The Greater Proportion of Born-Light Progeny from Sows Mated in Summer Contributes to Increased Carcass Fatness Observed in Spring

 , , , ,
, , , ,  , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Design
2.2. Housing Systems and Farrowing Outcomes of Sows
2.3. Physiological Monitoring and Injury Score of Gestating Sows
2.4. Placental Weight Collection
2.5. Plasma IGF-1 in New-Born Progeny
2.6. Growth Performance and Carcass Traits of Progeny Pigs
2.7. Tissue Composition of Primal Cuts Using Dual-Energy X-ray Absorptiometry (DXA)
2.8. Statistical Analysis
3. Results
3.1. Change of Body Condition during Gestation
3.2. Farrowing Outcomes
3.3. Placental Weight of Focal Sows
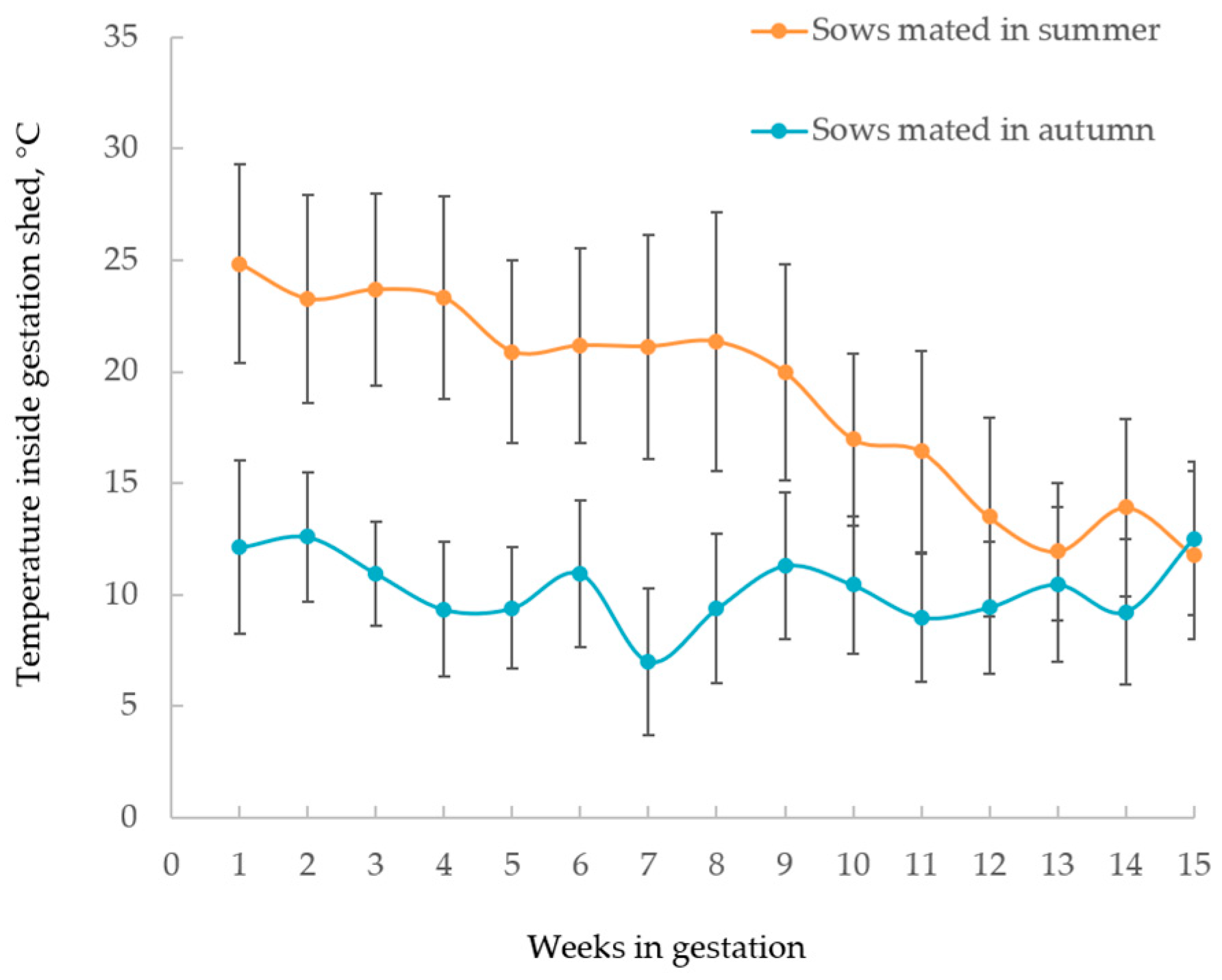
3.4. Physiological Signs of Heat Stress and Number of Scars on Sows
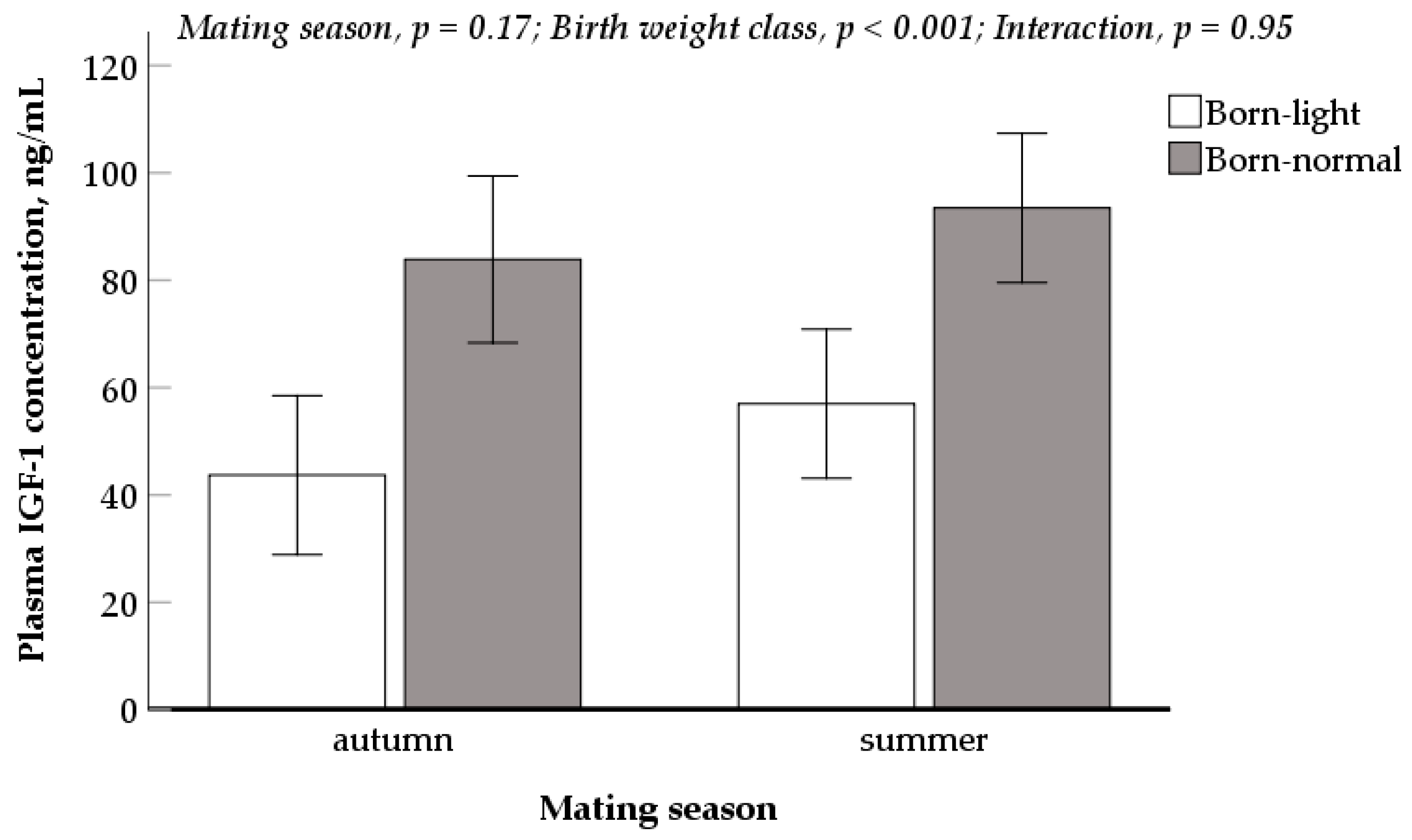
3.5. Plasma IGF-1 Concentration of Newborn Piglets
3.6. Growth Performance from 9 to 14 Weeks of Age and from 14 Weeks of Age to 101 kg Live Weight
3.7. Carcass Traits of Progeny
3.8. Tissue Composition of Primal Cuts and Half Carcasses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Trezona, M.; Mullan, B.P.; D’Antuono, M.; Wilson, R.H.; Williams, I.H. The causes of seasonal variation in backfat thickness of pigs in Western Australia. Aust. J. Agric. Res. 2004, 55, 273–277. [Google Scholar] [CrossRef]
- Boddicker, R.L.; Seibert, J.T.; Johnson, J.S.; Pearce, S.C.; Selsby, J.T.; Gabler, N.K.; Lucy, M.C.; Safranski, T.J.; Rhoads, R.P.; Baumgard, L.H.; et al. Gestational heat stress alters postnatal offspring body composition indices and metabolic parameters in pigs. PLoS ONE 2014, 9, e110859. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.S.; Baumgard, L.H. PHYSIOLOGY SYMPOSIUM: Postnatal consequences of in utero heat stress in pigs. J. Anim. Sci. 2019, 97, 962–971. [Google Scholar] [CrossRef] [Green Version]
- Poore, K.R.; Fowden, A.L. The effects of birth weight and postnatal growth patterns on fat depth and plasma leptin concentrations in juvenile and adult pigs. J. Physiol. 2004, 558, 295–304. [Google Scholar] [CrossRef] [PubMed]
