Zoonotic Viruses in Three Species of Voles from Poland

 ,
,  , , , and
, , , and 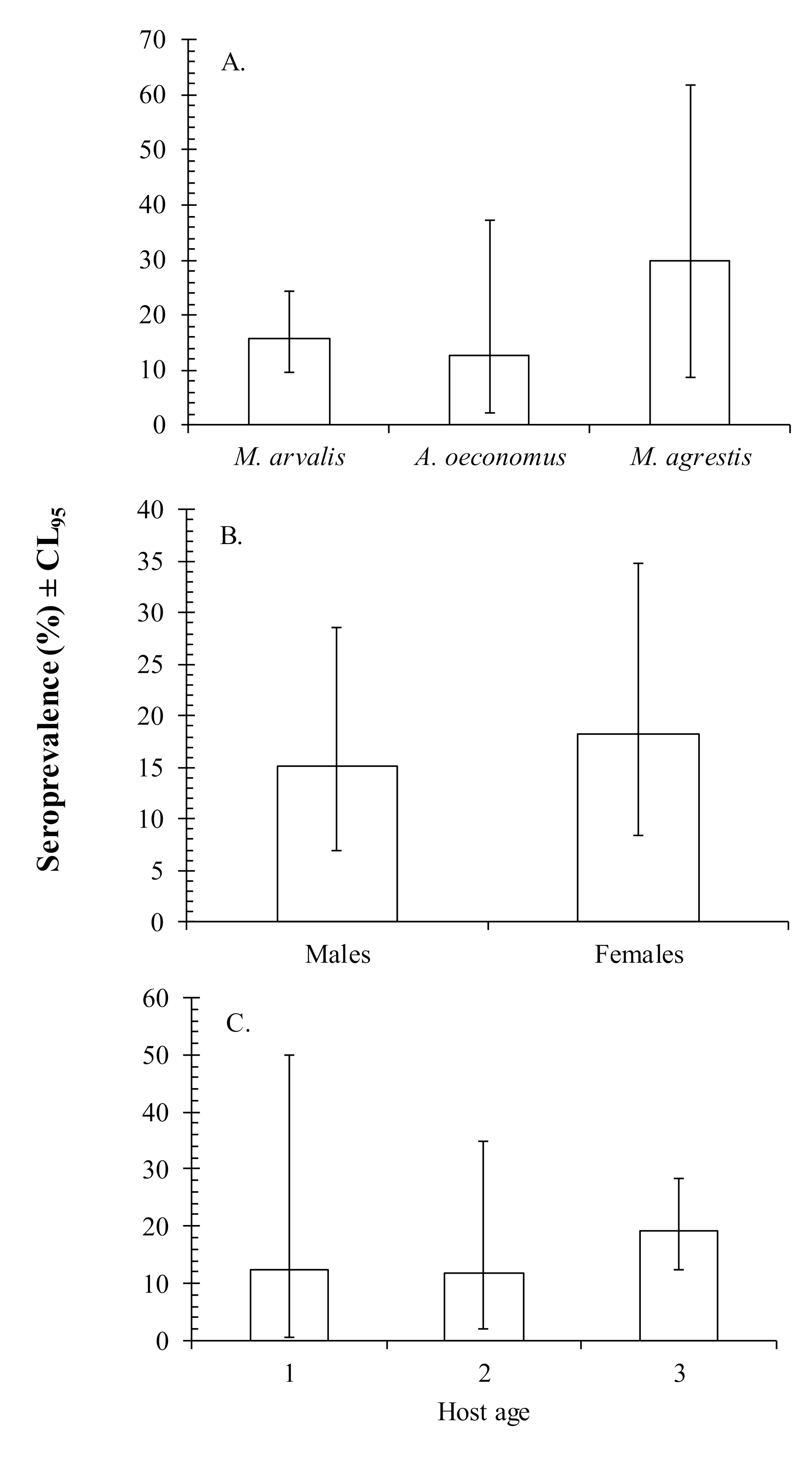{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Approval
2.2. Collection of Voles
2.3. Serological Screening ofAnti-Virus Antibodies
2.4. Statistical Analysis
3. Results
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Luis, A.D.; Hayman, D.T.S.; O’Shea, T.J.; Cryan, P.M.; Gilbert, A.T.; Pulliam, J.R.C.; Mills, J.N.; Timonin, M.E.; Willis, C.K.R.; Cunningham, A.A.; et al. A comparison of bats and rodents as reservoirs of zoonotic viruses: Are bats special? Proc. R. Soc. B Biol. Sci. 2013, 280, 20122753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mills, J. Ecologic Studies of Rodent Reservoirs: Their Relevance for Human Health. Emerg. Infect. Dis. 1998, 4, 529–537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bordes, F.; Blasdell, K.; Morand, S. Transmission ecology of rodent-borne diseases: New frontiers. Integr. Zool. 2015, 10, 424–435. [Google Scholar] [CrossRef] [PubMed]
- Kallio, E.R.; Begon, M.; Henttonen, H.; Koskela, E.; Mappes, T.; Vaheri, A.; Vapalahti, O. Cyclic hantavirus epidemics in humans—Predicted by rodent host dynamics. Epidemics 2009, 1, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Olsson, G.E.; Hjertqvist, M.; Lundkvist, Å.; Hörnfeldt, B. Predicting High Risk for Human Hantavirus Infections, Sweden. Emerg. Infect. Dis. 2009, 15, 104–106. [Google Scholar] [CrossRef] [PubMed]
- Carroll, K.; Hobden, J.; Miller, S.; Morse, S.; Mietzner, T.; Detrick, B.; Mitchell, T.; McKerrow, J.; Sakanari, J. Arthropod-Borne and Rodent-Borne Viral Diseases. In Jawetz Melnick & Adelbergs Medical Microbiology; McGraw-Hill: New York, NY, USA, 2015; ISBN 0071824987. [Google Scholar]
- Vapalahti, O.; Mustonen, J.; Lundkvist, A.; Henttonen, H.; Plyusnin, A.; Vaheri, A. Hantavirus infections in Europe. Lancet Infect. Dis. 2003, 3, 653–661. [Google Scholar] [CrossRef]
- Charrel, R.N.; de Lamballerie, X. Zoonotic aspects of arenavirus infections. Vet. Microbiol. 2010, 140, 213–220. [Google Scholar] [CrossRef] [Green Version]
- Kallio, E.R.; Voutilainen, L.; Vapalahti, O.; Vaheri, A.; Henttonen, H.; Koskela, E.; Mappes, T. Endemic hantavirus infection impairs the winter survival of its rodent host. Ecology 2007, 88, 1911–1916. [Google Scholar] [CrossRef] [Green Version]
