
Immunoregulatory Effects of Porcine Plasma Protein Concentrates on Rat Intestinal Epithelial Cells and Splenocytes

 ,
,  , , and
, , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Materials
2.3. Primary Splenocytes
2.4. IEC18 Experiments
2.5. Gene Silencing
2.6. Splenocyte Experiments
2.7. Isolation of Splenic T Lymphocytes and Monocytes
2.8. Statistical Analysis
3. Results
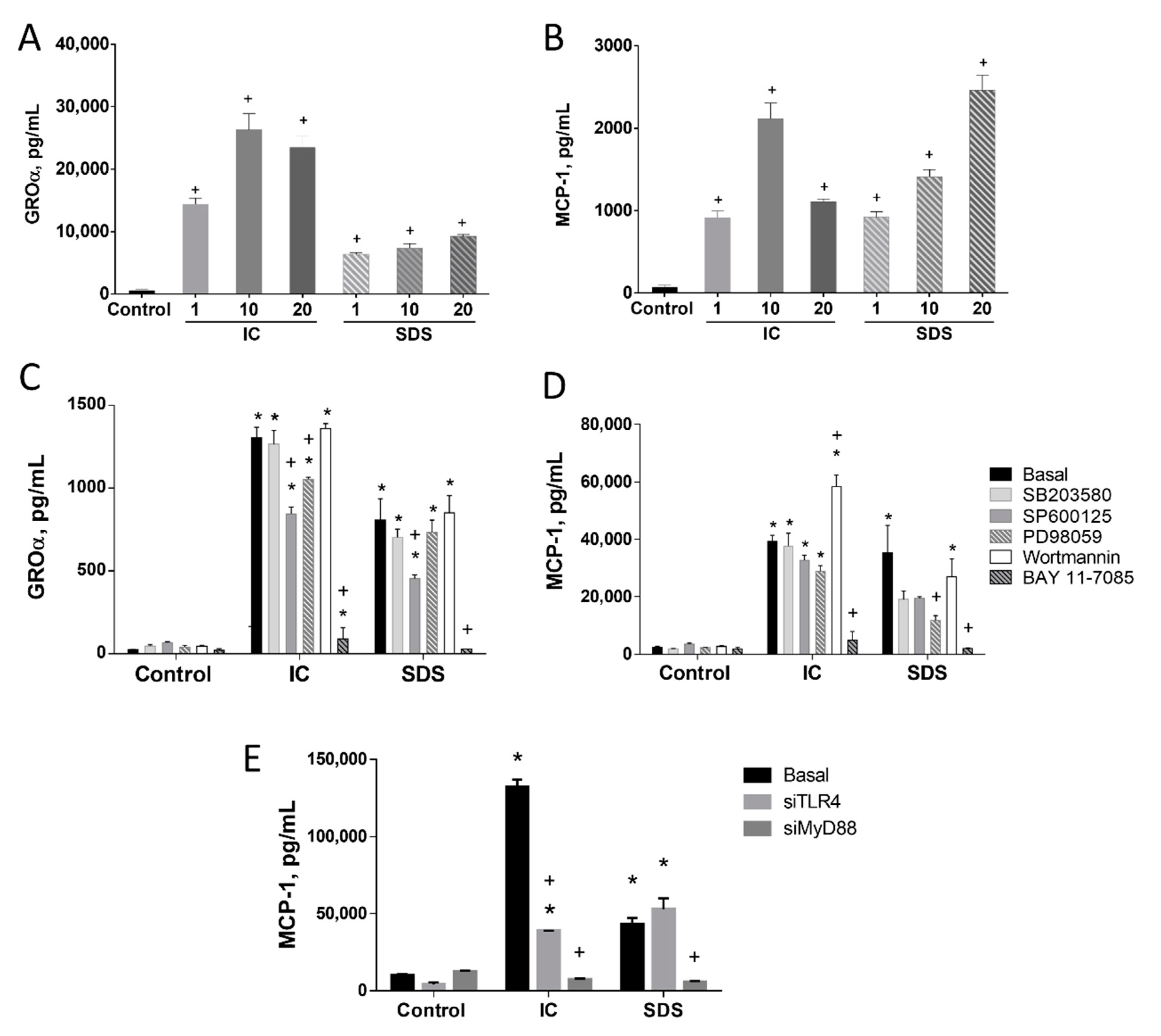
3.1. IC and SDS Induce Proinflammatory Cytokine Production in IEC18 Cells through a Pathway Involving the Activation of NF-κB
3.1.1. TLR4 Is Involved in the Induction of MCP-1 by SDS in IEC18 Cells
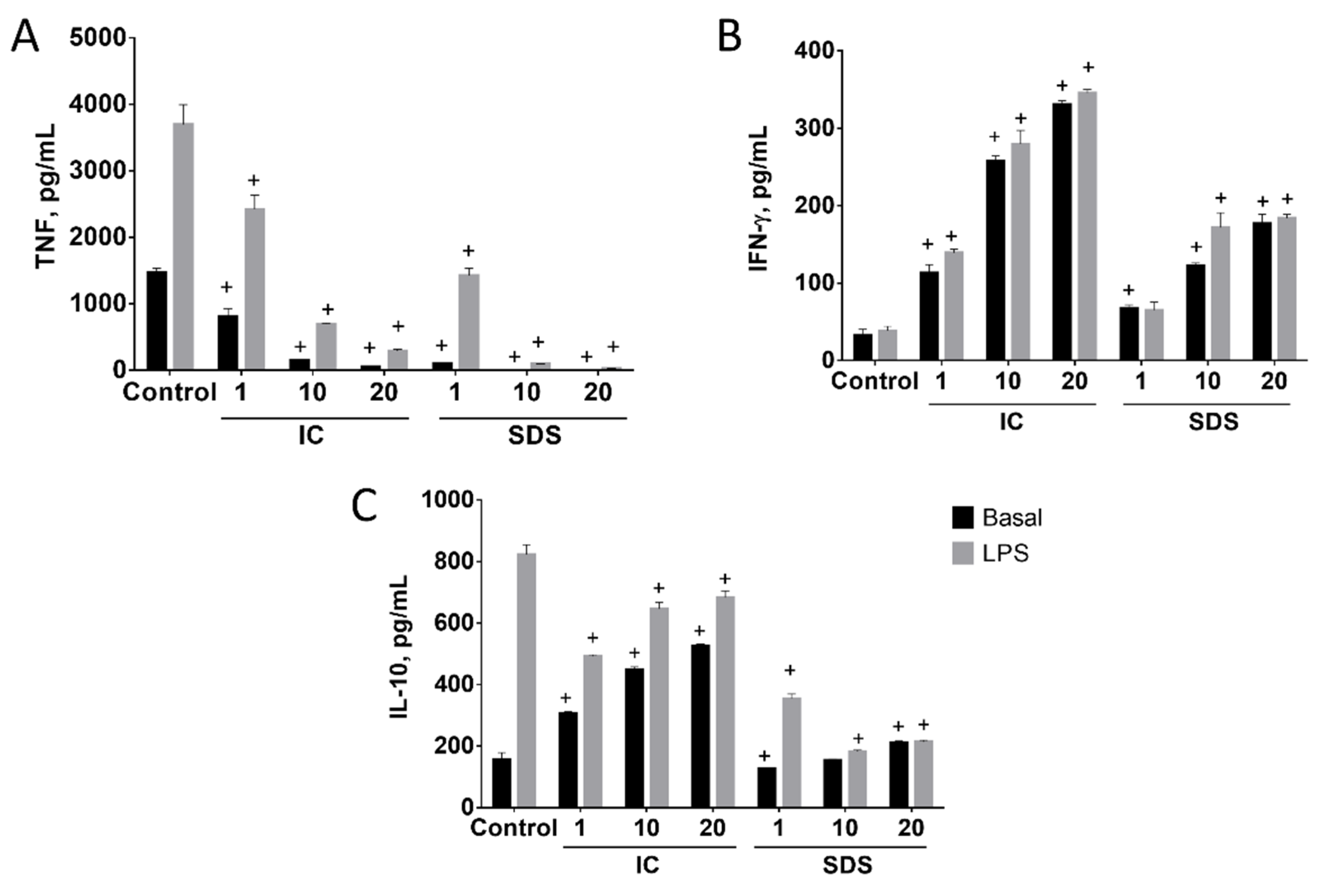
3.1.2. SDS and IC Regulate Cytokine Production in Spleen Cells
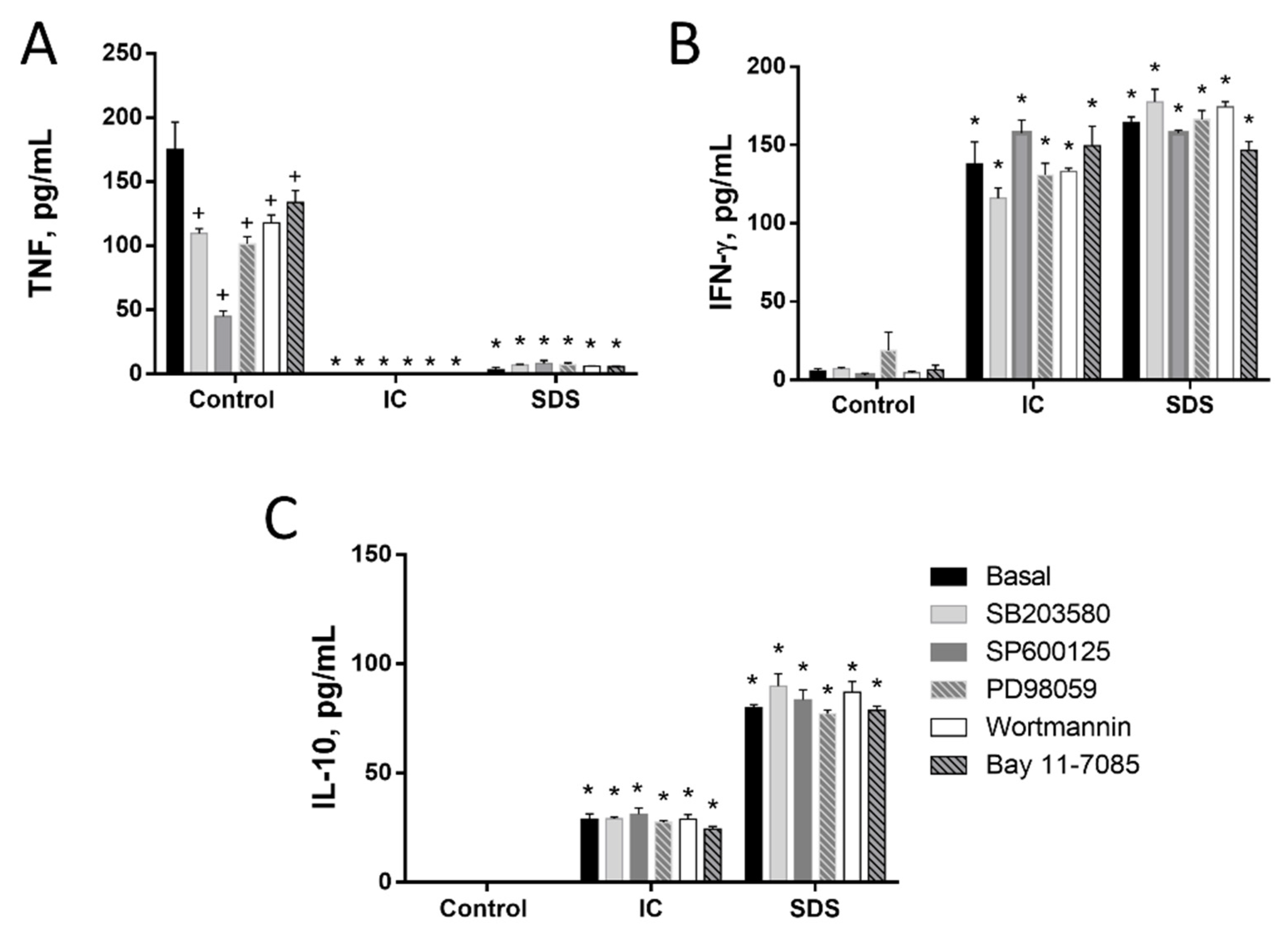
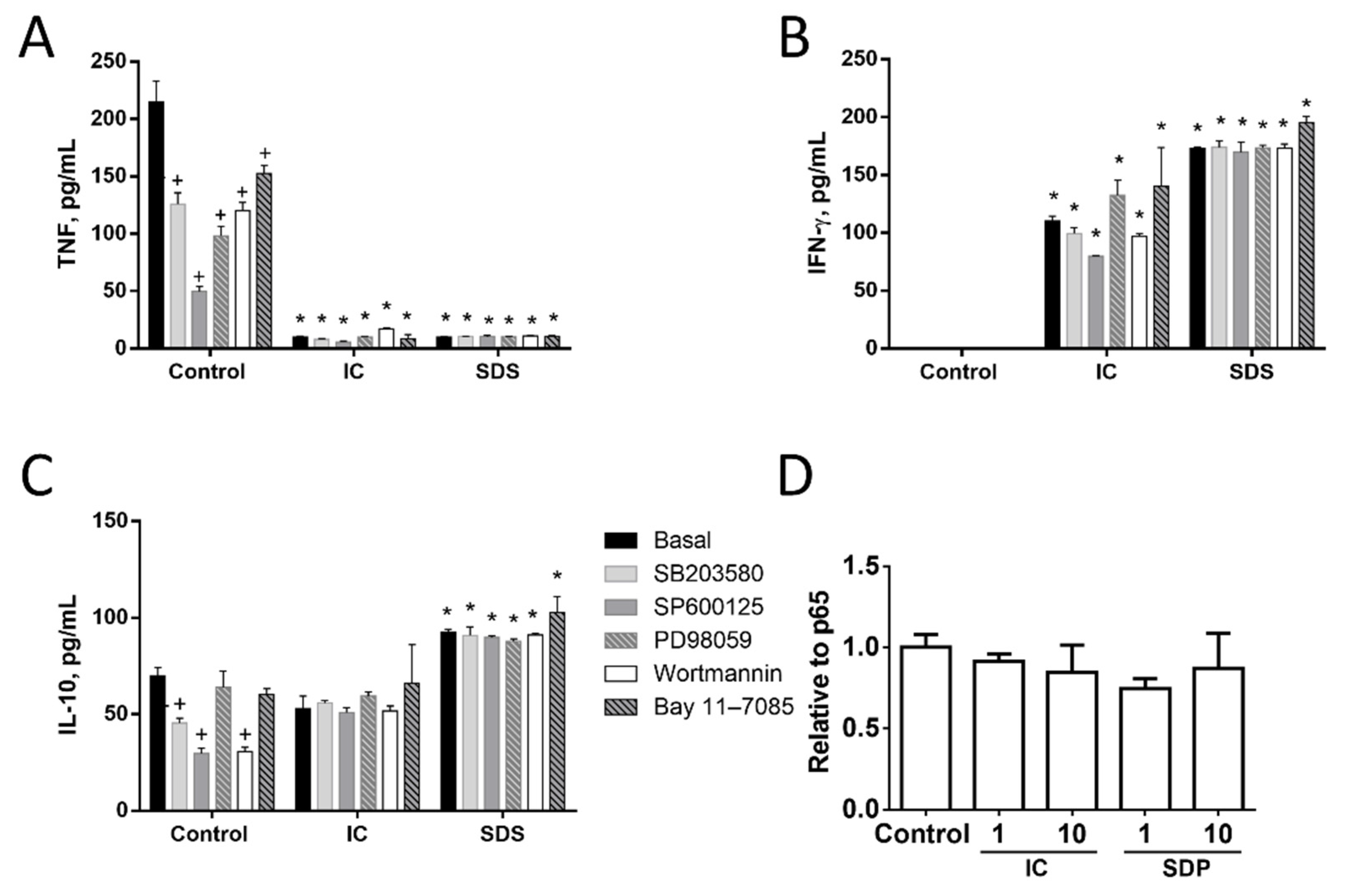
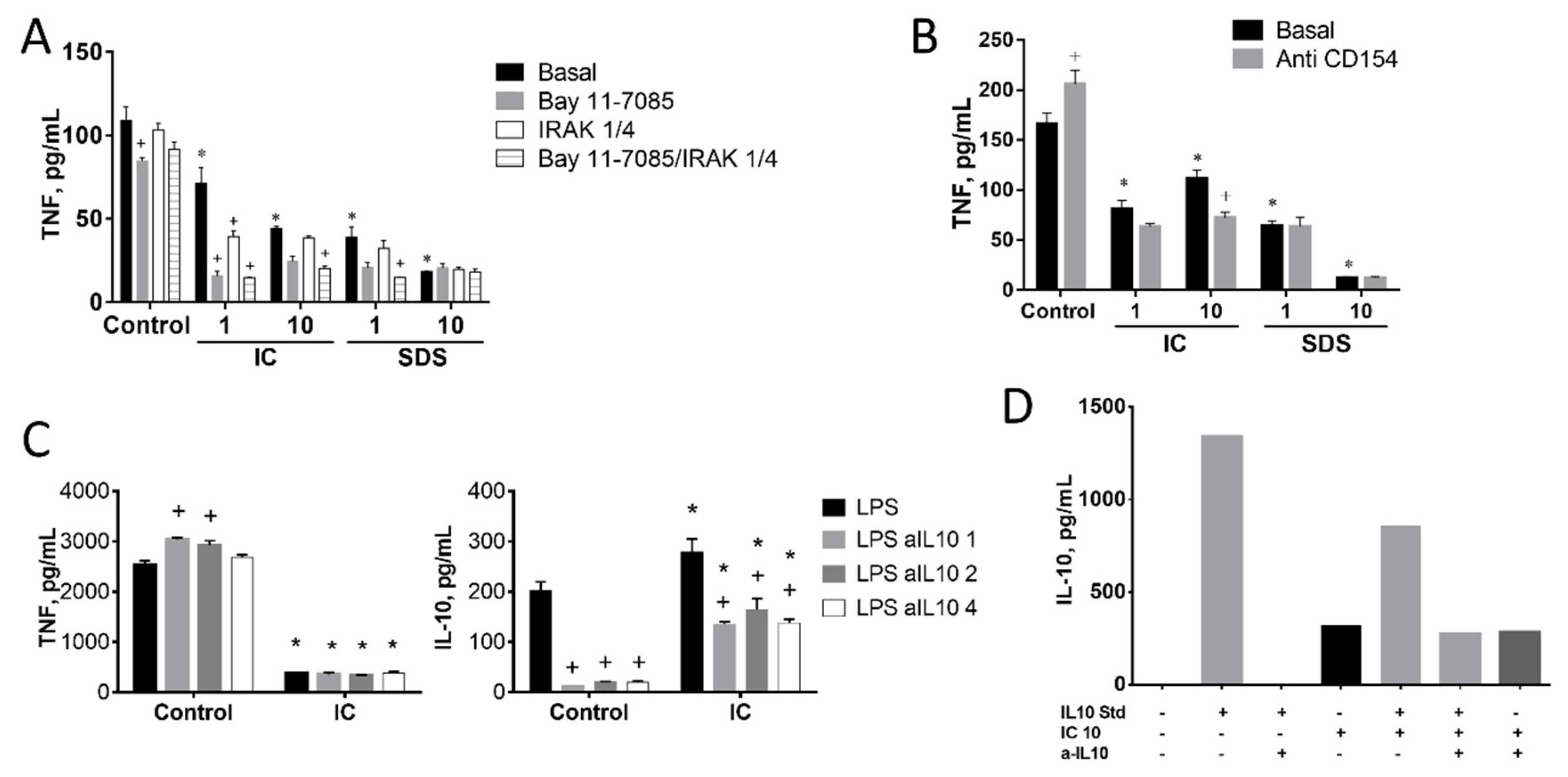
3.1.3. Signal Transduction Pathways Involving NF-κB, MAPK, AKT, or RAGE Do Not Contribute to Cytokine Regulation by IC or SPD in Spleen Cells
3.1.4. Induction of IL-10 Is Not Linked to TNF Inhibition by IC in Spleen Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Corl, B.A.; Harrell, R.J.; Moon, H.K.; Phillips, O.; Weaver, E.M.; Campbell, J.M.; Arthington, J.D.; Odle, J. Effect of animal plasma proteins on intestinal damage and recovery of neonatal pigs infected with rotavirus. J. Nutr. Biochem. 2007, 18, 778–784. [Google Scholar] [CrossRef] [PubMed]
- Perez-Bosque, A.; Miro, L.; Polo, J.; Russell, L.; Campbell, J.; Weaver, E.; Crenshaw, J.; Moreto, M. Dietary plasma protein supplements prevent the release of mucosal proinflammatory mediators in intestinal inflammation in rats. J. Nutr. 2010, 140, 25–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez-Bosque, A.; Moreto, M. A rat model of mild intestinal inflammation induced by Staphylococcus aureus enterotoxin B. Proc. Nutr. Soc. 2010, 69, 447–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maijo, M.; Miro, L.; Polo, J.; Campbell, J.; Russell, L.; Crenshaw, J.; Weaver, E.; Moreto, M.; Perez-Bosque, A. Dietary plasma proteins attenuate the innate immunity response in a mouse model of acute lung injury. Br. J. Nutr. 2012, 107, 867–875. [Google Scholar] [CrossRef] [Green Version]
