
Development of Colorimetric Reverse Transcription Loop-Mediated Isothermal Amplification Assay for Detecting Feline Coronavirus

 , and
, and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell and Virus Propagation
2.2. FCoV RNA Extraction
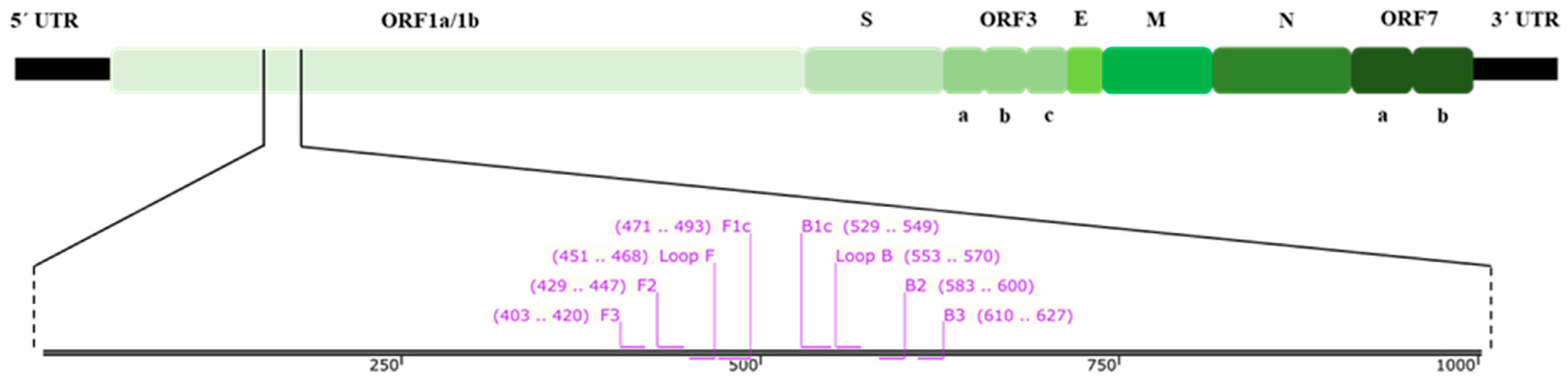
2.3. RT-LAMP Primer Design
2.4. RT-LAMP Reaction and Optimization
2.5. RT-LAMP Product Analysis
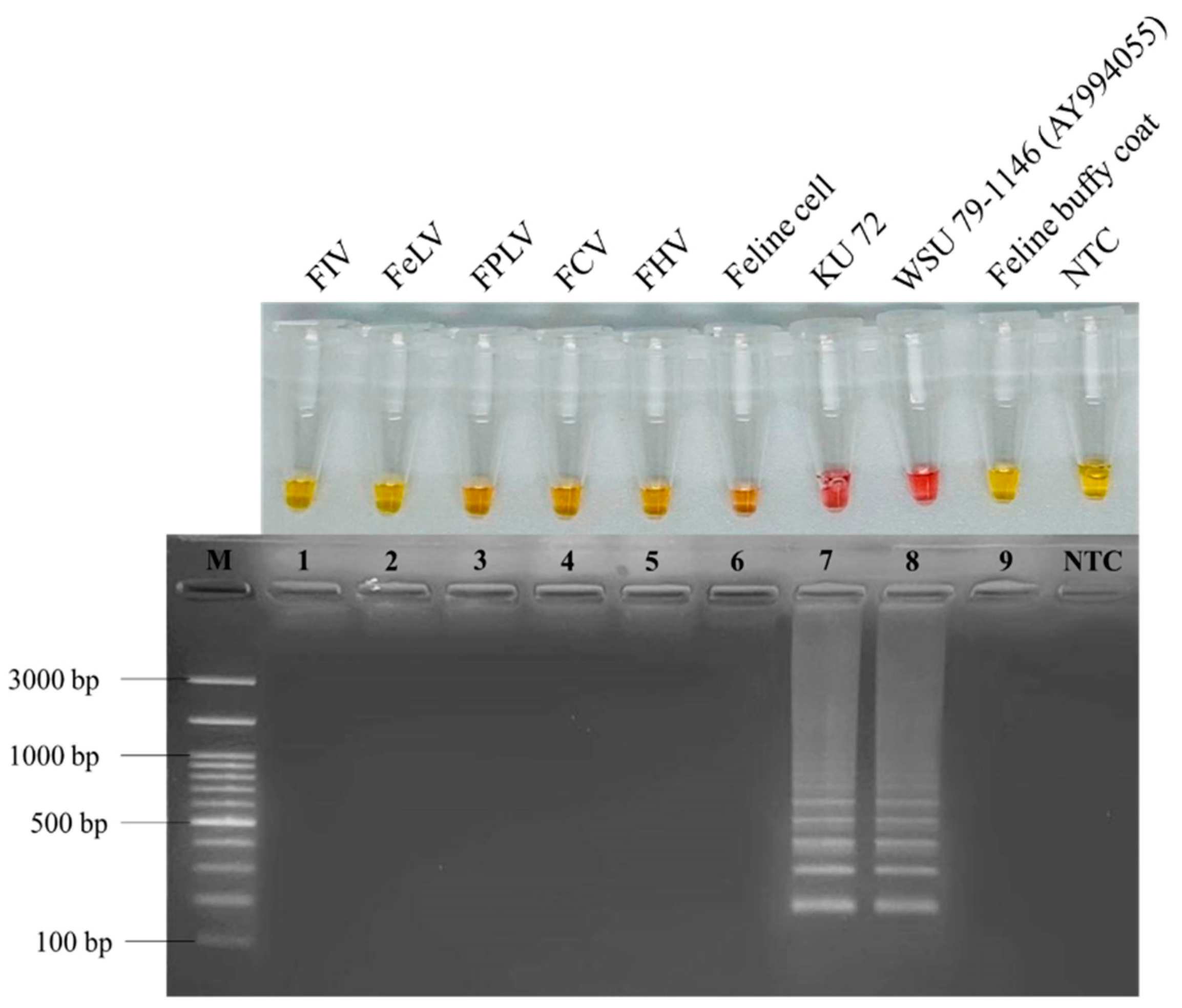
2.6. RT-LAMP Specificity Analysis
2.7. Reverse Transcription PCR (RT-PCR)
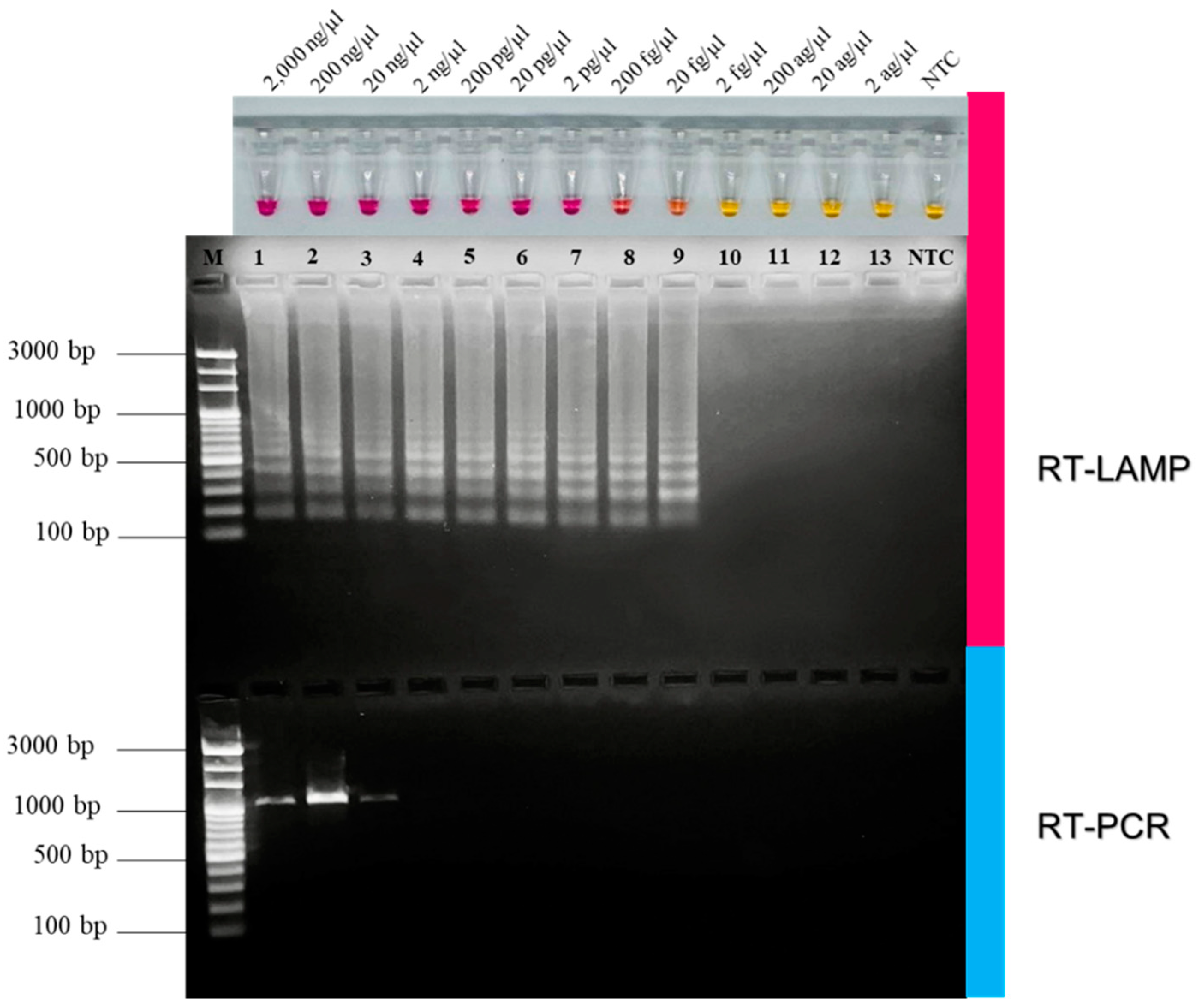
2.8. Comparison of RT-LAMP and RT-PCR on Sensitivity for FCoV Detection
2.9. Clinical Samples Test Using RT-LAMP Assay
2.10. Statistical Analysis
3. Results
3.1. Optimization of RT-LAMP Reaction
3.2. Specificity of Developed RT-LAMP for Detection of FCoV
3.3. Comparative Sensitivity of RT-LAMP and RT-PCR for FCoV Detection
3.4. Evaluation of RT-LAMP with Clinical Samples
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhou, Z.; Qiu, Y.; Ge, X. The Taxonomy, Host Range and Pathogenicity of Coronaviruses and Other Viruses in the Nidovirales Order. Anim. Dis. 2021, 1, 5. [Google Scholar] [CrossRef] [PubMed]
- Santana-Clavijo, N.F.; Reyes Romero, D.P.; Arango Fajardo, D.F.; Velandia Muñoz, A.; Taniwaki, S.A.; de Souza Silva, S.O.; Brandão, P.E. Molecular Diversity of Alphacoronavirus 1 in Dogs and Cats in Colombia. Heliyon 2020, 6, e04381. [Google Scholar] [CrossRef] [PubMed]
- Mavrodiev, E.V.; Tursky, M.L.; Mavrodiev, N.E.; Ebach, M.C.; Williams, D.M. On Classification and Taxonomy of Coronaviruses (Riboviria, Nidovirales, Coronaviridae) with Special Focus on Severe Acute Respiratory Syndrome-Related Coronavirus 2 (SARS-Cov-2) Short Title: On the Classification of Coronaviruses. bioRxiv 2020. [Google Scholar] [CrossRef]
- Kipar, A.; Meli, M.L. Feline Infectious Peritonitis: Still an Enigma? Vet. Pathol. 2014, 51, 505–526. [Google Scholar] [CrossRef] [PubMed]
- González, J.M.; Gomez-Puertas, P.; Cavanagh, D.; Gorbalenya, A.E.; Enjuanes, L. A Comparative Sequence Analysis to Revise the Current Taxonomy of the Family Coronaviridae. Arch. Virol. 2003, 148, 2207–2235. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Chen, C.; Liu, X.; Yang, K.; Xu, X.; Yang, H. Crystal Structure of Feline Infectious Peritonitis Virus Main Protease in Complex with Synergetic Dual Inhibitors. J. Virol. 2016, 90, 1910–1917. [Google Scholar] [CrossRef]
- Lewis, C.S.; Porter, E.; Matthews, D.; Kipar, A.; Tasker, S.; Helps, C.R.; Siddell, S.G. Genotyping Coronaviruses Associated with Feline Infectious Peritonitis. J. Gen. Virol. 2015, 96, 1358–1368. [Google Scholar] [CrossRef] [PubMed]
- Günther, S.; Felten, S.; Wess, G.; Hartmann, K.; Weber, K. Detection of Feline Coronavirus in Effusions of Cats with and without Feline Infectious Peritonitis Using Loop-Mediated Isothermal Amplification. J. Virol. Methods 2018, 256, 32–36. [Google Scholar] [CrossRef]