- Kallio-Kokko, H.; Uzcategui, N.; Vapalahti, O.; Vaheri, A. Viral zoonoses in Europe. FEMS Microbiol. Rev. 2005, 29, 1051–1077. [Google Scholar] [CrossRef] [Green Version]
- Kinnunen, P.M.; Henttonen, H.; Hoffmann, B.; Kallio, E.R.; Korthase, C.; Laakkonen, J.; Niemimaa, J.; Palva, A.; Schlegel, M.; Ali, H.S.; et al. Orthopox Virus Infections in Eurasian Wild Rodents. Vector-Borne Zoonotic Dis. 2011, 11, 1133–1140. [Google Scholar] [CrossRef]
- Tonteri, E.; Kipar, A.; Voutilainen, L.; Vene, S.; Vaheri, A.; Vapalahti, O.; Lundkvist, Å. The three subtypes of tick-borne encephalitis virus induce encephalitis in a natural host, the bank vole (Myodes glareolus). PLoS ONE 2013, 8, e81214. [Google Scholar] [CrossRef] [PubMed]
- Jääskeläinen, A.J.; Kolehmainen, P.; Voutilainen, L.; Hauffe, H.C.; Kallio-Kokko, H.; Lappalainen, M.; Tolf, C.; Lindberg, A.M.; Henttonen, H.; Vaheri, A.; et al. Evidence of Ljungan Virus specific antibodies in humans and rodents, Finland. J. Med. Virol. 2013, 85, 2001–2008. [Google Scholar] [CrossRef] [PubMed]
- Grzybek, M.; Sironen, T.; Mäki, S.; Tołkacz, K.; Alsarraf, M.; Strachecka, A.; Paleolog, J.; Biernat, B.; Szczepaniak, K.; Behnke-Borowczyk, J.; et al. Zoonotic Virus Seroprevalence among Bank Voles, Poland, 2002–2010. Emerg. Infect. Dis. 2019, 25, 1607–1609. [Google Scholar] [CrossRef] [PubMed]
- Goeijenbier, M.; Wagenaar, J.; Goris, M.; Martina, B.; Henttonen, H.; Vaheri, A.; Reusken, C.; Hartskeerl, R.; Osterhaus, A.; Van Gorp, E. Rodent-borne hemorrhagic fevers: Under-recognized, widely spread and preventable—Epidemiology, diagnostics and treatment. Crit. Rev. Microbiol. 2013, 39, 26–42. [Google Scholar] [CrossRef] [PubMed]
- Brummer-Korvenkontio, M.; Henttonen, H.; Vaheri, A. Hemorrhagic fever with renal syndrome in Finland: Ecology and virology of nephropathia epidemica. Scand. J. Infect. Dis. Suppl. 1982, 36, 88–91. [Google Scholar]
- Voutilainen, L.; Sironen, T.; Tonteri, E.; Bäck, A.T.; Razzauti, M.; Karlsson, M.; Wahlström, M.; Niemimaa, J.; Henttonen, H.; Lundkvist, Å. Life-long shedding of Puumala hantavirus in wild bank voles (Myodes glareolus). J. Gen. Virol. 2015, 96, 1238–1247. [Google Scholar] [CrossRef]
