conditions. Arab. J. Sci. Eng. 2018, 43, 3403–3415. [Google Scholar] [CrossRef]
- Pandya, N.; Desai, P. Gibberellic Acid Production by Bacillus cereus Isolated from the Rhizosphere of Sugarcane. J. Pure Appl. Microbiol. 2013, 7, 3239–3242. [Google Scholar]
- Shrivastava, P.; Kumar, R. Soil salinity: A serious environmental issue and plant growth promoting bacteria as one of the tools for its alleviation. Saudi J. Biol. Sci. 2015, 22, 123–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, S.; Khan, A.; Waqas, M.; You, Y.; Kim, J.; Lee, J. Plant growth-promoting rhizobacteria reduce adverse effects of salinity and osmotic stress by regulating phytohormones and antioxidants in Cucumis sativus. J. Plant Interact. 2014, 9, 673–682. [Google Scholar] [CrossRef] [Green Version]
- Patten, C.; Glick, B. Bacterial biosynthesis of indole-3-acetic acid. Can. J. Microbiol. 1996, 42, 207–220. [Google Scholar] [CrossRef] [PubMed]
- Gruodien, J.; Zvironaite, V. Effect of IAA on growth and synthesis of N compounds in Lucerne. Luk TSR Aukst. Mosklo Darb. Biol. 1971, 17, 77–87. [Google Scholar]
- Barazani, O.Z.; Friedman, J. Is IAA the major root growth factor secreted from plant-growth-mediating bacteria? J. Chem. Ecol. 1999, 25, 2397–2406. [Google Scholar] [CrossRef]
- Spaepen, S.; Vanderleyden, J. Auxin and plant-microbe interactions. Cold Spring Harb. Perspect. Biol. 2011, 3, a001438. [Google Scholar] [CrossRef] [Green Version]
- Shamseldin, A.; El-Saadani, M.; Sadowsky, A.C. Rapid identification and discrimination among Egyptian genotypes of Rhizobium leguminosarum bv. viciae and Sinorhizobium meliloti nodulating faba bean (Vicia faba L.) by analysis of nodC, ARDRA, and rDNA sequence analysis. Soil Biol. Biochem. 2009, 41, 45–53. [Google Scholar] [CrossRef]