- Pedersen, N.C.; Black, J.W.; Boyle, J.F.; Evermann, J.F.; McKeirnan, A.J.; Ott, R.L. Pathogenic Differences Between Various Feline Coronavirus Isolates. In Molecular Biology and Pathogenesis of Coronaviruses. Advances in Experimental Medicine and Biology; Springer: Boston, MA, USA, 1984; Volume 173, pp. 365–380. [Google Scholar]
- Barker, E.N.; Stranieri, A.; Helps, C.R.; Porter, E.L.; Davidson, A.D.; Day, M.J.; Knowles, T.; Kipar, A.; Tasker, S. Limitations of Using Feline Coronavirus Spike Protein Gene Mutations to Diagnose Feline Infectious Peritonitis. Vet. Res. 2017, 48, 60. [Google Scholar] [CrossRef]
- Emmler, L.; Felten, S.; Matiasek, K.; Balzer, H.J.; Pantchev, N.; Leutenegger, C.; Hartmann, K. Feline Coronavirus with and without Spike Gene Mutations Detected by Real-Time RT-PCRs in Cats with Feline Infectious Peritonitis. J. Feline Med. Surg. 2020, 22, 791–799. [Google Scholar] [CrossRef]
- Felten, S.; Klein-Richers, U.; Hofmann-Lehmann, R.; Bergmann, M.; Unterer, S.; Leutenegger, C.M.; Hartmann, K. Correlation of Feline Coronavirus Shedding in Feces with Coronavirus Antibody Titer. Pathogens 2020, 9, 598. [Google Scholar] [CrossRef] [PubMed]
- Le Poder, S.; Pham-Hung d’Alexandry d’Orangiani, A.L.; Duarte, L.; Fournier, A.; Horhogea, C.; Pinhas, C.; Vabret, A.; Eloit, M. Infection of Cats with Atypical Feline Coronaviruses Harbouring a Truncated Form of the Canine Type I Non-Structural ORF3 Gene. Infect. Genet. Evol. 2013, 20, 488–494. [Google Scholar] [CrossRef] [PubMed]
- Le Poder, S. Feline and Canine Coronaviruses: Common Genetic and Pathobiological Features. Adv. Virol. 2011, 2011, 609465. [Google Scholar] [CrossRef]
- Delaplace, M.; Huet, H.; Gambino, A.; le Poder, S. Feline Coronavirus Antivirals: A Review. Pathogens 2021, 10, 1150. [Google Scholar] [CrossRef]
- Pedersen, N.C.; Liu, H.; Dodd, K.A.; Pesavento, P.A. Significance of Coronavirus Mutants in Feces and Diseased Tissues of Cats Suffering from Feline Infectious Peritonitis. Viruses 2009, 1, 166–184. [Google Scholar] [CrossRef] [PubMed]
- Ehmann, R.; Kristen-Burmann, C.; Bank-Wolf, B.; König, M.; Herden, C.; Hain, T.; Thiel, H.J.; Ziebuhr, J.; Tekes, G. Reverse Genetics for Type I Feline Coronavirus Field Isolate to Study the Molecular Pathogenesis of Feline Infectious Peritonitis. mBio 2018, 9, e01422-18. [Google Scholar] [CrossRef] [PubMed]
- Tuanthap, S.; Chiteafea, N.; Rattanasrisomporn, J.; Choowongkomon, K. Comparative Sequence Analysis of the Accessory and Nucleocapsid Genes of Feline Coronavirus Strains Isolated from Cats Diagnosed with Effusive Feline Infectious Peritonitis. Arch. Virol. 2021, 166, 2779–2787. [Google Scholar] [CrossRef] [PubMed]