- Meyer, B.J.; Schmaljohn, C.S. Persistent hantavirus infections: Characteristics and mechanisms. Trends Microbiol. 2000, 8, 61–67. [Google Scholar] [CrossRef] [Green Version]
- Crouch, A.C.; Baxby, D.; Mccracken, C.M.; Gaskell, R.M.; Bennett, M. Serological evidence for the reservoir hosts of cowpox virus in British wildlife. Epidemiol. Infect. 1995, 115, 185–191. [Google Scholar] [CrossRef] [Green Version]
- Chantrey, J.; Meyer, H.; Baxby, D.; Begon, M.; Bown, K.J.; Hazel, S.M.; Jones, T.; Montgomery, W.I.; Bennett, M. Cowpox: Reservoir hosts and geographic range. Epidemiol. Infect. 1999, 122, 455–460. [Google Scholar] [CrossRef] [Green Version]
- Blasdell, K.R.; Becker, S.D.; Hurst, J.; Begon, M.; Bennett, M. Host Range and Genetic Diversity of Arenaviruses in Rodents, United Kingdom. Emerg. Infect. Dis. 2008, 14, 1455–1458. [Google Scholar] [CrossRef]
- Laakkonen, J.; Kallio-Kokko, H.; Öktem, M.A.; Blasdell, K.; Plyusnina, A.; Niemimaa, J.; Karataş, A.; Plyusnin, A.; Vaheri, A.; Henttonen, H. Serological Survey for Viral Pathogens in Turkish Rodents. J. Wildl. Dis. 2006, 42, 672–676. [Google Scholar] [CrossRef] [PubMed]
- Tagliapietra, V.; Rosà, R.; Hauffe, H.C.; Laakkonen, J.; Voutilainen, L.; Vapalahti, O.; Vaheri, A.; Henttonen, H.; Rizzoli, A. Spatial and temporal dynamics of lymphocytic choriomeningitis virus in wild rodents, northern Italy. Emerg. Infect. Dis. 2009, 15, 1019–1025. [Google Scholar] [CrossRef] [PubMed]
- Kallio, E.R.; Klingström, J.; Gustafsson, E.; Manni, T.; Vaheri, A.; Henttonen, H.; Vapalahti, O.; Lundkvist, Å. Prolonged survival of Puumala hantavirus outside the host: Evidence for indirect transmission via the environment. J. Gen. Virol. 2006, 87, 2127–2134. [Google Scholar] [CrossRef] [PubMed]
- Ledesma, J.; Fedele, C.G.; Carro, F.; Lledó, L.; Sánchez-Seco, M.P.; Tenorio, A.; Soriguer, R.C.; Saz, J.V.; Domínguez, G.; Rosas, M.F.; et al. Independent Lineage of Lymphocytic Choriomeningitis Virus in Wood Mice (Apodemus sylvaticus), Spain. Emerg. Infect. Dis. 2009, 15, 1677–1680. [Google Scholar] [CrossRef] [PubMed]
- ECDC. Annual Epidemiological Report for 2018. Stockholm. 2020. Available online: https://www.ecdc.europa.eu/sites/default/files/documents/hantavirus-annual-epidemiological-report-2018.pdf (accessed on 7 August 2020).