- Bee, G. Effect of early gestation feeding, birth weight, and gender of progeny on muscle fiber characteristics of pigs at slaughter. J. Anim. Sci. 2004, 82, 826–836. [Google Scholar] [CrossRef]
- Rehfeldt, C.; Tuchscherer, A.; Hartung, M.; Kuhn, G. A second look at the influence of birth weight on carcass and meat quality in pigs. Meat Sci. 2008, 78, 170–175. [Google Scholar] [CrossRef] [PubMed]
- Karlen, G.A.M.; Hemsworth, P.H.; Gonyou, H.W.; Fabrega, E.; David Strom, A.; Smits, R.J. The welfare of gestating sows in conventional stalls and large groups on deep litter. Appl. Anim. Behav. Sci. 2007, 105, 87–101. [Google Scholar] [CrossRef]
- Australian Pork Limited. Pork Training Manual; Australian Pork Limited: Canberra, ACT, Australia, 2018; p. 88. [Google Scholar]
- Suster, D.; Leury, B.J.; Hofmeyr, C.D.; D’Souza, D.N.; Dunshea, F.R. The accuracy of dual energy X-ray absorptiometry (DXA), weight, and P2 back fat to predict half-carcass and primal-cut composition in pigs within and across research experiments. Aust. J. Agric. Res. 2004, 55, 973–982. [Google Scholar] [CrossRef]
- Lucy, M.C.; Safranski, T.J.; Rhoades, J.N.; Ross, J.W.; Gabler, N.K.; Rhoads, R.P.; Baumgard, L.H. Litter characteristics and thermoregulatory behavior of first parity sows exposed to a controlled heat stress (HS) during gestation. J. Anim. Sci. 2012, 90, 731. [Google Scholar]
- Cruzen, S.M.; Boddicker, R.L.; Graves, K.L.; Johnson, T.P.; Arkfeld, E.K.; Baumgard, L.H.; Ross, J.W.; Safranski, T.J.; Lucy, M.C.; Lonergan, S.M. Carcass composition of market weight pigs subjected to heat stress in utero and during finishing. J. Anim. Sci. 2015, 93, 2587–2596. [Google Scholar] [CrossRef]
- Williams, A.M.; Safranski, T.J.; Spiers, D.E.; Eichen, P.A.; Coate, E.A.; Lucy, M.C. Effects of a controlled heat stress during late gestation, lactation, and after weaning on thermoregulation, metabolism, and reproduction of primiparous sows. J. Anim. Sci. 2013, 91, 2700–2714. [Google Scholar] [CrossRef] [PubMed]