- Herrewegh, A.A.P.M.; de Groot, R.J.; Cepica, A.; Egberink, H.F.; Horzinek, M.C.; Rottier, P.J.M. Detection of Feline Coronavirus RNA in Feces, Tissues, and Body Fluids of Naturally Infected Cats by Reverse Transcriptase PCR. J. Clin. Microbiol. 1995, 33, 684–689. [Google Scholar] [CrossRef]
- Rottier, P.J.M.; Nakamura, K.; Schellen, P.; Volders, H.; Haijema, B.J. Acquisition of Macrophage Tropism during the Pathogenesis of Feline Infectious Peritonitis Is Determined by Mutations in the Feline Coronavirus Spike Protein. J. Virol. 2005, 79, 14122–14130. [Google Scholar] [CrossRef]
- Vennema, H.; Poland, A.; Foley, J.; Pedersen, N.C. Feline Infectious Peritonitis Viruses Arise by Mutation from Endemic Feline Enteric Coronaviruses. Virology 1998, 243, 150–157. [Google Scholar] [CrossRef]
- Felten, S.; Hartmann, K. Diagnosis of Feline Infectious Peritonitis: A Review of the Current Literature. Viruses 2019, 11, 1068. [Google Scholar] [CrossRef] [PubMed]
- Moyadee, W.; Jaroensong, T.; Roytrakul, S.; Boonkaewwan, C.; Rattanasrisomporn, J. Characteristic Clinical Signs and Blood Parameters in Cats with Feline Infectious Peritonitis. Agric. Nat. Resour. 2019, 53, 433–438. [Google Scholar]
- Gut, M.; Leutenegger, C.M.; Huder, J.B.; Pedersen, N.C.; Lutz, H. One-Tube Fluorogenic Reverse Transcription-Polymerase Chain Reaction for the Quantitation of Feline Coronaviruses. J. Virol. Methods 1999, 77, 37–46. [Google Scholar] [CrossRef]
- Techangamsuwan, S.; Radtanakatikanon, A.; Purnaveja, S. Molecular Detection and Genotype Differentiation of Feline Coronavirus Isolates from Clinical Specimens in Thailand. Thai J. Vet. Med. 2012, 42, 413–422. [Google Scholar]
- Hornyák, Á.; Bálint, Á.; Farsang, A.; Balka, G.; Hakhverdyan, M.; Rasmussen, T.B.; Blomberg, J.; Belák, S. Detection of Subgenomic MRNA of Feline Coronavirus by Real-Time Polymerase Chain Reaction Based on Primer-Probe Energy Transfer (P-Sg-QPCR). J. Virol. Methods 2012, 181, 155–163. [Google Scholar] [CrossRef]
- Dye, C.; Helps, C.R.; Siddell, S.G. Evaluation of Real-Time RT-PCR for the Quantification of FCoV Shedding in the Faeces of Domestic Cats. J. Feline Med. Surg. 2008, 10, 167–174. [Google Scholar] [CrossRef]
- Doenges, S.J.; Weber, K.; Dorsch, R.; Fux, R.; Hartmann, K. Comparison of Real-Time Reverse Transcriptase Polymerase Chain Reaction of Peripheral Blood Mononuclear Cells, Serum and Cell-Free Body Cavity Effusion for the Diagnosis of Feline Infectious Peritonitis. J. Feline Med. Surg. 2017, 19, 344–350. [Google Scholar] [CrossRef]
- Fish, E.J.; Diniz, P.P.V.P.; Juan, Y.C.; Bossong, F.; Collisson, E.W.; Drechsler, Y.; Kaltenboeck, B. Cross-Sectional Quantitative RT-PCR Study of Feline Coronavirus Viremia and Replication in Peripheral Blood of Healthy Shelter Cats in Southern California. J. Feline Med. Surg. 2018, 20, 295–301. [Google Scholar] [CrossRef]