- Świtaj, K.; Kajfasz, P.; Kurth, A.; Nitsche, A. Cowpox after a cat scratch—case report from Poland. Ann. Agric. Environ. Med. 2015, 22, 456–458. [Google Scholar] [CrossRef]
- Hanski, I.; Henttonen, H. Predation on Competing Rodent Species: A Simple Explanation of Complex Patterns. J. Anim. Ecol. 1996, 65, 220–232. [Google Scholar] [CrossRef]
- Tołkacz, K.; Alsarraf, M.; Kowalec, M.; Dwużnik, D.; Grzybek, M.; Behnke, J.M.; Bajer, A. Bartonella infections in three species of Microtus: Prevalence and genetic diversity, vertical transmission and the effect of concurrent Babesia microti infection on its success. Parasites Vectors 2018, 11, 491. [Google Scholar] [CrossRef]
- Tołkacz, K.; Bednarska, M.; Alsarraf, M.; Dwużnik, D.; Grzybek, M.; Welc-Falęciak, R.; Behnke, J.M.; Bajer, A. Prevalence, genetic identity and vertical transmission of Babesia microti in three naturally infected species of vole, Microtus spp. (Cricetidae). Parasites Vectors 2017, 10, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Pawelczyk, A.; Bajer, A.; Behnke, J.M.; Gilbert, F.S.; Sinski, E. Factors affecting the component community structure of haemoparasites in common voles (Microtus arvalis) from the Mazury Lake District region of Poland. Parasitol. Res. 2004, 92, 270–284. [Google Scholar] [CrossRef]
- Haukisalmi, V.; Henttonen, H.; Tenora, F. Population Dynamics of Common and Rare Helminths in Cyclic Vole Populations. J. Anim. Ecol. 1988, 57, 807–825. [Google Scholar] [CrossRef]
- Bannikova, A.A.; Lebedev, V.S.; Lissovsky, A.A.; Matrosova, V.; Abramson, N.I.; Obolenskaya, E.V.; Tesakov, A.S. Molecular phylogeny and evolution of the Asian lineage of vole genus Microtus (Rodentia: Arvicolinae) inferred from mitochondrial cytochrome b sequence. Biol. J. Linn. Soc. 2010, 99, 595–613. [Google Scholar] [CrossRef] [Green Version]
- Lissovsky, A.A.; Petrova, T.V.; Yatsentyuk, S.P.; Golenishchev, F.N.; Putincev, N.I.; Kartavtseva, I.V.; Sheremetyeva, I.N.; Abramson, N.I. Multilocus phylogeny and taxonomy of East Asian voles Alexandromys (Rodentia, Arvicolinae). Zool. Scr. 2018, 47, 9–20. [Google Scholar] [CrossRef]
- Zorenko, T.A.; Atanasov, N.I. Copulatory behavior supports a new insight into taxonomic status of root vole Alexandromys oeconomus (Rodentia, Arvicolinae). Russ. J. Theriol. 2018, 17, 48–57. [Google Scholar] [CrossRef]
- Pucek, Z. Keys to Vertebrates of Poland. Mammals; Polish Scientific Publishers: Warsaw, Poland, 1981; ISBN 8301025530 9788301025533. [Google Scholar]
- Hedman, K.; Vaheri, A.; Brummer-Korvenkontio, M. Rapid diagnosis of hantavirus disease with an IgG-avidity assay. Lancet 1991, 338, 1353–1356. [Google Scholar] [CrossRef]
- Kallio-Kokko, H.; Laakkonen, J.; Rizzoli, A.; Tagliapietra, V.; Cattadori, I.; Perkins, S.E.; Hudson, P.J.; Cristofolini, A.; Versini, W.; Vapalahti, O.; et al. Hantavirus and arenavirus antibody prevalence in rodents and humans in Trentino, Northern Italy. Epidemiol. Infect. 2006, 134, 830–836. [Google Scholar] [CrossRef]