- Maijo, M.; Miro, L.; Polo, J.; Campbell, J.; Russell, L.; Crenshaw, J.; Weaver, E.; Moreto, M.; Perez-Bosque, A. Dietary plasma proteins modulate the adaptive immune response in mice with acute lung inflammation. J. Nutr. 2012, 142, 264–270. [Google Scholar] [CrossRef] [PubMed]
- Moreto, M.; Perez-Bosque, A. Dietary plasma proteins, the intestinal immune system, and the barrier functions of the intestinal mucosa. J. Anim. Sci. 2009, 87, E92–E100. [Google Scholar] [CrossRef] [PubMed]
- Perez-Bosque, A.; Miro, L.; Amat, C.; Polo, J.; Moreto, M. The Anti-Inflammatory Effect of Spray-Dried Plasma Is Mediated by a Reduction in Mucosal Lymphocyte Activation and Infiltration in a Mouse Model of Intestinal Inflammation. Nutrients 2016, 8, 657. [Google Scholar] [CrossRef] [Green Version]
- Miro, L.; Garcia-Just, A.; Amat, C.; Polo, J.; Moreto, M.; Perez-Bosque, A. Dietary Animal Plasma Proteins Improve the Intestinal Immune Response in Senescent Mice. Nutrients 2017, 9, 1346. [Google Scholar] [CrossRef] [Green Version]
- Perez-Bosque, A.; Amat, C.; Polo, J.; Campbell, J.M.; Crenshaw, J.; Russell, L.; Moreto, M. Spray-dried animal plasma prevents the effects of Staphylococcus aureus enterotoxin B on intestinal barrier function in weaned rats. J. Nutr. 2006, 136, 2838–2843. [Google Scholar] [CrossRef]
- Perez-Bosque, A.; Miro, L.; Maijo, M.; Polo, J.; Campbell, J.; Russell, L.; Crenshaw, J.; Weaver, E.; Moreto, M. Dietary intervention with serum-derived bovine immunoglobulins protects barrier function in a mouse model of colitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2015, 308, G1012–G1018. [Google Scholar] [CrossRef]
- Perez-Bosque, A.; Miro, L.; Maijo, M.; Polo, J.; Campbell, J.M.; Russell, L.; Crenshaw, J.D.; Weaver, E.; Moreto, M. Oral Serum-Derived Bovine Immunoglobulin/Protein Isolate Has Immunomodulatory Effects on the Colon of Mice that Spontaneously Develop Colitis. PLoS ONE 2016, 11, e0154823. [Google Scholar] [CrossRef] [PubMed]
- Miró, L.; Amat, C.; Rosell-Cardona, C.; Campbell, J.M.; Polo, J.; Pérez-Bosque, A.; Moretó, M. Dietary Supplementation with Spray-Dried Porcine Plasma Attenuates Colon Inflammation in a Genetic Mouse Model of Inflammatory Bowel Disease. Int. J. Mol. Sci. 2020, 21, 6760. [Google Scholar] [CrossRef]