- Bell, A.W.; McBride, B.W.; Slepetis, R.; Early, R.J.; Currie, W.B. Chronic heat stress and prenatal development in sheep: I. Conceptus growth and maternal plasma hormones and metabolites. J. Anim. Sci. 1989, 67, 3289–3299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Early, R.J.; McBride, B.W.; Vatnick, I.; Bell, A.W. Chronic heat stress and prenatal development in sheep: II. Placental cellularity and metabolism. J. Anim. Sci. 1991, 69, 3610–3616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Limesand, S.W.; Rozance, P.J.; Smith, D.; Hay, W.W. Increased insulin sensitivity and maintenance of glucose utilization rates in fetal sheep with placental insufficiency and intrauterine growth restriction. Am. J. Physiol. Endocrinol. Metab. 2007, 293, E1716–E1725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, R.; Yin, Y.; Pan, J.; Gao, Y.; Li, T. Expression profiling of IGFs and IGF receptors in piglets with intrauterine growth restriction. Livest. Sci. 2011, 136, 72–75. [Google Scholar] [CrossRef]
- Schoknecht, P.A.; Ebner, S.; Skottner, A.; Burrin, D.G.; Davis, T.A.; Ellis, K.; Pond, W.G. Exogenous insulin-like growth factor-I increases weight gain in intrauterine growth-retarded neonatal pigs. Pediatr. Res. 1997, 42, 201–207. [Google Scholar] [CrossRef] [Green Version]
- Van Barneveld, R.J.; Dunshea, F.R. Colostrum protein isolate increases gut and whole body growth and plasma IGF-I in neonatal pigs. Asian-Aust. J. Anim. Sci. 2011, 24, 670–677. [Google Scholar] [CrossRef]
- Yakar, S.; Liu, J.-L.; Fernandez, A.M.; Wu, Y.; Schally, A.V.; Frystyk, J.; Chernausek, S.D.; Mejia, W.; Le Roith, D. Liver-specific igf-1 gene deletion leads to muscle insulin insensitivity. Diabetes 2001, 50, 1110–1118. [Google Scholar] [CrossRef] [Green Version]
- Lewis, C.; Bunter, K. Effects of seasonality and ambient temperature on genetic parameters for production and reproductive traits in pigs. Anim. Prod. Sci. 2011, 51, 615–626. [Google Scholar] [CrossRef]
- Johnson, J.S.; Sanz Fernandez, M.V.; Patience, J.F.; Ross, J.W.; Gabler, N.K.; Lucy, M.C.; Safranski, T.J.; Rhoads, R.P.; Baumgard, L.H. Effects of in utero heat stress on postnatal body composition in pigs: II. Finishing phase1. J. Anim. Sci. 2015, 93, 82–92. [Google Scholar] [CrossRef] [Green Version]