- Sokal, R.R.; Rohlf, F.J. Biometry: The Principles and Practices of Statistics in Biological Research; W. H. Freeman: New York, NY, USA, 1994; ISBN 978-0716724117. [Google Scholar]
- Grzybek, M.; Bajer, A.; Bednarska, M.M.; Al-Sarraf, M.; Behnke-Borowczyk, J.; Harris, P.D.; Price, S.J.; Brown, G.S.; Osborne, S.-J.; Siński, E.; et al. Long-term spatiotemporal stability and dynamic changes in helminth infracommunities of bank voles (Myodes glareolus) in NE Poland. Parasitology 2015, 142, 1722–1743. [Google Scholar] [CrossRef]
- Grzybek, M.; Alsarraf, M.; Tołkacz, K.; Behnke-Borowczyk, J.; Biernat, B.; Stańczak, J.; Strachecka, A.; Guz, L.; Szczepaniak, K.; Paleolog, J.; et al. Seroprevalence of TBEV in bank voles from Poland—A long-term approach. Emerg. Microbes Infect. 2018, 7, 145. [Google Scholar] [CrossRef] [Green Version]
- Behnke, J.M.; Barnard, C.J.; Bajer, A.; Bray, D.; Dinmore, J.; Frake, K.; Osmond, J.; Race, T.; Sinski, E. Variation in the helminth community structure in bank voles (Clethrionomys glareolus) from three comparable localities in the mazury lake istrict region of Poland. Parasitology 2001, 123, 401–414. [Google Scholar] [CrossRef]
- Behnke, J.M.; Bajer, A.; Harris, P.D.; Newington, L.; Pidgeon, E.; Rowlands, G.; Sheriff, C.; Kuliś-Malkowska, K.; Siński, E.; Gilbert, F.S.; et al. Temporal and between-site variation in helminth communities of bank voles (Myodes glareolus) from N.E. Poland. 1. Regional fauna and component community levels. Parasitology 2008, 135, 985–997. [Google Scholar] [CrossRef]
- Bajer, A.; Behnke, J.M.; Pawełczyk, A.; Kuliś, K.; Sereda, M.J.; Siński, E. Medium-term temporal stability of the helminth component community structure in bank voles (Clethrionomys glareolus) from the Mazury Lake District region of Poland. Parasitology 2005, 130, 213–228. [Google Scholar] [CrossRef]
- Razzauti, M.; Plyusnina, A.; Sironen, T.; Henttonen, H.; Plyusnin, A. Analysis of Puumala hantavirus in a bank vole population in northern Finland: Evidence for co-circulation of two genetic lineages and frequent reassortment between strains. J. Gen. Virol. 2009, 90, 1923–1931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomason, A.G.; Begon, M.; Bradley, J.E.; Paterson, S.; Jackson, J.A. Endemic Hantavirus in Field Voles, Northern England. Emerg. Infect. Dis. 2017, 23, 1033–1035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forbes, K.M.; Voutilainen, L.; Jääskeläinen, A.; Sironen, T.; Kinnunen, P.M.; Stuart, P.; Vapalahti, O.; Henttonen, H.; Huitu, O. Serological Survey of Rodent-Borne Viruses in Finnish Field Voles. Vector-Borne Zoonotic Dis. 2014, 14, 278–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt-Chanasit, J.; Essbauer, S.; Petraityte, R.; Yoshimatsu, K.; Tackmann, K.; Conraths, F.J.; Sasnauskas, K.; Arikawa, J.; Thomas, A.; Pfeffer, M.; et al. Extensive Host Sharing of Central European Tula Virus. J. Virol. 2010, 84, 459–474. [Google Scholar] [CrossRef] [Green Version]