- Longstaff, L.; Porter, E.; Crossley, V.J.; Hayhow, S.E.; Helps, C.R.; Tasker, S. Feline Coronavirus Quantitative Reverse Transcriptase Polymerase Chain Reaction on Effusion Samples in Cats with and without Feline Infectious Peritonitis. J. Feline Med. Surg. 2017, 19, 240–245. [Google Scholar] [CrossRef]
- Manasateinkij, W.; Nilkumhang, P.; Jaroensong, T.; Noosud, J.; Lekcharoensuk, C.; Lekcharoensuk, P. Occurence of Feline Coronavirus and Feline Infectious Peritonitis Virus in Thailand. Agric. Nat. Resour. 2009, 43, 720–726. [Google Scholar]
- Gaffney, P.M.; Kennedy, M.; Terio, K.; Gardner, I.; Lothamer, C.; Coleman, K.; Munson, L. Detection of Feline Coronavirus in Cheetah (Acinonyx jubatus) Feces by Reverse Transcription-Nested Polymerase Chain Reaction in Cheetahs with Variable Frequency of Viral Shedding. J. Zoo Wildl. Med. 2012, 43, 776–786. [Google Scholar] [CrossRef] [PubMed]
- Felten, S.; Weider, K.; Doenges, S.; Gruendl, S.; Matiasek, K.; Hermanns, W.; Mueller, E.; Matiasek, L.; Fischer, A.; Weber, K.; et al. Detection of Feline Coronavirus Spike Gene Mutations as a Tool to Diagnose Feline Infectious Peritonitis. J. Feline Med. Surg. 2017, 19, 321–335. [Google Scholar] [CrossRef] [PubMed]
- Techangamsuwan, S.; Radtanakatikanon, A.; Thanawongnuwech, R. Development and Application of Reverse Transcription Loop-Mediated Isothermal Amplification (RT-LAMP) for Feline Coronavirus Detection. Thai J. Vet. Med. 2013, 43, 229–233. [Google Scholar]
- Stranieri, A.; Lauzi, S.; Giordano, A.; Paltrinieri, S. Reverse Transcriptase Loop-Mediated Isothermal Amplification for the Detection of Feline Coronavirus. J. Virol. Methods 2017, 243, 105–108. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, N.C. An Update on Feline Infectious Peritonitis: Diagnostics and Therapeutics. Vet. J. 2014, 201, 133–141. [Google Scholar] [CrossRef]
- Reed, L.J.; Muench, H. A Simple Method of Estimating Fifty per Cent Endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Stephensen, C.B.; Casebolt, D.B.; Gangopadhyay, N.N. Phylogenetic Analysis of a Highly Conserved Region of the Polymerase Gene from 11 Coronaviruses and Development of a Consensus Polymerase Chain Reaction Assay. Virus Res. 1999, 60, 181–189. [Google Scholar] [CrossRef]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.; Madden, T.L. Primer-BLAST: A Tool to Design Target-Specific Primers for Polymerase Chain Reaction. BMC Bioinform. 2012, 13, 134. [Google Scholar] [CrossRef]
- Wang, Y.; Yuan, X.; Li, Y.; Yu, K.; Yang, J.; Xu, H.; Zhang, Y.; Yu, K.; Liao, M.; Qin, Z. Rapid Detection of Newly Isolated Tembusu-Related Flavivirus by Reverse-Transcription Loop-Mediated Isothermal Amplification Assay. Virol. J. 2011, 8, 553. [Google Scholar] [CrossRef]