- Bateman, E.; Weaver, E.; Klein, G.; Wignall, A.; Wozniak, B.; Plews, E.; Mayo, B.; White, I.; Keefe, D. Serum-derived bovine immunoglobulin/protein isolate in the alleviation of chemotherapy-induced mucositis. Support. Care Cancer Off. J. Multinatl. Assoc. Support. Care Cancer 2016, 24, 377–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miró, L.; Amat, C.; Polo, J.; Moretó, M.; Pérez-Bosque, A. Anti-inflammatory effects of animal plasma protein supplementation in mice undergoing simultaneous gut and lung inflammation. Food Agric. Immunol. 2020, 31, 927–938. [Google Scholar] [CrossRef]
- Garcia-Just, A.; Miró, L.; Pérez-Bosque, A.; Amat, C.; Polo, J.; Pallàs, M.; Griñán-Ferré, C.; Moretó, M. Dietary Spray-Dried Porcine Plasma Prevents Cognitive Decline in Senescent Mice and Reduces Neuroinflammation and Oxidative Stress. J. Nutr. 2020, 150, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Choe, J.; Lee, J.J.; Kim, J.; Campbell, J.M.; Polo, J.; Crenshaw, J.D.; Pettigrew, J.E.; Song, M. Spray-dried plasma attenuates inflammation and lethargic behaviors of pregnant mice caused by lipopolysaccharide. PLoS ONE 2018, 13, e0203427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, T.H.; Choi, J.H.; Koh, K.C.; Jeon, W.M.; Han, K.S. Purification and Anti-pathogenic Properties of Immunoglobulin Concentrates from Porcine Blood. Korean J. Food Sci. Anim. Resour. 2017, 37, 743–751. [Google Scholar] [CrossRef] [Green Version]
- Han, K.S.; Boland, M.; Singh, H.; Moughan, P.J. The in vitro anti-pathogenic activity of immunoglobulin concentrates extracted from ovine blood. Appl. Biochem. Biotechnol. 2009, 157, 442–452. [Google Scholar] [CrossRef]
- Quigley, J.D.; Strohbehn, R.E.; Kost, C.J.; O’Brien, M.M. Formulation of colostrum supplements, colostrum replacers and acquisition of passive immunity in neonatal calves. J. Dairy Sci. 2001, 84, 2059–2065. [Google Scholar] [CrossRef]
- Good, L.; Rosario, R.; Panas, R. New therapeutic option for irritable bowel syndrome: Serum-derived bovine immunoglobulin. World J. Gastroenterol. 2015, 21, 3361–3366. [Google Scholar] [CrossRef]
- Petschow, B.W.; Burnett, B.P.; Shaw, A.L.; Weaver, E.M.; Klein, G.L. Dietary requirement for serum-derived bovine immunoglobulins in the clinical management of patients with enteropathy. Dig. Dis. Sci. 2015, 60, 13–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greenberg, P.D.; Cello, J.P. Treatment of severe diarrhea caused by Cryptosporidium parvum with oral bovine immunoglobulin concentrate in patients with AIDS. J. Acquir. Immune. Defic. Syndr. Hum. Retrovirol. 1996, 13, 348–354. [Google Scholar] [CrossRef] [PubMed]