- Benidire, L.; Lahrouni, M.; Daoui, K.; El-Abidine Fatemi, Z.; Carmona, R.; Göttfert, M.; Oufdou, K. Phenotypic and genetic diversity of Moroccan rhizobia isolated from Vicia faba and study of genes that are likely to be involved in their osmotolerance. Syst. Appl. Microbiol. 2018, 41, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Sandhya, V.; Grover, M.; Reddy, G.; Venkateswarlu, B. Alleviation of drought stress effects in sunflower seedlings by the exopolysaccharides producing Pseudomonas putida strain GAP-P45. Biol. Fertil. Soils 2009, 46, 17–26. [Google Scholar] [CrossRef]
- Alikhani, H.A.; Mohamadi, L. Assessing tolerance of rhizobial lentil symbiosis isolates to salinity and drought in dry land farming condition. In Proceedings of the 19th World Congress of Soil Science, Soil Solutions for a Changing World, Brisbane, Australia, 1–6 August 2010; pp. 1–6. [Google Scholar]
- Upadhyay, S.; Singh, J.; Singh, D. Exopolysaccharide-producing plant growth-promoting rhizobacteria under salinity condition. Pedosphere 2011, 21, 214–222. [Google Scholar] [CrossRef]
- Pandey, A.; Mann, M. Proteomics to study genes and genomes. Nature 2000, 405, 837. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Jiang, J.; Li, X.; Wang, L.; Yang, S. Isolation of salt-sensitive mutants from Sinorhizobium meliloti and characterization of genes involved in salt tolerance. Lett. Appl. Microbiol. 2004, 39, 278–283. [Google Scholar] [CrossRef] [PubMed]
- Mohammad, R.; Akhavan-Kharazian, M.; Campbell, W.; Rumbaugh, M. Identification of salt-and drought-tolerant Rhizobium meliloti L. strains. Plant Soil 1991, 134, 271–276. [Google Scholar] [CrossRef]
- Domínguez-Ferreras, A.; Pérez-Arnedo, R.; Becker, A.; Olivares, J.; Soto, M.; Sanjuán, J. Transcriptome profiling reveals the importance of plasmid pSymB for osmoadaptation of Sinorhizobium meliloti. J. Bacteriol. 2006, 188, 7617–7625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, J.; Wei, W.; Du, B.; Li, H.; Wang, L.; Yang, S. Salt-tolerance genes involved in cation efflux and osmoregulation of Sinorhizobium fredii RT19 detected by isolation and characterization of Tn5 mutants. FEMS Microbiol. Lett. 2004, 239, 139–146. [Google Scholar] [CrossRef] [Green Version]