- Song, C.; Wan, H.; Yu, S.; Han, X.; Qiu, X.; Hu, Q.; Tan, L.; Ding, C. Rapid Detection of Duck Hepatitis Virus Type-1 by Reverse Transcription Loop-Mediated Isothermal Amplification. J. Virol. Methods 2012, 182, 76–81. [Google Scholar] [CrossRef]
- Whelan, J.A.; Russell, N.B.; Whelan, M.A. A Method for the Absolute Quantification of cDNA Using Real-Time PCR. J. Immunol. Methods 2003, 278, 261–269. [Google Scholar] [CrossRef]
- Inaba, M.; Higashimoto, Y.; Toyama, Y.; Horiguchi, T.; Hibino, M.; Iwata, M.; Imaizumi, K.; Doi, Y. Diagnostic Accuracy of LAMP versus PCR over the Course of SARS-CoV-2 Infection. Int. J. Infect. Dis. 2021, 107, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Landis, J.R.; Koch, G.G. The Measurement of Observer Agreement for Categorical Data. Biometrics 1977, 33, 159. [Google Scholar] [CrossRef]
- McHugh, M.L. Interrater Reliability: The Kappa Statistic. Biochem. Med. 2012, 22, 276–282. [Google Scholar] [CrossRef]
- Pedersen, N.C. A Review of Feline Infectious Peritonitis Virus Infection: 1963-2008. J. Feline Med. Surg. 2009, 11, 225–258. [Google Scholar] [CrossRef]
- Chang, H.W.; Egberink, H.F.; Halpin, R.; Spiro, D.J.; Rottie, P.J.M. Spike Protein Fusion Peptide and Feline Coronavirus Virulence. Emerg. Infect. Dis. 2012, 18, 1089–1095. [Google Scholar] [CrossRef] [PubMed]
- Licitra, B.N.; Millet, J.K.; Regan, A.D.; Hamilton, B.S.; Rinaldi, V.D.; Duhamel, G.E.; Whittaker, G.R. Mutation in Spike Protein Cleavage Site and Pathogenesis of Feline Coronavirus. Emerg. Infect. Dis. 2013, 19, 1066–1073. [Google Scholar] [CrossRef]
- Bank-Wolf, B.R.; Stallkamp, I.; Wiese, S.; Moritz, A.; Tekes, G.; Thiel, H.J. Mutations of 3c and Spike Protein Genes Correlate with the Occurrence of Feline Infectious Peritonitis. Vet. Microbiol. 2014, 173, 177–188. [Google Scholar] [CrossRef]
- Notomi, T.; Okayama, H.; Masubuchi, H.; Yonekawa, T.; Watanabe, K.; Amino, N.; Hase, T. Loop-Mediated Isothermal Amplification of DNA. Nucleic Acids Res. 2000, 28, e63. [Google Scholar] [CrossRef]
- Parida, M.; Sannarangaiah, S.; Dash, P.K.; Rao, P.V.L.; Morita, K. Loop Mediated Isothermal Amplification (LAMP): A New Generation of Innovative Gene Amplification Technique; Perspectives in Clinical Diagnosis of Infectious Diseases. Rev. Med. Virol. 2008, 18, 407–421. [Google Scholar] [CrossRef]
- Cardoso, T.C.; Ferrari, H.F.; Bregano, L.C.; Silva-Frade, C.; Rosa, A.C.G.; Andrade, A.L. Visual Detection of Turkey Coronavirus RNA in Tissues and Feces by Reverse-Transcription Loop-Mediated Isothermal Amplification (RT-LAMP) with Hydroxynaphthol Blue Dye. Mol. Cell. Probes 2010, 24, 415–417. [Google Scholar] [CrossRef] [PubMed]
- Hanaki, K.I.; Ike, F.; Hatakeyama, R.; Hirano, N. Reverse Transcription-Loop-Mediated Isothermal Amplification for the Detection of Rodent Coronaviruses. J. Virol. Methods 2013, 187, 222–227. [Google Scholar] [CrossRef]