- Serviento, A.M.; Lebret, B.; Renaudeau, D. Chronic prenatal heat stress alters growth, carcass composition, and physiological response of growing pigs subjected to postnatal heat stress. J. Anim. Sci. 2020, 98, skaa161. [Google Scholar] [CrossRef] [PubMed]
- Renaudeau, D.; Gourdine, J.L.; St-Pierre, N.R. A meta-analysis of the effects of high ambient temperature on growth performance of growing-finishing pigs. J. Anim. Sci. 2011, 89, 2220–2230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alvarenga, A.L.N.; Chiarini-Garcia, H.; Cardeal, P.C.; Moreira, L.P.; Foxcroft, G.R.; Fontes, D.O.; Almeida, F.R.C.L. Intra-uterine growth retardation affects birthweight and postnatal development in pigs, impairing muscle accretion, duodenal mucosa morphology and carcass traits. Reprod. Fertil. Dev. 2013, 25, 387–395. [Google Scholar] [CrossRef] [PubMed]
- Schinckel, A.; Pas, M.; Einstein, M.; Jungst, S.; Booher, C.; Newman, S. Evaluation of the impact of pig birth weight on grow-finish performance, backfat depth, and loin depth. Prof. Anim. Sci. 2010, 26. [Google Scholar] [CrossRef]
- Gondret, F.; Lefaucheur, L.; Louveau, I.; Lebret, B.; Pichodo, X.; Le Cozler, Y. Influence of piglet birth weight on postnatal growth performance, tissue lipogenic capacity and muscle histological traits at market weight. Livest. Prod. Sci. 2005, 93, 137–146. [Google Scholar] [CrossRef]
- Stange, K.; Miersch, C.; Sponder, G.; Röntgen, M. Low birth weight influences the postnatal abundance and characteristics of satellite cell subpopulations in pigs. Sci. Rep. 2020, 10, 6149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greenwood, P.L.; Hunt, A.S.; Hermanson, J.W.; Bell, A.W. Effects of birth weight and postnatal nutrition on neonatal sheep: I. Body growth and composition, and some aspects of energetic efficiency3. J. Anim. Sci. 1998, 76, 2354–2367. [Google Scholar] [CrossRef] [PubMed]
- Greenwood, P.L.; Hunt, A.S.; Hermanson, J.W.; Bell, A.W. Effects of birth weight and postnatal nutrition on neonatal sheep: II. Skeletal muscle growth and development2. J. Anim. Sci. 2000, 78, 50–61. [Google Scholar] [CrossRef] [Green Version]