- Tacket, C.O.; Binion, S.B.; Bostwick, E.; Losonsky, G.; Roy, M.J.; Edelman, R. Efficacy of bovine milk immunoglobulin concentrate in preventing illness after Shigella flexneri challenge. Am. J. Trop Med. Hyg. 1992, 47, 276–283. [Google Scholar] [CrossRef] [PubMed]
- Tacket, C.O.; Losonsky, G.; Link, H.; Hoang, Y.; Guesry, P.; Hilpert, H.; Levine, M.M. Protection by milk immunoglobulin concentrate against oral challenge with enterotoxigenic Escherichia coli. N. Engl. J. Med. 1988, 318, 1240–1243. [Google Scholar] [CrossRef]
- Tawfeek, H.I.; Najim, N.H.; Al-Mashikhi, S. Efficacy of an infant formula containing anti-Escherichia coli colostral antibodies from hyperimmunized cows in preventing diarrhea in infants and children: A field trial. Int. J. Infect. Dis. 2003, 7, 120–128. [Google Scholar] [CrossRef] [Green Version]
- Roos, N.; Mahé, S.; Benamouzig, R.; Sick, H.; Rautureau, J.; Tomé, D. 15N-labeled immunoglobulins from bovine colostrum are partially resistant to digestion in human intestine. J. Nutr. 1995, 125, 1238–1244. [Google Scholar] [CrossRef]
- Warny, M.; Fatimi, A.; Bostwick, E.F.; Laine, D.C.; Lebel, F.; LaMont, J.T.; Pothoulakis, C.; Kelly, C.P. Bovine immunoglobulin concentrate-clostridium difficile retains C difficile toxin neutralising activity after passage through the human stomach and small intestine. Gut 1999, 44, 212–217. [Google Scholar] [CrossRef] [Green Version]
- Kelly, C.P.; Chetham, S.; Keates, S.; Bostwick, E.F.; Roush, A.M.; Castagliuolo, I.; LaMont, J.T.; Pothoulakis, C. Survival of anti-Clostridium difficile bovine immunoglobulin concentrate in the human gastrointestinal tract. Antimicrob. Agents Chemother. 1997, 41, 236–241. [Google Scholar] [CrossRef] [Green Version]
- Henderson, A.L.; Brand, M.W.; Darling, R.J.; Maas, K.J.; Detzel, C.J.; Hostetter, J.; Wannemuehler, M.J.; Weaver, E.M. Attenuation of Colitis by Serum-Derived Bovine Immunoglobulin/Protein Isolate in a Defined Microbiota Mouse Model. Dig. Dis. Sci. 2015, 60, 3293–3303. [Google Scholar] [CrossRef] [Green Version]
- Ustunol, Z.; Wong, C. Effect of nonfat dry milk and major whey components on interleukin-6 and interleukin-8 production in human intestinal epithelial-like Caco-2 cells. J. Dairy Sci. 2010, 93, 2311–2314. [Google Scholar] [CrossRef]