- Zahran, H. Rhizobium-legume symbiosis and nitrogen fixation under severe conditions and in an arid climate. Microbiol. Mol. Biol. Rev. 1999, 63, 968–989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piccoli, P.; Bottini, R. Abiotic stress tolerance induced by endophytic PGPR. In Symbiotic Endophytes; Springer: Berlin/Heidelberg, Germany, 2013; pp. 151–163. [Google Scholar]
- Naveed, M.; Hussain, M.; Mehboob, I.; Zahir, Z. Rhizobial Amelioration of Drought Stress in Legumes. In Microbes for Legume Improvement; Springer: Cham, Switzerland, 2017; pp. 341–365. [Google Scholar]
- Rüberg, S.; Tian, Z.; Krol, E.; Linke, B.; Meyer, F.; Wang, Y.; Becker, A. Construction and validation of a Sinorhizobium meliloti whole genome DNA microarray: Genome-wide profiling of osmoadaptive gene expression. J. Biotechnol. 2003, 106, 255–268. [Google Scholar] [CrossRef] [PubMed]
- Zahran, H. Legume-microbe interactions under stressed environments. In Microbes for Legume Improvement; Springer: Cham, Switzerland, 2017; pp. 301–339. [Google Scholar]
- Vance, C. Symbiotic nitrogen fixation and phosphorus acquisition. Plant nutrition in a world of declining renewable resources. Plant Physiol. 2001, 127, 390–397. [Google Scholar] [CrossRef] [PubMed]
- Williams, P. Quorum sensing, communication and cross-kingdom signaling in the bacterial world. Microbiology 2007, 153, 3923–3938. [Google Scholar] [CrossRef] [PubMed] [Green Version]













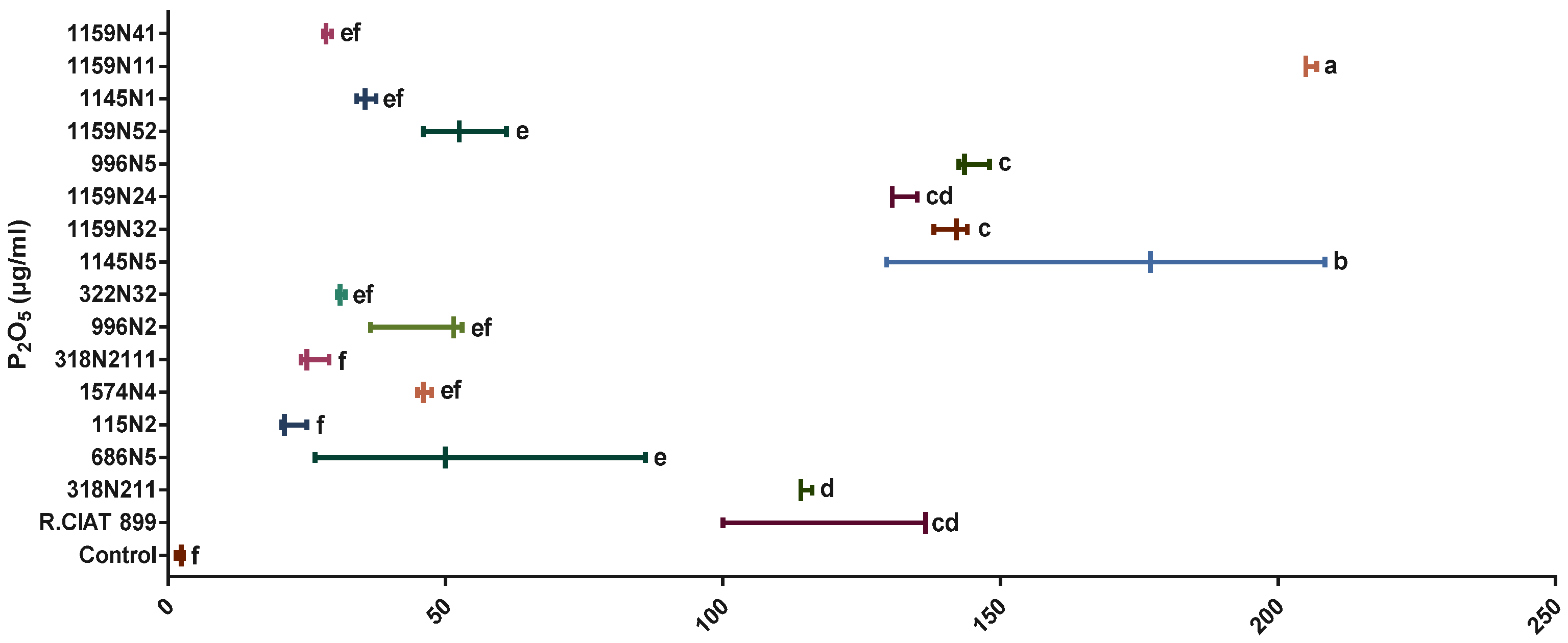{kind=link}
{kind=link}
{kind=link}
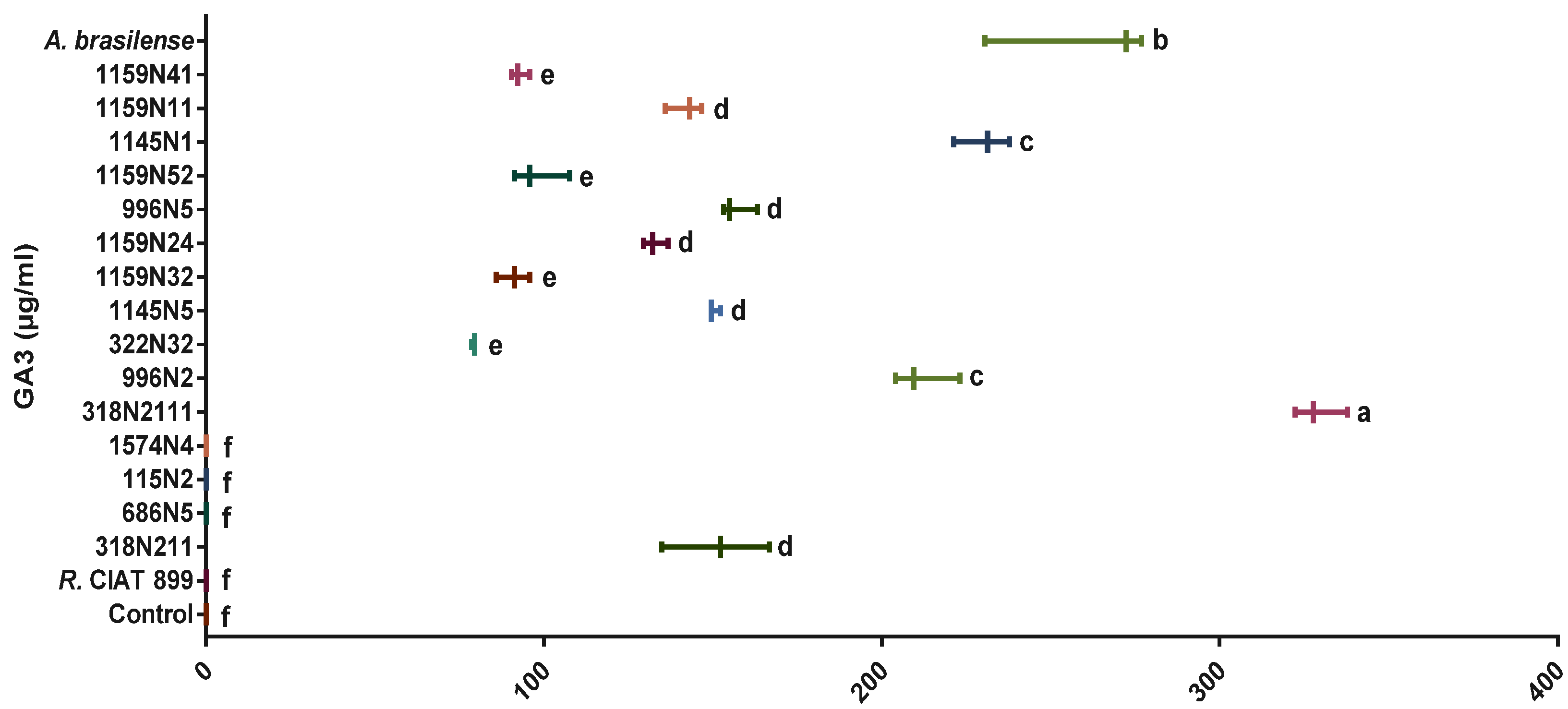{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | 16S rDNA AN | SS | PGPA | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0% | 0.5% | 1% | 1.50% | 2% | 3% | 4% | 5% | IAA | PS | S | ||
| R. tropici CIAT 899 | NR102511 | +++ | +++ | +++ | +++ | +++ | ++ | ++ | ++ | + | 0.97 ± 0.057b | 1.45 ± 0.201bc |
| 318N211 | MK483120 | +++ | +++ | +++ | +++ | ++ | - | - | - | +++ | - | 1.18 ± 0.681d |
| 686N5 | MK483121 | +++ | +++ | +++ | +++ | +++ | ++ | ++ | ++ | + | 0.83 ± 0.046c | 1.41 ± 0.243cd |
| 115N2 | MK483122 | +++ | ++ | + | - | - | - | - | - | - | - | - |