- Johnston, L.J.; Ellis, M.; Libal, G.W.; Mayrose, V.B.; Weldon, W.C. Effect of room temperature and dietary amino acid concentration on performance of lactating sows. NCR-89 Committee on Swine Management. J. Anim. Sci. 1999, 77, 1638–1644. [Google Scholar] [CrossRef] [Green Version]
- Gourdine, J.L.; Bidanel, J.P.; Noblet, J.; Renaudeau, D. Effects of breed and season on performance of lactating sows in a tropical humid climate. J. Anim. Sci. 2006, 84, 360–369. [Google Scholar] [CrossRef]
- Liu, F.; Hogg, J.; Kracht, S.; Brewster, C.J.; Henman, D.J.; Athorn, R.Z.; Morrison, R.S.; Smits, R.J.; Campbell, R.G. Supplementing 2 g per day bovine lactoferrin from late gestation until weaning did not improve lactation performance of mixed parity sows. Anim. Prod. Sci. 2019, 59, 2191–2195. [Google Scholar] [CrossRef]
- Liu, F.; de Ruyter, E.M.; Athorn, R.Z.; Brewster, C.J.; Henman, D.J.; Morrison, R.S.; Smits, R.J.; Cottrell, J.J.; Dunshea, F.R. Effects of L-citrulline supplementation on heat stress physiology, lactation performance and subsequent reproductive performance of sows in summer. J. Anim. Physiol. Anim. Nutr. 2019, 103, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Wadhwa, P.D.; Garite, T.J.; Porto, M.; Glynn, L.; Chicz-DeMet, A.; Dunkel-Schetter, C.; Sandman, C.A. Placental corticotropin-releasing hormone (CRH), spontaneous preterm birth, and fetal growth restriction: A prospective investigation. Am. J. Obstet. Gynecol. 2004, 191, 1063–1069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lesage, J.; Del-Favero, F.; Leonhardt, M.; Louvart, H.; Maccari, S.; Vieau, D.; Darnaudery, M. Prenatal stress induces intrauterine growth restriction and programmes glucose intolerance and feeding behaviour disturbances in the aged rat. J. Endocrinol. 2004, 181, 291–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Mated in Summer (n = 15 Sows) | Mated in Autumn (n = 15 Sows) | SEM | p-Value |
|---|---|---|---|---|
| Parity of focal sows | 4.1 | 4.1 | 0.39 | 1.00 |
| Body weight (day 2), kg | 250 | 270 | 4.1 | 0.003 |
| Body weight (day 110), kg | 286 | 288 | 3.7 | 0.74 |
| Body weight gain, kg | 36 | 18 | 2.2 | <0.001 |
| Backfat thickness (day 2), mm | 24.1 | 27.3 | 1.23 | 0.080 |