- Lee, Y.Z.; Sim, J.S.; Al-Mashikhi, S.; Nakai, S. Separation of immunoglobulins from bovine blood by polyphosphate precipitation and chromatography. J. Agric. Food Chem. 1988, 36, 922–928. [Google Scholar] [CrossRef]
- Cian, R.E.; Hernandez-Chirlaque, C.; Gamez-Belmonte, R.; Drago, S.R.; Sanchez de Medina, F.; Martinez-Augustin, O. Green Alga Ulva spp. Hydrolysates and Their Peptide Fractions Regulate Cytokine Production in Splenic Macrophages and Lymphocytes Involving the TLR4-NFkappaB/MAPK Pathways. Mar. Drugs 2018, 16, 235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daddaoua, A.; Martinez-Plata, E.; Ortega-Gonzalez, M.; Ocon, B.; Aranda, C.J.; Zarzuelo, A.; Suarez, M.D.; de Medina, F.S.; Martinez-Augustin, O. The nutritional supplement Active Hexose Correlated Compound (AHCC) has direct immunomodulatory actions on intestinal epithelial cells and macrophages involving TLR/MyD88 and NF-kappaB/MAPK activation. Food Chem. 2013, 136, 1288–1295. [Google Scholar] [CrossRef]
- Foey, A.D.; Feldmann, M.; Brennan, F.M. CD40 ligation induces macrophage IL-10 and TNF-alpha production: Differential use of the PI3K and p42/44 MAPK-pathways. Cytokine 2001, 16, 131–142. [Google Scholar] [CrossRef] [PubMed]
- Cian, R.E.; Garzón, A.G.; Martínez-Augustin, O.; Botto, C.C.; Drago, S.R. Antithrombotic Activity of Brewers’ Spent Grain Peptides and their Effects on Blood Coagulation Pathways. Plant. Foods Hum. Nutr. 2018, 73, 241–246. [Google Scholar] [CrossRef]
- Reyes-López, F.E.; Ibarz, A.; Ordóñez-Grande, B.; Vallejos-Vidal, E.; Andree, K.B.; Balasch, J.C.; Fernández-Alacid, L.; Sanahuja, I.; Sánchez-Nuño, S.; Firmino, J.P.; et al. Skin Multi-Omics-Based Interactome Analysis: Integrating the Tissue and Mucus Exuded Layer for a Comprehensive Understanding of the Teleost Mucosa Functionality as Model of Study. Front. Immunol. 2020, 11, 613824. [Google Scholar] [CrossRef] [PubMed]
- Günther, C.; Buchen, B.; Neurath, M.F.; Becker, C. Regulation and pathophysiological role of epithelial turnover in the gut. Semin. Cell Dev. Biol. 2014, 35, 40–50. [Google Scholar] [CrossRef]
- Menendez, A.; Willing, B.P.; Montero, M.; Wlodarska, M.; So, C.C.; Bhinder, G.; Vallance, B.A.; Finlay, B.B. Bacterial stimulation of the TLR-MyD88 pathway modulates the homeostatic expression of ileal Paneth cell alpha-defensins. J. Innate Immun. 2013, 5, 39–49. [Google Scholar] [CrossRef]