- Nemoto, M.; Morita, Y.; Niwa, H.; Bannai, H.; Tsujimura, K.; Yamanaka, T.; Kondo, T. Rapid Detection of Equine Coronavirus by Reverse Transcription Loop-Mediated Isothermal Amplification. J. Virol. Methods 2015, 215, 13–16. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Hu, X.; Wang, X.; Yang, J.; Zhang, L.; Deng, Q.; Zhang, X.; Wang, Z.; Hou, T.; Li, S. A Novel One-Pot Rapid Diagnostic Technology for COVID-19. Anal. Chim. Acta 2021, 1154, 338310. [Google Scholar] [CrossRef] [PubMed]
- Pyrc, K.; Milewska, A.; Potempa, J. Development of Loop-Mediated Isothermal Amplification Assay for Detection of Human Coronavirus-NL63. J. Virol. Methods 2011, 175, 133–136. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Wang, L.; An, X.; Tong, Y. All-in-One in Situ Colorimetric RT-LAMP Assay for Point-of-Care Testing of SARS-CoV-2. Analyst 2021, 146, 6026–6034. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, W.; Matsumura, Y.; Thongchankaew-Seo, U.; Yamazaki, Y.; Nagao, M. Development of a Point-of-Care Test to Detect SARS-CoV-2 from Saliva Which Combines a Simple RNA Extraction Method with Colorimetric Reverse Transcription Loop-Mediated Isothermal Amplification Detection. J. Clin. Virol. 2021, 136, 104760. [Google Scholar] [CrossRef]
- Sila, T.; Sunghan, J.; Laochareonsuk, W.; Surasombatpattana, S.; Kongkamol, C.; Ingviya, T.; Siripaitoon, P.; Kositpantawong, N.; Kanchanasuwan, S.; Hortiwakul, T.; et al. Suspected Cat-to-Human Transmission of SARS-CoV-2, Thailand, July–September 2021. Emerg. Infect. Dis. 2022, 28, 1485. [Google Scholar] [CrossRef]
- Nagamine, K.; Watanabe, K.; Ohtsuka, K.; Hase, T.; Notomi, T. Loop-Mediated Isothermal Amplification Reaction Using a Nondenatured Template. Clin. Chem. 2001, 47, 1742–1743. [Google Scholar] [CrossRef]
- Quyen, T.L.; Ngo, T.A.; Bang, D.D.; Madsen, M.; Wolff, A. Classification of Multiple DNA Dyes Based on Inhibition Effects on Real-Time Loop-Mediated Isothermal Amplification (LAMP): Prospect for Point of Care Setting. Front. Microbiol. 2019, 10, 2234. [Google Scholar] [CrossRef]
- Mori, Y.; Nagamine, K.; Tomita, N.; Notomi, T. Detection of Loop-Mediated Isothermal Amplification Reaction by Turbidity Derived from Magnesium Pyrophosphate Formation. Biochem. Biophys. Res. Commun. 2001, 289, 150–154. [Google Scholar] [CrossRef]
- Tomita, N.; Mori, Y.; Kanda, H.; Notomi, T. Loop-Mediated Isothermal Amplification (LAMP) of Gene Sequences and Simple Visual Detection of Products. Nat. Protoc. 2008, 3, 877–882. [Google Scholar] [CrossRef] [PubMed]
- Saechue, B.; Kamiyama, N.; Wang, Y.; Fukuda, C.; Watanabe, K.; Soga, Y.; Goto, M.; Dewayani, A.; Ariki, S.; Hirose, H.; et al. Development of a Portable Reverse Transcription Loop-Mediated Isothermal Amplification System to Detect the E1 Region of Chikungunya Virus in a Cost-Effective Manner. Genes Cells 2020, 25, 615–625. [Google Scholar] [CrossRef] [PubMed]