| Backfat thickness (day 110), mm | 23.2 | 25.7 | 1.29 | 0.033 |
| Backfat thickness change, kg | −1.1 | −0.7 | 0.94 | 0.75 |
| Average daily feed intake, kg | 2.2 | 2.2 | 0.01 | 0.58 |
| Variables | Mated in Summer (n = 119 Sows) | Mated in Autumn (n = 118 Sows) | SEM | p-Value |
|---|---|---|---|---|
| Parity of sows | 3.7 | 3.8 | 0.13 | 0.83 |
| Farrowing rate | 75% | 89% | <0.001 | |
| Number of piglets born | 14.0 | 13.5 | 0.41 | 0.46 |
| Number of born-alive piglets | 12.6 | 12.5 | 0.29 | 0.73 |
| Number of stillborn piglets | 1.40 | 1.37 | 0.18 | 0.92 |
| Number of mummified piglets | 0.20 | 0.15 | 0.045 | 0.48 |
| Litter birth weight (incl. stillborn), kg | 18.5 | 19.6 | 0.040 | 0.044 |
| Litter birth weight (born-alive), kg | 17.3 | 18.4 | 0.36 | 0.024 |
| Piglet birth weight (incl. stillborn), kg | 1.37 | 1.54 | 0.026 | <0.001 |
| Piglet birth weight (born-alive), kg | 1.39 | 1.55 | 0.026 | <0.001 |
| Proportion of piglets (born-alive) ≤1.1 kg, % | 24.2 | 15.8 | 1.5 | <0.001 |
| Variables | Mated in Summer (n = 38 L) | Mated in Autumn (n = 38 L) | SEM | p-Value |
|---|---|---|---|---|
| Parity of sows | 3.7 | 3.7 | 0.24 | 1.00 |
| Number of piglets born | 15.5 | 14.7 | 0.62 | 0.37 |
| Number of born-alive piglets | 13.8 | 12.9 | 0.51 | 0.25 |
| Number of stillborn piglets | 1.4 | 1.6 | 0.36 | 0.60 |
| Number of mummified piglets | 0.3 | 0.1 | 0.10 | 0.19 |
| Placental weight, kg * | 0.26 | 0.31 | 0.012 | 0.011 |
| Piglet birth weight, kg * | 1.37 | 1.48 | 0.038 | 0.040 |
| Piglet: placental weight * | 5.72 | 5.11 | 0.230 | 0.054 |
| Variables | Progeny from Sows Mated in Summer | Progeny from Sows Mated in Autumn | SEM | p-Values | ||||
|---|---|---|---|---|---|---|---|---|
| Born Light | Born Normal | Born Light | Born Normal | Mating Season | Birth Weight | Interaction | ||
| Body weight, 9 weeks, kg | 19.6 | 20.4 | 19.6 | 20.4 | 0.27 | 0.96 | 0.005 | 0.92 |
| ADFI, kg | 1.45 | 1.50 | 1.35 | 1.36 | 0.029 | <0.001 | 0.29 | 0.67 |
| ADG, kg | 0.79 | 0.82 | 0.74 | 0.73 | 0.015 | <0.001 | 0.38 | 0.18 |
| FCR | 1.85 | 1.82 | 1.82 | 1.86 | 0.026 | 0.78 | 0.64 | 0.18 |
| Body weight, 14 weeks, kg | 47.2 | 49.2 | 45.5 | 46.2 | 0.56 | <0.001 | 0.014 | 0.25 |