- Olsson, G.E.; Leirs, H.; Henttonen, H. Hantaviruses and Their Hosts in Europe: Reservoirs Here and There, But Not Everywhere? Vector-Borne Zoonotic Dis. 2010, 10, 549–561. [Google Scholar] [CrossRef]
- Vaheri, A.; Henttonen, H.; Voutilainen, L.; Mustonen, J.; Sironen, T.; Vapalahti, O. Hantavirus infections in Europe and their impact on public health. Rev. Med. Virol. 2013, 23, 35–49. [Google Scholar] [CrossRef]
- Sironen, T.; Plyusnina, A.; Andersen, H.K.; Lodal, J.; Leirs, H.; Niemimaa, J.; Henttonen, H.; Vaheri, A.; Lundkvist, Å.; Plyusnin, A. Distribution of Puumala Hantavirus in Denmark: Analysis of Bank Voles (Clethrionomys glareolus) from Fyn and Jutland. Vector-Borne Zoonotic Dis. 2002, 2, 37–45. [Google Scholar] [CrossRef]
- Vaheri, A.; Vapalahti, O.; Plyusnin, A. How to diagnose hantavirus infections and detect them in rodents and insectivores. Rev. Med. Virol. 2008, 18, 277–288. [Google Scholar] [CrossRef]
- Vieth, S.; Drosten, C.; Lenz, O.; Vincent, M.; Omilabu, S.; Hass, M.; Becker-Ziaja, B.; ter Meulen, J.; Nichol, S.T.; Schmitz, H.; et al. RT-PCR assay for detection of Lassa virus and related Old World arenaviruses targeting the L gene. Trans. R. Soc. Trop. Med. Hyg. 2007, 101, 1253–1264. [Google Scholar] [CrossRef]
- Jonsson, C.B.; Figueiredo, L.T.M.; Vapalahti, O. A Global Perspective on Hantavirus Ecology, Epidemiology, and Disease. Clin. Microbiol. Rev. 2010, 23, 412–441. [Google Scholar] [CrossRef] [Green Version]
- Yee, J.; Wortman, I.A.; Nofchissey, R.A.; Goade, D.; Bennett, S.G.; Webb, J.P.; Irwin, W.; Hjelle, B. Rapid and simple method for screening wild rodents for antibodies to Sin Nombre hantavirus. J. Wildl. Dis. 2003, 39, 271–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, B.A.; Schmidt, J.P.; Bowden, S.E.; Drake, J.M. Rodent reservoirs of future zoonotic diseases. Proc. Natl. Acad. Sci. USA 2015, 112, 7039–7044. [Google Scholar] [CrossRef] [PubMed] [Green Version]

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grzybek, M.; Tołkacz, K.; Sironen, T.; Mäki, S.; Alsarraf, M.; Behnke-Borowczyk, J.; Biernat, B.; Nowicka, J.; Vaheri, A.; Henttonen, H.; et al. Zoonotic Viruses in Three Species of Voles from Poland. Animals 2020, 10, 1820. https://doi.org/10.3390/ani10101820
Grzybek M, Tołkacz K, Sironen T, Mäki S, Alsarraf M, Behnke-Borowczyk J, Biernat B, Nowicka J, Vaheri A, Henttonen H, et al. Zoonotic Viruses in Three Species of Voles from Poland. Animals. 2020; 10(10):1820. https://doi.org/10.3390/ani10101820
Chicago/Turabian StyleGrzybek, Maciej, Katarzyna Tołkacz, Tarja Sironen, Sanna Mäki, Mohammed Alsarraf, Jolanta Behnke-Borowczyk, Beata Biernat, Joanna Nowicka, Antti Vaheri, Heikki Henttonen, and et al. 2020. "Zoonotic Viruses in Three Species of Voles from Poland" Animals 10, no. 10: 1820. https://doi.org/10.3390/ani10101820