| 1574N4 | MK483123 | +++ | +++ | +++ | +++ | +++ | ++ | ++ | + | - | - | - |
| 318N2111 | MK483124 | +++ | +++ | +++ | +++ | ++ | ++ | + | ++ | + | - | - |
| 996N2 | MK483125 | +++ | +++ | +++ | +++ | +++ | ++ | ++ | + | ++ | - | - |
| 322N32 | MK483126 | +++ | ++ | + | - | - | - | - | - | - | - | - |
| 1145N5 | MK483127 | +++ | +++ | +++ | +++ | +++ | ++ | ++ | - | - | 0.97 ± 0.560bc | 1.38 ± 0.142bc |
| 1159N32 | MK483128 | +++ | +++ | +++ | +++ | +++ | +++ | ++ | + | + | 0.82 ± 0.751bc | 2.07 ± 0.533 b |
| 1159N24 | MK483129 | +++ | +++ | +++ | +++ | +++ | ++ | ++ | - | ++ | - | - |
| 996N5 | MK483130 | +++ | +++ | +++ | +++ | +++ | ++ | ++ | - | + | 0.72 ± 0.284c | 1.83 ± 0.393d |
| 1159N52 | MK483131 | +++ | +++ | +++ | +++ | +++ | ++ | + | - | + | - | 1.58 ± 0.148b |
| 1145N1 | MK483132 | +++ | +++ | +++ | +++ | +++ | ++ | ++ | + | +++ | - | - |
| 1159N11 | MK483133 | +++ | ++ | + | - | - | - | - | - | - | 2.1 ± 0.617c | - |
| 1159N41 | MK483134 | +++ | +++ | +++ | + | - | - | - | - | ++ | - | - |
| A. brasilense DSM-1690 | NR117478 | +++ | +++ | +++ | +++ | +++ | ++ | ++ | + | +++ | - | 2.07 ± 0.716a |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sijilmassi, B.; Filali-Maltouf, A.; Fahde, S.; Ennahli, Y.; Boughribil, S.; Kumar, S.; Amri, A. In-Vitro Plant Growth Promotion of Rhizobium Strains Isolated from Lentil Root Nodules under Abiotic Stresses. Agronomy 2020, 10, 1006. https://doi.org/10.3390/agronomy10071006
Sijilmassi B, Filali-Maltouf A, Fahde S, Ennahli Y, Boughribil S, Kumar S, Amri A. In-Vitro Plant Growth Promotion of Rhizobium Strains Isolated from Lentil Root Nodules under Abiotic Stresses. Agronomy. 2020; 10(7):1006. https://doi.org/10.3390/agronomy10071006
Chicago/Turabian StyleSijilmassi, Badreddine, Abdelkarim Filali-Maltouf, Sara Fahde, Youness Ennahli, Said Boughribil, Shiv Kumar, and Ahmed Amri. 2020. "In-Vitro Plant Growth Promotion of Rhizobium Strains Isolated from Lentil Root Nodules under Abiotic Stresses" Agronomy 10, no. 7: 1006. https://doi.org/10.3390/agronomy10071006