| ADFI, kg | 2.83 | 2.80 | 2.40 | 2.60 | 0.045 | <0.001 | 0.070 | 0.008 |
| ADG, kg | 1.16 | 1.20 | 1.02 | 1.13 | 0.018 | <0.001 | <0.001 | 0.039 |
| FCR | 2.45 | 2.35 | 2.39 | 2.31 | 0.038 | 0.22 | 0.025 | 0.77 |
| Days to slaughter | 144.7 | 142.2 | 154.8 | 148.9 | 1.22 | <0.001 | <0.001 | 0.16 |
| Body weight, slaughter, kg | 101.0 | 101.4 | 102.1 | 102.7 | 0.69 | 0.079 | 0.47 | 0.90 |
| Variables | Progeny from Sow Mated in Summer | Progeny from Sow Mated in Autumn | SEM | p-Values | ||||
|---|---|---|---|---|---|---|---|---|
| Born Light (n = 56) | Born Normal (n = 56) | Born Light (n = 56) | Born Normal (n = 56) | Mating Season | Birth Weight | Interaction | ||
| Carcass weight, kg | 77.0 | 77.4 | 77.5 | 78.1 | 0.58 | 0.29 | 0.41 | 0.83 |
| Backfat, P2 site #, mm | 15.6 | 14.0 | 14.4 | 14.0 | 0.33 | 0.060 | 0.003 | 0.073 |
| Loin depth #, mm | 53.6 | 54.1 | 54.4 | 52.2 | 0.70 | 0.47 | 0.26 | 0.068 |
| Head weight †, kg | 6.9 | 6.9 | 7.0 | 7.0 | 0.06 | 0.27 | 0.89 | 0.41 |
| Dressing, % | 76.3 | 76.4 | 76.1 | 76.1 | 0.31 | 0.38 | 0.79 | 0.91 |
| Variables | Progeny from the Sow Mated in Summer | Progeny from the Sow Mated in Autumn | SEM | p-Values | ||||
|---|---|---|---|---|---|---|---|---|
| Born Light (n = 12) | Born Normal (n = 12) | Born Light (n = 12) | Born Normal (n = 12) | Mating Season | Birth Weight | Interaction | ||
| Age at slaughter, days | 137.5 | 136.4 | 140.2 | 139.3 | 0.44 | <0.001 | 0.027 | 0.85 |
| Live weight at slaughter, kg | 100.9 | 100.0 | 99.3 | 100.4 | 0.84 | 0.49 | 0.89 | 0.25 |
| HSCW, kg | 76.4 | 76.1 | 76.0 | 77.2 | 0.66 | 0.65 | 0.49 | 0.28 |
| Backfat thickness, mm | 14.7 | 13.4 | 13.9 | 13.8 | 0.65 | 0.69 | 0.26 | 0.36 |
| Shoulder | ||||||||
| Fat, % | 32.2 | 29.8 | 30.2 | 28.9 | 1.49 | 0.35 | 0.22 | 0.70 |
| Lean, % | 47.1 | 49.2 | 49.0 | 50.0 | 1.42 | 0.34 | 0.28 | 0.71 |
| Bone, % | 20.7 | 21.0 | 20.8 | 21.0 | 0.21 | 0.85 | 0.20 | 0.78 |
| Fat: Lean ratio | 0.70 | 0.62 | 0.63 | 0.59 | 0.049 | 0.33 | 0.23 | 0.61 |
| Loin | ||||||||
| Fat, % | 44.2 | 35.4 | 34.5 | 34.0 | 2.30 | 0.023 | 0.054 | 0.087 |
| Lean, % | 37.2 | 44.8 | 44.8 | 45.2 | 2.08 | 0.067 | 0.066 | 0.10 |