- Omagari, D.; Takenouchi-Ohkubo, N.; Endo, S.; Ishigami, T.; Sawada, A.; Moro, I.; Asano, M.; Komiyama, K. Nuclear factor kappa B plays a pivotal role in polyinosinic-polycytidylic acid-induced expression of human beta-defensin 2 in intestinal epithelial cells. Clin. Exp. Immunol. 2011, 165, 85–93. [Google Scholar] [CrossRef]
- Gong, J.; Xu, J.; Zhu, W.; Gao, X.; Li, N.; Li, J. Epithelial-specific blockade of MyD88-dependent pathway causes spontaneous small intestinal inflammation. Clin. Immunol. 2010, 136, 245–256. [Google Scholar] [CrossRef]
- Martinez-Augustin, O.; Rivero-Gutierrez, B.; Mascaraque, C.; Sanchez de Medina, F. Food derived bioactive peptides and intestinal barrier function. Int. J. Mol. Sci. 2014, 15, 22857–22873. [Google Scholar] [CrossRef] [Green Version]
- Sanchez de Medina, F.; Romero-Calvo, I.; Mascaraque, C.; Martinez-Augustin, O. Intestinal inflammation and mucosal barrier function. Inflamm. Bowel. Dis. 2014, 20, 2394–2404. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Chirlaque, C.; Aranda, C.J.; Ocon, B.; Capitan-Canadas, F.; Ortega-Gonzalez, M.; Carrero, J.J.; Suarez, M.D.; Zarzuelo, A.; Sanchez de Medina, F.; Martinez-Augustin, O. Germ-free and Antibiotic-treated Mice are Highly Susceptible to Epithelial Injury in DSS Colitis. J. Crohn’s Colitis 2016, 10, 1324–1335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capitan-Canadas, F.; Ocon, B.; Aranda, C.J.; Anzola, A.; Suarez, M.D.; Zarzuelo, A.; de Medina, F.S.; Martinez-Augustin, O. Fructooligosaccharides exert intestinal anti-inflammatory activity in the CD4+ CD62L+ T cell transfer model of colitis in C57BL/6J mice. Eur. J. Nutr. 2016, 55, 1445–1454. [Google Scholar] [CrossRef] [PubMed]
- Ortega-Gonzalez, M.; Capitan-Canadas, F.; Requena, P.; Ocon, B.; Romero-Calvo, I.; Aranda, C.; Suarez, M.D.; Zarzuelo, A.; Sanchez de Medina, F.; Martinez-Augustin, O. Validation of bovine glycomacropeptide as an intestinal anti-inflammatory nutraceutical in the lymphocyte-transfer model of colitis. Br. J. Nutr. 2014, 111, 1202–1212. [Google Scholar] [CrossRef]