- Jaroenram, W.; Cecere, P.; Pompa, P.P. Xylenol Orange-Based Loop-Mediated DNA Isothermal Amplification for Sensitive Naked-Eye Detection of Escherichia coli. J. Microbiol. Methods 2019, 156, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Tanner, N.A.; Zhang, Y.; Evans, T.C. Visual Detection of Isothermal Nucleic Acid Amplification Using pH-Sensitive Dyes. Biotechniques 2015, 58, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Frisch, L.M.; Niessen, L. Development and Optimization of a Group-Specific Loop-Mediated Isothermal Amplification (LAMP) Assay for the Detection of Patulin-Producing Penicillium Species. Int. J. Food Microbiol. 2019, 298, 20–30. [Google Scholar] [CrossRef]
- Zhang, F.; Wu, J.; Wang, R.; Wang, L.; Ying, Y. Portable pH-Inspired Electrochemical Detection of DNA Amplification. Chem. Commun. 2014, 50, 8416–8419. [Google Scholar] [CrossRef]
- Jothikumar, P.; Narayanan, J.; Hill, V.R. Visual Endpoint Detection of Escherichia coli O157:H7 Using Isothermal Genome Exponential Amplification Reaction (GEAR) Assay and Malachite Green. J. Microbiol. Methods 2014, 98, 122–127. [Google Scholar] [CrossRef]
- Wang, D.; Yu, J.; Wang, Y.; Zhang, M.; Li, P.; Liu, M.; Liu, Y. Development of a Real-Time Loop-Mediated Isothermal Amplification (LAMP) Assay and Visual LAMP Assay for Detection of African Swine Fever Virus (ASFV). J. Virol. Methods 2020, 276, 113775. [Google Scholar] [CrossRef]
- Frisch, L.M.; Mann, M.A.; Marek, D.N.; Niessen, L. Development and Optimization of a Loop-Mediated Isothermal Amplification (LAMP) Assay for the Species-Specific Detection of Penicillium expansum. Food Microbiol. 2021, 95, 103681. [Google Scholar] [CrossRef]
- Wang, Y.; Dai, J.; Liu, Y.; Yang, J.; Hou, Q.; Ou, Y.; Ding, Y.; Ma, B.; Chen, H.; Li, M.M.; et al. Development of a Potential Penside Colorimetric LAMP Assay Using Neutral Red for Detection of African Swine Fever Virus. Front. Microbiol. 2021, 12, 516. [Google Scholar] [CrossRef] [PubMed]
- Shokrollahi, A.; Firoozbakht, F. Determination of the Acidity Constants of Neutral Red and Bromocresol Green by Solution Scanometric Method and Comparison with Spectrophotometric Results. Beni-Suef Univ. J. Basic Appl. Sci. 2016, 5, 13–20. [Google Scholar] [CrossRef]
- Fuertes-Perez, S.; Hilgarth, M.; Vogel, R.F. Development of a Rapid Detection Method for Photobacterium spp. Using Loop-Mediated Isothermal Amplification (LAMP). Int. J. Food Microbiol. 2020, 334, 108805. [Google Scholar] [CrossRef] [PubMed]