| Bone, % | 20.7 | 21.0 | 20.8 | 21.0 | 0.21 | 0.85 | 0.20 | 0.78 |
| Fat: Lean ratio | 1.34 | 0.83 | 0.80 | 0.78 | 0.128 | 0.024 | 0.040 | 0.058 |
| Belly | ||||||||
| Fat, % | 38.1 | 35.6 | 36.3 | 37.2 | 1.89 | 0.94 | 0.39 | 0.66 |
| Lean, % | 48.2 | 50.1 | 49.0 | 49.6 | 1.89 | 0.94 | 0.50 | 0.75 |
| Bone, % | 13.7 | 14.3 | 13.9 | 14.0 | 0.34 | 0.99 | 0.28 | 0.51 |
| Fat: Lean ratio | 0.81 | 0.76 | 0.77 | 0.75 | 0.069 | 0.70 | 0.59 | 0.84 |
| Leg | ||||||||
| Fat, % | 24.2 | 21.2 | 23.4 | 24.4 | 1.66 | 0.49 | 0.54 | 0.23 |
| Lean, % | 54.9 | 57.8 | 55.8 | 54.7 | 1.58 | 0.50 | 0.57 | 0.22 |
| Bone, % | 20.9 | 21.0 | 20.8 | 20.9 | 0.20 | 0.73 | 0.56 | 0.86 |
| Fat: Lean ratio | 0.46 | 0.38 | 0.43 | 0.46 | 0.047 | 0.55 | 0.64 | 0.23 |
| Half carcass | ||||||||
| Fat, % | 32.9 | 28.7 | 29.2 | 29.2 | 1.55 | 0.29 | 0.19 | 0.19 |
| Lean, % | 47.8 | 51.4 | 50.9 | 50.9 | 1.45 | 0.39 | 0.22 | 0.22 |
| Bone, % | 19.3 | 19.8 | 20.0 | 20.0 | 0.19 | 0.041 | 0.15 | 0.14 |
| Fat: Lean ratio | 0.71 | 0.57 | 0.58 | 0.59 | 0.054 | 0.29 | 0.22 | 0.19 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, F.; Ford, E.M.; Morrison, R.S.; Brewster, C.J.; Henman, D.J.; Smits, R.J.; Zhao, W.; Cottrell, J.J.; Leury, B.J.; Dunshea, F.R.; et al. The Greater Proportion of Born-Light Progeny from Sows Mated in Summer Contributes to Increased Carcass Fatness Observed in Spring. Animals 2020, 10, 2080. https://doi.org/10.3390/ani10112080
Liu F, Ford EM, Morrison RS, Brewster CJ, Henman DJ, Smits RJ, Zhao W, Cottrell JJ, Leury BJ, Dunshea FR, et al. The Greater Proportion of Born-Light Progeny from Sows Mated in Summer Contributes to Increased Carcass Fatness Observed in Spring. Animals. 2020; 10(11):2080. https://doi.org/10.3390/ani10112080
Chicago/Turabian StyleLiu, Fan, Erin M. Ford, Rebecca S. Morrison, Chris J. Brewster, David J. Henman, Robert J. Smits, Weicheng Zhao, Jeremy J. Cottrell, Brian J. Leury, Frank R. Dunshea, and et al. 2020. "The Greater Proportion of Born-Light Progeny from Sows Mated in Summer Contributes to Increased Carcass Fatness Observed in Spring" Animals 10, no. 11: 2080. https://doi.org/10.3390/ani10112080