- Ortega-Gonzalez, M.; Ocon, B.; Romero-Calvo, I.; Anzola, A.; Guadix, E.; Zarzuelo, A.; Suarez, M.D.; Sanchez de Medina, F.; Martinez-Augustin, O. Nondigestible oligosaccharides exert nonprebiotic effects on intestinal epithelial cells enhancing the immune response via activation of TLR4-NFkappaB. Mol. Nutr. Food Res. 2014, 58, 384–393. [Google Scholar] [CrossRef] [PubMed]
- Mascaraque, C.; Aranda, C.; Ocon, B.; Monte, M.J.; Suarez, M.D.; Zarzuelo, A.; Marin, J.J.; Martinez-Augustin, O.; Sanchez de Medina, F. Rutin has intestinal antiinflammatory effects in the CD4+ CD62L+ T cell transfer model of colitis. Pharm. Res. 2014, 90, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Mascaraque, C.; Suarez, M.D.; Zarzuelo, A.; Sanchez de Medina, F.; Martinez-Augustin, O. Active hexose correlated compound exerts therapeutic effects in lymphocyte driven colitis. Mol. Nutr. Food Res. 2014, 58, 2379–2382. [Google Scholar] [CrossRef]
- Lopez-Posadas, R.; Requena, P.; Gonzalez, R.; Suarez, M.D.; Zarzuelo, A.; Sanchez de Medina, F.; Martinez-Augustin, O. Bovine glycomacropeptide has intestinal antiinflammatory effects in rats with dextran sulfate-induced colitis. J. Nutr. 2010, 140, 2014–2019. [Google Scholar] [CrossRef]
- Requena, P.; Daddaoua, A.; Guadix, E.; Zarzuelo, A.; Suarez, M.D.; Sanchez de Medina, F.; Martinez-Augustin, O. Bovine glycomacropeptide induces cytokine production in human monocytes through the stimulation of the MAPK and the NF-kappaB signal transduction pathways. Br. J. Pharm. 2009, 157, 1232–1240. [Google Scholar] [CrossRef] [Green Version]
- Requena, P.; Daddaoua, A.; Martínez-Plata, E.; González, M.; Zarzuelo, A.; Suárez, M.D.; Sánchez de Medina, F.; Martínez-Augustin, O. Bovine glycomacropeptide ameliorates experimental rat ileitis by mechanisms involving downregulation of interleukin 17. Br. J. Pharmacol. 2008, 154, 825–832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daddaoua, A.; Puerta, V.; Zarzuelo, A.; Suarez, M.D.; Sanchez de Medina, F.; Martinez-Augustin, O. Bovine glycomacropeptide is anti-inflammatory in rats with hapten-induced colitis. J. Nutr. 2005, 135, 1164–1170. [Google Scholar] [CrossRef] [Green Version]