- Penning, L.C.; Vrieling, H.E.; Brinkhof, B.; Riemers, F.M.; Rothuizen, J.; Rutteman, G.R.; Hazewinkel, H.A.W. A Validation of 10 Feline Reference Genes for Gene Expression Measurements in Snap-Frozen Tissues. Vet. Immunol. Immunopathol. 2007, 120, 212–222. [Google Scholar] [CrossRef]
- Ashwell, M.; Freire, M.; O’Nan, A.T.; Benito, J.; Hash, J.; McCulloch, R.S.; Lascelles, B.D.X. Characterization of Gene Expression in Naturally Occurring Feline Degenerative Joint Disease-Associated Pain. Vet. J. 2019, 243, 42–47. [Google Scholar] [CrossRef]
- Seetaha, S.; Khamplong, P.; Wanaragthai, P.; Aiebchun, T.; Ratanabunyong, S.; Krobthong, S.; Yingchutrakul, Y.; Rattanasrisomporn, J.; Choowongkomon, K. KERRA, Mixed Medicinal Plant Extracts, Inhibits SARS-CoV-2 Targets Enzymes and Feline Coronavirus. COVID 2022, 2, 621–632. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequence (5′ → 3′) | Ref. |
|---|---|---|
| F3 | GTACTRCTWCCGTAACGG | This work |
| B3 | CGGGCRAHTTTAAGGTCA | |
| FIP (F1c-F2) | GACGTAGTGATCCTTACGATYRC-CTATGTYTTCGTGCCTGA | |
| BIP (B1c-B2) | ACCTGTYCTCMTTACCGAACC-AGAACGCCATTGCAGTTR | |
| Loop F | TCAACTAGGTCACGACGG | |
| Loop B | CGTCATGTTGCARGGYTT | |
| F2 | GCTATGTYTTCGTGCCTGA | |
| B2 | AGAACGCCATTGCAGTTR | |
| F9N | CGTCAACTGGGGAGATGAAC | [18] |
| R9N | CATCTCAACCTGTGTGTCATC | |
| F7 | TAAAATGGCCKTGGTATGTGT | |
| R7Sc | ACTTCTCATRAACGGTGC |
| Detection Method | Detection Limit | |
|---|---|---|
| g/µL | Copy Molecules/µL | |
| NR-based RT-LAMP | 20 fg/µL | 1.5 × 105 |
| Conventional RT-PCR | 20 ng/µL | 3.5 × 1010 |
| RT-LAMP | Conventional RT-PCR | Sensitivity (%) (95% CI) | Specificity (%) (95% CI) | ĸ Value | ||
|---|---|---|---|---|---|---|
| Positive | Negative | Total | ||||
| Positive | 60 | 1 | 61 | 94 (61.1–100.0) | 94.12 (55.8–100.0) | 0.79 |
| Negative | 4 | 16 | 20 | |||
| Total | 64 | 17 | 81 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rapichai, W.; Saejung, W.; Khumtong, K.; Boonkaewwan, C.; Tuanthap, S.; Lieberzeit, P.A.; Choowongkomon, K.; Rattanasrisomporn, J. Development of Colorimetric Reverse Transcription Loop-Mediated Isothermal Amplification Assay for Detecting Feline Coronavirus. Animals 2022, 12, 2075. https://doi.org/10.3390/ani12162075
Rapichai W, Saejung W, Khumtong K, Boonkaewwan C, Tuanthap S, Lieberzeit PA, Choowongkomon K, Rattanasrisomporn J. Development of Colorimetric Reverse Transcription Loop-Mediated Isothermal Amplification Assay for Detecting Feline Coronavirus. Animals. 2022; 12(16):2075. https://doi.org/10.3390/ani12162075
Chicago/Turabian StyleRapichai, Witsanu, Wichayet Saejung, Kotchaporn Khumtong, Chaiwat Boonkaewwan, Supansa Tuanthap, Peter A. Lieberzeit, Kiattawee Choowongkomon, and Jatuporn Rattanasrisomporn. 2022. "Development of Colorimetric Reverse Transcription Loop-Mediated Isothermal Amplification Assay for Detecting Feline Coronavirus" Animals 12, no. 16: 2075. https://doi.org/10.3390/ani12162075