- Inoue, M.; Arikawa, T.; Chen, Y.H.; Moriwaki, Y.; Price, M.; Brown, M.; Perfect, J.R.; Shinohara, M.L. T cells down-regulate macrophage TNF production by IRAK1-mediated IL-10 expression and control innate hyperinflammation. Proc. Natl. Acad. Sci. USA 2014, 111, 5295–5300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bayry, J.; Thirion, M.; Misra, N.; Thorenoor, N.; Delignat, S.; Lacroix-Desmazes, S.; Bellon, B.; Kaveri, S.; D Kazatchkine, M. Mechanisms of Action of Intravenous Immunoglobulin in Autoimmune and Inflammatory Diseases. Transfus. Clin. Biol. 2003, 24 (Suppl. S4), S217–S221. [Google Scholar]
- Requena, P.; Gonzalez, R.; Lopez-Posadas, R.; Abadia-Molina, A.; Suarez, M.D.; Zarzuelo, A.; de Medina, F.S.; Martinez-Augustin, O. The intestinal antiinflammatory agent glycomacropeptide has immunomodulatory actions on rat splenocytes. Biochem. Pharm. 2010, 79, 1797–1804. [Google Scholar] [CrossRef] [PubMed]
- Cian, R.E.; Lopez-Posadas, R.; Drago, S.R.; Medina, F.S.; Martinez-Augustin, O. Immunomodulatory Properties of the Protein Fraction from Phorphyra columbina. J. Agric. Food Chem. 2012, 60, 8146–8154. [Google Scholar] [CrossRef]
- Cian, R.E.; Lopez-Posadas, R.; Drago, S.R.; Sanchez de Medina, F.; Martinez-Augustin, O. A Porphyra columbina hydrolysate upregulates IL-10 production in rat macrophages and lymphocytes through an NF-kappaB, and p38 and JNK dependent mechanism. Food Chem. 2012, 134, 1982–1990. [Google Scholar] [CrossRef]
- Moretó, M.; Miró, L.; Amat, C.; Polo, J.; Manichanh, C.; Pérez-Bosque, A. Dietary supplementation with spray-dried porcine plasma has prebiotic effects on gut microbiota in mice. Sci. Rep. 2020, 10, 2926. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hernández-Chirlaque, C.; Aranda, C.J.; Ocón, B.; Polo, J.; Martínez-Augustin, O.; Sánchez de Medina, F. Immunoregulatory Effects of Porcine Plasma Protein Concentrates on Rat Intestinal Epithelial Cells and Splenocytes. Animals 2021, 11, 807. https://doi.org/10.3390/ani11030807
Hernández-Chirlaque C, Aranda CJ, Ocón B, Polo J, Martínez-Augustin O, Sánchez de Medina F. Immunoregulatory Effects of Porcine Plasma Protein Concentrates on Rat Intestinal Epithelial Cells and Splenocytes. Animals. 2021; 11(3):807. https://doi.org/10.3390/ani11030807
Chicago/Turabian StyleHernández-Chirlaque, Cristina, Carlos J. Aranda, Borja Ocón, Javier Polo, Olga Martínez-Augustin, and Fermín Sánchez de Medina. 2021. "Immunoregulatory Effects of Porcine Plasma Protein Concentrates on Rat Intestinal Epithelial Cells and Splenocytes" Animals 11, no. 3: 807. https://doi.org/10.3390/ani11030807